")
Back to Journals » Drug Design, Development and Therapy » Volume 14
High ACTN1 Is Associated with Poor Prognosis, and ACTN1 Silencing Suppresses Cell Proliferation and Metastasis in Oral Squamous Cell Carcinoma
Authors Xie G, Zhao L, Chen Q , Tang D , Chen Q , Lu H , Cai J , Chen Z
Received 2 January 2020
Accepted for publication 31 March 2020
Published 5 May 2020 Volume 2020:14 Pages 1717—1727
DOI https://doi.org/10.2147/DDDT.S244516
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qiongyu Guo
Guo-feng Xie,1,* Luo-dan Zhao,2,3,* Qiang Chen,4,* Dong-xiao Tang,4 Qiong-yu Chen,4 Hong-fei Lu,4 Jia-rong Cai,5 Zheng Chen4
1Department of Radiation Oncology, Affiliated Cancer Hospital & Institute of Guangzhou Medical University, Guangzhou, Guangdong Province 510095, People’s Republic of China; 2Department of Oral and Maxillofacial Surgery, Sun Yat-sen Memorial Hospital, Sun Yat-sen University, Guangzhou, Guangdong Province, People’s Republic of China; 3Key Laboratory of Malignant Tumor Gene Regulation and Target Therapy of Guangzhou Higher Education Institutes, Sun Yat-sen University, Guangzhou, Guangdong Province, People’s Republic of China; 4Department of Stomatology, The Third Affiliated Hospital of Sun Yat-sen University, Guangzhou 510630, Guangdong Province, People’s Republic of China; 5Department of Urology, The Third Affiliated Hospital of Sun Yat-sen University, Guangzhou 510630, Guangdong Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jia-rong Cai
Department of Urology, The Third Affiliated Hospital of Sun Yat-sen University, No. 600 Tianhe Road, Guangzhou, Guangdong Province, People’s Republic of China
Tel +86-20-82179727
Fax +86-20-85253336
Email [email protected]
Zheng Chen
Department of Stomatology, The Third Affiliated Hospital of Sun Yat-sen University, No. 600 Tianhe Road, Guangzhou, Guangdong Province, People’s Republic of China
Fax +86-20-85253336
Email [email protected]
Purpose: Oral squamous cell carcinoma (OSCC) is a common malignancy of the oral cavity. As the survival rate of OSCC patients is low, it is crucial to explore new markers and therapeutic targets for early diagnosis of the disease. A high level of actinin alpha 1 (ACTN1) in patients could serve as an independent prognostic factor of acute myeloid leukemia. However, the role of ACTN1 in OSCC remains unclear. In the present study, we aimed to investigate the role of ACTN1 in OSCC.
Methods: ACTN1 protein levels in tissues were determined by immunohistochemical (IHC) staining. The correlation of ACTN1 expression with clinicopathological features and prognosis was analyzed. Univariate and multivariate analyses were performed. The effect of ACTN1 knockdown on cell proliferation, migration, invasion, apoptosis, epithelial-mesenchymal transition (EMT), and the cell cycle was evaluated using Western blotting, Cell Counting Kit‑8 (CCK8) assays, flow cytometry analysis, transwell assays, wound-healing assays, and nude mouse models of subcutaneous xenograft and pulmonary metastasis.
Results: Based on the total score of ACTN1 IHC staining analysis, ACTN1 expression was found to be low in 10 normal mucosal tissues, 48 normal mucosal tissues adjacent to OSCC, and 19 OSCC tissues, but high in 29 OSCC tissues. ACTN1 protein levels were significantly associated with the clinical stage and node metastasis, and a high ACTN1 protein level indicated poor prognosis. Moreover, ACTN1 expression was an independent predictor of poor prognosis of OSCC. Using in vitro assays, we found that ACTN1 knockdown could induce cell cycle arrest, promote apoptosis, and inhibit EMT and cell proliferation, migration, and invasion in the OSCC cell lines, SCC-15 and HSC-3. Moreover, ACTN1 knockdown inhibited subcutaneous tumor growth and pulmonary metastasis in vivo.
Conclusion: ACTN1 levels were significantly associated with the clinical stage and node metastasis, and a high ACTN1 protein level indicated poor prognosis. Moreover, ACTN1 knockdown could suppress cell proliferation and metastasis of OSCC. Our results suggested that ACTN1 may serve as a diagnostic and prognostic marker of OSCC.
Keywords: actinin alpha 1, poor prognosis, oral squamous cell carcinoma, knockdown, proliferation, metastasis
Introduction
Oral cancer is a malignant neoplasm of the oral cavity.1 Oral cancer is one of the 10 most common cancers, and a significant public health threat worldwide.1 More than 90% of oral cancers are estimated to be oral squamous cell carcinoma (OSCC).2,3 Although local OSCC can be effectively controlled by surgical excision and radiotherapy, metastasis to the lymph nodes and other distant organs significantly decreases the survival rate.4 The cause of the low survival rate may be attributed to a delay in early diagnosis or lack of specific markers to predict tumor development and patient prognosis. Therefore, it is necessary to explore the key factors and elucidate the mechanisms involved in the occurrence and development of OSCC to identify new markers and therapeutic targets of OSCC diagnosis and provide a theoretical basis for the early diagnosis, prognostic judgment, and development of personalized therapeutic targets of OSCC.
Alpha actinins (ACTNs) are major cytoskeletal proteins and perform important non-muscle functions, such as regulation of cytokinesis, cell adhesion, and migration, besides mediating sarcomere function.5,6 Four isoforms of ACTNs are known in mammals: ACTN1, ACTN2, ACTN3, and ACTN4. ACTN4 plays a key role in promoting tumorigenesis, metastasis, and epithelial-mesenchymal transition (EMT).7 Gene amplification of ACTN4 is a potential prognostic biomarker of the overall survival (OS) of oral tongue cancer patients,8 and promotes metastasis in gastric cancer.9 However, few studies have reported the roles of ACTN1, ACTN2, and ACTN3 in cancer cells. High ACTN1 and ACTN3 expression levels are associated with shorter event-free survival (EFS) and OS of patients with acute myeloid leukemia (AML). In these patients, ACTNs serve as independent poor prognostic factors.10 These results indicate that ACTNs may be important factors in the occurrence and developmental progression of malignant tumors. However, their role in OSCC remains elusive.
In the present study, we aimed to investigate the role of ACTN1 in OSCC. Firstly, we analyzed ACTN1 expression in OSCC tissues and the correlation of ACTN1 expression with the clinicopathological features and prognosis. Finally, the effects of ACTN1 knockdown on OSCC cells were evaluated using in vitro and in vivo models.
