")
Back to Journals » International Journal of General Medicine » Volume 15
HSPB11 is a Prognostic Biomarker Associated with Immune Infiltrates in Hepatocellular Carcinoma
Received 25 February 2022
Accepted for publication 5 April 2022
Published 13 April 2022 Volume 2022:15 Pages 4017—4027
DOI https://doi.org/10.2147/IJGM.S363679
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Hui Liu,1,* Mei Yang,1,* Zhiwei Dong2
1Department of Gastroenterology, Second Hospital of Dalian Medical University, Dalian, Liaoning, People’s Republic of China; 2Department of General Surgery, Air Force Medical Center, PLA, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiwei Dong, Department of General Surgery, Air Force Medical Center, PLA, Beijing, People’s Republic of China, Tel +8617611408626, Fax +86 411-84671291-3106, Email [email protected]
Purpose: Heat shock proteins (HSPs) play important roles in oncogenesis and malignant progression. HSPB11 is highly expressed in many malignant tumors, but research on its role in hepatocellular carcinoma (HCC) is insufficient.
Patients and Methods: A comprehensive analysis of HSPB11 in HCC was performed based on data of patients with HCC and those from online public databases.
Results: HSPB11 was overexpressed in HCC, with a high discrimination ability between tumor and normal tissues (area under the curve =0.923). HSPB11 overexpression correlated with advanced tumor stage, poorer tumor differentiation, and worse prognosis and was an independent risk factor for HCC prognosis. The nomogram and calibration models composed of HSPB11, T stage, and M stage had good abilities to predict the 1-, 3-, and 5-year survival rates of patients. HSPB11 was determined to be involved in multiple oncogenic processes, including cell cycle checkpoints, the G2M checkpoint, E2F targets, Rho GTPases, and KRAS signaling. HSPB11 expression was related to immune cell infiltration, especially that of Th2 cells and dendritic cells.
Conclusion: HSPB11 is involved in oncogenesis and immune regulation in HCC and is a potential prognostic biomarker and therapeutic target.
Keywords: heat shock protein B11, biomarker, immune cell infiltration, Th2 cell, dendritic cell
Introduction
Hepatocellular carcinoma (HCC) is characterized by high morbidity and mortality, making it the sixth most common cancer and one of the most common causes of cancer-related death.1,2 Patients with early-stage HCC can be effectively treated with surgical resection, radiofrequency ablation, or liver transplantation, but most patients are not diagnosed until advanced disease stages, resulting in a poor prognosis.3,4 Systemic drug therapies, such as sorafenib, lenvatinib, regorafenib, and cabozantinib, are recommended for advanced-stage HCC.5–8 However, the mortality of patients with advanced HCC remains very high.9 Therefore, researchers have been exploring valuable prognostic biomarkers and more effective treatment methods.10
Heat shock proteins (HSPs) are a family of highly conserved proteins that are expressed at low basal levels under normal physiological conditions, but their expression is upregulated upon exposure to heat stress and tumorigenic environments.11–14 Several members of the HSP family, such as HSP27, HSP70, and HSP90, play important roles in the oncogenesis, therapeutic resistance, invasion, and metastasis of HCC.15–20 Various inhibitors of HSPs have been proposed as possible treatment strategies for HCC, which could be used alone or in combination with other treatment approaches.21 HSPB11 has been investigated in many animal models, such as fish and birds.22,23 Its expression is closely related to several diseases, such as tumors and degenerative lesions of the nervous system.15,24–29 For example, HSPB11 is overexpressed in high-grade gliomas, and its expression level correlates with the degree of malignancy and prognosis.27,29,30 In esophageal cancer, the expression level of HSPB11 is closely related to the survival and prognosis of patients. Combined with other biomarkers, it can be used to predict the response to neoadjuvant radiochemotherapy in patients with esophageal cancer.28,31 Norouzinia et al found that the expression of HSPB11 in gastric cancer tissues of patients infected with Helicobacter pylori is lower than that in tissues of individuals not infected with H. pylori, suggesting that it plays a role in the mechanism of H. pylori infection.32 Yang et al reported that HSPB11 is highly expressed in HCC, but its roles in oncogenesis and immune regulation are still not clear.15 Thus, we comprehensively analyzed the role of HSPB11 in HCC, including its diagnostic and prognostic values, biological functions, and association with the immune microenvironment.
Materials and Methods
Comparison of HSPB11 Expression Using Data from Online Public Databases
The expression level of HSPB11 was investigated in 17 common tumors using data from The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx) databases. Additionally, 225 liver tumor tissues and 220 non-tumor tissues from GSE14520, as well as 81 tumor tissues and 10 non-tumor tissues from GSE62232 were retrieved to evaluate the expression level of HSPB11. The receiver operating characteristic (ROC) curve was created to explore the diagnostic value of HSPB11 in HCC.
Validation of HSPB11 Expression in Clinical Samples
Thirty tumor tissues and their adjacent normal tissues were collected from HCC patients in the Second Hospital of Dalian Medical University. Quantitative reverse-transcription polymerase chain reaction (qRT-PCR) was performed using the ThermoScript RT-PCR system (Invitrogen, Carlsbad, CA, USA) and StepOnePlus apparatus (Applied Biosystems, Foster City, CA, USA). The amplification primers used were as follows: HSPB11 forward, TGATGGCTCCGCTACTTACTT and reverse, GCAGAAACGCTATGCACAGAT; glyceraldehyde-3-phosphate dehydrogenase (GADPH, the internal reference gene) forward, CAGCCTCAAGATCATCAGCAAT and reverse, ATGAGTCCTTCCACGATACCAAA. Three technical replicates were performed for each sample. The Ethics Committee of Second Hospital of Dalian Medical University approved this research (no. 0202159). Written informed consent was obtained from all patients before specimen collection.
Association Between HSPB11 and Cancer Stage and Prognosis
Correlation analyses were performed using the R ggplot2 package based on data derived from TCGA to investigate the association between HSPB11 expression and tumor size (T stage), pathological stage, tumor differentiation (histological grade), and adjacent hepatic tissue inflammation. Kaplan–Meier plots and univariate and multivariate Cox proportional hazards models were created using the survival package to evaluate the effect of HSPB11 on the prognosis of HCC and to identify risk factors for HCC based on data available in TCGA database. Another online database (Kaplan–Meier Plotter, https://kmplot.com/analysis/) was also used to evaluate the effect of HSPB11 on the prognosis of patients with HCC.
