")
Back to Journals » Biologics: Targets and Therapy » Volume 14
Galanin Receptors as Drug Target for Novel Antidepressants: Review
Authors Demsie DG , Altaye BM , Weldekidan E , Gebremedhin H, Alema NM , Tefera MM , Bantie AT
Received 1 December 2019
Accepted for publication 19 March 2020
Published 21 April 2020 Volume 2020:14 Pages 37—45
DOI https://doi.org/10.2147/BTT.S240715
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Doris Benbrook
Desalegn Getnet Demsie,1 Birhanetensay Masresha Altaye,2 Etsay Weldekidan,1 Hagazi Gebremedhin,1 Niguse Meles Alema,3 Mulugeta Mihrete Tefera,3 Abere Tilahun Bantie4
1College of Medicine and Health Sciences, Department of Pharmacy, Adigrat University, Adigrat, Ethiopia; 2College of Health Sciences, Debre Berhan University, Debre Berhan, Ethiopia; 3Bahir Dar Health Science College, Bahir Dar, Ethiopia; 4College of Medicine and Health Sciences, Department of Anesthesia, Adigrat University, Adigrat, Ethiopia
Correspondence: Desalegn Getnet Demsie Tel +251 937404956
Email [email protected]
Abstract: Galanin (GAL) is a 29-amino-acid neuropeptide that serves multiple physiological functions throughout the central and peripheral nervous system. Its role involves in a range of physiological and pathological functions including control of food intake, neuro-protection, neuronal regeneration, energy expenditure, reproduction, water balance, mood, nociception and various neuroendocrine functions. The use of currently available antidepressant drugs raises concerns regarding efficacy and onset of action; therefore, the need for antidepressants with novel mechanisms is increasing. Presently, various studies revealed the link between GAL and depression. Attenuation of depressive symptoms is achieved through inhibition of GalR1 and GalR3 and activation of GalR2. However, lack of receptor selectivity of ligands has limited the complete elucidation of effects of different receptors in depression-like behavior. Studies have suggested that GAL enhances the action of selective serotonin reuptake inhibitors (SSRIs) and promotes availability of transcription proteins. This review addresses the role of GAL, GAL receptors (GALRs) ligands including selective peptides, and the mechanism of ligand receptor interaction in attenuating depressive symptoms.
Keywords: Galanin (GAL), depression, Galanin (GAL) receptors
Introduction
Galanin (GAL) is a neuropeptide with 29 amino acids in length (30 in humans) that co-localize with noradrenaline in the locus coeruleus (LC) neurons and serotonin in the raphe nucleus;1–4 it is found in a wider distribution in the central and peripheral nervous systems.5 The receptors mediating its action have also a different distribution in the central nervous system and the peripheral nervous system as well as distinctive signaling pathways with different biological and pathological effects.
Several studies so far, indicated that GAL is involved in numerous functions such as metabolism, endocrine secretions, intestinal motility, nociception, cognition, feeding behavior, hormone secretion (insulin), nerve regeneration, memory, neuroendocrine release, gut secretion, contractility, and addiction.6–9 Pharmacological and genetic pre-clinical data have also linked the neuroprotective and neuronal regenerative capacity of this neuropeptide and to a range of physiological and pathological functions including epilepsy, chronic anxiety, depression, and pain.9–11 These multitude regulatory effects are mediated through interaction with three G-protein coupled receptors, namely Galanin receptor 1 (GalR1), Galanin receptor 2 (GalR2) and Galanin receptor 3 (GalR3); their roles have been the focus of intense investigation.12–14 The pertussis toxin (PTX) sensitive guanine nucleotide binding proteins, inhibitory G-proteins (Gi/Go) are chiefly involved in the signal transduction mechanism of these receptors.15
Investigations of the mechanism through which GAL acts and the receptor subtypes may represent tremendous potential in neuroscience research and novel drug lead development.16 One of the areas of research interest is depression.17 Stress induces the synthesis and release of GAL with other co-transmitters including serotonin and norepinephrine. A cohort study that enrolled a European white population revealed that variants in genes for GAL and its receptors conferred an increased risk of depression and anxiety in subjects with a history of childhood adversity or recent negative life events. According to the studies, the neuropeptide is involved in the pathogenesis of depressive disorders by maximizing psychosocial stress conditions.16