Patients and Methods
Collection of OSCC Tissues and Follow-Up Information
Forty-eight OSCC patients, who were diagnosed at the Third Affiliated Hospital of Sun Yat-sen University from 2014 to 2018, were enrolled in this study according to the inclusion and exclusion criteria. The inclusion criteria were: 1) clinically and pathologically diagnosed as OSCC; 2) age 18–90 years; 3) no immune system disease or HIV infection; and 4) not received any anticancer treatments for at least 3 months before the study. Exclusion criteria were: 1) age below 18 or above 90 years; 2) pregnant or nursing women; 3) having severe heart, lung, liver, kidney, or other systemic diseases; 4) other tumors or mental disorder; 5) having diseases of the immune system or HIV infection; and 6) have received anticancer treatment 3 months before the study. OSCC tissues (n = 48) and clinically normal mucosal tissues adjacent to the OSCC (at a distance of about 2 cm, n = 48) were obtained during surgeries. The detailed clinical information of all enrolled OSCC patients is provided in Table 1. Survival time was recorded by conducting regular follow-up. Ten normal mucosal samples were obtained from patients who received surgical treatment for trauma. All patients understood the purpose of our study and signed the informed consent. Our study was approved by the ethics committee of the Third Affiliated Hospital of Sun Yat-sen University (No.[2020]02-017-01).
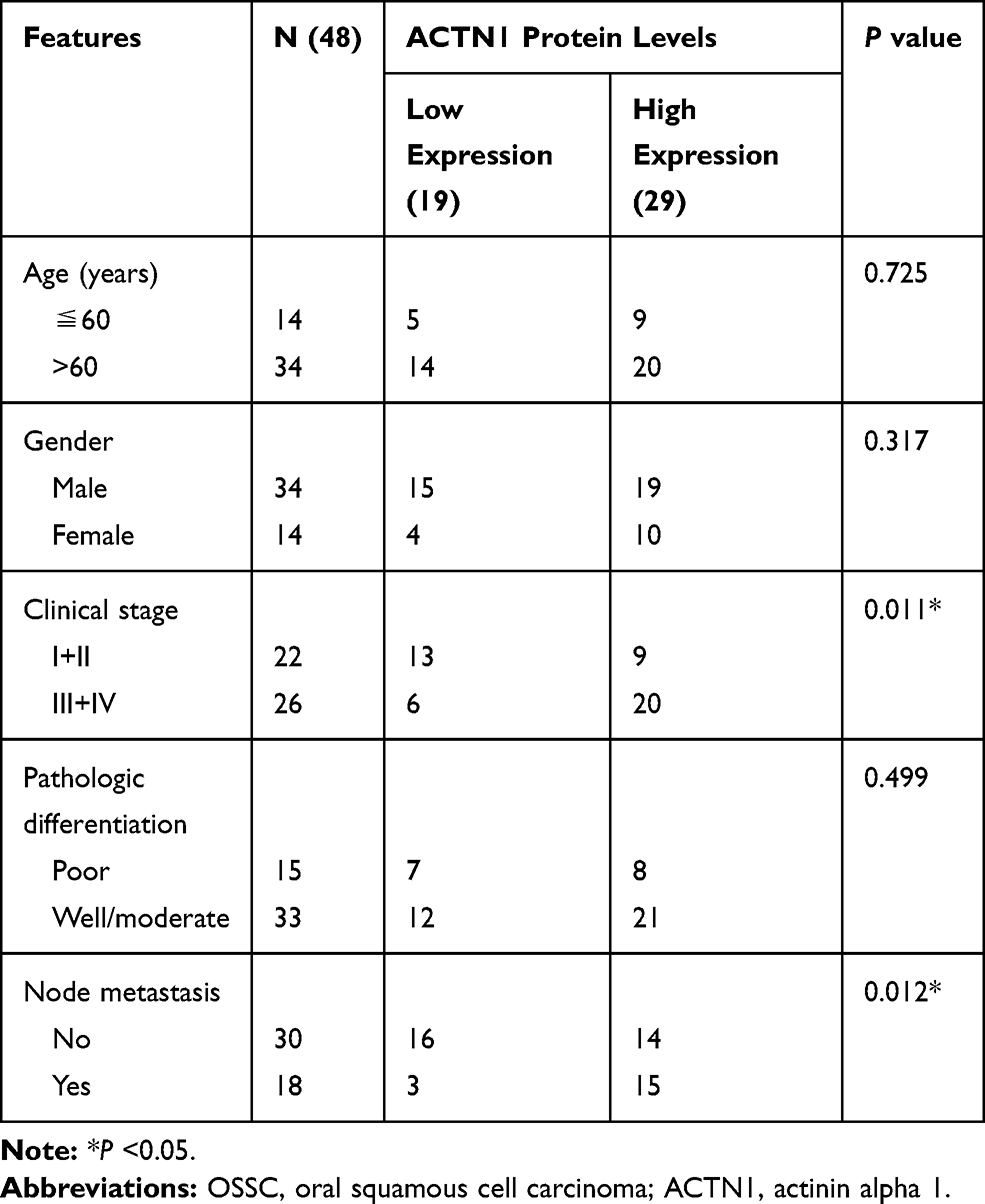 |
Table 1 Correlation Between the ACTN1 Protein Levels and Clinicopathological Features of Patients with OSCC |
Immunohistochemical (IHC) Staining and Analysis
All collected tissues were embedded in paraffin and cut into sections (4 µm). The ACTN1 expression level in all sections was examined by IHC analysis. Briefly, dewaxed sections were blocked with goat serum at 22–25°C for 30 min, after antigen repair in sodium citrate buffer (pH 6.0, ≥92~95°C) in a microwave oven for 15 min. Following this, sections were incubated with anti-ACTN1 antibodies (dilution, 1:100; ab68194, Abcam, Cambridge, MA, USA) at 4°C overnight. The next day, these sections were incubated with secondary antibodies at 37°C for 30 min. Finally, the sections were incubated in 2% 3,3′-diaminobenzidine (DAB) chromogenic solution at 22–25°C for 15 min. After mounting, the ACTN1 expression levels in all sections were judged independently by two senior pathologists, according to the following method. The total score of ACTN1 staining = staining intensity score (0: negative; 1: light yellow; 2: brown; 3: tan) × percentage of positive cells (0: <5%; 1: 5%-25%; 2: 25%-50%; 3: 51%–75%: 4: >75%). The total scores of the results of ACTN1 staining were grouped into three categories- a score of 1–4 was regarded as low expression, 5–8 as moderate expression, and 9–12 as high expression.