Establishment of Prognostic Models for HCC
The time-dependent survival ROC curve was created using the R timeROC package to evaluate the predictive ability of HSPB11 for the prognosis of patients with HCC. By integrating risk factors identified in the aforementioned Cox regression model, the nomogram and calibration models were built using the R rms and survival packages to predict the survival probabilities.
Enrichment of HSPB11-Associated Genes in HCC
Patients with HCC from TCGA were divided into HSPB11-high and HSPB11-low groups, and the differentially expressed genes and their corresponding logFC values were screened using R with the DESeq2 package. The gene set enrichment analysis (GSEA) was performed using the clusterProfiler package based on MSigDB Collections (https://www.gsea-msigdb.org/). A false discovery rate <0.25 and adjusted P-value <0.05 were used as criteria for statistical significance. h.all.v7.2.symbols.gmt Hallmarks and c2.cp.v7.2.symbols.gmt Curated were used as the reference gene sets.
Correlation Analyses of HSPB11 and Immune Cell Infiltration
The ssGSEA analysis was performed using the R GSVA package to demonstrate the association between HSPB11 and the level of infiltration of 24 types of immune cells. Spearman correlation and Wilcoxon rank-sum tests were applied to investigate the relationship between HSPB11 expression and the immune cell infiltration level.
Statistical Analyses
R (v.3.6.3) and RStudio software were used for the statistical analyses, and P < 0.05 was set as the criterion for statistical significance.
Results
Expression Level of HSPB11 in Pan-Cancers and HCC
HSPB11 was more highly expressed in 13 types of tumors (breast infiltrating carcinoma, bladder urothelial carcinoma, cholangiocarcinoma, colon adenocarcinoma, esophageal carcinoma, pleomorphic glioma, renal clear cell carcinoma, renal papillary cell carcinoma, liver hepatocellular carcinoma, lung adenocarcinoma, lung squamous cell carcinoma, prostate cancer, and gastric cancer) than in their adjacent normal tissues (Figure 1A). The hyperexpression of HSPB11 was observed in HCC tissues in both unmatched (Figure 1B) and paired (Figure 1C) comparative studies based on TCGA database. Furthermore, in the ROC curve, the area under the curve (AUC) was 0.923, indicating a high ability of HSPB11 to differentiate tumor and normal tissues (Figure 1D). The comparative analysis of public databases of GSE14520 (Figure 1E) and GSE62232 (Figure 1F), accompanied by the qRT-PCR analysis of clinical samples from our hospital (Figure 1G), all confirmed higher HSPB11 mRNA expression in HCC than in the adjacent normal liver tissues.
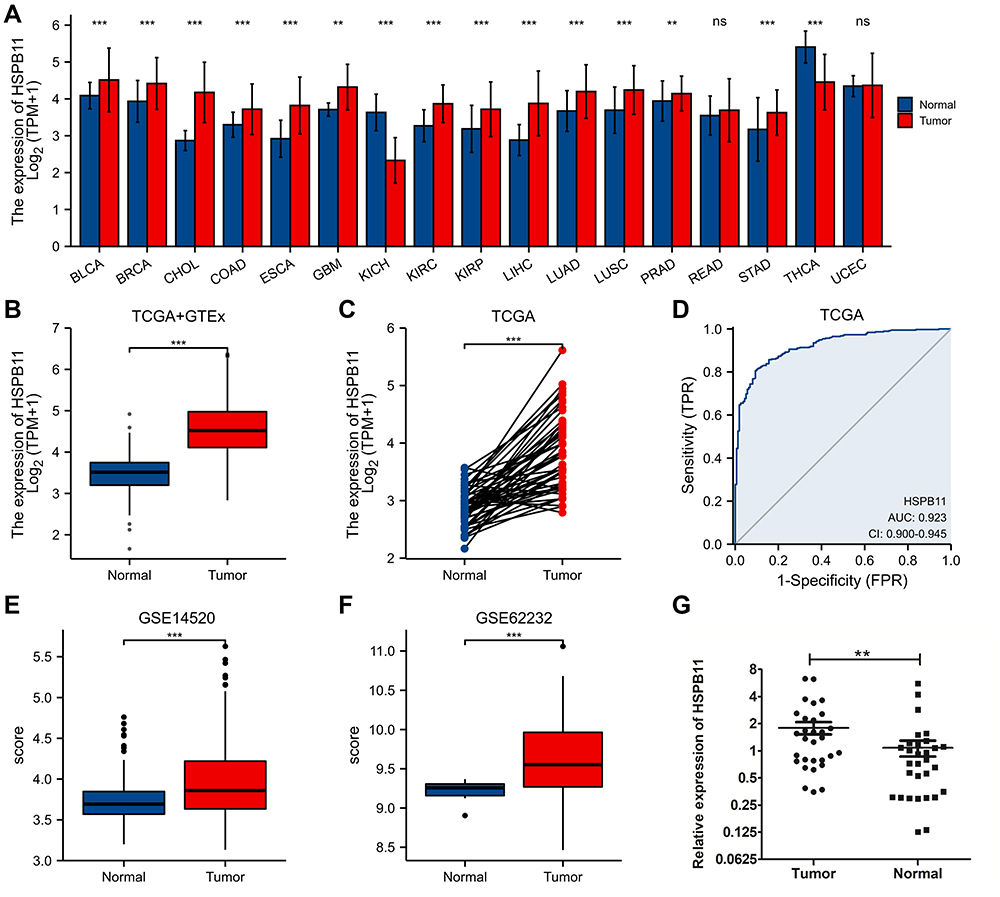 |
Figure 1 Expression level of HSPB11 in pan-cancers and hepatocellular carcinoma (HCC). (A) HSPB11 expression in pan-cancers compared with that in normal tissues in The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx) databases. (B–E) Expression levels of HSPB11 in HCC in several online public databases. (F) Receiver operating characteristic curve to test the value of HSPB11 to identify HCC tissues. (G) The quantitative polymerase chain reaction of clinical HCC samples confirmed that the mRNA level of HSPB11 in tumor tissues was higher than that in the adjacent normal liver tissue. **p < 0.01, ***p < 0.001, ns, no significance. |
HSPB11 Correlates with HCC Stage and Prognosis
The clinical data used to identify the relationship between HSPB11 and the cancer stage and prognosis of HCC were retrieved from TCGA database, and the baseline characteristics of the 374 HCC patients are presented in Table S1. The expression level of HSPB11 was positively correlated with tumor size, pathological stage, and histological grade, indicating that patients with advanced stage and poorer tumor differentiation tend to express higher level of HSPB11. (Figure 2A–C). Higher HSPB11 expression was also observed in HCC patients with adjacent hepatic tissue inflammation (Figure 2D).
