")
Back to Journals » Infection and Drug Resistance » Volume 14
Heterogeneity of Helicobacter pylori Strains Isolated from Patients with Gastric Disorders in Guiyang, China
Authors Mi M, Wu F, Zhu J , Liu F, Cui G, Wen X, Hu Y, Deng Z, Wu X , Zhang Z, Qi T, Chen Z
Received 19 October 2020
Accepted for publication 9 January 2021
Published 11 February 2021 Volume 2021:14 Pages 535—545
DOI https://doi.org/10.2147/IDR.S287631
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Mengheng Mi,1,2,* Fangcao Wu1 12 2,* Jian Zhu3 3, Fang Liu1 12 2, Guzhen Cui1 14 4, Xueqing Wen1 12 2, Yue Hu5 5, Zhaohui Deng3 3, Xiaojuan Wu1 12 2, Zhengrong Zhang1 14 4, Tingna Qi1 14 4, Zhenghong Chen1 12 2
1Key Laboratory of Microbiology and Parasitology of Education Department of Guizhou, School of Basic Medical Science, Guizhou Medical University/Joint Laboratory of Helicobacter Pylori and Intestinal Microecology of Affiliated Hospital of Guizhou Medical University, Guiyang, People’s Republic of China; 2Key Laboratory of Environmental Pollution Monitoring and Disease Control, Ministry of Education, Guizhou Medical University, Guiyang, People’s Republic of China; 3Department of Gastroenterology, Guiyang Hospital of Guizhou Aviation Industry Group, Guiyang, People’s Republic of China; 4Key Laboratory of Endemic and Ethnic Diseases (Guizhou Medical University), Ministry of Education, Guiyang, People’s Republic of China; 5Department of Gastroenterology, Affiliated Hospital of Guizhou Medical University, Guiyang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhenghong Chen
Department of Microbiology, School of Basic Medical Science, Guizhou Medical University, Guiyang, People’s Republic of China
Tel + 86-13985006815
Fax + 86-85188174015
Email [email protected]
Purpose: Chronic Helicobacter pylori infection causes peptic ulcers in a subpopulation of individuals and is a risk factor for the development of gastric cancer. Multiple infections and heteroresistant H. pylori contribute to poor treatment efficacy. Here, we investigated the extent of genetic diversity among H. pylori strains within a given host and its influence on the results of antibiotic (metronidazole, levofloxacin, clarithromycin, amoxicillin, and tetracycline) susceptibility testing.
Materials and Methods: Gastric mucosa biopsy samples were obtained from patients with gastric disorders, including 48 H. pylori positive patients, who were never previously treated for H. pylori infection. Five potential H. pylori colonies isolated from each sample were subcultured for enrichment. Enriched H. pylori colonies were identified through Gram staining and assays for urease, oxidase, and catalase. For each H. pylori monoclonal colony, the antibiotic susceptibility was assessed, genomic DNA was sequenced, and the cytotoxin-associated gene A (cagA) genotype was verified. Co-infection with multiple H. pylori strains was determined using random amplified polymorphic DNA (RAPD)-polymerase chain reaction (PCR).
Results: Thirteen gastric mucosa biopsy samples were positive for H. pylori. Five monoclonal strains isolated from each of these 13 patients were identified as H. pylori. RAPD-PCR indicated that intra-patient monoclonal strains of H. pylori in 10 of the 13 samples exhibited heterogeneity. Among the 13 patients, intra-patient monoclonal strains isolated from 4 patients had identical cagA genotype, whereas intra-patient monoclonal strains isolated from the other 9 patients harbored more than one cagA genotype. The antibiotic susceptibility of five intra-patient monoclonal strains from seven patients was inconsistent.
Conclusion: The existence of heterogeneous H. pylori strains with resistance to different drugs and virulence were common within the gastric mucosa of an individual patient.
Keywords: Helicobacter pylori, heterogeneity, antibiotic resistance, cagA typing
Introduction
Helicobacter pylori is a spiral, microaerobic, Gram-negative bacterium and among the most common chronic pathogens associated with human diseases.1 Helicobacter pylori infection can last for decades and is the main cause of chronic active gastritis, peptic ulcer, and gastric mucosa-associated lymphoma. Moreover, the WHO has classified H. pylori as a class I carcinogen for gastric cancer.2,3 The average H. pylori infection rate in the global population is >50% but varies greatly among countries,4 with infection rates in developing countries higher than those in developed countries. In 2017, Hooi et al5 reported that Nigeria had the highest H. pylori infection rate (87.7%), whereas Switzerland had the lowest infection rate (18.9%). In 2014, Wang et al6 reported that the average infection rate of H. pylori in China was 55.8%, with the highest rate in the Tibet region (84.6%).
Triple therapy, which involves the use of one proton-pump inhibitor (PPI) and two antibiotics (usually amoxicillin and metronidazole/clarithromycin), was introduced 30 years ago for the eradication of H. pylori and showed great success at that time. However, due to the widespread use of antibiotics globally, the antibiotic resistance of H. pylori strains is significantly higher than it was in previous decades and multi-resistant strains have become more common.7–11 The success rate of triple therapy has declined significantly in recent years.12
The consensus of the Kyoto global report on H. pylori gastritis published in 201513 suggests that H. pylori exhibits a large geographical difference in resistance to common antibiotics; therefore, the preferred eradication programs in different regions vary accordingly. In 2016, the Toronto Consensus Group proposed a strategy involving a 14-day administration of bismuth quadruple therapy (PPI + bismuth + two antibiotics) for the treatment of H. pylori infection in the updated treatment guidelines.14 Many European countries have abandoned the use of triple therapy and replaced it with quadruple therapy.