Bioinformatics Analysis
The ACTN1 expression in head and neck squamous cell carcinoma (HNSC) tissues and the relationship between OS of HNSC patients and ACTN1 expression levels were analyzed using gene expression profiling interactive analysis (GEPIA). This is a web-based tool used to deliver fast and customizable functionalities based on The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx) data.11
Cell Culture
Human keratinocytes of the HaCaT and human OSCC cell lines (SCC-15, HSC-3, OSC-19, and HSC-4) were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). All the culture media were prepared according to the instructions from the supplier. Cells were cultured at 37°C in a 5% CO2 incubator.
Construction of ACTN1 Stable Knockdown Cells
The short hairpin RNA (shRNA, 5ʹgatccCCTCAGGAGATCAATGGCAAACTCGAGTTTGCCATTGATCTCCTGAGGTTTTTg) that targets ACTN1 (shACTN1) was designed and cloned into an L202 plasmid (and named L202-shACTN1). L202-shACTN1, PMD2.G, and pSPAX2 were transfected into 293T cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) to package lentiviruses that express shACTN1 (lv-shACTN1). Following this, lv-shACTN1 were produced. Empty plasmids L202, PMD2.G, and pSPAX2 were transfected into 293T cells to package negative control lentiviruses (lv-NC). SCC-15 and HSC-3 cells were transfected with lv-shACTN1 or lv-NC (multiplicity of infection = 50) and screened by puromycin. Screened ACTN1 stable knockdown cells were named SCC-15–shACTN1 and HSC-3-shACTN1. lv-NC transfected cells were named SCC-15-NC and HSC-3-NC.
Western Blotting
SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells were harvested, and the total protein was isolated with RIPA buffer. After protein quantification using a BCA Protein Assay kit (Thermo Scientific Pierce, Rockford, IL, USA), total protein (30 µg) was loaded for SDS-PAGE electrophoresis and transferred onto a PVDF membrane. After being blocked with 5% (w/v) nonfat dried milk in tris buffered saline (TBS), membranes were incubated with diluted primary antibodies (anti-caspase 3, 1:5000; anti-caspase 9, 1:3000; anti-bax, 1:3000; anti-Bcl2, 1:2000; anti-E-cadherin, 1:500; anti-vimentin, 1:2000; anti-ACTN1, 1:1000; all purchases were made from Abcam, Cambridge, MA, USA). After washing with TBS containing 1‰ Tween (TBST) thrice for 5 min, membranes were incubated with secondary antibodies (Goat Anti-Rabbit IgG(H+L)-HRP, 1:20,000; Rabbit Anti-Mouse IgG(H+L)-HRP, 1:10,000; all purchased were made from Southern Biotech, Birmingham, AL, USA). After washing with TBST thrice for 5 min, membranes were incubated with Immobilon Western Chemilum HRP Substrate (Millipore Corporation, Billerica, MA, USA).
Cell Counting Kit‑8 (CCK8) Assay
SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells (1×104) were seeded into 96‐well plates. After 24, 48, and 72 h of cell seeding, CCK8 solution (10 µL) (Beyotime Institute of Biotechnology, Shanghai, China) was added into each well. After incubating for 4 h, the optical density was detected at 450 nm using a measured Multiskan MK3 microplate reader (Thermo Fisher Scientific).
Flow Cytometry Analysis for Apoptosis and the Cell Cycle
SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells (1×106) were harvested for apoptosis and cell cycle assays. Apoptosis detection was performed using the Annexin V‐FITC/PI apoptosis detection kit (KeyGen Biotech, Co, Ltd, Nanjing, China). Cell cycle analysis was performed using a Cell Cycle Analysis Kit (KeyGen Biotech, Co, Ltd).
Transwell Assay
For transwell migration assay, SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells (1 × 105) were suspended in serum-free medium (200 µL) and seeded into the upper chamber of a 12-well transwell plate (Becton Dickinson, Franklin Lakes, NJ, USA). Culture medium (600 µL), with 20% fetal bovine serum, was added into the lower chamber. After 24 h of incubation, cells in the upper chamber were removed with a cotton swab, and cells on the underside were fixed with 4% paraformaldehyde for 15 min. After being washed with PBS, cells were stained with 0.1% crystal violet for 10 min. After being washed again with PBS, stained cells were randomly photographed under a phase contrast microscope. The cell number was counted using Image Pro-Plus 6.0 software (Media Cybernetics, Rockville, MD, USA), and the average number of cells per field was calculated from five fields. For invasion, the upper chambers were pre-coated with Matrigel (Becton Dickinson), while the other procedures were conducted in a manner similar to that of the transwell migration assay.
Wound-Healing Assay
Straight lines were drawn at the bottom of 6-well plates, and SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells (5 × 105) were seeded into the wells. On the 2nd day, a wound was generated along the straight line, using a sterile pipette (200 µL) tip. After removing the fallen cells by washing the plates with PBS, cells were cultured in a serum-free medium. The wound was photographed at 6, 24, and 48 h. The wound area was analyzed using Image Pro-Plus 6.0 software (Media Cybernetics). The percentage of wound closure was calculated using the formula: 1-wound area 6 h, 24 h, or 48 h ÷ wound area 0 h × 100%.
Construction of Nude Mouse Models of Subcutaneous Xenograft and Pulmonary Metastasis
Six-week-old athymic nude mice (n = 24) were purchased from Beijing Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China). After adaptive feeding for 3 days, all mice were randomly divided into eight groups (n = 3). All animal experiments in this study were approved by the ethics committee of the Third Affiliated Hospital of Sun Yat-sen University (No.00207797), in accordance with the Guide for the Care and Use of Laboratory Animals published by the National Institute of Health (NIH Publication No 85–23, revised 1996).
To construct subcutaneous xenograft models, SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells (4×106 cells in 0.2 mL of PBS) were subcutaneously injected in the right armpit region of different nude mice. After injection for 7, 10, 13, 16, 19, 22, 25, 28, 31, 34, and 37 days, tumor length and width were measured using calipers, and tumor volume was calculated using the formula: 1/2×L×W2 (L is the length of the tumor; W is the width of the tumor).
To construct pulmonary metastasis models, SCC-15-NC, SCC-15–shACTN1, HSC-3-NC, and HSC-3-shACTN1 cells (5 × 105) were re-suspended in Hank’s Balanced Salt Solution (100 µL) and injected intravenously into the tails of nude mice (n = 4). Six weeks later, all mice were euthanized by administration of an intraperitoneal injection of sodium pentobarbital (130 mg/kg). Lung tissues were isolated for H&E staining to evaluate the status of lung metastasis.