The role of GAL in learning and memory is implicated by its co-localization in the nucleus basalis neurons of acetylcholine and induction of hyperpolarization in these neurons, accompanying decrease release of acetylcholine in the ventral hippocampus. Centrally administered GAL antagonists attenuate the inhibitory actions and their potential role in dementia has been investigated.18 The neuropeptide, which is thought to be an adrenergic neurotransmitter in the pancreas, potently inhibits insulin secretion by activating ATP sensitive potassium channels.19 The prior addition of receptor antagonist, galantide (500 nmol/l) abolished the changes in [Ca2+] and insulin concentrations.20
Galanin Receptors
Galanin receptors (GALRs) belong to G-protein-couple receptor (GPCR) superfamily that consists of three subfamilies namely GalR1, GalR2 and GalR3.21 All the three receptors are classified under rhodopsin like (class A) GPCRs,17,22,23 alternatively named as seven trans membrane receptors or 7TM based on the notion that seven membrane spanning proteins are constituted in their general structure.23 Their structure is composed of three intercellular loops (ICLs), an extracellular N-terminus, three intracellular loops and three extracellular loops (ECLs).21,24 The helix eight at the C-termini which is located parallel to the membrane is present in all models;21 it is found in the structure of three receptors and acts as a conformational switch.25
Although the three receptors show similarities in their structure as GPCR subclass, distinctly different binding properties to other molecules and effects on intracellular signaling has been documented.21,24 For instance, the lengths of C- and N-termini vary between the three receptor subtypes. The residues in the C-terminal vary within the three receptor subtypes ie, 37 residues in GalR1, 30 residues in GalR2 and 13 residues in GalR3. Likewise, N-terminal which plays an important role in ligand binding is 80 residues long in GalR2, 62 residues in GalR3 but only 47 residues long in GalR1. The receptor subtype is determined by a region between ECL2 and TM helix 7; this region is highly variable and affects ligand binding, cavity size and changing the pattern of ligand interacting residues. Visual inspection revealed the binding cavity in GalR3 as much narrower than in the other two Receptors.21
GalR1 was first cloned from human bowes melanoma cells; it has 7TM domains with extracellular ligand binding N-terminus and intracellular C-terminus.15 Several amino acid residues located in the first and second extracellular TM domains (a pair of cysteine residues presumed to form a disulfide), in the fourth (proline residue), in the fifth and seventh TM domains are thought to introduce a bend in the a-helical structure of the TM and participate in the formation of the binding pocket.15,26 High levels of receptor expression were observed in nucleus of the olfactory tract and olfactory bulb of the olfactory system, cerebral cortex, amygdaloid nucleus, dorsal dentate gyrus and ventral subiculum of hippocampus, thalamus and in some areas of the hypothalamus.
GAL binds to the extracellular N- terminus of GalR1 receptors and elicit conformational change to the signaling molecule. The activated receptors will then bind to G-proteins to form a ternary-complex with GAL and the receptor. Then inhibition of adenyl cyclase (AC) production via the Gi/0 pathway (PTX sensitive Gi/o-pathway) follows and leads to decrease production of a second messenger, cyclic Adenosine Monophosphate (cAMP).15,27 In addition, receptors activation results in the stimulation of the mitogen activated protein kinase (MAPK) pathway, however, activation of the MAPK is independent of protein kinas C (PKC), rather the βγ-subunit is responsible for activation of this pathway.27 This receptor regulates G-protein coupled inwardly rectifying potassium channels.27
Cloning of GalR2 was first done in rat hypothalamus cells12 and then cloned from spleen cells in mice.28 More recently, the receptors have been cloned from a variety of human tissues. Molecular characterization of the receptor indicated four residues, His252 and His253 located in transmembrane domain 6 and Phe264 and Tyr271 in the extracellular loop 3, to be of great significance in the interactions responsible for ligand binding and receptor activation. Receptor occupancy by an agonist results in activation of an effector, phospholipase C (PLC) by which phosphatidylinositol, 4, 5-bisphosphate is converted in to diacylglycerol (DAG) and inositol triphosphate (IP3). IP3 mediated intracellular calcium increment promotes calcium-calmodulin complex formation through which subsequent substrates are activated. On the other hand, DAG located in the cell membrane catalyzes the activation of PKC.29 Generally, an increase in cAMP levels is associated with activation of the stimulatory G-protein (Gs) signaling pathway, leading to activation of AC followed by increased cAMP levels.