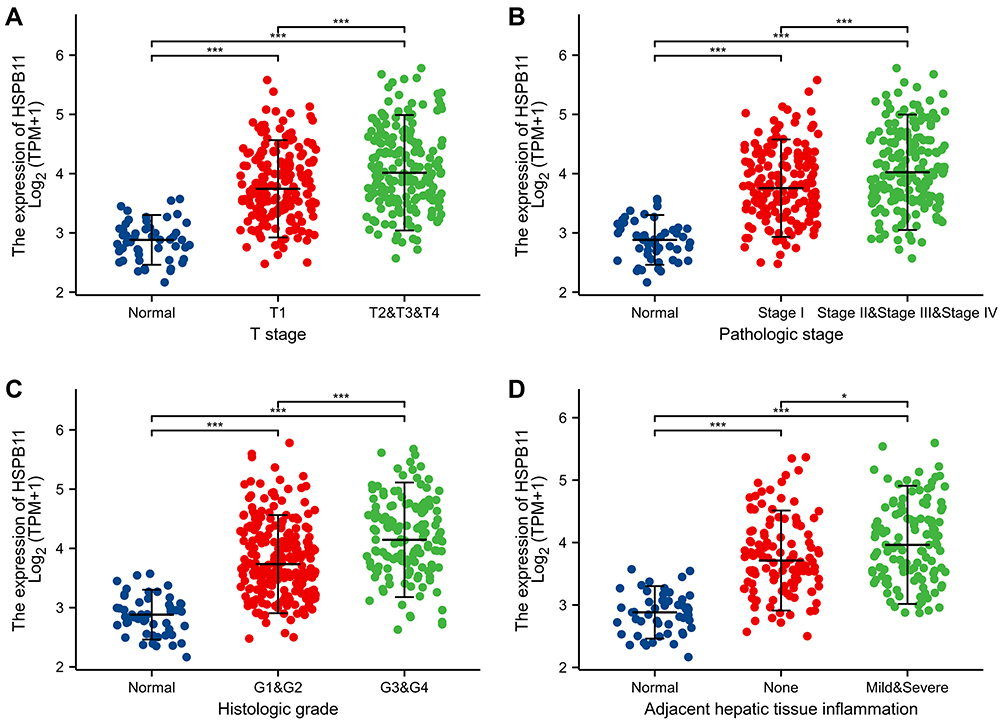 |
Figure 2 Relationship between the expression level of HSPB11 and stage of hepatocellular carcinoma. The expression level of HSPB11 was highly correlated with (A) T stage, (B) pathological stage, (C) histological grade, and (D) adjacent hepatic tissue inflammation. *p < 0.05, ***p < 0.001. |
The association between the HSPB11 level and the prognosis of HCC was evaluated. According to the Kaplan–Meier survival curves based on TCGA, HCC patients with higher HSPB11 expression showed a lower overall survival (OS) (hazard ratio (HR) = 2.09, 95% confidence interval (CI) = 1.45–2.99, p < 0.001), poorer disease-specific survival (DSS; HR = 2.19, CI = 1.38–3.48, p = 0.001), and a worse progression-free interval (PFI; HR = 1.70, CI = 1.25–2.32, p = 0.001) (Figure 3A–C). An analysis of the Kaplan–Meier Plotter database confirmed the poorer prognosis of HCC patients with high HSPB11 expression (Figure 3D–F). The multivariate Cox models showed that tumor size, distant metastasis, and the expression level of HSPB11 were all independent risk factors for the prognosis of HCC (Table 1).
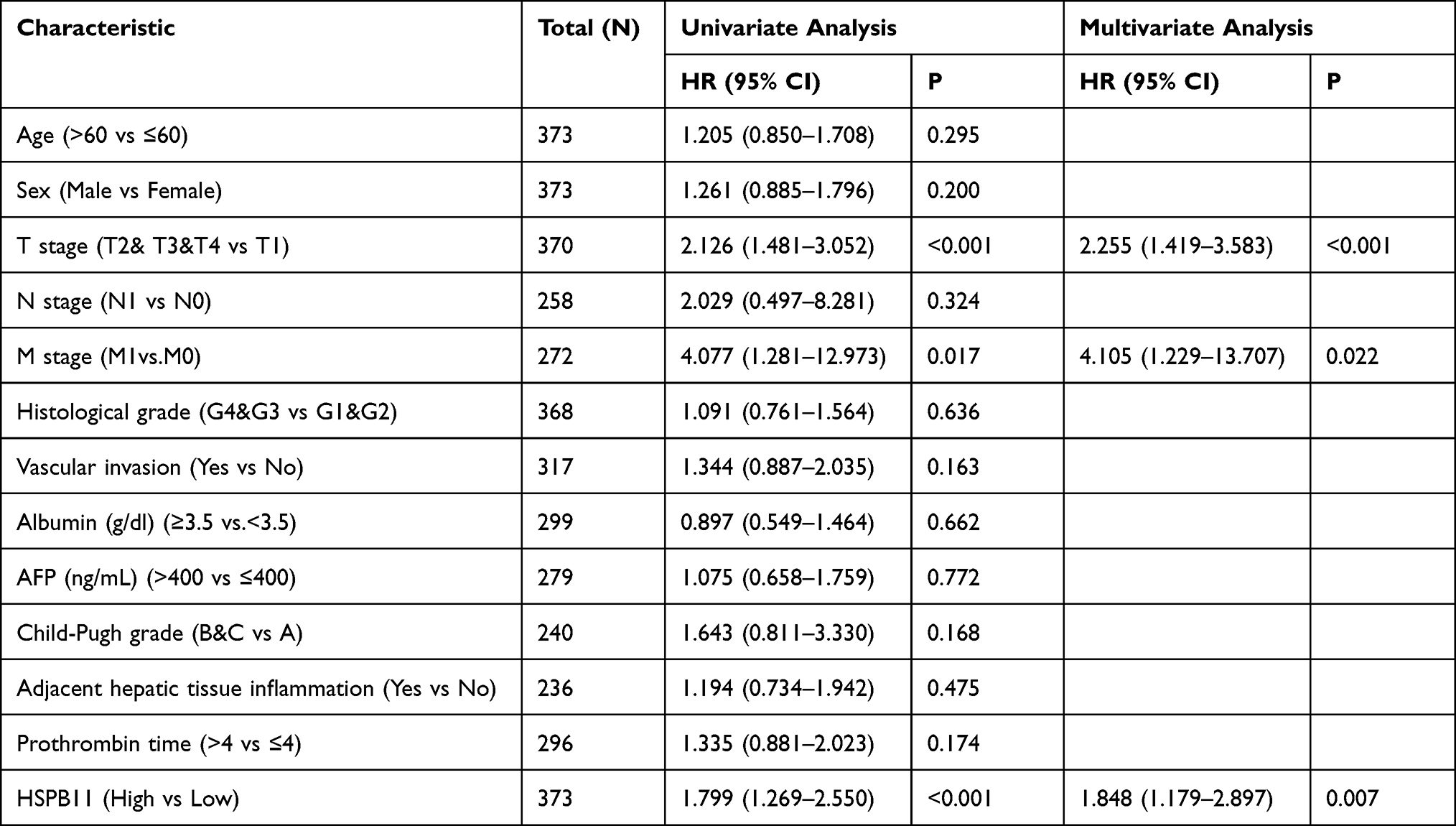 |
Table 1 Univariate and Multivariate Survival Analysis (Overall Survival) of Prognostic Covariates in Patients with Hepatocellular Carcinoma |
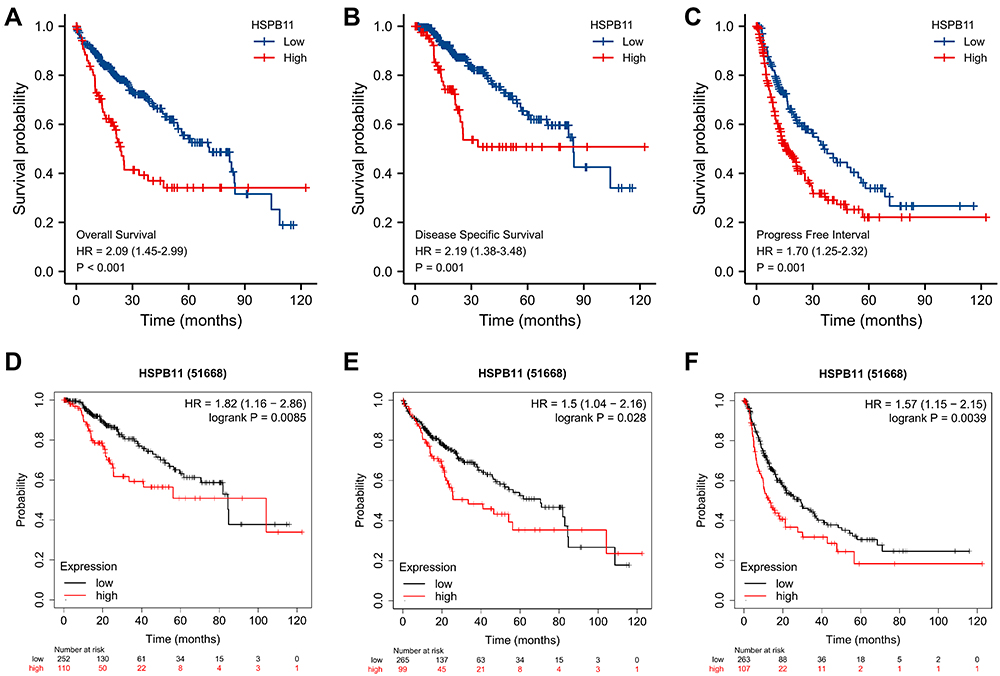 |
Figure 3 Prognostic value of the mRNA level of HSPB11 in patients with hepatocellular carcinoma (HCC). (A) Overall survival (OS), (B) disease-specific survival (DSS), and (C) progression-free interval (PFI) survival curves comparing patients with high (red) and low (blue) HSPB11 expression based on TCGA database. Analysis of the Kaplan–Meier Plotter database confirmed the poorer prognosis of HCC patients with a high HSPB11 expression level (D–F). HSPB11 expression in HCC was plotted at the threshold of p < 0.05. |