Statistical Analysis
The results are presented as mean ± standard deviation. Statistical analysis was performed using SPSS 19.0 software (IBM, Chicago, IL, USA). Statistical comparison between the two groups was analyzed using t-tests. Statistical analysis for more than two groups was performed using one-way ANOVA, followed by post-hoc LSD test. The significance of the correlation of ACTN1 expression with clinicopathological features was determined by a Pearson χ2 test. OS was analyzed using the Kaplan–Meier method. Receiver operating characteristic (ROC) analysis was conducted using GraphPad Prism version 7.0 (GraphPad Software, San Diego, CA, USA). A value of P <0.05 was considered statistically significant.
Results
ACTN1 Expression Level in OSCC and Normal Tissues
First, the expression profiles of ACTN1 mRNA in GEPIA were analyzed. As shown in Figure 1A, ACTN1 mRNA expression was significantly higher in HNSC tissues than in normal tissues. Following this, we evaluated the ACTN1 protein levels in 48 OSCC tissues and 48 normal mucosal tissues adjacent to OSCC using IHC analysis. Representative pictures of ACTN1 expression in normal mucosal tissues, OSCC tissues, and normal mucosal tissues adjacent to OSCC are shown in Figure 1B. Based on the total score of ACTN1 staining analyzed by two senior pathologists, we found that ACTN1 was expressed at low levels in 10 normal mucosal tissues from healthy volunteers, 48 normal mucosal tissues adjacent to OSCC, and 19 OSCC tissues, whereas it was highly expressed in 29 OSCC tissues. As shown in Figure 1C, the total score of ACTN1 staining was significantly higher in OSCC tissues than in normal mucosal tissues or normal mucosal tissues adjacent to OSCC (P <0.0001).
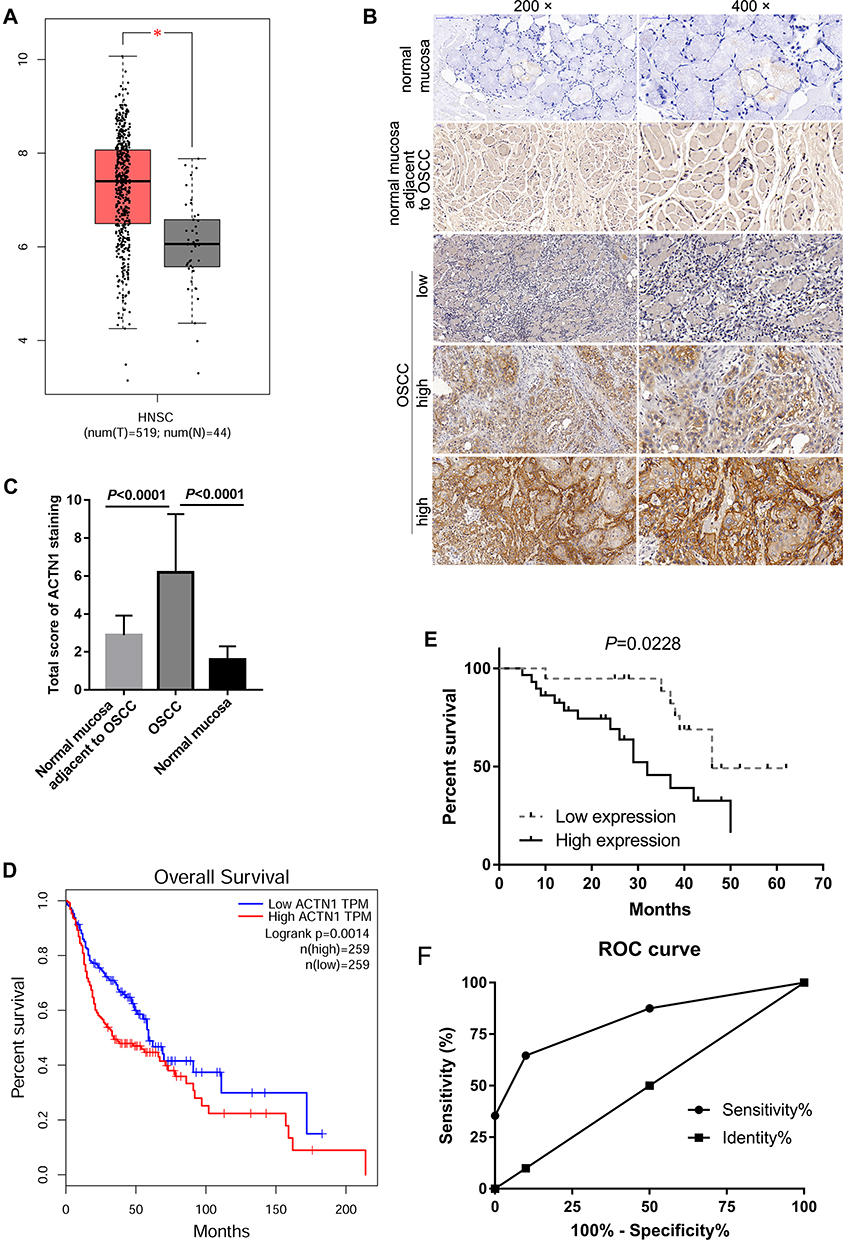 |
Figure 1 The expression level and prognostic significance of actinin alpha 1 (ACTN1) in patients with oral squamous cell carcinoma (OSCC). (A) ACTN1 mRNA levels were higher in head and neck squamous cell carcinoma (HNSC) tissues than in normal tissues. GEPIA, a web-based tool to deliver fast and customizable functionalities based on The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx) data, was performed. T: tumor, red bar; N: normal, grey bar. *P <0.05. (B) Representative pictures of ACTN1 expression in normal mucosal tissues, OSCC tissues, and normal mucosal tissues adjacent to OSCC. (C) Statistical result of the total score of ACTN1 immunohistochemical (IHC) staining analyzed by two senior pathologists. (D) Correlation between the ACTN1 mRNA levels and overall survival time of patients with OSCC, as analyzed by GEPIA. (E) Correlation between the ACTN1 protein levels and overall survival time of patients with OSCC, based on our IHC results and follow-up information. (F) Receiver operating characteristic (ROC) analysis curve analyzed by GraphPad Prism. |
Clinicopathological and Prognostic Significance of ACTN1 in OSCC Patients
We analyzed the correlation between ACTN1 mRNA expression and OS on the basis of GEPIA. As shown in Figure 1B, ACTN1 mRNA levels were significantly associated with OS time, and a high ACTN1 mRNA level indicated poor prognosis (Figure 1D). Furthermore, we analyzed the relationship between the ACTN1 protein levels and OS time on the basis of IHC results and follow-up information. The median survival time in the low and high expression groups was 46 months and 32 months, respectively (Figure 1E). These results revealed that the ACTN1 protein level was significantly associated with the OS time, and a high ACTN1 protein level indicated poor prognosis (Figure 1E). Furthermore, we analyzed the performance of ACTN1 in diagnosing OSCC using ROC curve analysis. Area under the ROC curve was 0.8362. At the optimal expression cutoff value of 4.5, the sensitivity was 60.42% and specificity was 100% (Figure 1F). Next, we analyzed the correlation between the ACTN1 protein levels and clinicopathological features of OSCC patients. The results showed that the ACTN1 protein levels were not significantly associated with age, gender, and pathologic differentiation, but with clinical stage and node metastasis (Table 1). Moreover, univariate and multivariate analyses were performed to determine whether ACTN1 expression and the clinicopathological parameters (Table 1) were independent prognostic parameters of OSCC patient outcomes. The results indicated that ACTN1 expression was an independent predictor of poor prognosis of OSCC (Table 2).