According to the European Maastricht Consensus,15 failure to eradicate H. pylori should be followed by rescue treatment guided by drug-susceptibility testing of the H. pylori strain. Therefore, the rational use of antibiotics based on drug-susceptibility tests has received increasing attention. Farzi et al16 found that strains exhibiting different drug resistance could be isolated from the same patient, and other reports indicated that H. pylori isolated from the gastric antrum and corpus of the same patient showed different drug susceptibilities.17,18
The presence of multiple H. pylori strains colonizing a patient with gastric cancer was reported previously.19 Besides exhibiting multi-drug resistance; the bacteria had different genotypes for cytotoxin-associated gene A (cagA) and vacuolating cytotoxin gene A (vacA), the main virulence factors, in those intra-patient isolates. Genetic variability may result in the evolution of more virulent and adapted strains. CagA is an oncogenic protein, which comprises a conserved N-terminal region, a variable EPIYA (Glu-Pro-Ile-Tyr-Ala)-repeat region, and a C-terminal tail. Based on the flanking amino acid sequences, EPIYA can be further classified based on the four motifs: EPIYA-A, -B, -C, and -D. According to the composition of the EPIYA-A, -B, -C, and D motifs, H. pylori can be classified into different cagA subtypes, such as cagA-AB, -ABC, -ABD or-BD.20,21 The presence of complex and mixed EPIYA pattern may increase the risk of gastric cancer.
Our previous studies demonstrated the occurrence of mixed H. pylori strains within the same patient,22 suggesting co-infection with multiple H. pylori strains, perhaps with different antibiotic susceptibilities. When choosing antibiotics for rescue therapy, antibiotics should be selected based on the results of drug-susceptibility testing; however, the only way to obtain H. pylori isolates is via endoscopy examination. Therefore, susceptibility testing is not performed for majority of patients. Furthermore, co-infection with different H. pylori strains with different phenotypes and genotypes could potentially contribute to difficulty in eradication and further complications. This study explored the heteroresistance, genotypic diversity, and CagA typing of H. pylori strains isolated from a single host and the prevalence of mixed infection in individual patients in Guiyang, China.
Materials and Methods
Gastric Samples
A total of 48 patients were recruited from Guiyang Hospital of Guizhou Aviation Industry Group between January and December 2018. The exclusion criteria included: 1) anti-H. pylori treatment in the previous 15 days; 2) complications such as massive gastrointestinal bleeding and/or pyloric obstruction; or 3) severe heart, lung, or kidney disease. Gastric mucosa biopsy samples (two each) were obtained through endoscopy from patients with gastric disorders. One of the samples was sent to the pathology department for pathological examination, while the other was used for the isolation of H. pylori. All protocols, including the protocol for obtaining informed consent, were approved by the Human Medical Ethics Committee of Guizhou Medical University, and all procedures contributing to the study complied with the Declaration of Helsinki. All patients were informed as to the purpose of this study, unless incapacitated; informed consent was obtained from the patients or from the guardians of the incapacitated patients.
H. pylori Isolation and Identification
Gastric antrum biopsy samples were homogenized and inoculated in selective BHI agar base (Oxoid, Ltd., Basingstoke, UK) containing 10% sheep blood (Qingdao Hope Biol-Technology Co., Ltd., Qingdao, China) and Skirrow’s supplement (vancomycin 10 mg/L, trimethoprim 5 mg/L, cefsulodin 5 mg/L, and amphotericin B; Oxoid, Ltd.). Agar plates were incubated at 37°C for 3 to 5 days under microaerophilic conditions (10% CO2, 85% N2, and 5% O2) with 95% relative humidity (Micro Aerobic Bag; Mitsubishi Chemical Corporation, Tokyo, Japan). The potential H. pylori colonies were subcultured on fresh blood agar plates and incubated for two to four days to obtain monoclonal cultures which were then identified through assays for urease,23 catalase,24 and oxidase.25 Subcultures with positive results for all three enzymatic tests were transferred to a single blood agar plate and incubated for another three to five days. They were then harvested for drug-susceptibility testing and genomic DNA extraction, followed by species confirmation using H. pylori‑specific 16S rRNA gene fragment polymerase chain reaction (PCR).
