")
Back to Journals » Cancer Management and Research » Volume 12
Honokiol Suppressed Pancreatic Cancer Progression via miR-101/Mcl-1 Axis
Authors Wang Y, Liu Z, Liu Q, Han Y, Zang Y, Zhang H, Du X, Qin T, Wu Y
Received 5 November 2019
Accepted for publication 11 March 2020
Published 1 July 2020 Volume 2020:12 Pages 5243—5254
DOI https://doi.org/10.2147/CMAR.S237323
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Yishuo Wang,1,2 Zhongyong Liu,1,2 Qinrong Liu,1,2 Yongguang Han,1,2 Yuncai Zang,1,2 Huichao Zhang,1,2 Xuzhao Du,1,2 Tao Qin,3 Yuquan Wu1,2
1College of Pharmacy, Henan University of Chinese Medicine, Zhengzhou, People’s Republic of China; 2Henan Integrated Engineering Technology Research Center of Traditional Chinese Medicine Production, Zhengzhou, People’s Republic of China; 3Department of Rheumatology, Xinmi Hospital of Traditional Chinese Medicine, Xinmi, People’s Republic of China
Correspondence: Yishuo Wang
College of Pharmacy, Henan University of Chinese Medicine, Zhengzhou 450046, People’s Republic of China
Tel +86-13619847844
Email [email protected]
Background: Pancreatic cancer is one of the most aggressive malignancies. The present study aimed to examine the anti-tumor effects of honokiol in pancreatic cancer and to explore the underlying molecular mechanisms.
Materials and Methods: In vitro functional assays determined pancreatic cancer cell proliferation, apoptosis and invasion. Xenograft nude mice model determined the in vivo anti-cancer effects of honokiol. Luciferase reporter assay determined the interaction between miR101 and myeloid cell leukemia-1 (Mcl-1).
Results: Honokiol concentration-dependently suppressed pancreatic cancer cell viability. In addition, honokiol increased the caspase-3 activity and cell apoptotic rates, induced cell cycle arrest at G0/G1 phase, and inhibited cell invasion in pancreatic cancer. Interestingly, honokiol treatment induced up-regulation of miR-101 in pancreatic cancer cells. Knockdown of miR-101 attenuated the honokiol-induced cell apoptosis and inhibition in cell invasion of pancreatic cancer cells. On the other hand, miR-101 overexpression induced cell apoptosis and inhibited cell viability and invasion in pancreatic cancer. Further mechanistic study verified that Mcl-1 was negatively regulated by miR-101, and Mcl-1 overexpression counteracted the tumor-suppressive effects of honokiol on the pancreatic cancer cells. In vivo studies showed that honokiol dose-dependently suppressed tumor growth of pancreatic cancer in the nude mice and up-regulated miR-101 expression but down-regulated Mcl-1 expression in tumor tissues.
Conclusion: Our data showed that honokiol suppressed pancreatic cancer progression via miR-101-Mcl-1 axis. Honokiol could be a promising candidate for cancer prevention and/or therapeutic treatment for pancreatic cancer.
Keywords: pancreatic cancer, honokiol, miR-101, apoptosis, invasion, Mcl-1
Introduction
Pancreatic cancer is one of the most aggressive malignancies and is one of the main causes of cancer-related deaths worldwide. Pancreatic cancer has ranked the 11th most common cancer counting ~0.5 million new cases and causing ~0.4 million deaths (4.5% of all cancer-related deaths).1 The five-year survival rate after the diagnosis of pancreatic cancer is only about 4%, and 80% to 85% of patients with pancreatic cancer are at the metastatic stage when diagnosed and not suitable for surgery treatments, resulting in poor prognosis and lower survival rate.2,3 For patients with locally advanced or metastatic pancreatic cancer, gemcitabine-based combination chemotherapy is the main treatment, but most patients with pancreatic cancer developed gemcitabine resistance, which is a main obstacle for the chemotherapy treatment in pancreatic cancer.4 Therefore, there is an urgent need to develop new drugs for a better treatment of pancreatic cancer.
Studies have shown that natural products play a critical role in the discovery and the development of numerous drugs for the treatment of various cancers via different mechanisms.5–7 Honokiol is the main active ingredient of Chinese herbal medicine Magnolia, which has the diverse functions of gasification and dampness, warming and relieving pain, and reducing anti-asthma.8 Honokiol has diverse pharmacologic effects including anti-free radical and lipid peroxidation, anti-inflammatory, anti-bacterial, anti-diabetes, and cardiovascular protection. In recent years, its pharmacological activities in tumors have attracted more and more attention. Studies have shown that honokiol inhibited cancer cell invasion and epithelial–mesenchymal transition (EMT) in various types of cancer, indicating that honokiol may be as a promising active compound against various tumor malignancies including pancreatic cancer.9,10
MicroRNAs (miRNAs) are a class of small RNAs (about 20–22 bases) that are complementary to the target mRNA and degrade or repress the target mRNA translation.3,11 Recent studies have confirmed miRNAs are crucial for the development and maintenance of tumor status and act as tumor-suppressive miRNAs or oncogenic miRNAs.12–15 For example, miR-203 was identified as a oncogenic miRNA and regulated estrogen receptor-positive breast cancer growth and stemness via targeting suppressor of cytokine signaling 3;16 miR-146b promoted human bladder cancer progression via enhancing ETS2-mediated mmp2 mRNA transcription.17 Recently, numerous miRNAs such as miR-20a, miR-21, miR-24, miR-25 and miR-99a, etc. were found to be aberrantly expressed in pancreatic cancer,18 and these miRNAs are found to play important roles in regulating pancreatic cancer progression via distinct mechanisms.
In this study, we examined the anti-cancer effects of honokiol on pancreatic cancer both in vitro and in vivo. The effects of honokiol on the miRNA expression and the molecular mechanisms underlying honokiol-mediated anti-tumor effects were also explored.
Materials and Methods
Cell Culture and Transfection
The pancreatic cancer cell lines PANC-1 and SW1990 were purchased from ATCC company (Manassas, USA), and were maintained in DMEM medium (Thermo Fisher Scientific, Waltham, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone Laboratories, South Logan, USA). Cells were maintained in a humidified atmosphere with 5% CO2 at 37 °C. The mimics and inhibitors for miR-101 and the negative controls were purchased from Ribobio (Guangzhou, China). The pcDNA3.1 and the vector for overexpressing myeloid cell leukemia-1 (Mcl-1; pcDNA3.1-Mcl-1) were purchased from GenePharma (Shanghai, China). All the vectors and miRNA were transfected into cells using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, USA) according to the manufacturer’s protocol.