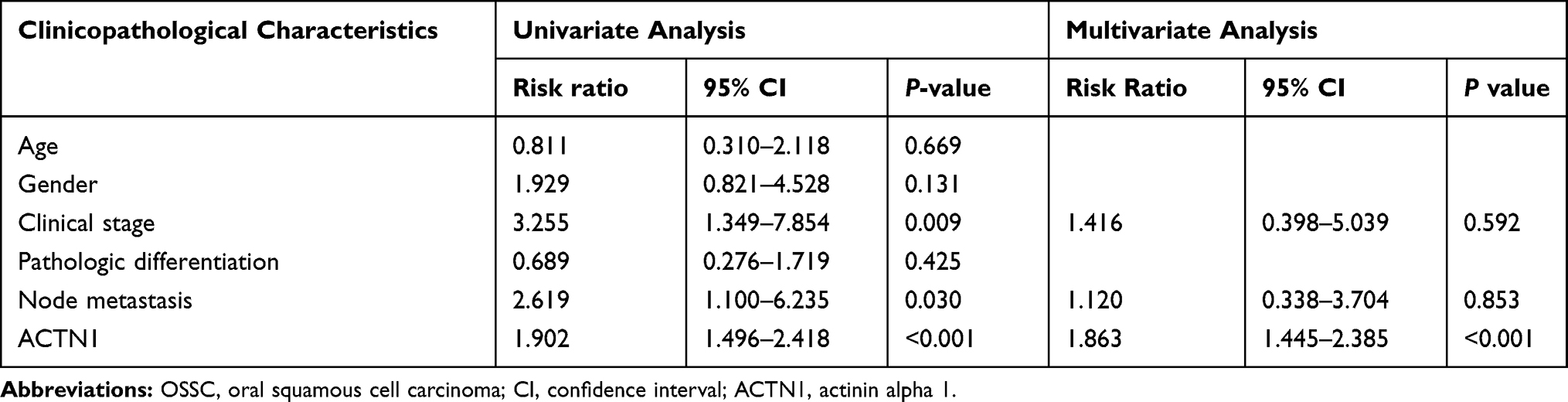 |
Table 2 Univariate and Multivariate Cox Proportional Hazard Models for Overall Survival of OSCC Patients |
ACTN1 Knockdown Inhibited Cell Proliferation, Induced Cell Cycle Arrest, and Promoted Apoptosis in vitro
As shown in Figure 2A, ACTN1 expression was higher in the OSCC cell lines SCC-15, HSC-3, OSC-19, and HSC-4, than in HaCaT, and the highest expression was found in SCC-15 and HSC-3. Following this, ACTN1 in SCC-15 and HSC-3 cells was stably knocked down by transfecting these cells with lv-shACTN1. As shown in Figure 2B, ACTN1 levels in the shACTN1 group cells decreased compared to those in the NC group cells, indicating that the ACTN1 knockdown cells were successfully constructed. Following this, we evaluated the effect of ACTN1 knockdown on cell proliferation, apoptosis, and the cell cycle. As shown in Figure 2C, the proliferation rate of the shACTN1 group cells was lower than that of the NC group cells, after culturing for 2, 3, and 4 days. As shown in Figure 2D, the total cellular apoptotic rate of the shACTN1 group cells was higher than that of the NC group cells. In addition, cleaved caspase 3, cleaved caspase 9, and Bax levels were lower and Bcl-2 level was higher in the shACTN1 group cells than in the NC group cells (Figure 2E). Moreover, the percentage of cells in the G1 stage was higher whereas in the S and G2 stages was lower in the shACTN1 group than in the NC group (Figure 2F). Therefore, ACTN1 knockdown could inhibit cell proliferation, induce cell cycle arrest, and promote apoptosis in OSCC cell lines, SCC-15 and HSC-3.
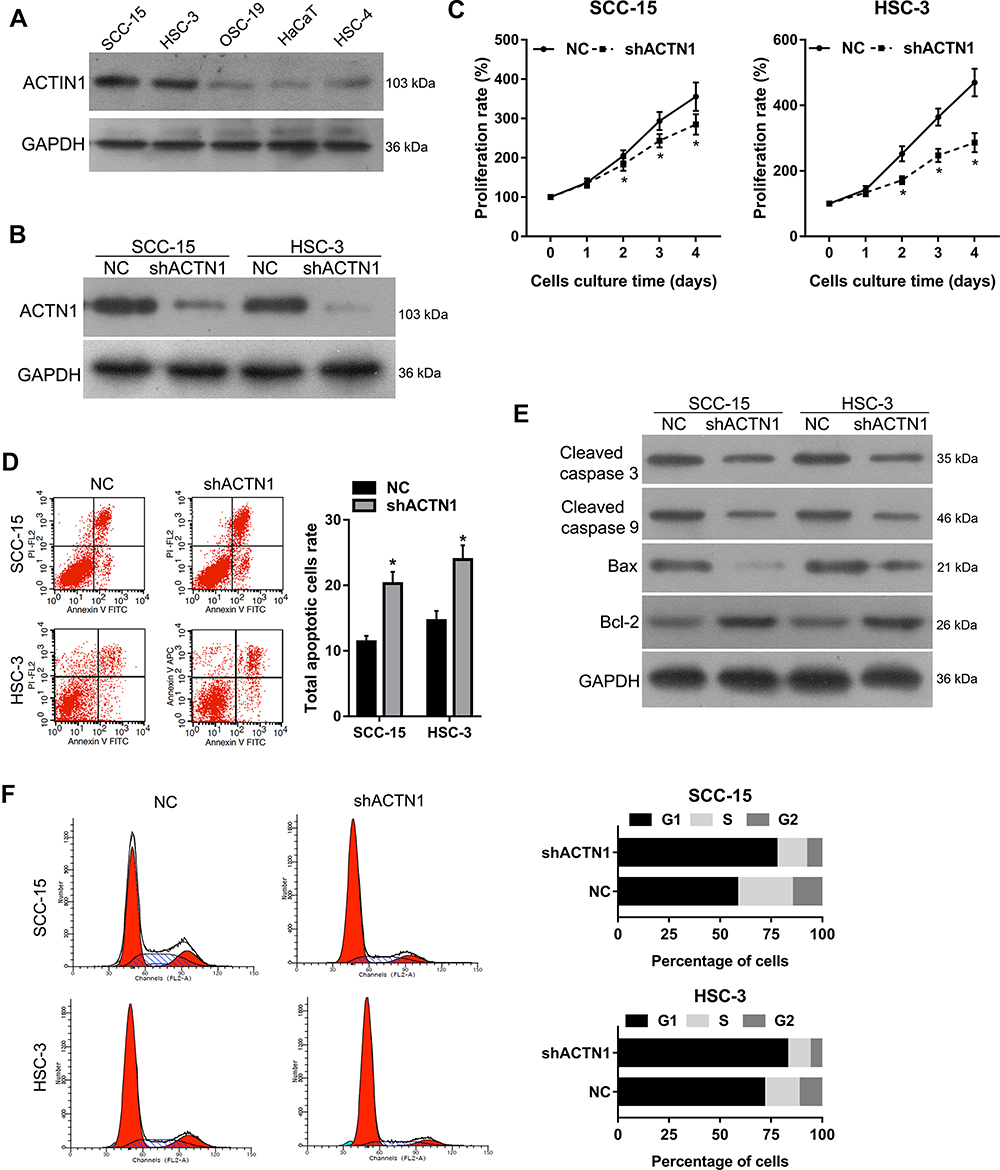 |
Figure 2 Effects of actinin alpha 1 (ACTN1) knockdown on cell proliferation, apoptosis, and the cell cycle. (A) ACTN1 expression in the oral squamous cell carcinoma (OSCC) cell lines, SCC-15, HSC-3, OSC-19, and HSC-4, and immortalized human keratinocytes, HaCaT. (B) ACTN1 in SCC-15 and HSC-3 cells was stably knocked down by transfecting the cells with lentivirus-expressing short hairpin ACTN1 and verified by Western blotting. The cells with stably knocked down ACTN1 were named shACTN1, and the negative control cells were named NC. The effects of ACTN1 knockdown on cell proliferation (C), apoptosis (D, E), and the cell cycle (F) were evaluated by Cell Counting Kit‑8 assays, flow cytometry analysis, and Western blotting to detect apoptosis related proteins, cleaved caspase 3, cleaved caspase 9, Bax, and Bcl-2. For panels D and E, the left side shows representative images, and the right side shows statistical results. *P <0.05. |