Genomic DNA was extracted using an Ezup column bacteria genomic DNA purification kit (Sangon Biotech Co., Ltd.), according to the manufacturer’s instructions. The DNA purity and concentration were measured using a NanoDrop 2000 ultra-micro UV spectrophotometer (Thermo Fisher Scientific., Waltham, MA, USA). Universal primer sequences26 and amplification conditions for 16S rRNA gene PCR are shown in Table 1. Primers were purchased from Sangon Biotech Co., Ltd. (Shanghai, China). 16S rRNA gene PCR products were sent to Sangon Biotech Co., Ltd. for sequencing. Species were identified using NCBI blast.
 |
Table 1 Primer Sequences and PCR Conditions |
H. pylori Drug-Susceptibility Testing
Breakpoint susceptibility testing was performed to examine the antibiotic-susceptibility of H. pylori, as per the method proposed by the Clinical Laboratory Standard Institute.27 Five antibiotics (metronidazole, levofloxacin, clarithromycin, amoxicillin, and tetracycline) were used for the antibiotic-susceptibility testing, and the breakpoint concentrations27 are listed in Table 2. For H. pylori breakpoint susceptibility testing, a simplified version of the agar dilution method was used. Inocula were prepared from 3-day-old agar plates and colonies were harvested in 2 mL sterile saline. The fresh bacterial suspension was adjusted to an opacity equivalent to a 2 Mcfarland turbidity standard (approximately 6 × 108 CFU/mL) and inoculated directly onto an agar plate containing an antibiotic concentration equal to the breakpoint concentration, which defines resistance, and also inoculated on an agar plate without antibiotics as a growth control.
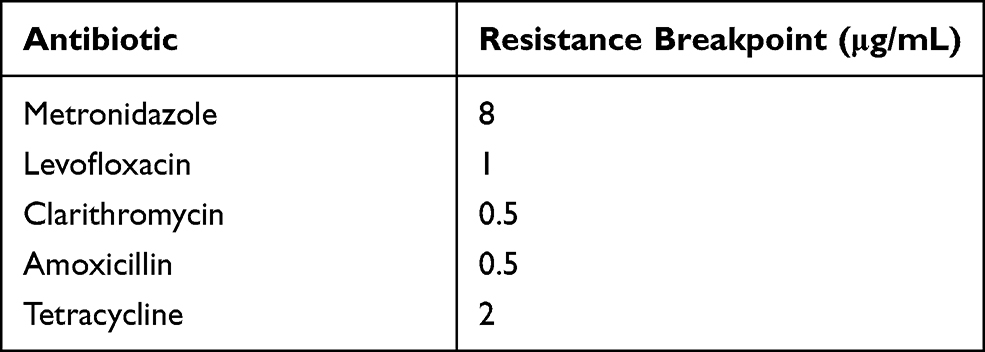 |
Table 2 Antibiotic Breakpoints |
All strains are expected to grow on the agar plates without antibiotics. H. pylori ATCC26,695 (provided by the Institute of Pathogenic Biology of Qilu Medical College, Shandong University), which is susceptible to all five antibiotics in this study, was used as the control strain. All plates were incubated at 37°C for 72 h under microaerophilic conditions. Results of the antibiotic-susceptibility test for each single colony (monoclonal strain) and the pool of five colonies from the same patient were recorded.
PCR Amplification and Sequencing of the Variable Region of cagA in Each Monoclonal Strain
Genomic DNA was prepared as described above. PCR primers22,28 (Sangon Biotech Co., Ltd.) used for amplification of the cagA gene are shown in Table 2. PCR assays were performed in a volume of 50 µL; the reaction mixtures contained 2 μL of forward primer (10 μM), 2 μL of reverse primer (10 μM), 1 μL of genomic DNA (~10 ng/μL), 25 µL of 2× Taq PCR master mix (Jiangsu CWBiotech Science and Technology Co., Ltd., Jiangsu, China), and 20 µL of Milli-Q water. All PCR amplification runs included one negative control (Milli-Q water instead of genomic DNA) and one positive control (H. pylori 26,695). After PCR, 6 μL of the amplification products and DNA ladder markers (Tiangen Biotech Co., Ltd., Beijing, China) were electrophoretically separated on 1% agarose gels in Tris-acetate EDTA buffer, followed by staining with GoldView 1 nucleic acid stain (Solarbio Science and Technology Co., Ltd., Beijing, China) and visualization using a BG-gdaAUTO gel imaging analysis system (Beijing Baygenebiotech Biotech Co., Ltd., Beijing, China). PCR products were sequenced at Sangon Biotech Co., Ltd., and the gene sequences were translated into amino acid sequences using BioEdit software (v.7.1.3.0; https://www.bioedit.com/). In the intra-patient strains, CagA genotypes (the EPIYA motif patterns) were classified based on amino acid sequences flanking the EPIYA motif.20
Random Amplified Polymorphic DNA (RAPD)-PCR Analysis
RAPD-PCR typing was performed to detect the related and distinct strains of H. pylori that colonized the stomach of each patient. Accordingly, the random primers 124728,29 and 128328 were used (Table 1). The amplification was performed in a 100 µL volume containing 50 µL of 2x Taq PCR master mix (Jiangsu CWBiotech Science and Technology Co., Ltd.), 1 µL of genomic DNA (~20 ng), 2 µL of primer 1247 or primer 1283, 10 mM Tris-HCl, 50 mM KCl, and Milli-Q water. Amplification conditions were as follows: 94°C for 2 min, 37°C for 1 min, and 72°C for 4 min, followed by 29 cycles of 94°C for 2 min, 37°C for 3 min, and 72°C for 7 min. PCR products were then electrophoresed in 2% agarose gels. The similarity between the RAPD banding profiles of five distinct strains from the same patient was analyzed, and the identical or indistinguishable RAPD band pattern was considered to represent infection by a single strain, whereas two or more RAPD band patterns indicated infections by different strains.29–31
Results
H. pylori Isolation
Helicobacter pylori were isolated from 13 out of 48 patients (27%). From the samples obtained from each of the 13 H. pylori positive patients, five single colonies were subcultured that resulted in the isolation of 65 monoclonal strains in total. We have used serial number G1-G13 to represent the 13 H. pylori positive samples. All of these strains were positive for oxidase, urease, and catalase tests and the 16S rDNA sequences showed ~99% to 100% identity to H. pylori 26695. The antibiotic-susceptibility results for the 65 monoclonal strains are listed in Table 3. Five intra-patient monoclonal strains from G5, G6, G7, G9, and G12 exhibited identical antibiotic-susceptibility results. However, intra-patient monoclonal strains isolated from the other seven patients showed varying antibiotic-susceptibility profiles.