By far, the highest levels of GALR2 mRNA expression are observed in the dorsal root ganglia (DRG), hippocampal formation (selectively distributed over the granule cell layer of the dentate gyrus), hypothalamus (present virtually in all hypothalamic nuclei), and cerebellar cortex. Instead of Gs, the high concentrations of Ca2+ intracellularly, βγ G-protein subunit and PKC were presented as a possible candidate to be able to stimulate AC and cause an increase in cAMP levels. GalR2 also mediates a modest inhibition of cAMP Production, demonstrating coupling to Gi.30
GalR3 was first isolated from cDNA libraries of the rat hypothalamus by both homology and expression cloning techniques.13,27,31 Rat GalR3 has 370 amino acids and shares 52–54% and (35–36%) resemblance with GalR2 and GalR1 respectively. Whereas, human GalR3 is comprised of 368 amino acids and shares 90% sequence identity with rat GalR3. GalR3 is widely distributed throughout the median, ventromedian, and ventrolateral preoptic nuclei, medulla oblongata (at the level of the dorsal paragigantocellular field [DPGi]), and medial preoptic region (in the subfornical organ).13,32
Galanin in Depression
Depression is among the most disabling of all medical disorders33 and the most common of the affective disorders.34 Although the exact cause is not exactly known, genetic predisposition, psychosocial stressors and biologic pathophysiology are associated with depressive disorders.35 The symptoms may range from a very mild condition bordering on normality to severe (psychotic) depression which may be accompanied by hallucinations and delusions.34 The pathophysiology of depression remains unclear, but accumulated evidence implicates disturbances in monoaminergic transmission (the monoamine hypothesis) in the brain.36 Currently available medications employed in the treatment of major depression increase the synaptic concentration of monoamines especially norepinephrine and serotonin. For instance, selective serotonin reuptake inhibitors (SSRIs), most widely used to treat depression, enhance the level of a monoamine, serotonin, in the synaptic space.37 In addition to the older idea that a deficit in function or amount of monoamines, there is evidence that neurotrophic and endocrine factors play a major role (the neurotrophic hypothesis).38
Studies focused on the regulatory effects of antidepressants on synaptic function and cellular proteins, less emphasis has been placed on monoaminergic regulation at a more global systemic level, or how such systemic alterations in monoaminergic function might alleviate the behavioral, cognitive, emotional and physiological manifestations of depression and anxiety disorders.39 Although certain highly selective drugs may prove effective, and remain indispensable in the experimental (and clinical) evaluation of the significance of novel mechanisms, a diversity of multi-target strategies are proposed for the more efficacious and rapid control of core and co-morbid symptoms of depression, together with improved tolerance relative to currently available agents.40
Therefore, there is an ongoing search for new medications targeting different sites and potentially modify mood disorders, such as major depression and anxiety.40,41 Neuropeptides and their receptors, the most diverse family of neurotransmitters in the brain, have been extensively explored.26,42 In this regard the neuropeptide GAL is under extensive preclinical investigation.37
The neuropeptide, GAL, upon its action on its three receptors (GalR1-3), participates in numerous physiological and different disease conditions and thus represent tremendous potential in neuroscience research and novel drug lead development.6–8 One of the areas where GAL is involved is depression.43–47 Recent studies have shown the interaction of serotonergic system and neuropeptides to be a key aspect of implication to treat major depression in rodents.48–50
In depression, the neuropeptide modulates the release of neurotransmitters from the LC51 and raphe nucleus52,53 through signal transduction mediated by GalRs (1–3).54 Involvement of GalR1 was explored in a rat model of depression based on chronic mild stress. GAL’s action on GalR1 in ventral periaqueductal gray area was associated with depressive like behavior, possibly representing a novel target for antidepressant therapy.23 Moreover, it modulates the function of serotonin receptor (5-HT1AR) at autoreceptor and postsynaptic levels in the brain areas.53,55–59