ACTN1 Knockdown Inhibited Cell Migration, Invasion, and EMT in vitro
To evaluate the effect of ACTN1 knockdown on cell migration, invasion, and EMT in vitro, transwell and wound-healing assays were performed. The results of the transwell assays showed that number of migrated or invasive cells that passed through the membrane onto the lower chamber, per field, was lesser in the shACTN1 group than in the NC group (Figure 3A and B). The results of the wound-healing assays showed that the percentage of wound closure in the shACTN1 group was lower than that in the NC group at 6 h, 24 h, and 48 h after wound generation (Figure 3C and D). In addition, epithelial marker E-cadherin levels were higher and mesenchymal marker, vimentin, levels were lower in the shACTN1 group cells than in the NC group cells (Figure 3E).
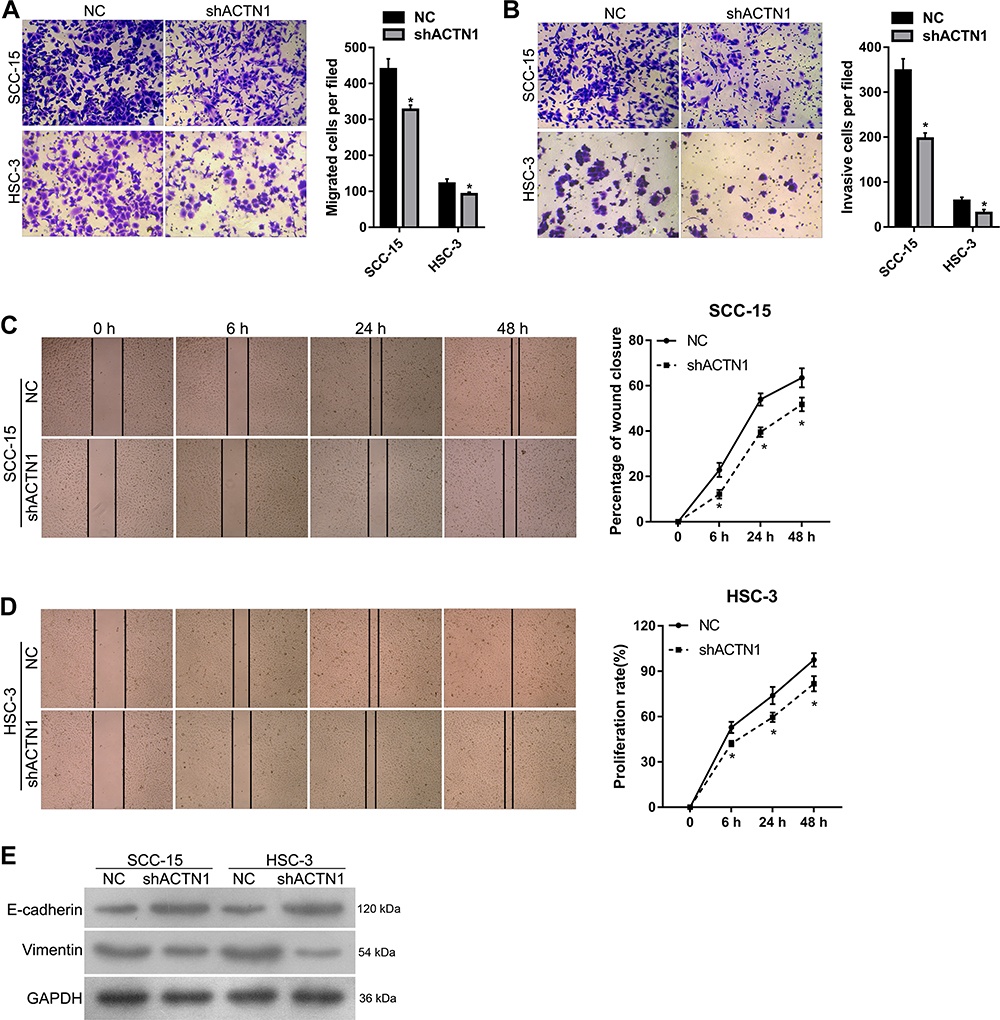 |
Figure 3 Effects of actinin alpha 1 (ACTN1) knockdown on cell migration, invasion, and epithelial-mesenchymal transition (EMT) in vitro. ACTN1 cells that were stably knocked down were named shACTN1, and negative control cells were named NC. The migration and invasion capabilities of the shACTN1 and NC group cells were evaluated by transwell (A, B) and wound-healing (C,D) assays. E-cadherin and vimentin protein levels in the shACTN1 and NC group cells were detected by Western blotting (E). For panel A and C, the left side shows representative images, and the right side shows statistical results. *P <0.05. |
ACTN1 Knockdown Inhibited Subcutaneous Tumor Growth and Pulmonary Metastasis in vivo
To verify our in vitro results, nude mouse models of subcutaneous xenograft and pulmonary metastasis were constructed to evaluate the effect of ACTN1 knockdown on subcutaneous tumor growth and pulmonary metastasis in vivo. As shown in Figure 4A and B, subcutaneous tumor volume generated by the shACTN1 group cells was lower than that generated by the NC group cells. Moreover, HE staining showed that the nude mouse lung tissues injected with shACTN1 group cells had lesser metastatic foci than those injected with NC group cells (Figure 4C). These results indicated that ACTN1 knockdown inhibited subcutaneous tumor growth and pulmonary metastasis in vivo.