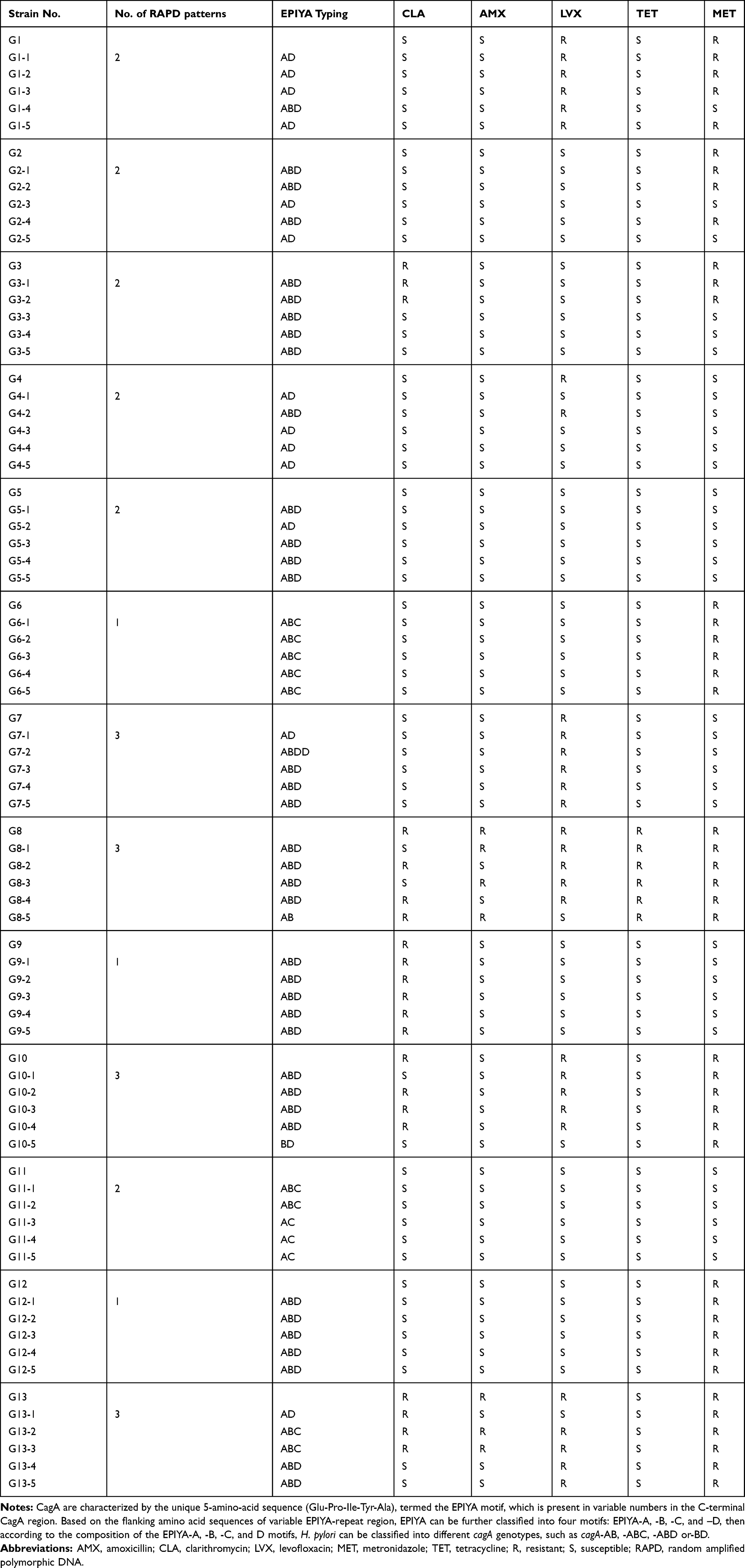 |
Table 3 Antibiotic Susceptibility and Typing of 65 Monoclonal Strains Isolated from 13 Patients |
Results of cagA Typing of 65 Monoclonal H. pylori Strains from 13 Patients
The results of cagA typing of 65 monoclonal H. pylori colonies isolated from 13 patients are listed in Table 3. CagA was present in all 65 colonies and five intra-patient monoclonal strains isolated from four patients (G3, G6, G9, and G12) displayed the same cagA genotype. However, intra-patient monoclonal strains isolated from two other patients (G7 and G13) displayed three cagA genotypes, and those from seven other patients (G1, G2, G4, G5, G8, G10, and G11) displayed two cagA genotypes.