MTT Assay
MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay was used to determine the cell viability. PANC-1 and SW1990 cells were seeded in a 96-well plate at a density of 1500 cells/well. After an overnight incubation, various concentrations (0–200 µM) honokiol were then added. At 24 h post-incubation, the medium was removed and cells were washed with phosphate buffered saline (PBS). Then, the cells were incubated with MTT (5 mg/mL) for 3 h at 37°C. After that, dimethyl sulfoxide was added into each well to dissolve formazan crystals. Cell viability was determined by measuring optical density (OD) values at 490 nm using a microplate reader (BioTek Instruments, Winooski, USA).
Caspase-3 Activity
The caspase-3 activity of the honokiol-treated PANC-1 and SW1990 cells was measured using the Caspase-3 activity assay kit (Abcam, Cambridge, USA) according to the manufacturer’s protocol.
Cell Cycle Analysis
PANC-1 or SW1990 cells were seeded onto the 6-well plates at a density of 27 cells/well. After being treated with 50 µM honokiol or vehicle for 24 h, cells were then fixed in 70% (v/v) cold ethanol at 4°C overnight. After washing, the fixed cells were collected and re-suspended in propidium iodide (PI)/RNase Staining Buffer (Cell Cycle Detection Kit, Signalway Antibody (SAB) Co. Ltd., Maryland, USA) and cell cycle was analyzed by using a flow cytometer (BD Biosciences, San Jose, USA).
Cell Apoptosis Analysis
After being treated with 50 µM honokiol or vehicle for 24 h, PANC-1 or SW1990 cells were collected and washed twice with cold PBS and re-suspended in binding buffer. Annexin V-FITC and PI solution were then added to stain the cells prior to flow cytometry analysis.
Transwell Invasion Assay
After being treated with 50 µM honokiol or vehicle for 24 h, 4×104 PANC-1 or SW1990 cells seeded in the upper chamber with 8 μm pore membrane coated with Matrigel (BD Biosciences, USA). Serum-free medium was added to the upper chamber and full medium was added to the bottom well. After incubation at 37 °C for 24 h, the non-invaded cells on the upper surface of chamber were removed and the invaded cells in the lower surface were fixed with 4% paraformaldehyde for 10 min and were stained with 0.1% crystal violet for 5 min. The number of stained cells was counted under a light microscope by randomly selecting three fields.
Quantitative Real-Time PCR (qRT-PCR)
Total RNA from cells or tissues were extracted using Trizol reagent (Invitrogen) according to the manufacturer’s protocol. For miR-101, cDNA was synthesized using TaqMan microRNA Reverse Transcription Kit (Applied Biosystems, Foster City, USA) and PrimeScript RT Master Mix (Takara, Dalian, China) was used for mRNA reverse transcription. Then, real-time PCR reactions were performed on the ABI7900 systems (Applied Biosystems) using SYBR Green Master Mix Kit (Takara). The expression of miRNA and mRNA was normalized to U6 and β-actin, respectively, and was analysed using 2−ΔΔCt method. Primers for qRT-PCR were shown as follows: miR-101, forward, 5ʹ-GTATTTCGTAGGACAGG-3ʹ and reverse, 5ʹ-CAGTACTGTG ATAACTGAA-3ʹ; Mcl-1, forward, 5ʹ-TAAGGACAAAAC GGGACTGG-3ʹ and reverse, 5ʹ-ACCAGCTCCTACTCCAGCAA-3ʹ; GAPDH, forward, 5ʹ-TGATGACATCAAGAAGGTGGTGAAG-3ʹ and reverse, 5ʹ-TCCTTGGAGGCCATGTGGGCCAT-3ʹ.
Luciferase Reporter Assay
The wild type or mutant 3ʹ-untranslated region (3ʹ-UTR) sequence of Mcl-1was synthesized and inserted into the pGL3 reporter vector (Promega, Madison, USA). For the reporter assay, cells were plated onto 24-well plates and transfected with the above constructs and miR-101 or mimics control using Lipofectamine 2000 reagent. After 48 h, the cells were harvested and the luciferase activity was determined using the dual-luciferase reporter assay system (Promega).
Western Blot Analysis
Total protein of cells and tissues was extracted by using RIPA lysis buffer supplemented with protease inhibitors (Roche, Basel, Switzerland). Then, equal amounts of proteins were separated on a 10% SDS-PAGE and transferred to the PVDF membranes (Sigma, St. Louis, USA). After blocking with 5% non-fat milk at room temperature for 1h, the membranes were incubated with Mcl-1 primary antibody (dilution 1:500; Santa Cruz, San Diego, USA), active caspase-3 (dilution 1:1000; Santa Cruz), active caspase-9 (dilution 1:1000; Santa Cruz), Bax (dilution 1:1000; Santa Cruz) and β-actin (dilution 1:2000; Cell Signaling Technology, Danvers, USA) overnight at 4 °C. After washing, the membranes were again incubated with horseradish peroxidase-conjugated secondary antibodies (dilution 1:2500; Cell Signaling Technology). The bands were visualized by ECL Detection kit (HANNOTECH Biosciences, Dongguan, China).
In vivo Tumor Xenograft Study
Six-week-old male athymic nude mice were used for in vivo study and the study was approved by the Animal Ethics Committee of Henan University of Chinese Medicine and the experiments were performed in accordance with the National Guidelines for Experimental Animal Welfare (the Ministry of Science and Technology, China). A 5 × 106 cells in 0.1 mL 50% Matrigel were injected subcutaneously into the right flank of nude mice. Seven days after cell inoculation, the animals with palpable tumors were randomly divided into three experimental groups: (1) honokiol, 20 mg/kg in 20% Intralipid; (2) honokiol, 40 mg/kg in 20% Intralipid; and (3) vehicle control, 20% Intralipid. The nude mice were subjected to intraperitoneal injections every other day for 5 weeks. Tumor volume was measured every week. At the endpoint, the mice were sacrificed and tumors were weighed and processed for further analysis.
Statistical Analysis
All the data analysis was performed using GraphPad Prism V5.0 (GraphPad Software, La Jolla, USA). The data from this study were presented as mean ± standard deviation. Significant differences between/among different treatment groups were evaluated by unpaired Student’s t-test or One-way ANOVA followed by Bonferroni’s post-hoc test. P < 0.05 was considered to be statistically significant.
Results
Effects of Honokiol on the Cell Viability of Pancreatic Cancer Cells
The effects of honokiol on the cell viability were examined in PANC-1 and SW1990 cells. Different concentrations of honokiol (0–200 µM) were used to treat PANC-1 or SW1990 cells. At 24 h after treatment, MTT assay results showed that honokiol significantly reduced cell viability in a concentration-dependent manner (Figure 1A and B). Since 50 µM honokiol caused more than 50% suppression of the viability, we chose this concentration for further study.