As to previous studies, investigations were underway to confirm the mechanism through which receptors mediate signal transductions.15,27,29 Interestingly, a recent study confirmed the inhibitory action of GalR1 and GalR3 on receptor knockdown from a brain slice of Sprague Dawley rats after neurons failed to exhibit a significant neural firing. In addition, the inhibitory effect of GAL was abolished by treatment of PTX, indicating that inhibitory g-protein (Gi) is required for GalR1 mediated hyperpolarization; its effect is also blocked by GIRK blockers.29,51,54 Bai and colleagues also used siRNA technique to knockdown GalR1 and GalR2 in a rat's brain slice. They confirmed that induced hyperpolarization was significantly decreased in GalR1 knockdown brain slices while there was not significant change in hyperpolarization in GalR2 knockdown and the control group.51
GalR2 induced excitatory signaling is mediated via coupling to Gq/G11 thereby increasing neural reactivity in different brain regions.32 On the contrary, GalR1 and GalR3 couples to Gi/Go to induce cellular hyperpolarization.27,60 Gi/o induced hyperpolarization leads to increased potassium conductance through inwardly rectifying potassium channels and GIRK channels.51 Consequently, neural cell hyperpolarization inhibits neurotransmitter release from the LC and raphe nucleus, notably norepinephrine and serotonin.61,62
The pro-depressant effect of GalR1 is attributed to its ability to modulate 5-Hydroxytryptamine-1A (5-HT1AR) signaling owing to its hyperpolarizing and ability of forming a complex with 5-HT1AR to inhibit release and binding, and reduction of serotonin metabolism in ventral limbic cortex, the hippocampal formation, and frontal parietal cortex.52 The interaction of GALRs with 5- HT1A receptors was found in the paraventricular thalamic nucleus, the nucleus reuniens and rhomboideus, the zona incerta, the medial and the lateral hypothalamus, and the medial and the lateral amygdaloid area, but not in the pars compacta of the substantia nigra, which lacks 5-HT1AR binding sites.63
The binding characteristics of the neuropeptide were analyzed on 5-HT receptor subtypes in the ventral limbic cortex of the rat;52 and it has been demonstrated that GalR1-5-HT1AR heteromerize.64 It was found that GAL markedly decreased the affinity value of 5-HT1AR binding, while there were no effects on the binding characteristics of the 5-HT1B or 5-HT2 radio ligands. This GalR1-5-HT1AR interaction is located in dorsal raphe 5-HT1AR cells that innervate large parts of forebrain.52,53
Thus, GAL upon receptor–receptor interaction with 5-HT1AR induces hyperpolarization of serotonergic neurons it reduces the binding affinity of post-junctional and serotonin autoreceptor. Its binding to GalR1 receptor subtype causes GalR1-5-HT1AR activation via Gi/o-AC-PKA cascades, thereby decreasing the phosphorylation of cyclic AMP response element binding protein (CREB) that leads to a reduced gene transcription such as anti-apoptotic proteins.23,64 Regulation of CREB expression by members of the diverse family of neuropeptides, including GAL, are co-expressed with noradrenaline and serotonin, modulates synaptic and other neuronal structure activities by enhancing the firing, neurogenesis and neuronal plasticity.23
The implication of GAL in the regulation of affective behavior, including modulation of 5-HT signaling was also investigated in various central and peripheral nervous system regions including, limbic and basal forebrain, hypothalamus/mammillary bodies, midbrain, pons and medulla and lamina I–V.65 The investigation of GAL on 5-HT level in specific brain parts indicated a 79% decrease in serotonin levels in the medial prefrontal cortex (mPFC) and the ventral hippocampus (vHPC) to 72% after intracerebroventricular (ICV) infusion. Local infusions caused a long-lasting decrease in 5-HT levels in vHPC to 88%, and a moderate decrease in central amygdaloid nucleus (CeA), whereas the 5-HT levels in mPFC significantly increased to 121%. These effects of ICV GAL correlated well with the density of 5-HT and GALnerve terminals, and GALRs autoradiography in mPFC, vHPC, and CeA. GAL was often found to coexist in NA, but could never be detected in 5-HT terminals.66