Establishment of Prognostic Models for HCC
The AUC values of the time-dependent survival ROC curve for 1-, 3-, and 5-year survival were all greater than 0.6 (Figure 4A). By integrating risk factors for prognosis identified in the Cox regression models, including tumor size, distant metastasis, and the expression level of HSPB11, nomogram and calibration models were built to predict the survival probabilities at 1, 3, and 5 years. The prediction models showed good prediction ability; the dotted line predicted by the model was close to the ideal line (Figure 4B and C).
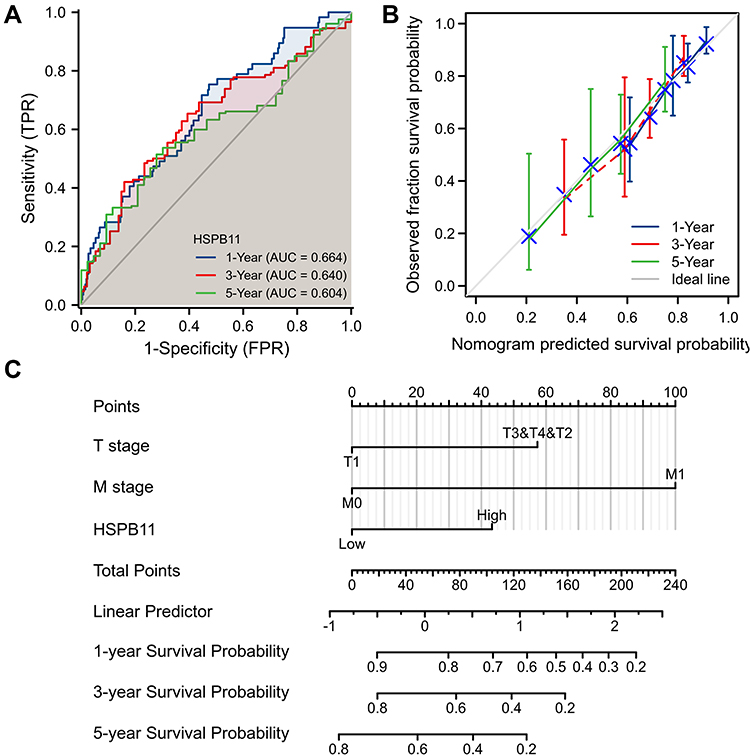 |
Figure 4 Establishment of prediction models to evaluate the probability of 1-, 3-, and 5-year overall survival (OS) of hepatocellular carcinoma patients. (A) Time-dependent survival receiver operating characteristic curve to evaluate the expression of HSPB11 for 1-, 3-, and 5-year OS. (B) Nomogram for estimating the probability of 1-, 3-, and 5-year OS. (C) Calibration plots of the nomogram for evaluating the probability of OS at 1, 3, and 5 years. |
Enrichment Analysis of HSPB11 and Associated Genes in HCC
The GSEA between the HSPB11-high and HSPB11-low patients was performed to explore HSPB11-associated pathways. The GSEA, based on h.all.v7.2.symbols.gmt, revealed that the G2M checkpoint, E2F targets, mitotic spindle, and KRAS signaling were significantly enriched. Cell cycle checkpoints, the G2M checkpoint, mitotic prometaphase, and signaling via Rho GTPases were significantly enriched based on the c2.cp.v7.2.symbols.gmt database (Figure 5).