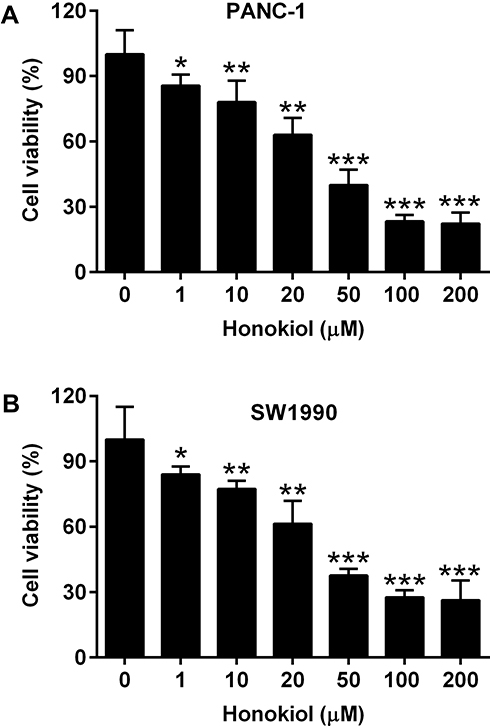 |
Figure 1 Effects of Honokiol on the cell viabilities of pancreatic cancer cells. (A) PANC-1 and (B) SW1990 cells were treated with Honokiol (1–200 µM) for 24 h, and cell viability of these cells was determined by MTT assay. N = 3. *P<0.05, **P<0.01 and ***P<0.001. |
Effects of Honokiol on the Cell Apoptosis, Cell Cycle and Cell Invasion of Pancreatic Cancer Cells
Cell apoptosis is a normal physiological process and caspase-3 enzyme is a member of the family that regulates apoptosis signaling networks.19 Treatment with 50 µM honokiol for 24 h markedly enhanced the caspase-3 activity in both PANC-1 and SW1990 cells (Figure 2A and B). Western blot results showed that 50 µM honokiol treatment increased the protein levels of active caspase-3, caspase-9 and Bax in both PANC-1 and SW1990 cells (Figure 2C and D). Cell apoptosis and cell cycle analysis results showed that 50 µM honokiol resulted in increase of apoptotic rates and the percentage of cells in the G0/G1 phase. Moreover, the S-phase cells showed an obvious decrease after honokiol treatment (Figure 2E–H). These data indicated that cell cycle distribution was blocked in the G0/G1 phase. Furthermore, the invasive activity of treated PANC-1 and SW1990 cells was also examined and 50 µM honokiol caused markedly decline when compared with the untreated cells (Figure 2I–J).
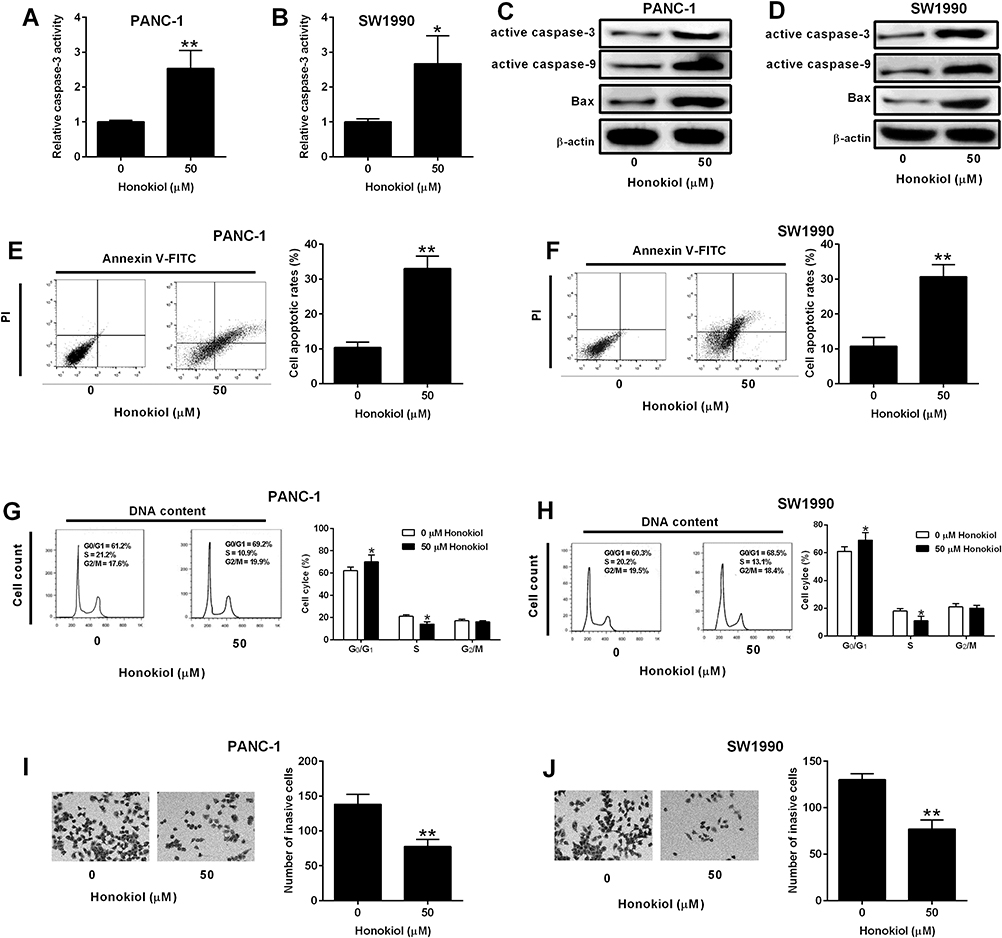 |
Figure 2 Effects of Honokiol on the cell apoptosis, cell cycle and cell invasion of pancreatic cancer cells. (A, B) Caspase-3 activity of PANC-1 and SW1990 cells after being treated with 50 µM Honokiol or vehicle for 24 h was determined by Caspase-3 activity Assay Kit. (C, D) Active caspase-3, caspase-9 and Bax protein levels of PANC-1 and SW1990 cells after being treated with 50 µM Honokiol or vehicle for 24 h was determined by Western blot assay. (E–H) Flow cytometry determined cell apoptotic rates (E, F) and cell cycle (G, H) of PANC-1 and SW-1990 cells after being treated with 50 µM Honokiol or vehicle for 24 h. (I, J) Transwell invasion assay determined cell invasive ability of PANC-1 and SW1990 cells after being treated with 50 µM Honokiol or vehicle for 24 h. N = 3. *P<0.05 and **P<0.01. |
Honokiol Suppressed Pancreatic Cell Viability, Apoptosis and Invasion via Regulating miR-101 Expression
Previous studies showed that miR-101 inhibited pancreatic cancer cell migration and invasion.20,21 To examine whether miR-101 is required for honokiol-induced anti-cancer effects, we have checked the expression level of miR-101 in honokiol-treated cells. Honokiol-induced concentration-dependently an increase of miR-101 level in both PANC-1 and SW1990 cells. On the other hand, miR-101 inhibitors transfected down-regulated miR-101 expression markedly (Figure 3A–D). Silence of miR-101 promoted cell viability and invasive abilities, while blocked caspase 3, thus decreasing the apoptosis rate (Figure 3E–L). The rescue experiments showed under 50 µM honokiol treatment, miR-101 inhibition could partially restore the anti-cancer effects caused by honokiol (Figure 3E–L).