RAPD-PCR Typing
RAPD-PCR fingerprinting was performed on DNA extracts from the 65 monoclonal strains obtained from 13 patients, and the patterns of the five intra-patient strains isolated from an individual patient shown in Figure 1. Monoclonal strains from 3 patients (G6, G9, and G12) showed identical intra-patient molecular patterns, whereas the other 10 patients harbored two or three distinct strains. Typing results are shown in Table 3.
 |
Figure 1 Electropherogram of RAPD-PCR typing of Helicobacter pylori strains from 13 patients.Notes: M: 2K DNA ladder; G1–G13: sample/patient number; bands 1–5: monoclonal strains from a single patient amplified using primer 1247; and bands 6–10: monoclonal strains from a single patient amplified using primer 1283. |
Discussion
H. pylori infection is the most common bacterial infection of the digestive system.1,3,32
In local hospitals, some patients’ symptoms were relieved or even disappeared after eradication treatment with amoxicillin and clarithromycin combined with esomeprazole, but symptoms recurred after a few months. The infection recurrence may be due to spontaneous mutation for drug resistance. Under the effects of antibiotics, susceptible wild strains could have been eradicated while drug-resistant clones were screened out, with the growth of resistant clones leading to the recurrence of symptoms. Co-infection with susceptible and resistant strains in the stomachs of patients might be another reason for the recurrence of symptoms. During treatment with antibiotics, the susceptible strains are removed. With the reduction in the number of bacteria, the symptoms are temporarily relieved, but with the proliferation of drug-resistant strains, obvious symptoms later reappear. In any case, mixed infections caused by different clones are difficult to detect and can likely impact the effectiveness of eradication treatment if the resistant strain is not detected.11,29,33,34
H. pylori is among the most genetically diverse bacterial species studied to date. Although most genomes range from 1.5 Mb to 1.8 Mb, diversity among strains includes variations in the complement of genes, chromosomal gene order, deployment of repetitive DNA, sequence variation in conserved genes, homoplasies, status of phase-variable genes, complement of restriction-modification loci, and mobile DNA.35–37 These genetic traits might promote the coexistence of several strains with different antibiotic susceptibilities or different genotypes colonizing a single host, which is defined as H. pylori heterogeneity, and has been reported in several studies.29,31,38,39 In this study, RAPD-PCR and cagA typing was used for distinguishing the heterogeneity of intra-patient strains of H. pylori.
Among several genotyping methods applied to H. pylori, RAPD-PCR is considered to be ideal, because it is a simple, rapid, and low-cost means of distinguishing H. pylori isolates.40 Although multilocus sequence typing and whole-genome sequencing are well suited for phylogenic studies, they are time-consuming and very expensive for emerging countries. In any case, RAPD typing has conclusively indicated high discriminatory power and made significant contributions to several studies.18,31,33,34,41–47 Bacterial infection resulting in more than one RAPD-PCR banding pattern within an individual patient is considered to indicate mixed infections.28,31 Farzi et al39 reported 40 intra-patient H. pylori strains isolated from the antrum and corpus of 97 patients, with isolate diversity in resistance to clarithromycin detected in 17.5% (7/40) of the patients. Additionally, they found that the heteroresistant strains were mostly derived from pre-existing strains in each patient rather than from mixed-type infection. Similarly, Kao et al34 reported 19 intra-patient strains exhibiting antibiotic heteroresistance, whereas only an isolate from one patient (5.3%) showed a different RAPD-PCR banding pattern relative to the identical or similar fingerprinting patterns displayed by the others. These heteroresistant strains are considered to exhibit distinct microevolution derived from pre-existing strains during chronic infection. Furthermore, other studies reported colonization of individuals with more than a single H. pylori strain. Qureshi et al48 isolated 10 single colonies from 30 patients, respectively, with gastro-duodenal problems, with characterization of each colony using DNA fingerprinting, as well as genotyping of several important genetic markers and two other genetic segments, revealing that all 30 patients were infected with more than one strain and sometimes strains with five to six genetic variants. Interestingly, Mansour et al29 reported that multiple H. pylori infection is more prevalent in Tunisia than in France and demonstrated a higher prevalence of multiple infections in a developing country when compared to a developed country. However, this could also be explained based on the differing prevalence of infection between the two countries.
CagA of H. pylori is an oncogenic protein and is a major virulence factor associated with gastric cancer. The prevalence of cagA among H. pylori in different regions varies between 50% and 60% in certain western countries to almost 100% in East Asia.49 A lot of studies have shown that the cagA genotype varies markedly worldwide. These genotyping were carried out using strains from different patients, and not from intra-patient strains.
In this study, 13 intra-patient H. pylori strains were isolated from 13 patients, and according to cagA typing and RAPD-PCR fingerprinting, intra-patient strains isolated from 10 patients showed heterogeneity and were considered to indicate multiple infections. Among these patients, G13 had gastric cancer and harbored three strains with three different CagA EPIYA motifs and different RAPD-PCR typing. Moreover, susceptibility testing showed different antibiotic resistance spectra for these three strains. The heterogeneity of H. pylori has important clinical significance and suggests that the antibiotic-susceptibility of a single strain isolated from a biopsy might not be absolutely representative. We identified amoxicillin-resistant H. pylori strains in two out of the 13 patients, including the patient who had gastric cancer. Amoxicillin-resistance H. pylori is relatively rare, although it has already been reported.48,50,51 Based on our results, we propose that the lack of knowledge about heterogeneous strains in an individual may result in underestimated H. pylori drug resistance, which further complicates its eradication.