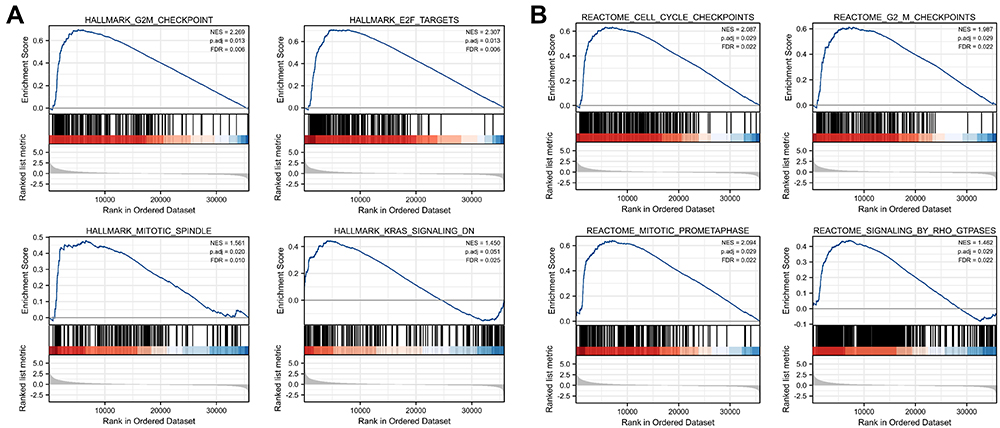 |
Figure 5 Plots from gene set enrichment analyses based on (A) h.all.v7.2symbols.gmt and (B) cp. v7.2. symbols.gmt. HSPB11 is involved in multiple oncogenic processes including the G2M checkpoint, mitotic spindle, E2F targets, KRAS signaling, cell cycle checkpoints, mitotic prometaphase, and signaling via Rho GTPase. NES, normalized enrichment score; p. adj, adjusted P value; FDR, false discovery rate. |
Association Between HSPB11 Expression and Immune Cell Infiltration
As shown in Figure 6A, the expression level of HSPB11 was correlated with several immune cells including NK cells, neutrophils, mast cells, T cells, B cells, and macrophages. Significantly, the expression of HSPB11 was positively correlated with the abundance of Th2 cells (R = 0.426, P < 0.001) and negatively associated with the abundance of DCs (R = −0.222, P < 0.001). HCC with high HSPB11 expression presented higher infiltration levels of Th2 cells and lower infiltration levels of DCs (Figure 6B–E).
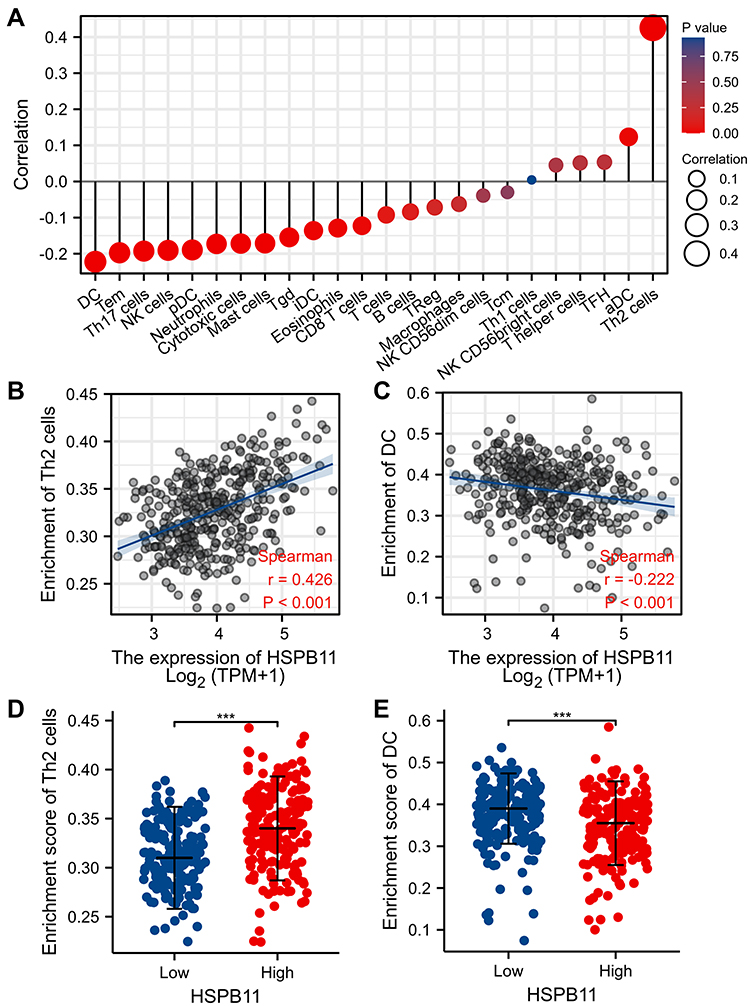 |
Figure 6 Expression level of HSPB11 is related to immune cell infiltration. (A) Association between the expression level of HSPB11 and relative abundance of 24 immune cell types. The size of dots reflects the absolute values of Spearman R. (B–E) Correlation diagrams and scatter plots indicate the differentiation of T helper (Th)2 and dendritic cell (DC) infiltration levels between HSPB11-high and HSPB11-low groups. ***p < 0.001. |
Discussion
As molecular chaperones, HSPs can protect cells to ensure survival under stress conditions, and increased expression of HSPs is observed in pathophysiological processes of cancer.33 Studies have found that HSPs can enhance the survival and aggressiveness of cancer cells under stress conditions,34 and specific inhibitors of HSPs or combined treatment with chemotherapy drugs can improve the prognosis of HCC.21 In recent years, few studies have reported that HSPB11 is overexpressed in a variety of malignant tumors.15,27–29,35 However, research on HSPB11 in HCC is insufficient. Based on data from public databases and those of clinical samples, we confirmed that HSPB11 is highly expressed in HCC, which is consistent with the findings of a previous study,15 and the expression level had a high diagnostic value. The expression level of HSPB11 was significantly correlated with a more advanced tumor stage and worse prognosis. In multivariate Cox survival analysis models, the expression level of HSPB11 was an independent risk factor for the prognosis of patients with HCC. By combining the expression levels of HSPB11 with T stage and M stage, we established a nomogram and calibration models that could be used to predict the 1-, 3-, and 5-year survival probabilities of patients. The results showed that HSPB11 has good prediction ability, and the predicted dotted line was close to the ideal line. These results indicate that HSPB11 might be a useful prognostic and diagnostic marker for HCC.