Moreover, effects of receptor agonists on LC neurons are significant for its role in participation of depressive disorders.1,67 It can be synthesized in several ascending systems including cholinergic neurons, serotonergic dorsal raphe neurons, and the noradrenergic LC systems. The LC neurons express the highest levels of GAL and that in cortex and hippocampus GAL peptide can only be detected in the noradrenergic projections.68,69 Electro-physiologic studies show that the neuropeptide hyperpolarizes both serotonergic dorsal raphe neurons and noradrenergic LC neurons at fairly very high concentrations (106−10−7 M). In addition, it enhances the 5-HT and noradrenaline induced hyperpolarization at low concentrations (10−9 M). Consequently, a receptor antagonist could attenuate an inhibitory tone on both dorsal raphe and LC neurons and thus perhaps exert antidepressant activity.67
The inhibitory effect on LC neurons is mainly GalR1 receptor mediated. In in vitro recording technique, the action of the two receptor agonists namely, AR-M961 and AR-M1896 was investigated. Accordingly, application of AR-M961, an agonist both at GalR1, GalR2, evoked a reversible membrane hyperpolarization and inhibition of spike discharge in all LC neurons, whereas AR-M961, the selective GalR2 agonist (AR-M1896) only caused a slight hyperpolarization as compared to AR-M961.70 Immunohistochemical staining of intracellular filled neurons indicate that the neuropeptide exerts an inhibitory effect on norepinephrine neurons of the LC via increase in potassium conductance.71
Not only GAL, but also Galanin N-terminal fragments like Galanin 1–15 (GAL1-15) are active at the central level to elicit GAL like effects.47,49,72 Interaction of GAL (1–15) with GalR1-GalR2 isoreceptor dimers results in depression like and anxiogenic effects to a greater extent than GAL.46,73 GALRs and neuropeptide Y Y1 (NPYY1) receptor interaction may also play a role in the pathophysiology of mood disorders, including depression and anxiety.9,74–76 Narvaez et al confirmed the interaction between GalR2 and NPYY1R in the dentate gyrus (DG) with enhancement of the antidepressive-like behavior mediated by NPY Y1R77 and anxiolytic behavior.78 Moreover, GalR1-GalR2 heteromer interaction with Neuropeptide Y Y2 (NPYY2) may be a key molecular mechanism for GAL and its GAL1-15.79
Furthermore, GAL1-15 fragments facilitate GalR1-5-HT1AR heteroreceptor complexes formation in the raphe-hippocampal 5-HT neurons and affects serotonin release; GAL1–15 induces stronger effects than GAL to cause depression.72 The presence of these heteromers in the discrete brain regions help to explore possible novel therapeutic strategies for treatment of depression by targeting the GalR1-5-HT1AR heteromers.80 The inhibition of CREB by 50 nM of GAL1–15 and GAL1–29 was fully counteracted by the non-selective receptor antagonist M35 and the selective GalR2 antagonist, M871.This misbalance in the signaling of the GalR1–GalR2 heteroreceptor complexes induced by GAL1–15 may contribute to depression-like actions since GalR1 agonists produce such effects.79 The absence of an additive or a synergistic interaction upon coactivation of the two receptors suggests the existence of an allosteric inhibitory communication in the interface between the two receptors of the heteromer.79,80
Molecular studies showed that GAL1-15 increased post-junctional mRNA levels of 5-HT1AR while the density of autoreceptors is decreased.46,49,81 In line with this, the existence of GAL-5HT1AR heterorecptor complex dysfunction leads to disturbance in mesolimbic neurotransmission of 5-HT.82,83 Indeed, the modulation of auto-receptor function is distinctly regulated by the GalR1-GalR2-5-HT1AR heterotrimeric complex to elicit antidepressant effects.46,83