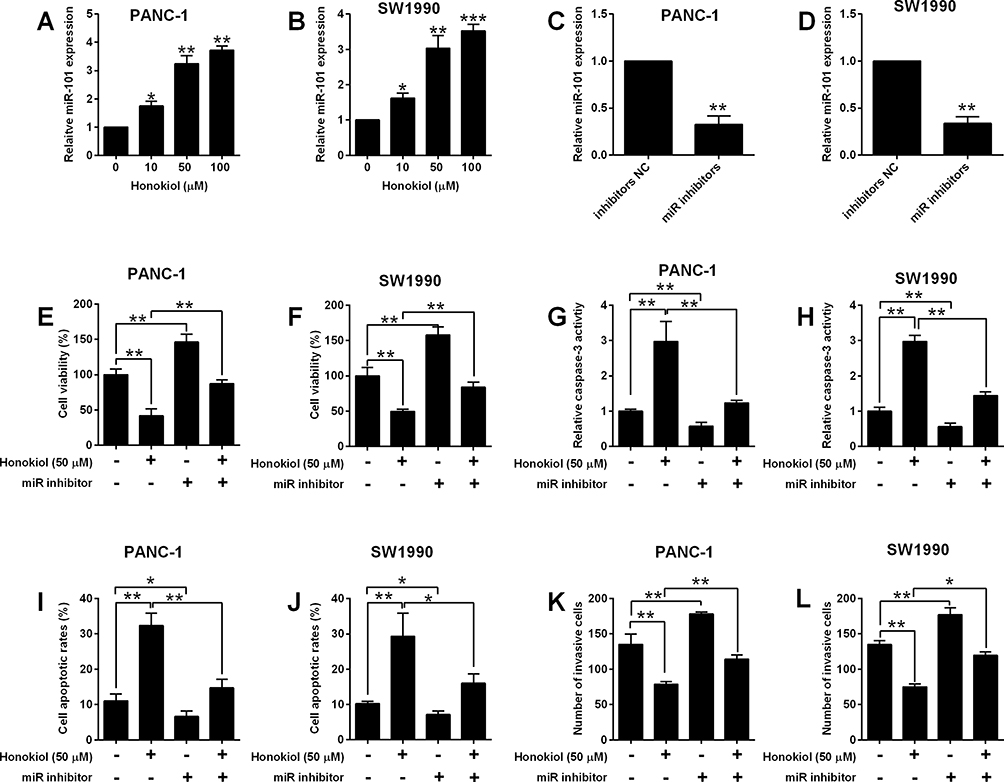 |
Figure 3 Honokiol suppressed pancreatic cell viability, apoptosis and invasion via regulating miR-101 expression. (A, B) MiR-101 expression in PANC-1 and SW1990 cells after being treated 10, 50 and 100 µM Honokiol or vehicle was determined by qRT-PCR. (C, D) MiR-101 expression in PANC-1 and SW1990 cells after being transfected with inhibitors NC or miR inhibitors. (E–L) PANC-1 and SW1990 cells that were treated with Honokiol (50 µM) or vehicle were transfected with miR inhibitors or inhibitors NC, and 24 h later, cell viability was determined by MTT assay (E, F), cell apoptosis was evaluated with Caspase-3 Activity Assay Kit (G, H) and flow cytometry (I, J), and cell invasive ability was assessed with Transwell invasion assay (K, L). N = 3. *P<0.05, **P<0.01 and ***P<0.001. |
Effects of miR-101 Overexpression on Cell Viability, Apoptosis and Invasion of Pancreatic Cancer Cells
To explore the anti-cancer effects of miR-101 in pancreatic cancer, miR-101 mimics was transiently transfected into PANC-1 and SW1990 cells, which increased the expression of more than 10 folds (Figure 4A–B). Consistently, overexpression of miR-101 reduced cell viability increased the caspase 3 activity and apoptotic rates as well as inhibited cell invasion in both PANC-1 and SW1990 cells (Figure 4C–J).