A GSEA using data from patients with HSPB11-high and HSPB11-low tumors was performed to explore HSPB11-associated functions and pathways. The results revealed that HSPB11 is involved in multiple oncogenic processes, including cell cycle checkpoints, the G2M checkpoint, E2F targets, Rho GTPase signaling, and KRAS signaling. Inactivation of cell cycle checkpoints has been reported to correlate with the progression of multiple malignancies.36,37 The G2/M checkpoint can inhibit cells with genomic DNA damage from entering the M phase, thereby facilitating the DNA damage repair process, which is critical for protection against malignant transformation.38,39 The E2F family is critical in regulating the cell cycle, proliferation, apoptosis, DNA damage signaling, and tumorigenesis. E2F1 is a proto-oncogenic gene that could induce both cell proliferation and tumor suppression by promoting apoptosis in the liver.40 Rho GTPases are involved in regulating cell proliferation, differentiation, and apoptosis, and are closely related to the infiltration and distant metastasis of liver cancer.41–46 KRAS participates in the RAS/RAF/MEK/ERK signaling pathway and is an important regulator of cell growth. Studies have confirmed KRAS mutations and activation of the Ras signaling pathway in HCC.47–49 Our GSEA revealed that HSPB11 might be involved in these signaling pathways and functions in HCC, but further studies are needed to confirm the specific mechanism.
HCC is an inflammation-induced cancer, and the immune microenvironment plays a central role in regulation of the anti-tumor immune response.50,51 In the current study, we found that the hyperexpression of HSPB11 is related to the infiltration level of immune cells in HCC. More specifically, HSPB11 was positively correlated with the abundance of Th2 cells and negatively associated with the abundance of DCs. Th2 cells secrete interleukin-4 and 10, promoting tumor growth and inducing metastasis via immunosuppression.52–55 A Th1/Th2 imbalance has been observed in HCC patients with elevated Th2-released cytokines.56 The dysfunction of DCs will result in the suppression of CD8+ T cell responses, leading to immune tolerance and cancer immunosurveillance failure.57,58 DC-based vaccines have emerged as potential cancer immunotherapeutics.59,60 Considering the significant correlations among HSPB11, Th2 cells, and DCs, we believe that HSPB11 could be involved in the immune regulation of HCC through its interaction with tumor immune cells.
The clinical data used in this study were mainly retrieved from public databases, and thus, some limitations are unavoidable. First, large clinical trials are needed to validate its diagnostic and prognostic values. Second, the mechanism through which HSPB11 affects oncogenesis and immune regulation in HCC should be confirmed through further basic molecular and animal experiments.
Conclusion
In this study, we comprehensively analyzed the role of HSPB11 in HCC. The value of HSPB11 in the diagnosis and prediction of HCC prognosis was confirmed. HSPB11 may not only be involved in the development but also in the immune regulation of HCC. However, cell line-based experiments and clinical studies are still needed to validate the potential diagnostic and prognostic value in HCC.
Abbreviations
HSPB11, heat shock protein B11; HCC, hepatocellular carcinoma; TCGA, The Cancer Genome Atlas; GTEx, Genotype-Tissue Expression (GTEx); OS, overall survival; DSS, disease-specific survival; PFI, progression-free interval; HR, hazard ratio; AUC, area under the curve; GSEA, Gene Set Enrichment Analysis; DCs, dendritic cells.
Research Ethics and Consent
The study complies with the Declaration of Helsinki, and the Ethics Committee of Second Hospital of Dalian Medical University approved this research (no. 0202159). Written informed consent was obtained from all patients before specimen collection.
Funding
This study was supported by the National Nature Science Foundation of China (81800473) and the “Young Eagle Project” of Air Force Medical University (KT2021DX007).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rahib L, Smith BD, Aizenberg R, Rosenzweig AB, Fleshman JM, Matrisian LM. Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014;74(11):2913–2921. doi:10.1158/0008-5472.CAN-14-0155
2. Parkin DM, Bray F, Ferlay J, Pisani P. Estimating the world cancer burden: globocan 2000. Int J Cancer. 2001;94(2):153–156. doi:10.1002/ijc.1440
3. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
4. Muhammad H, Tehreem A, Ting PS, et al. Hepatocellular carcinoma and the role of liver transplantation: a review. J Clin Transl Hepatol. 2019;9(5):738–748.