Besides increasing hippocampal mRNA levels of post junctional serotonin receptors, co-administration of GAL1-15 and fluoxetine (FLX) help to enhance the agonist binding affinity of FLX in the dentate gyrus.81 According to the findings by Flores-Burgess et al the combination use of the three sc injections of FLX (10 mg/kg) and a single ICV injection of GAL1–15 (1 nmol) produced a significant increase in the 5-HT1AR mRNA levels in the median prefrontal cortex with a significant increase in the Kd value (F3,20 = 14.36, p < 0.001; post hoc p < 0.01) in mPFC (F3,19 = 6.418, p < 0.01; post hoc p < 0.01).84
The existence of 5-HT1AR-5-HT2A isoreceptor complexes has also been considered as a potential drug target for antidepressants. 5-HT2A agonist, TCB2, significantly reduced the binding affinity of ipsapirone (5-HT1AR agonist); this action was blocked by the 5-HT2A antagonist ketanserin.81 Of course, previous studies showed that some antidepressants block 5-HT2A receptors while others elicit antidepressant action via activation of 5-HT1AR.85 In line with the aforementioned explanations, various ligands, models and their effects, including the action of synthetic peptide, J1817 are presented in Table 1.
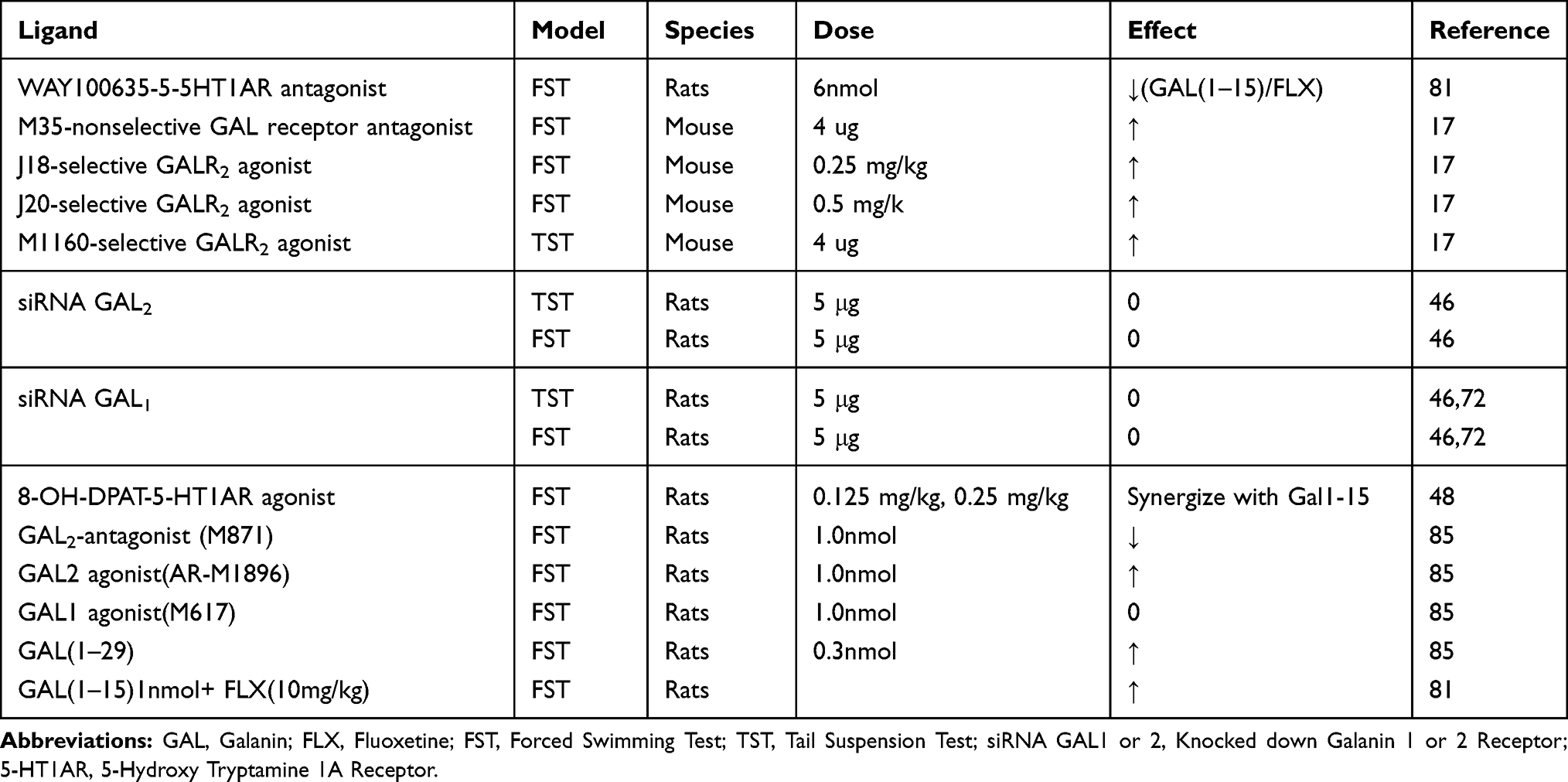 |
Table 1 Effects of Galanin Receptor Ligands and Animal Models in Rodent Test of Depression |
Conclusion
GAL produces mixed depressive and anti-depressant effects in preclinical studies. The existence of iso and hetero receptor dimers, and difference in the distribution of receptor subtypes in discrete brain regions confers the neuropeptide inhibitory or stimulatory actions on the function of neurons. GalR1 and GalR3 mediate antidepressant action while GAL binding to GalR2 elicits depressive like effects. Likewise, GAL1-15 has been considered as a promising novel ligand in mood disorders. It increases expression of 5-HT1R and enhance binding affinity of SSRIs. Interaction between GalR2 and NPYY1R results in enhancement of the antidepressive-like behavior mediated by NPY Y1R and anxiolytic behavior. Moreover, GalR1-GalR2 heteromer interaction with NPYY2 may be a key molecular target for GAL and GAL1-15.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Melander T, Hökfelt T, Rökaeus A. Distribution of galaninlike immunoreactivity in the rat central nervous system. J Comp Neurol. 1986;248(4):475–517. doi:10.1002/cne.v248:4
2. Fuxe K, Agnati LF, Von Euler G, et al. Galanin/5-HT receptor interactions. A new integrative mechanism in the control of 5-HT neurotransmission in the central nervous system. In: Paoletti R,Vonhoutte PM,Brunello N, Maggi FM, editors. Serotonin. Dordrecht: Springer; 1990:169–185.
3. Xu IS, Grass S, Xu XJ, Wiesenfeld-Hallin Z. On the role of galanin in mediating spinal flexor reflex excitability in inflammation. Neurosci. 1998;85(3):827–835. doi:10.1016/S0306-4522(97)00676-3
4. Lu X, Barr AM, Kinney JW, et al. A role for galanin in antidepressant actions with a focus on the dorsal raphe nucleus. Proc Natl Acad Sci. 2005;102(3):874–879. doi:10.1073/pnas.0408891102
5. Kask K, Langel Ü, Bartfai T. Galanin—a neuropeptide with inhibitory actions. Cell Mol Neurobiol. 1995;15(6):653–673. doi:10.1007/BF02071130
6. Lundkvist J, Land T, Kahl U, Bedecs K, Bartfai T. cDNA sequence, ligand binding, and regulation of galanin/GMAP in mouse brain. Neurosci Lett. 1995;200(2):121–124. doi:10.1016/0304-3940(95)12094-K
7. Kakuyama H, Kuwahara A, Mochizuki T, Hoshino M, Yanaihara N. Role of N-terminal active sites of galanin in neurally evoked circular muscle contractions in the guinea-pig ileum. Eur J Pharmacol. 1997;329(1):85–91. doi:10.1016/S0014-2999(97)10109-1
8. Smith KE, Walker MW, Artymyshyn R, et al. Cloned human and rat galanin GALR3 receptors pharmacology and activation of G-protein inwardly rectifying K+ channels. J Biol Chem. 1998;273(36):23321–23326. doi:10.1074/jbc.273.36.23321
9. Lang R, Gundlach AL, Kofler B. The galanin peptide family: receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharmacol Ther. 2007;115(2):177–207. doi:10.1016/j.pharmthera.2007.05.009
10. Ögren SO, Kuteeva E, Elvander-Tottie E, Hökfelt T. Neuropeptides in learning and memory processes with focus on galanin. Eur J Pharmacol. 2010;626(1):9–17. doi:10.1016/j.ejphar.2009.09.070
11. Picciotto MR, Brabant C, Einstein EB, Kamens HM, Neugebauer NM. Effects of galanin on monoaminergic systems and HPA axis: potential mechanisms underlying the effects of galanin on addiction-and stress-related behaviors. Brain Res. 2010;1314:206–218.