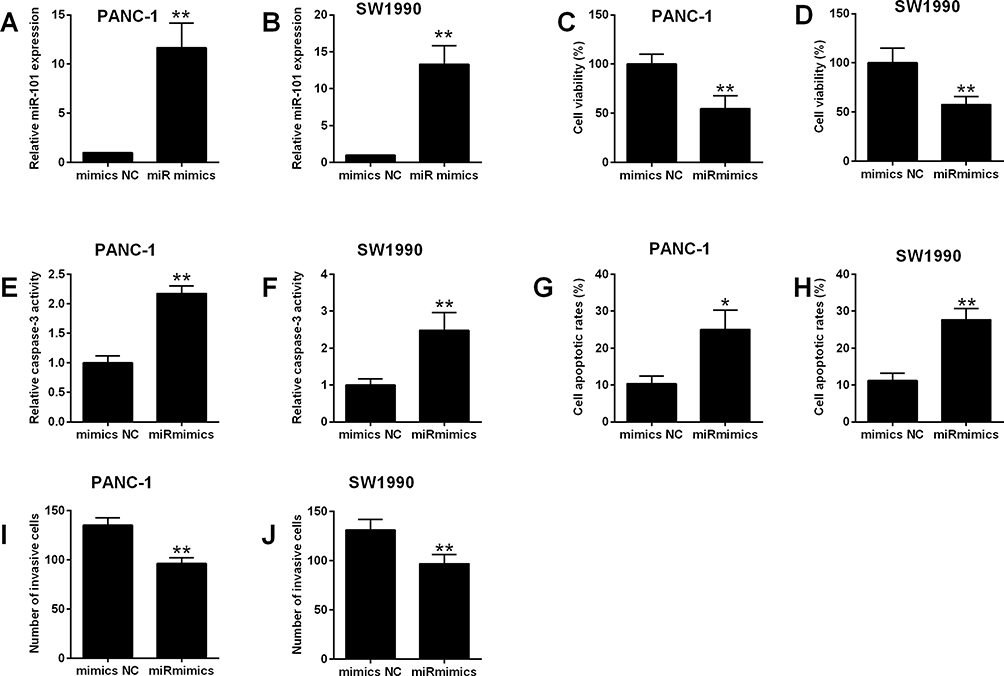 |
Figure 4 Effects of miR-101 overexpression on cell viability, apoptosis and invasion of pancreatic cancer cells. (A, B) PANC-1 and SW1990 cells were transfected with mimics NC or miR mimics, and 24 h later, miR-101 expression in these transfected cells was determined by qRT-PCR. (C–J) PANC-1 and SW1990 cells were transfected with mimics NC or miR mimics, and 24 h later, cell viability was determined by MTT assay (C, D), cell apoptosis was evaluated with Caspase-3 Activity Assay Kit (E, F) and flow cytometry (G, H), and cell invasive ability was assessed with Transwell invasion assay (I, J). N = 3. *P<0.05 and **P<0.01. |
MiR-101 Targeted Mcl-1 3ʹUTR and Inversely Regulated the Expression of Mcl-1
The screening results from online databases suggested that miR-101 may target the 3′-UTR of Mcl-1. To verify this, Luciferase reporter assays were performed. After dual transfection, the luciferase activity of wild type reporter vector was significantly downregulated by miR-101 mimics. On the other hand, the mimics failed to affect the luciferase activity of the mutant one (Figure 5A–C). Furthermore, miR-101 mimics suppressed Mcl-1 expression at both mRNA and protein levels, while the inhibitor led to a surge in Mcl-1 expression. Rescue experiments showed that under honokiol treatment, the Mcl-1 expression was decreased, which was partially restored by miR-101 inhibitor transfection (Figure 5D–G).
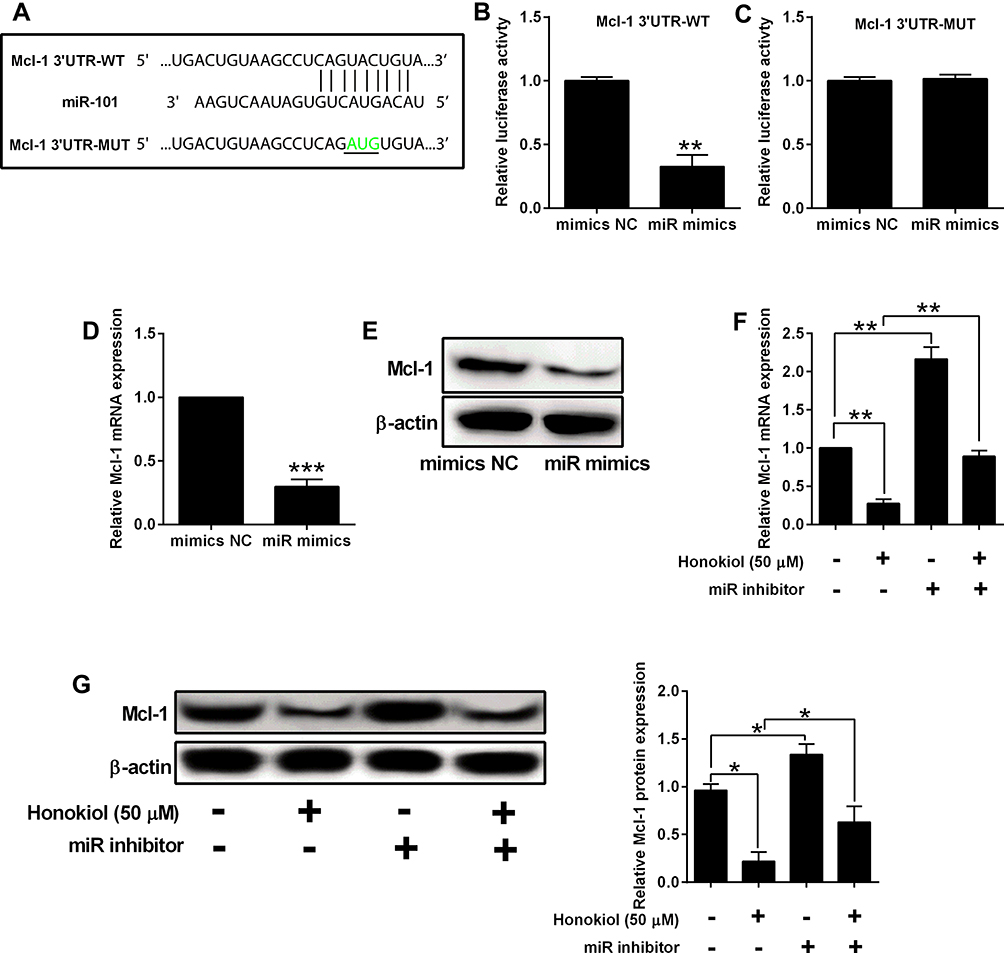 |
Figure 5 MiR-101 targeted Mcl-1 3ʹUTR and inversely regulated the expression of Mcl-1. (A) Predicted binding sequences between miR-101 and Mcl-1 3ʹUTR. The nucleotides highlighted in green colour were indicated as mutated sites. (B, C) Panc-1 cells were co-transfected with miRNAs (mimics NC or miR mimics) and reporter vectors (Mcl-1 3ʹUTR-WT or Mcl-1 3ʹUTR-MUT), and 48 h later, relative luciferase activity was evaluated by Dual-Luciferase Reporter Assay System. (D, E) PANC-1 cells were transfected with mimics NC or miR mimics, and 24 h later, the mRNA and protein expression of Mcl-1 was determined by qRT-PCR and Western blot assay, respectively. (F, G) PANC-1 and SW1990 cells that were treated with Honokiol (50 µM) or vehicle were transfected with miR inhibitors or inhibitors NC, and 24 h later, the mRNA and protein expression of Mcl-1 was determined by qRT-PCR and Western blot assay, respectively. N = 3. *P<0.05, **P<0.01 and ***P<0.001. |
Mcl-1 Overexpression Counteracted Tumor-Suppressive Effects of Honokiol on Pancreatic Cancer Cells
To test the role of Mcl-1 in pancreatic cancer, we overexpressed Mcl-1 in the rescue experiments. Overexpression of Mcl-1 in both mRNA and protein levels was achieved by transfecting cells with pcDNA3.1-Mcl-1 (Figure 6A and B). Under honokiol treatment, cell viability and invasive ability were decreased, and caspase 3 activity and apoptosis were enhanced. In addition, overexpressing Mcl-1 partially resorted to the tumor-suppressive effects of honokiol (Figure 6C–F).