This research has some limitations. The number of samples is small, with H. pylori isolated only from 13 patients. In this study, it usually took 3–5 h from sample collection to delivery to the microbiology research laboratory for inoculation. Though selective medium containing antibiotic additives (nalidixic acid, polymyxin B, and bacitracin) was used for H. pylori isolation, non-H. pylori still grew and affected the growth of H. pylori in some cases, and the positive rate of H. pylori isolation was only approximately 30%, which is similar to that reported previously.52–54 To improve the positive rate of isolation, inoculation was performed immediately after sampling or transporting samples at low temperatures, using dry ice to keep them cold, and adding nalidixic and bacitracin along with H. pylori-selective supplements in the medium to avoid the growth of non-H. pylori. This is a small sample-size study, but we did find a general heterogeneity of H. pylori in the stomachs of local patients. Thus, we consider this as a pilot study, and a larger study will be conducted in future.
Conclusion
Multiple strain infection with H. pylori is common. Therefore, when selecting H. pylori colonies for antibiotic-susceptibility testing, it is important to select more than one colony.
Ethics Approval and Informed Consent
All protocols were approved by the Human Medical Ethics Committee of Guizhou Medical University (Ethics review approval number: 2017-52), and all procedures, including the protocol for obtaining informed consent, were carried out in accordance with the principles of the Declaration of Helsinki. Informed consent was obtained from all patients or from the guardians of incapacitated patients prior to the initiation of the study.
Acknowledgments
We would like to thank Prof. Barry J. Marshall for his guidance on isolation and culture, and Dr. Chin Yen Tay for their suggestions on the revision of the paper. We would also like to thank Editage [www.editage.cn] for their help with English-language editing.
Co-first authors: Mengheng Mi and Fangcao Wu.
Funding
This work was funded by the National Natural Science Foundation of China (Nos. 81860353 and 81460314) and Natural Science of Foundation of Guizhou ([2019]1441) and the United Foundation of Guiyang Medical University-Science and Technology Department of Guizhou Province (Nos. [2012]010 and [2018]5779–17).
Disclosure
The authors declare no conflicts of interest for this work.
References
1. NIH Consensus Conference. Helicobacter pylori in peptic ulcer disease. NIH consensus development panel on helicobacter pylori in peptic ulcer disease. JAMA. 1994;272(1):65–69. doi:10.1001/jama.272.1.65
2. Malfertheiner P, Megraud F, O’Morain CA, et al. Management of Helicobacter pylori infection–the Maastricht IV/Florence consensus report. Gut. 2012;61:646–664. doi:10.1136/gutjnl-2012-302084
3. Correa P. Helicobacter pylori as a pathogen and carcinogen. J Physiol Pharmacol. 1997;48(5):19–24.
4. Khalifa MM, Sharaf RR, Aziz RK. Helicobacter pylori: a poor man’s gut pathogen? Gut Pathog. 2010;2(1):2. doi:10.1186/1757-4749-2-2
5. Hooi JKY, Lai WY, Ng WK, et al. Global prevalence of Helicobacter pylori infection: systematic review and meta-analysis. Gastroenterology. 2017;153(2):420–429. doi:10.1053/j.gastro.2017.04.022
6. Wang R, Chen X-Z. High mortality from hepatic, gastric and esophageal cancers in mainland China: 40 years of experience and development. Clin Res Hepatol Gastroenterol. 2015;21(3):751–756. doi:10.1016/j.clinre.2014.04.014
7. Liu DS, Wang YH, Zeng ZR, et al. Primary antibiotic resistance of Helicobacter pylori in Chinese patients: a multiregion prospective 7-year study. Clin Microbiol Infect. 2018;24(7):
8. Boyanova L, Hadzhiyski P, Kandilarov N, Markovska R, Mitov I. Multidrug resistance in Helicobacter pylori: current state and future directions. Expert Rev Clin Pharmacol. 2019;12(9):909–915. doi:10.1080/17512433.2019.1654858
9. Wang D, Guo Q, Yuan Y, Gong Y. The antibiotic resistance of Helicobacter pylori to five antibiotics and influencing factors in an area of China with a high risk of gastric cancer. BMC Microbiol. 2019;19(1):152. doi:10.1186/s12866-019-1517-4
10. Zhang YX, Zhou LY, Song ZQ, Zhang JZ, He LH, Ding Y. Primary antibiotic resistance of Helicobacter pylori strains isolated from patients with dyspeptic symptoms in Beijing: a prospective serial study. World J Gastroenterol. 2015;21(9):2786–2792. doi:10.3748/wjg.v21.i9.2786
11. Savoldi A, Carrara E, Graham DY, Conti M, Tacconelli E. Prevalence of antibiotic resistance in helicobacter pylori: a systematic review and meta-analysis in World Health Organization regions. Gastroenterology. 2018;155(5):1372–1382.e1317. doi:10.1053/j.gastro.2018.07.007
12. Iwanczak F, Iwanczak B. Treatment of Helicobacter pylori infection in the aspect of increasing antibiotic resistance. Adv Clin Exp Med. 2012;21(5):671–680.