12. Smith KE, Forray C, Walker MW, et al. Expression cloning of a rat hypothalamic galanin receptor coupled to phosphoinositide turnover. J Biol Chem. 1997;272(39):24612–24616. doi:10.1074/jbc.272.39.24612
13. Smith KE, Walker MW, Artymyshyn R, et al. Cloned human and rat galanin GALR3 receptors Pharmacology and activation of G-protein inwardly rectifying K+ channels. Journal of Biological Chemistry. 1998;273(36):23321-23326.
14. Iismaa TP, Shine J. Galanin and galanin receptors. In:Richter D, editor. Regulatory Peptides and Cognate Receptors. Berlin, Heidelberg: Springer; 1999;26:257–291.
15. Habert-Ortoli E, Amiranoff B, Loquet I, Laburthe M, Mayaux JF. Molecular cloning of a functional human galanin receptor. Proc Natl Acad Sci. 1994;91(21):9780–9783. doi:10.1073/pnas.91.21.9780
16. Juhasz G, Hullam G, Eszlari N, et al. Brain galanin system genes interact with life stresses in depression-related phenotypes. Proc Natl Acad Sci. 2014;111(16):E1666–73. doi:10.1073/pnas.1403649111
17. Saar I, Lahe J, Langel K, et al. Novel systemically active galanin receptor 2 ligands in depression‐like behavior. J Neurochem. 2013;127(1):114–123. doi:10.1111/jnc.12274
18. Crawley JN, Wenk GL. Co-existence of galanin and acetylcholine: is galanin involved in memory processes and dementia? Trends Neurosci. 1989;12(8):278–282. doi:10.1016/0166-2236(89)90003-9
19. Dunne MJ, Bullett MJ, Li GD, Wollheim CB, Petersen OH. Galanin activates nucleotide‐dependent K+ channels in insulin‐secreting cells via a pertussis toxin‐sensitive G‐protein. EMBO J. 1989;8(2):413–420. doi:10.1002/embj.1989.8.issue-2
20. Fridolf T, Ahrén B. Dual action of the neuropeptide galanin on the cytoplasmic free calcium concentration in RIN m5F cells. Biochem Biophys Res Commun. 1993;191(3):1224–1229. doi:10.1006/bbrc.1993.1348
21. Jurkowski W, Yazdi S, Elofsson A. Ligand binding properties of human galanin receptors. Mol Membr Biol. 2013;30(2):206–216. doi:10.3109/09687688.2012.750384
22. Guipponi M, Chentouf A, Webling KE, et al. Galanin pathogenic mutations in temporal lobe epilepsy. Hum Mol Genet. 2015;24(11):3082–3091.
23. Wang P, Li H, Barde S, et al. Depression-like behavior in rat: involvement of galanin receptor subtype 1 in the ventral periaqueductal gray. Proc Natl Acad Sci. 2016;113(32):E4726–35. doi:10.1073/pnas.1609198113
24. Probst WC, Snyder LA, Schuster DI, Brosius J, Sealfon SC. Sequence alignment of the G-protein coupled receptor superfamily. DNA Cell Biol. 1992;11(1):1–20. doi:10.1089/dna.1992.11.1
25. Krishna AG, Menon ST, Terry TJ, Sakmar TP. Evidence that helix 8 of rhodopsin acts as a membrane-dependent conformational switch. Biochemistry. 2002;41(26):8298–8309. doi:10.1021/bi025534m
26. Burbach JP. Neuropeptides from concept to online database www.neuropeptides.nl. Eur J Pharmacol. 2010;626(1):27–48. doi:10.1016/j.ejphar.2009.10.015
27. Wang S, He C, Maguire MT, et al. Genomic organization and functional characterization of the mouse GalR1 galanin receptor. FEBS Lett. 1997;411(2–3):225–230. doi:10.1016/S0014-5793(97)00695-9
28. Pang L, Hashemi T, Lee HJ, et al. The mouse GalR2 galanin receptor: genomic organization, cDNA cloning, and functional characterization. J Neurochem. 1998;71(6):2252–2259. doi:10.1046/j.1471-4159.1998.71062252.x
29. Pan J, Singh US, Takahashi T, et al. PKC mediates cyclic stretch‐induced cardiac hypertrophy through Rho family GTPases and mitogen‐activated protein kinases in cardiomyocytes. J Cell Physiol. 2005;202(2):536–553. doi:10.1002/jcp.20151
30. Mons N, Decorte L, Jaffard R, Cooper DM. Ca2+-sensitive adenylyl cyclases, key integrators of cellular signalling. Life Sci. 1998;62(17–18):1647–1652. doi:10.1016/S0024-3205(98)00122-2
31. Wang S, Hashemi T, Fried S, Clemmons AL, Hawes BE. Differential intracellular signaling of the GalR1 and GalR2 galanin receptor subtypes. Biochemistry. 1998;37(19):6711–6717. doi:10.1021/bi9728405
32. Kolakowski LF
33. Charney DS, Manji HK. Life stress, genes, and depression: multiple pathways lead to increased risk and new opportunities for intervention. Sci Stke. 2004;2004(225):re5.