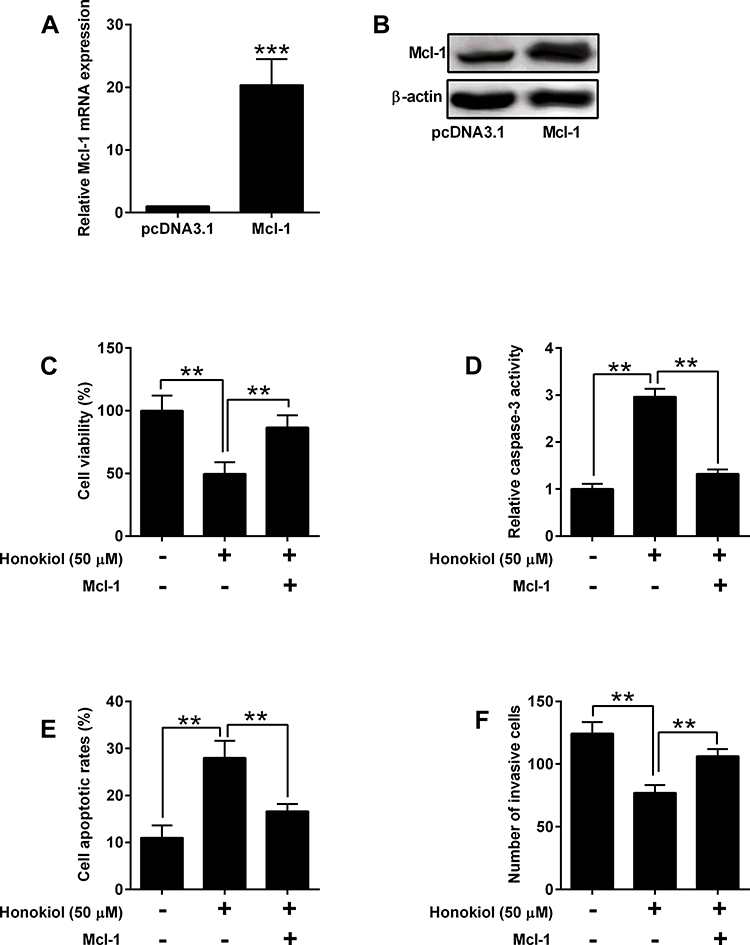 |
Figure 6 Mcl-1 overexpression counteracted tumor-suppressive effects of Honokiol on pancreatic cancer cells. (A, B) PANC-1 cells were transfected with pcDNA3.1 or pcDNA3.1-Mcl-1, and 24 h later, the mRNA and protein expression of Mcl-1 was determined by qRT-PCR and Western blot, respectively. (C–F) PANC-1 and SW1990 cells that were treated with Honokiol (50 µM) or vehicle were transfected with pcDNA3.1 or pcDNA3.1-Mcl-1, and 24 h later, cell viability was determined by MTT assay (C), cell apoptosis was evaluated with Caspase-3 Activity Assay Kit (D) and flow cytometry (E), and cell invasive ability was assessed with Transwell invasion assay (F). N = 3. **P<0.01 and ***P<0.001. |
Honokiol Suppressed in vivo Tumor Growth of PANC-1 Cells
To evaluate the anti-tumor effects of honokiol in vivo, xenografts were generated in the nude mice. After 5-week treatment, the tumor progression of both 20 mg/kg and 40 mg/kg groups was markedly slower than that of vehicle control group and also presented as a dose-dependent effect. Tumor weight in the honokiol treatment groups was also lower than that of control group (Figure 7A–B). Honokiol had no effect on the body weight of the nude mice (Figure 7C). Furthermore, the expression level of miR-101 and Mcl-1 were determined in the tumor tissues and honokiol up-regulated the miR-101 expression and down-regulated Mcl-1 expression dose-dependently (Figure 7D and E).
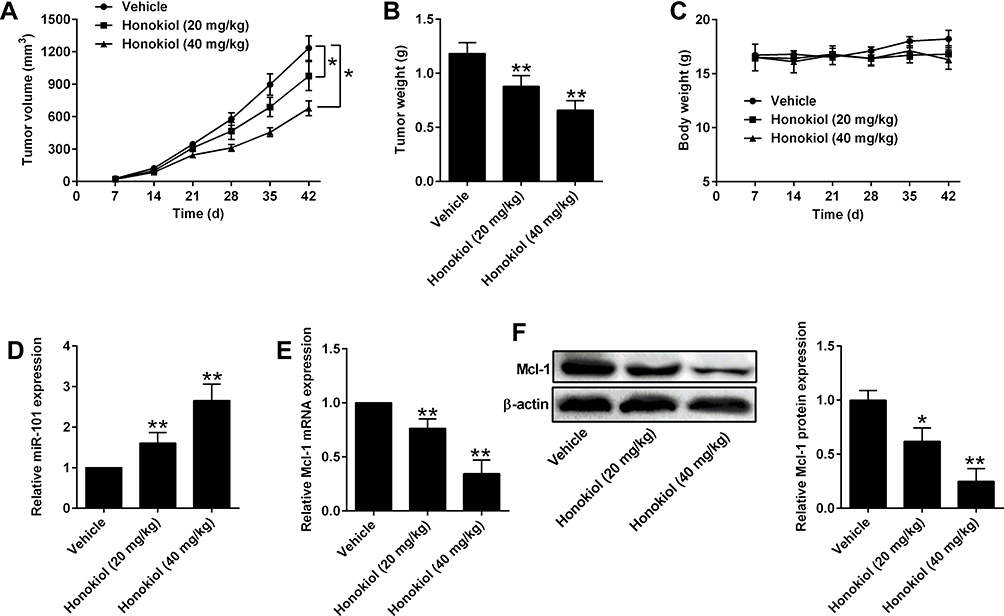 |
Figure 7 Honokiol suppressed in vivo tumor growth of PANC-1 cells. (A) Tumor growth curve and (B) tumor weight from vehicle or Honokiol (20 mg/kg and 40 mg/kg) treatment groups. (C) Body weight of the nude mice from vehicle or Honokiol (20 mg/kg and 40 mg/kg) treatment groups. (D) MiR-101 expression in harvested tumor tissues was determined by qRT-PCR. (E, F) The mRNA and protein expression of Mcl-1 in harvested tumor tissues was determined by qRT-PCR and Western blot, respectively. N = 5. *P<0.05 and **P<0.01. |
Discussion
With the continuous development of new technologies, the treatment of pancreatic cancer has made great progress. However, the 5-year survival rate has remained unchanged and very low. Therefore, it is necessary to uncover novel therapeutics or alternative treatments for pancreatic cancer. In this study, we have examined the anticancer effects of honokiol in pancreatic cancer cells. The study demonstrated that honokiol decreased cell viability and invasive ability, induced cell apoptosis cell cycle arrest at G0/G1 phase in pancreatic cancer cells. Furthermore, miR-101/Mcl-1 axis was proved to contribute to honokiol-induced anticancer effects.