5. Llovet JM, Ricci S, Mazzaferro V, et al. Sorafenib in advanced hepatocellular carcinoma. N Engl J Med. 2008;359(4):378–390. doi:10.1056/NEJMoa0708857
6. Kudo M, Finn RS, Qin S, et al. Lenvatinib versus sorafenib in first-line treatment of patients with unresectable hepatocellular carcinoma: a randomised Phase 3 non-inferiority trial. Lancet. 2018;391(10126):1163–1173. doi:10.1016/S0140-6736(18)30207-1
7. Bruix J, Qin S, Merle P, et al. Regorafenib for patients with hepatocellular carcinoma who progressed on sorafenib treatment (RESORCE): a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet. 2017;389(10064):56–66. doi:10.1016/S0140-6736(16)32453-9
8. Abou-Alfa GK, Meyer T, Cheng AL, et al. Cabozantinib in patients with advanced and progressing hepatocellular carcinoma. N Engl J Med. 2018;379(1):54–63. doi:10.1056/NEJMoa1717002
9. Forner A, Reig M, Bruix J. Hepatocellular carcinoma. Lancet. 2018;391(10127):1301–1314.
10. Yim SH, Chung YJ. An overview of biomarkers and molecular signatures in HCC. Cancers. 2010;2(2):809–823. doi:10.3390/cancers2020809
11. Azad AA, Zoubeidi A, Gleave ME, Chi KN. Targeting heat shock proteins in metastatic castration-resistant prostate cancer. Nat Rev Urol. 2015;12(1):26–36. doi:10.1038/nrurol.2014.320
12. Ischia J, So AI. The role of heat shock proteins in bladder cancer. Nat Rev Urol. 2013;10(7):386–395. doi:10.1038/nrurol.2013.108
13. Zhang L, Fok JH, Davies FE. Heat shock proteins in multiple myeloma. Oncotarget. 2014;5(5):1132–1148. doi:10.18632/oncotarget.1584
14. Calderwood SK, Khaleque MA, Sawyer DB, Ciocca DR. Heat shock proteins in cancer: chaperones of tumorigenesis. Trends Biochem Sci. 2006;31(3):164–172. doi:10.1016/j.tibs.2006.01.006
15. Yang Z, Zhuang L, Szatmary P, et al. Upregulation of heat shock proteins (HSPA12A, HSP90B1, HSPA4, HSPA5 and HSPA6) in tumor tissues is associated with poor outcomes from HBV-related early-stage hepatocellular carcinoma. Int J Med Sci. 2015;12(3):256–263. doi:10.7150/ijms.10735
16. Sharma A, Upadhyay AK, Bhat MK. Inhibition of Hsp27 and Hsp40 potentiates 5-fluorouracil and carboplatin mediated cell killing in hepatoma cells. Cancer Biol Ther. 2009;8(22):2106–2113. doi:10.4161/cbt.8.22.9687
17. Yang N, Li S, Li G, et al. The role of extracellular vesicles in mediating progression, metastasis and potential treatment of hepatocellular carcinoma. Oncotarget. 2017;10(2):3683–3695. doi:10.18632/oncotarget.12465
18. Chen R, Dai RY, Duan CY, et al. Unfolded protein response suppresses cisplatin-induced apoptosis via autophagy regulation in human hepatocellular carcinoma cells. Folia Biol. 2011;57(3):87–95.
19. Lang SA, Moser C, Fichnter-Feigl S, et al. Targeting heat-shock protein 90 improves efficacy of rapamycin in a model of hepatocellular carcinoma in mice. Hepatology. 2009;49(2):523–532. doi:10.1002/hep.22685
20. Liu CC, Jan YJ, Ko BS, et al. 14-3-3σ induces heat shock protein 70 expression in hepatocellular carcinoma. BMC Cancer. 2014;14:425. doi:10.1186/1471-2407-14-425
21. Wang C, Zhang Y, Guo K, et al. Heat shock proteins in hepatocellular carcinoma: molecular mechanism and therapeutic potential. Int J Cancer. 2016;138(8):1824–1834. doi:10.1002/ijc.29723
22. Heikkila JJ. The expression and function of hsp30-like small heat shock protein genes in amphibians, birds, fish, and reptiles. Comp Biochem Physiol a Mol Integr Physiol. 2017;203:179–192. doi:10.1016/j.cbpa.2016.09.011
23. Franck E, Madsen O, Van Rheede T, Ricard G, Huynen MA, De Jong WW. Evolutionary diversity of vertebrate small heat shock proteins. J Mol Evol. 2004;59(6):792–805. doi:10.1007/s00239-004-0013-z
24. Kirbach BB, Golenhofen N. Differential expression and induction of small heat shock proteins in rat brain and cultured hippocampal neurons. J Neurosci Res. 2011;89(2):162–175. doi:10.1002/jnr.22536
25. Peferoen LA, Gerritsen WH, Breur M, et al. Small heat shock proteins are induced during multiple sclerosis lesion development in white but not grey matter. Acta Neuropathol Commun. 2015;3:87. doi:10.1186/s40478-015-0267-2
26. Zhu Z, Reiser G. The small heat shock proteins, especially HspB4 and HspB5 are promising protectants in neurodegenerative diseases. Neurochem Int. 2018;115:69–79. doi:10.1016/j.neuint.2018.02.006
27. Cheng W, Li M, Jiang Y, et al. Association between small heat shock protein B11 and the prognostic value of MGMT promoter methylation in patients with high-grade glioma. J Neurosurg. 2016;125(1):7–16. doi:10.3171/2015.5.JNS142437
28. Zoltan L, Farkas R, Schally AV, et al. Possible predictive markers of response to therapy in esophageal squamous cell cancer. Pathol Oncol Res. 2019;25(1):279–288. doi:10.1007/s12253-017-0342-z
29. Turi Z, Hocsak E, Racz B, et al. Role of mitochondrial network stabilisation by a human small heat shock protein in tumour malignancy. J Cancer. 2015;6(5):470–476. doi:10.7150/jca.11494
30. Pozsgai E, Gomori E, Szigeti A, et al. Correlation between the progressive cytoplasmic expression of a novel small heat shock protein (Hsp16.2) and malignancy in brain tumors. BMC Cancer. 2007;7:233. doi:10.1186/1471-2407-7-233
31. Farkas R, Pozsgai E, Bellyei S, et al. Correlation between tumor-associated proteins and response to neoadjuvant treatment in patients with advanced squamous-cell esophageal cancer. Anticancer Res. 2011;31(5):1769–1775.
32. Norouzinia M, Zamanian Azodi M, Najafgholizadeh Seyfi D, Kardan A, Naseh A, Akbari Z. Predication of hub target genes of differentially expressed microRNAs contributing to Helicobacter pylori infection in gastric non-cancerous tissue. Gastroenterol Hepatol Bed Bench. 2019;12(Suppl1):S44–s50.
33. Boliukh I, Rombel-Bryzek A, Żuk O, Radecka B. The role of heat shock proteins in neoplastic processes and the research on their importance in the diagnosis and treatment of cancer. Contemp Oncol. 2021;25(2):73–79.
34. Yamada N, Matsushima-Nishiwaki R, Kobayashi K, et al. Cellular Functions of Small Heat Shock Proteins (HSPB) in hepatocellular carcinoma. Curr Mol Med. 2021;21(10):872–887. doi:10.2174/1573405617666210204211252
35. Strik HM, Weller M, Frank B, et al. Heat shock protein expression in human gliomas. Anticancer Res. 2000;20(6b):4457–4462.