13. Sugano K, Tack J, Kuipers EJ, et al. Kyoto global consensus report on Helicobacter pylori gastritis. Gut. 2015;64(9):1353–1367. doi:10.1136/gutjnl-2015-309252
14. O’Connor A, Lamarque D, Gisbert JP, O’Morain C. Treatment of Helicobacter pylori infection 2017. Helicobacter. 2017;22(Suppl1). doi:10.1111/hel.12410
15. Malfertheiner P, Megraud F, O’Morain CA, et al. Management of Helicobacter pylori infection - the maastricht v/florence consensus report. Gut. 2017;66(1):6–30.
16. Farzi N, Malekian T, Alebouyeh M, Vaziri F, Zali MR. Genotype diversity and quasispecies development of Helicobacter pylori in a single host. Jpn J Infect Dis. 2015;68(4):176–180. doi:10.7883/yoken.JJID.2014.165
17. Ayala G, Galvan-Portillo M, Chihu L, et al. Resistance to antibiotics and characterization of Helicobacter pylori strains isolated from antrum and body from adults in Mexico. Microb Drug Resist. 2011;17(2):149–155. doi:10.1089/mdr.2010.0154
18. Selgrad M, Tammer I, Langner C, et al. Different antibiotic susceptibility between antrum and corpus of the stomach, a possible reason for treatment failure of Helicobacter pylori infection. World J Gastroenterol. 2014;20(43):16245–16251. doi:10.3748/wjg.v20.i43.16245
19. Grande R, Di Giulio M, Di Campli E, et al. A model of Helicobacter pylori persistence in a case of gastric cancer. New Microbiol. 2010;33(4):343–349.
20. Hatakeyama M. Anthropological and clinical implications for the structural diversity of the Helicobacter pylori CagA oncoprotein. Cancer Sci. 2011;102(1):36–43. doi:10.1111/j.1349-7006.2010.01743.x
21. Jones K, Joo Y, Jang S, et al. Polymorphism in the CagA EPIYA motif impacts development of gastric cancer. J Clin Microbiol. 2009;47(4):959–968. doi:10.1128/JCM.02330-08
22. Yin L, Liu F, Guo C, et al. Analysis of virulence diversity of 73 Helicobacter pylori strains isolated in Guizhou province, China. Mol Med Rep. 2018;18(5):4611–4620. doi:10.3892/mmr.2018.9462
23. Hazell SL, Borody TJ, Gal A, Lee A. Campylobacter pyloridis gastritis I: detection of urease as a marker of bacterial colonization and gastritis. Am J Gastroenterol. 1987;82(4):292–296.
24. Hazell SL, Evans DJ
25. Castillo-Juarez I, Rangel-Vega A, Romero I. Rapid paper disk test for identification of Helicobacter pylori in mixed cultures of gerbil gastric homogenates. J Microbiol Methods. 2010;83(1):20–25. doi:10.1016/j.mimet.2010.07.007
26. Thomas P. Isolation of Bacillus pumilus from in vitro grapes as a long-term alcohol-surviving and rhizogenesis inducing covert endophyte. J Appl Microbiol. 2004;97(1):114–123. doi:10.1111/j.1365-2672.2004.02279.x
27. Mégraud F, Lehours P. Helicobacter pylori detection and antimicrobial susceptibility testing. Clin Microbiol Rev. 2007;20(2):280–322.
28. Akopyanz N, Bukanov NO, Westblom TU, Kresovich S, Berg DE. DNA diversity among clinical isolates of Helicobacter pylori detected by PCR-based RAPD fingerprinting. Nucleic Acids Res. 1992;20(19):5137–5142. doi:10.1093/nar/20.19.5137
29. Ben Mansour K, Fendri C, Battikh H, et al. Multiple and mixed Helicobacter pylori infections: comparison of two epidemiological situations in Tunisia and France. Infect Genet Evol. 2016;37:43–48. doi:10.1016/j.meegid.2015.10.028
30. Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233–2239. doi:10.1128/JCM.33.9.2233-2239.1995
31. Sheu SM, Sheu BS, Lu CC, Yang HB, Wu JJ. Mixed infections of Helicobacter pylori: tissue tropism and histological significance. Clin Microbiol Infect. 2009;15(3):253–259. doi:10.1111/j.1469-0691.2008.02666.x
32. Zagari RM, Rabitti S, Eusebi LH, Bazzoli F. Treatment of Helicobacter pylori infection: a clinical practice update. Eur J Clin Invest. 2017;48(1):e12857. doi:10.1111/eci.12857
33. Wong BC, Wang WH, Berg DE, et al. High prevalence of mixed infections by Helicobacter pylori in Hong Kong: metronidazole sensitivity and overall genotype. Aliment Pharmacol Ther. 2001;15(4):493–503. doi:10.1046/j.1365-2036.2001.00949.x
34. Kao CY, Lee AY, Huang AH, et al. Heteroresistance of Helicobacter pylori from the same patient prior to antibiotic treatment. Infect Genet Evol. 2014;23:196–202. doi:10.1016/j.meegid.2014.02.009
35. Kraft C, Suerbaum S. Mutation and recombination in Helicobacter pylori: mechanisms and role in generating strain diversity. Int J Med Microbiol. 2005;295(5):299–305. doi:10.1016/j.ijmm.2005.06.002
36. Atherton JC, Blaser MJ. Coadaptation of Helicobacter pylori and humans: ancient history, modern implications. J Clin Invest. 2009;119(9):2475–2487. doi:10.1172/JCI38605
37. Cooke CL, Huff JL, Solnick JV. The role of genome diversity and immune evasion in persistent infection with. Helicobacter Pylori FEMS Immunol Med Microbiol. 2005;45(1):11–23.