Epigenetic modification is crucial for cancer mechanisms. miRNA-mediated mRNA regulation is considered to be a key factor involved in cancer progression. MiRNAs can act as oncogenes, tumor suppressor genes, and regulators of cancer stem cells and metastasis. With an in-depth understanding of miRNA-targeted genes and their effects on cells, regulating miRNA expression may provide an exciting opportunity for cancer treatment. Previous studies reported that miR-101 acted as a tumor suppressor in various types of cancers.22–25 In the pancreatic cancer studies, miR-101 suppressed pancreatic cancer cell EMT via targeting high mobility group AT-hook 2;21 miR-101 could target stathmin 1 to suppress pancreatic cancer cell proliferation and invasion.20 Moreover, miR-101 sensitized pancreatic cancer cells to gemcitabine by silencing DNA-PKcs.26 Consistently, miR-101 acted as a tumor suppressor miRNA in pancreatic cancer in this study. Overexpression of miR-101 reduced cell viability increased the caspase 3 activity and apoptosis rate as well as inhibited cell invasion. To explore the mechanism of miR-101 involved in honokiol-induced anticancer effects, we had screened its target through on-line software. Mcl-1 was found to be the potential target and Luciferase reporter assays demonstrated the direct interaction between miR-101 and Mcl-1. Furthermore, miR-101 down-regulated Mcl-1 expression.
As the downstream target of miR-101, Mcl-1 exerts anti-apoptotic function. Its overexpression has been reported in lung cancer, ovarian cancer, breast cancer and pancreatic cancer.27–29 In breast cancer, high Mcl-1 level was associated with poor prognosis and inhibition of Mcl-1 restricted the growth of breast cancer xenografts in vivo.29 Apart from anti-apoptotic actions, Mcl-1showed resistance ability against the antitumor agents, thus limiting the efficiency of antitumor drugs such as taxol and cisplatin.30,31 In non-small cell lung cancer, Mcl-1 was upregulated in cisplatin-resistant cancer cells. Inhibition of Mcl-1 molecule sensitized cisplatin’s effects and induced apoptotic death of lung cancer cells.32 In our study, Mcl-1 overexpression counteracted tumor-suppressive effects of honokiol on pancreatic cancer cells. Therefore, silence of Mcl-1 might be a promising approach for cancer treatment.
Based on the above findings, the anti-cancer effects of honokiol were through miR-101-Mcl-1 axis. However, silence of miR-101 or Mcl-l partially inhibited honokiol-induced effects, which indicated that there might be other regulators or mechanisms. Recent studies reported honokiol inhibits cancer metastasis by blocking EMT through modulation of Snail/Slug protein translation or induced autophagic apoptosis through a P53-dependent pathway,10,33 and the effects of honokiol on other signaling pathways may require further investigations. In addition, further studies may examine if miR-101 can affect the in vivo tumor growth of the nude mice received honokiol treatment, which may consolidate our current findings.
Conclusion
In summary, our data showed that honokiol suppressed pancreatic cancer progression via miR-101-Mcl-1 axis (see Figure 8 for the summarized diagram). Honokiol may be a promising candidate for chemoprevention and/or therapeutic treatment for pancreatic cancer treatment.
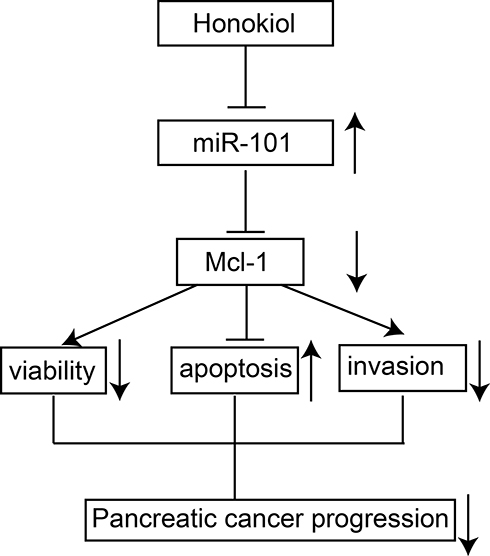 |
Figure 8 A diagram of the honokiol-mediated tumor-suppressive actions in pancreatic cancer. |
Abbreviations
3ʹUTR, 3ʹ-untranslated region; EMT, epithelial–mesenchymal transition; FBS, fetal bovine serum; Mcl-1, myeloid cell leukemia-1; miRNA, microRNA; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; OD, optical density; PBS, phosphate buffered saline.
Data Sharing Statement
All the data in the manuscript are available upon reasonable request.