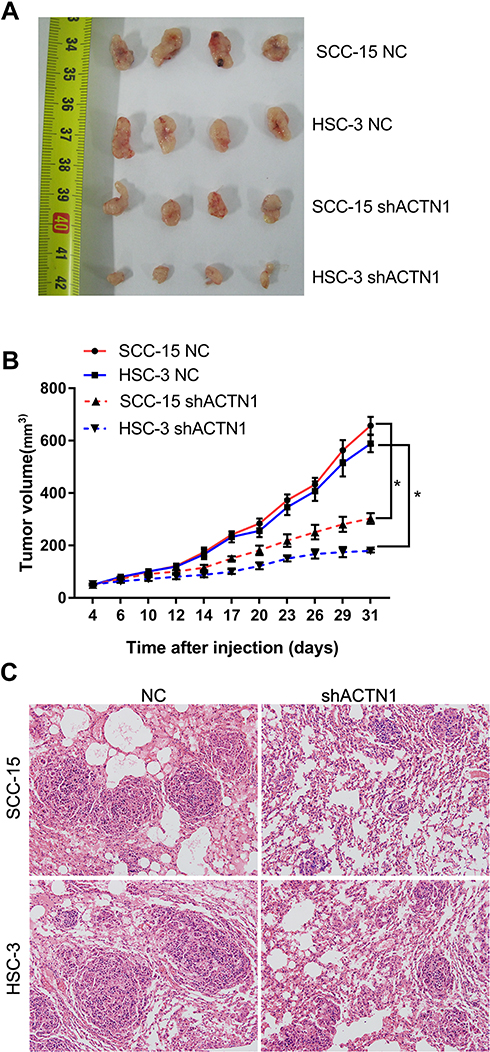 |
Figure 4 Effects of actinin alpha 1 (ACTN1) knockdown on subcutaneous tumor growth and pulmonary metastasis in vivo. Nude mouse models of subcutaneous xenograft and pulmonary metastasis were constructed using the shACTN1 and NC group SCC-15 and HSC-3 cells. (A–B) Photograph (A) and tumor volume curve (B) of subcutaneous tumors. *P <0.05. (C) Hematoxylin and eosin staining results of lung tissues obtained from nude mouse models of pulmonary metastasis. |
Discussion
ACTNs may serve as new potential diagnostic and prognostic markers of many types of cancers.7–10 Therefore, it is important to further investigate their role in other cancers. Based on GEPIA, a web-based tool to deliver fast and customizable functionalities based on TCGA and GTEx data, we found that among the four isoforms of ACTNs, only ACTN1 demonstrated an mRNA level that was significantly higher in HNSC tissues than in normal tissues. A high ACTN1 mRNA level indicated poor prognosis. Therefore, we aimed to investigate the role of ACTN1 in OSCC, the commonest head and neck cancer.
Firstly, we analyzed the ACTN1 protein levels in 10 normal mucosal tissues, 48 OSCC tissues, and 48 normal mucosal tissues adjacent to OSCC by IHC analysis. The statistical result of the total score of IHC staining showed that the ACTN1 protein level in the OSCC tissues was significantly higher than that in the normal tissues, indicating that the ACTN1 proteins were highly expressed in OSCC. This was consistent with the result that ACTN1 mRNA was highly expressed in HNSC tissues, which were analyzed by GEPIA. Following this, we found that ACTN1 protein levels were significantly associated with the clinical stage and node metastasis, and a high ACTN1 protein level indicated poor prognosis. Moreover, ACTN1 expression was an independent predictor of poor prognosis of OSCC. Therefore, our results suggested that ACTN1 may be a diagnostic and prognostic marker of OSCC. This conclusion is consistent with that of the study on ACTN1 in AML, suggesting ACTN1 as an independent poor prognostic factor.10 Our results could also be supported by Xu et al, who reported that ACTN1 level was higher in OSCC tissues than in normal tissues.12 In our future study, we will verify our hypothesis by enrolling more patients with OSCC and other types of HNSC.
Our results from clinical specimens suggested that ACTN1 might play a vital role in the tumorigenesis and development of OSCC. As ACTN1 level was highest in OSCC cell lines, SCC-15 and HSC-3, ACTN1 in SCC-15 and HSC-3 cells was stably knocked down by transfecting the cells with lv-shACTN1. Following this, we found that the proliferation rate was lower and the subcutaneous tumor volume was lesser in the shACTN1 group cells than in the NC group cells. These results suggested that ACTN1 knockdown inhibited cell proliferation in the OSCC cells. To further illuminate the mechanisms underlying the involvement of ACTN1 in cell proliferation, we analyzed the effect of ACTN1 knockdown on the cell cycle. We found that the number of cells in the G1 stage was higher and in the S and G2 stages was lower in the shACTN1 group than in the NC group. This indicated that ACTN1 knockdown induced G1 arrest. The cell cycle is the complex sequence of events by which eukaryotic cells grow and divide.13 Therefore, we hypothesized that ACTN1 was involved in regulating OSCC cell proliferation by affecting the cell cycle.