36. Braig M, Lee S, Loddenkemper C, et al. Oncogene-induced senescence as an initial barrier in lymphoma development. Nature. 2005;436(7051):660–665. doi:10.1038/nature03841
37. Chen Z, Trotman LC, Shaffer D, et al. Crucial role of p53-dependent cellular senescence in suppression of Pten-deficient tumorigenesis. Nature. 2005;436(7051):725–730. doi:10.1038/nature03918
38. Yin L, Chang C, Xu C. G2/M checkpoint plays a vital role at the early stage of HCC by analysis of key pathways and genes. Oncotarget. 2017;8(44):76305–76317. doi:10.18632/oncotarget.19351
39. Paulovich AG, Toczyski DP, Hartwell LH. When checkpoints fail. Cell. 1997;88(3):315–321. doi:10.1016/S0092-8674(00)81870-X
40. Huntington JT, Tang X, Kent LN, Schmidt CR, Leone G. The spectrum of E2F in liver disease–mediated regulation in biology and cancer. J Cell Physiol. 2016;231(7):1438–1449. doi:10.1002/jcp.25242
41. Stanley A, Thompson K, Hynes A, Brakebusch C, Quondamatteo F. NADPH oxidase complex-derived reactive oxygen species, the actin cytoskeleton, and Rho GTPases in cell migration. Antioxid Redox Signal. 2014;20(13):2026–2042. doi:10.1089/ars.2013.5713
42. Wong CM, Wei L, Au SL, et al. MiR-200b/200c/429 subfamily negatively regulates Rho/ROCK signaling pathway to suppress hepatocellular carcinoma metastasis. Oncotarget. 2015;6(15):13658–13670. doi:10.18632/oncotarget.3700
43. Schmidt VA. Watch the GAP: emerging roles for IQ motif-containing GTPase-activating proteins IQGAPs in hepatocellular carcinoma. Int J Hepatol. 2012;2012:958673. doi:10.1155/2012/958673
44. Brown K, Bhowmick NA. Linking TGF-beta-mediated Cdc25A inhibition and cytoskeletal regulation through RhoA/p160(ROCK) signaling. Cell Cycle. 2004;3(4):408–410. doi:10.4161/cc.3.4.778
45. Fu Y, Li J, Feng MX, et al. Cytohesin-3 is upregulated in hepatocellular carcinoma and contributes to tumor growth and vascular invasion. Int J Clin Exp Pathol. 2014;7(5):2123–2132.
46. Ng L, Kwan V, Chow A, et al. Overexpression of Pin1 and rho signaling partners correlates with metastatic behavior and poor recurrence-free survival of hepatocellular carcinoma patients. BMC Cancer. 2019;19(1):713. doi:10.1186/s12885-019-5919-3
47. Calvisi DF, Ladu S, Gorden A, et al. Ubiquitous activation of Ras and Jak/Stat pathways in human HCC. Gastroenterology. 2006;130(4):1117–1128. doi:10.1053/j.gastro.2006.01.006
48. Challen C, Guo K, Collier JD, Cavanagh D, Bassendine MF. Infrequent point mutations in codons 12 and 61 of ras oncogenes in human hepatocellular carcinomas. J Hepatol. 1992;14(2–3):342–346. doi:10.1016/0168-8278(92)90181-N
49. Tsuda H, Hirohashi S, Shimosato Y, Ino Y, Yoshida T, Terada M. Low incidence of point mutation of c-Ki-ras and N-ras oncogenes in human hepatocellular carcinoma. Jpn J Cancer Res. 1989;80(3):196–199. doi:10.1111/j.1349-7006.1989.tb02290.x
50. Hato T, Goyal L, Greten TF, Duda DG, Zhu AX. Immune checkpoint blockade in hepatocellular carcinoma: current progress and future directions. Hepatology. 2014;60(5):1776–1782. doi:10.1002/hep.27246
51. Chen KJ, Lin SZ, Zhou L, et al. Selective recruitment of regulatory T cell through CCR6-CCL20 in hepatocellular carcinoma fosters tumor progression and predicts poor prognosis. PLoS One. 2011;6(9):e24671. doi:10.1371/journal.pone.0024671
52. Schipilliti FM, Garajová I, Rovesti G, et al. The growing skyline of advanced hepatocellular carcinoma treatment: a review. Pharmaceuticals. 2021;14(1):43. doi:10.3390/ph14010043
53. Budhu A, Forgues M, Ye QH, et al. Prediction of venous metastases, recurrence, and prognosis in hepatocellular carcinoma based on a unique immune response signature of the liver microenvironment. Cancer Cell. 2006;10(2):99–111. doi:10.1016/j.ccr.2006.06.016
54. Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454(7203):436–444. doi:10.1038/nature07205
55. Lee HL, Jang JW, Lee SW, et al. Inflammatory cytokines and change of Th1/Th2 balance as prognostic indicators for hepatocellular carcinoma in patients treated with transarterial chemoembolization. Sci Rep. 2019;9(1):3260. doi:10.1038/s41598-019-40078-8
56. Ji L, Gu J, Chen L, Miao D. Changes of Th1/Th2 cytokines in patients with primary hepatocellular carcinoma after ultrasound-guided ablation. Int J Clin Exp Pathol. 2017;10(8):8715–8720.
57. Dudek AM, Martin S, Garg AD, Agostinis P. Immature, semi-mature, and fully mature dendritic cells: toward a DC-cancer cells interface that augments anticancer immunity. Front Immunol. 2013;4:438. doi:10.3389/fimmu.2013.00438
58. Harimoto H, Shimizu M, Nakagawa Y, et al. Inactivation of tumor-specific CD8+ CTLs by tumor-infiltrating tolerogenic dendritic cells. Immunol Cell Biol. 2013;91(9):545–555. doi:10.1038/icb.2013.38
59. Filley AC, Dey M. Dendritic cell based vaccination strategy: an evolving paradigm. J Neurooncol. 2017;133(2):223–235. doi:10.1007/s11060-017-2446-4
60. Constantino J, Gomes C, Falcão A, Neves BM, Cruz MT. Dendritic cell-based immunotherapy: a basic review and recent advances. Immunol Res. 2017;65(4):798–810. doi:10.1007/s12026-017-8931-1
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.