Author Contributions
YW and ZL designed the whole study; ZL, QL, YH, YZ and HZ performed the experiments and analyzed the data; XD, TQ and YW performed the statistical analysis; YW wrote the manuscript; all authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Hidalgo M, Cascinu S, Kleeff J, et al. Addressing the challenges of pancreatic cancer: future directions for improving outcomes. Pancreatology. 2015;15(1):8–18. doi:10.1016/j.pan.2014.10.001
3. Falasca M, Kim M, Casari I. Pancreatic cancer: current research and future directions. Biochim Biophys Acta. 2016;1865(2):123–132. doi:10.1016/j.bbcan.2016.01.001
4. Marasini B, Sahu RP. Natural anti-cancer agents: implications in gemcitabine-resistant pancreatic cancer treatment. Mini Rev Med Chem. 2017;17(11):920–927. doi:10.2174/1389557517666170315124438
5. Muhammad N, Steele R, Isbell TS, Philips N, Ray RB. Bitter melon extract inhibits breast cancer growth in preclinical model by inducing autophagic cell death. Oncotarget. 2017;8(39):66226–66236. doi:10.18632/oncotarget.19887
6. Bhattacharya S, Muhammad N, Steele R, Kornbluth J, Ray RB. Bitter melon enhances natural killer-mediated toxicity against head and neck cancer cells. Cancer Prev Res (Phila). 2017;10(6):337–344. doi:10.1158/1940-6207.CAPR-17-0046
7. Bhattacharya S, Muhammad N, Steele R, Peng G, Ray RB. Immunomodulatory role of bitter melon extract in inhibition of head and neck squamous cell carcinoma growth. Oncotarget. 2016;7(22):33202–33209. doi:10.18632/oncotarget.8898
8. Lin D, Yan Z, Chen A, et al. Anti-proliferative activity and structure-activity relationship of honokiol derivatives. Bioorg Med Chem. 2019;27(16):3729–3734. doi:10.1016/j.bmc.2019.06.042
9. Zhang J, Zhang Y, Shen W, et al. Cytological effects of honokiol treatment and its potential mechanism of action in non-small cell lung cancer. Biomed Pharmacother. 2019;117:109058. doi:10.1016/j.biopha.2019.109058
10. Wang WD, Shang Y, Li Y, Chen SZ. Honokiol inhibits breast cancer cell metastasis by blocking EMT through modulation of snail/slug protein translation. Acta Pharmacol Sin. 2019;40(9):1219–1227. doi:10.1038/s41401-019-0240-x
11. Previdi MC, Carotenuto P, Zito D, Pandolfo R, Braconi C. Noncoding RNAs as novel biomarkers in pancreatic cancer: what do we know? Future Oncol. 2017;13(5):443–453. doi:10.2217/fon-2016-0253
12. Neureiter D, Jager T, Ocker M, Kiesslich T. Epigenetics and pancreatic cancer: pathophysiology and novel treatment aspects. World J Gastroenterol. 2014;20(24):7830–7848. doi:10.3748/wjg.v20.i24.7830
13. Li Y, Sarkar FH. MicroRNA targeted therapeutic approach for pancreatic cancer. Int J Biol Sci. 2016;12(3):326–337. doi:10.7150/ijbs.15017
14. Chitkara D, Mittal A, Mahato RI. miRNAs in pancreatic cancer: therapeutic potential, delivery challenges and strategies. Adv Drug Deliv Rev. 2015;81:34–52. doi:10.1016/j.addr.2014.09.006
15. Vorvis C, Koutsioumpa M, Iliopoulos D. Developments in miRNA gene signaling pathways in pancreatic cancer. Future Oncol. 2016;12(9):1135–1150. doi:10.2217/fon-2015-0050
16. Muhammad N, Bhattacharya S, Steele R, Ray RB. Anti-miR-203 suppresses ER-positive breast cancer growth and stemness by targeting SOCS3. Oncotarget. 2016;7(36):58595–58605. doi:10.18632/oncotarget.11193
17. Zhu J, Xu C, Ruan L, Wu J, Li Y, Zhang X. MicroRNA-146b overexpression promotes human bladder cancer invasion via enhancing ETS2-mediated mmp2 mRNA transcription. Mol Ther Nucleic Acids. 2019;16:531–542. doi:10.1016/j.omtn.2019.04.007
18. Abreu FB, Liu X, Tsongalis GJ. miRNA analysis in pancreatic cancer: the Dartmouth experience. Clin Chem Lab Med. 2017;55(5):755–762. doi:10.1515/cclm-2017-0046
19. Modi S, Kir D, Banerjee S, Saluja A. Control of apoptosis in treatment and biology of pancreatic cancer. J Cell Biochem. 2016;117(2):279–288. doi:10.1002/jcb.25284
20. Zhu L, Chen Y, Nie K, Xiao Y, Yu H. MiR-101 inhibits cell proliferation and invasion of pancreatic cancer through targeting STMN1. Cancer Biomark. 2018;23(2):301–309. doi:10.3233/CBM-181675
21. Jiang W, Gu W, Qiu R, et al. miRNA-101 suppresses epithelial-to-mesenchymal transition by targeting HMGA2 in pancreatic cancer cells. Anticancer Agents Med Chem. 2016;16(4):432–439. doi:10.2174/1871520615666150507122142
22. Wang CZ, Deng F, Li H, et al. MiR-101: a potential therapeutic target of cancers. Am J Transl Res. 2018;10(11):3310–3321.
23. Teiten MH, Dicato M, Diederich M. Curcumin as a regulator of epigenetic events. Mol Nutr Food Res. 2013;57(9):1619–1629. doi:10.1002/mnfr.201300201
24. Lima CR, Gomes CC, Santos MF. Role of microRNAs in endocrine cancer metastasis. Mol Cell Endocrinol. 2017;456:62–75. doi:10.1016/j.mce.2017.03.015
25. Bao B, Li Y, Ahmad A, et al. Targeting CSC-related miRNAs for cancer therapy by natural agents. Curr Drug Targets. 2012;13(14):1858–1868. doi:10.2174/138945012804545515
26. Hu H, He Y, Wang Y, Chen W, Hu B, Gu Y. microRNA-101 silences DNA-PKcs and sensitizes pancreatic cancer cells to gemcitabine. Biochem Biophys Res Commun. 2017;483(1):725–731. doi:10.1016/j.bbrc.2016.12.074
27. Akgul C. Mcl-1 is a potential therapeutic target in multiple types of cancer. Cell Mol Life Sci. 2009;66(8):1326–1336. doi:10.1007/s00018-008-8637-6
28. Reiner T, de Las Pozas A, Parrondo R, et al. Mcl-1 protects prostate cancer cells from cell death mediated by chemotherapy-induced DNA damage. Oncoscience. 2015;2(8):703–715. doi:10.18632/oncoscience.231
29. Campbell KJ, Dhayade S, Ferrari N, et al. MCL-1 is a prognostic indicator and drug target in breast cancer. Cell Death Dis. 2018;9(2):19. doi:10.1038/s41419-017-0035-2
30. Balko JM, Giltnane JM, Wang K, et al. Molecular profiling of the residual disease of triple-negative breast cancers after neoadjuvant chemotherapy identifies actionable therapeutic targets. Cancer Discov. 2014;4(2):232–245. doi:10.1158/2159-8290.CD-13-0286
31. Wertz IE, Kusam S, Lam C, et al. Sensitivity to antitubulin chemotherapeutics is regulated by MCL1 and FBW7. Nature. 2011;471(7336):110–114. doi:10.1038/nature09779
32. Zhang F, Shen M, Yang L, et al. Simultaneous targeting of ATM and Mcl-1 increases cisplatin sensitivity of cisplatin-resistant non-small cell lung cancer. Cancer Biol Ther. 2017;18(8):606–615. doi:10.1080/15384047.2017.1345391
33. Lin MC, Lee YW, Tseng YY, et al. Honokiol induces autophagic apoptosis in neuroblastoma cells through a P53-dependent pathway. Am J Chin Med (Gard City N Y). 2019;47(4):895–912. doi:10.1142/S0192415X19500472
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.