38. Marzio L, Cellini L, Amitrano M, et al. Helicobacter pylori isolates from proximal and distal stomach of patients never treated and already treated show genetic variability and discordant antibiotic resistance. Eur J Gastroenterol Hepatol. 2011;23(6):467–472. doi:10.1097/MEG.0b013e328345d40f
39. Farzi N, Behzad C, Hasani Z, Alebouyeh M, Zojaji H, Zali MR. Characterization of clarithromycin heteroresistance among Helicobacter pylori strains isolated from the antrum and corpus of the stomach. Folia Microbiol (Praha). 2019;64(2):143–151. doi:10.1007/s12223-018-0637-9
40. Burucoa C, Lhomme V, Fauchere JL. Performance criteria of DNA fingerprinting methods for typing of Helicobacter pylori isolates: experimental results and meta-analysis. J Clin Microbiol. 1999;37(12):4071–4080. doi:10.1128/JCM.37.12.4071-4080.1999
41. Morales-Espinosa R, Castillo-Rojas G, Gonzalez-Valencia G, et al. Colonization of Mexican patients by multiple Helicobacter pylori strains with different vacA and cagA genotypes. J Clin Microbiol. 1999;37(9):3001–3004. doi:10.1128/JCM.37.9.3001-3004.1999
42. Patra R, Chattopadhyay S, De R, et al. Multiple infection and microdiversity among Helicobacter pylori isolates in a single host in India. PLoS One. 2012;7(8):e43370. doi:10.1371/journal.pone.0043370
43. Yakoob J, Fan XG, Hu GL, et al. Polycolonization of Helicobacter pylori among Chinese subjects. Clin Microbiol Infect. 2001;7(4):187–192. doi:10.1046/j.1198-743x.2001.00226.x
44. Ren L, Liao YL, Song Y, et al. High frequency variations of Helicobacter pylori isolates in individual hosts in a Chinese population. Int J Infect Dis. 2012;16(5):e358–363. doi:10.1016/j.ijid.2012.01.006
45. Hua J, Ng HC, Yeoh KG, Ho B. Predominance of a single strain of Helicobacter pylori in gastric antrum. Helicobacter. 1999;4(1):28–32. doi:10.1046/j.1523-5378.1999.09043.x
46. Toita N, Yokota S, Fujii N, Konno M. Clonality analysis of helicobacter pylori in patients isolated from several biopsy specimens and gastric juice in a Japanese urban population by random amplified polymorphic DNA fingerprinting. Gastroenterol Res Pract. 2013;2013:721306. doi:10.1155/2013/721306
47. Enroth H, Nyrén O, Engstrand L. One stomach–one strain: does Helicobacter pylori strain variation influence disease outcome? Dig Dis Sci. 1999;44(1):102–107. doi:10.1023/A:1026658301825
48. Qureshi NN, Gallaher B, Schiller NL. Evolution of amoxicillin resistance of Helicobacter pylori in vitro: characterization of resistance mechanisms. Microb Drug Resist. 2014;20(6):509–516. doi:10.1089/mdr.2014.0019
49. Yamaoka Y. Mechanisms of disease: helicobacter pylori virulence factors. Nat Rev Dis Primers. 2010;7(11):629–641.
50. Adam M, Murali B, Glenn NO, Potter SS. Epigenetic inheritance based evolution of antibiotic resistance in bacteria. BMC Evol Biol. 2008;8:52. doi:10.1186/1471-2148-8-52
51. Okamoto T, Yoshiyama H, Nakazawa T, et al. A change in PBP1 is involved in amoxicillin resistance of clinical isolates of. Helicobacter Pylori J Antimicrob Chemother. 2002;50(6):849–856. doi:10.1093/jac/dkf140
52. Saniee P, Hosseini F, Kadkhodaei S, Siavoshi F, Khalili-Samani S. Helicobacter pylori multidrug resistance due to misuse of antibiotics in Iran. Arch Iran Med. 2018;21(7):283–288.
53. Raaf N, Amhis W, Saoula H, et al. Prevalence, antibiotic resistance, and MLST typing of Helicobacter pylori in Algiers, Algeria. Helicobacter. 2017;22:6. doi:10.1111/hel.12446
54. Gong YN, Li YM, Yang NM, et al. Centralized isolation of Helicobacter pylori from multiple centers and transport condition influences. World J Gastroenterol. 2015;21(3):944–952. doi:10.3748/wjg.v21.i3.944
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.