We also found that the total apoptotic cell rate of the shACTN1 group cells was higher than that of the NC group cells. These results indicated that ACTN1 knockdown could promote apoptosis in the OSCC cells. This conclusion was further supported by the expression profiles of cleaved caspase 3, cleaved caspase 9, Bax, and Bcl-2. Our results showed that ACTN1 knockdown could increase the levels of cleaved caspase 3, cleaved caspase 9, and Bax and decrease the Bcl-2 level in OSCC cells. Bax is a pro-apoptotic protein, while Bcl-2 is an anti-apoptotic protein.14–16 The increase in the levels of cleaved caspase 3 and caspase 9 showed that pro-apoptotic proteins were activated.14–16 Therefore, all of our results suggested that ACTN1 was involved in apoptosis of the OSCC cells, and a decrease in ACTN1 might promote the apoptosis of OSCC cells.
Finally, we evaluated the role of ACTN1 in regulating metastasis of OSCC. The results of the transwell assay showed that ACTN1 knockdown could decrease the number of migrated and invasive cells that passed through the membrane onto the lower chamber. The results of the wound-healing assay showed that ACTN1 knockdown could decrease the percentage of wound closure. All these in vitro results suggested that ACTN1 knockdown inhibited cell migration and invasion in OSCC, and this suggestion was supported by the in vivo results that showed that ACTN1 knockdown inhibited pulmonary metastasis of OSCC in nude mice. To further investigate the mechanism of ACTN1 in regulating metastasis, we examined the effect of ACTN1 knockdown on the EMT-related proteins. Our results showed that ACTN1 knockdown increased the level of the epithelial marker,17 E-cadherin, but decreased the level of the mesenchymal marker,17 vimentin, indicating that ACTN1 knockdown inhibited EMT. The contribution of EMT to tumor invasion, migration, and metastatic outgrowth has been reported by many researchers.18–20 Therefore, we hypothesized that ACTN1 knockdown might suppress OSCC metastasis by inhibiting EMT.
However, our present study had one limitation. The underlying mechanism of regulation of cell proliferation and metastasis of OSCC by ACTN1 remains unclear, and hence, in our future studies, we will focus on the regulatory mechanisms by measuring the mRNA, non-coding RNA, and protein levels.
Conclusion
In conclusion, ACTN1 level was higher in the OSCC tissues than in the normal tissues. ACTN1 levels were significantly associated with the clinical stage and node metastasis, and a high ACTN1 protein level indicated poor prognosis. Moreover, ACTN1 knockdown could suppress cell proliferation and metastasis of OSCC in in vitro and in vivo models. Therefore, our results suggested that ACTN1 may be a diagnostic and prognostic marker of OSCC. Our study provides a potential target for novel therapeutic strategies involving ACTN1 inhibition and control of OSCC proliferation and metastasis.
Acknowledgments
This study was supported by the Natural Science Foundation of Guangdong Province (2018A030310164), the Fundamental Research Funds for the Central Universities (18zxxt58), and Guangzhou key medical discipline construction project fund.
Disclosure
We wish to confirm that there are no known conflicts of interest associated with this publication and no significant financial support that could have influenced the work and its outcome was extended.
References
1. Rivera C. Essentials of oral cancer. Int J Clin Exp Pathol. 2015;8(9):11884–11894.
2. Choi S, Myers JN. Molecular pathogenesis of oral squamous cell carcinoma: implications for therapy. J Dent Res. 2008;87(1):14–32. doi:10.1177/154405910808700104
3. Zini A, Czerninski R, Sgan-Cohen HD. Oral cancer over four decades: epidemiology, trends, histology, and survival by anatomical sites. J Oral Pathol Med. 2010;39(4):299–305. doi:10.1111/j.1600-0714.2009.00845.x
4. de Aguiar A, Kowalski LP, de Almeida OP. Clinicopathological and immunohistochemical evaluation of oral squamous cell carcinoma in patients with early local recurrence. Oral Oncol. 2007;43(6):593–601. doi:10.1016/j.oraloncology.2006.07.003
5. Foley KS, Young PW. The non-muscle functions of actinins: an update. Biochem J. 2014;459(1):1–13. doi:10.1042/BJ20131511
6. Shao H, Wang JH, Pollak MR, Wells A. α-actinin-4 is essential for maintaining the spreading, motility and contractility of fibroblasts. PLoS One. 2010;5(11):e13921. doi:10.1371/journal.pone.0013921
7. Tentler D, Lomert E, Novitskaya K, Barlev NA. Role of ACTN4 in tumorigenesis, metastasis, and EMT. Cells. 2019;8(11):E1427. doi:10.3390/cells8111427
8. Kakuya T, Mori T, Yoshimoto S, et al. Prognostic significance of gene amplification of ACTN4 in stage I and II oral tongue cancer. Int J Oral Maxillofac Surg. 2017;46(8):968–976. doi:10.1016/j.ijom.2017.03.001
9. Liu X, Chu K-M. α-Actinin-4 promotes metastasis in gastric cancer. Lab Invest. 2017;97(9):1084–1094. doi:10.1038/labinvest.2017.28
10. Yang X, Pang Y, Zhang J, et al. High expression levels of ACTN1 and ACTN3 indicate unfavorable prognosis in acute myeloid leukemia. J Cancer. 2019;10(18):4286–4292. doi:10.7150/jca.31766
11. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
12. Xu J, Lv H, Zhang B, et al. miR-30b-5p acts as a tumor suppressor microRNA in esophageal squamous cell carcinoma. J Thorac Dis. 2019;11(7):3015–3029. doi:10.21037/jtd.2019.07.50
13. McGowan CH. Regulation of the eukaryotic cell cycle. Prog Cell Cycle Res. 2003;5:1–4.
14. Pistritto G, Trisciuoglio D, Ceci C, Garufi A, D’Orazi G. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging. 2016;8(4):603–619. doi:10.18632/aging.100934
15. Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495–516. doi:10.1080/01926230701320337
16. Goldar S, Khaniani MS, Derakhshan SM, Baradaran B. Molecular mechanisms of apoptosis and roles in cancer development and treatment. Asian Pac J Cancer Prev. 2015;16(6):2129–2144. doi:10.7314/APJCP.2015.16.6.2129
17. Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196.
18. Yeung KT, Yang J. Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol. 2017;11(1):28–39. doi:10.1002/1878-0261.12017
19. Diepenbruck M, Christofori G. Epithelial-mesenchymal transition (EMT) and metastasis: yes, no, maybe? Curr Opin Cell Biol. 2016;43:7–13. doi:10.1016/j.ceb.2016.06.002
20. Liao TT, Yang MH. Revisiting epithelial-mesenchymal transition in cancer metastasis: the connection between epithelial plasticity and stemness. Mol Oncol. 2017;11(7):792–804. doi:10.1002/1878-0261.12096
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.