34. Rang HP, Ritter JM, Flower RJ, Henderson G. Rang and Dale’s Pharmacology.
35. Kendler KS. The dappled nature of causes of psychiatric illness: replacing the organic–functional/hardware–software dichotomy with empirically based pluralism. Mol Psychiatry. 2012;17(4):377–388. doi:10.1038/mp.2011.182
36. Liu M, Simon MI. Regulation by cAMP-dependent protein kinease of a G-protein-mediated phospholipase C. Nature. 1996;382(6586):83–87. doi:10.1038/382083a0
37. Tatemoto K, Rökaeus Å, Jörnvall H, McDonald TJ, Mutt V. Galanin—a novel biologically active peptide from porcine intestine. FEBS Lett. 1983;164(1):124–128. doi:10.1016/0014-5793(83)80033-7
38. Katzung BG. Basic and Clinical Pharmacology. Mc Graw Hill; 2012.
39. Morilak DA, Frazer A. Antidepressants and brain monoaminergic systems: a dimensional approach to understanding their behavioural effects in depression and anxiety disorders. Int J Neuropsychopharmacol. 2004;7(2):193–218. doi:10.1017/S1461145704004080
40. Millan MJ. Multi-target strategies for the improved treatment of depressive states: conceptual foundations and neuronal substrates, drug discovery and therapeutic application. Pharmacol Ther. 2006;110(2):135–370. doi:10.1016/j.pharmthera.2005.11.006
41. Maes M, Fišar Z, Medina M, Scapagnini G, Nowak G, Berk M. New drug targets in depression: inflammatory, cell-mediated immune, oxidative and nitrosative stress, mitochondrial, antioxidant, and neuroprogressive pathways. And new drug candidates—Nrf2 activators and GSK-3 inhibitors. Inflammopharmacology. 2012;20(3):127–150.
42. Trivedi MH, Rush AJ, Wisniewski SR, et al. Evaluation of outcomes with citalopram for depression using measurement-based care in STAR* D: implications for clinical practice. Am J Psychiatry. 2006;163(1):28–40. doi:10.1176/appi.ajp.163.1.28
43. Diaz-Cabiale Z, Parrado C, Fuxe K, Agnati L, Narvaez JA. Receptor–receptor interactions in central cardiovascular regulation. Focus on neuropeptide/α 2-adrenoreceptor interactions in the nucleus tractus solitarius. J Neural Transm. 2007;114(1):115–125. doi:10.1007/s00702-006-0559-6
44. Díaz-Cabiale Z, Parrado C, Narváez M, et al. Neurochemical modulation of central cardiovascular control: the integrative role of galanin. In: Hokfelt T, editor. Galanin. Basel: Springer; 2010;102:113–131.
45. Díaz-Cabiale Z, Parrado C, Narváez M, et al. Galanin receptor/neuropeptide Y receptor interactions in the dorsal raphe nucleus of the rat. Neuropharmacol. 2011;61(1–2):80–86. doi:10.1016/j.neuropharm.2011.03.002
46. Millón C, Flores-Burgess A, Narváez M, et al. A role for galanin N-terminal fragment (1–15) in anxiety-and depression-related behaviors in rats. Int J Neuropsychopharmacol. 2015;18(3):pyu064. doi:10.1093/ijnp/pyu064
47. Weiss JM, Bonsall RW, Demetrikopoulos MK, Emery MS, West CH. Galanin: a significant role in depression? Ann N Y Acad Sci. 1998;863(1):364–382. doi:10.1111/j.1749-6632.1998.tb10707.x
48. Millón C, Flores-Burgess A, Narváez M, et al. (1–15) enhances the antidepressant effects of the 5-HT1A receptor agonist 8-OH-DPAT: involvement of the raphe-hippocampal 5-HT neuron system. Brain Struct Funct. 2016;221(9):4491–4504. doi:10.1007/s00429-015-1180-y
49. Jackowski A, Crockard A, Burnstock G, Lincoln J. Alterations in serotonin and neuropeptide Y content of cerebrovascular sympathetic nerves following experimental subarachnoid hemorrhage. J Cereb Blood Flow Metab. 1989;9(3):271–279. doi:10.1038/jcbfm.1989.45
50. Terenina NB, Kreshchenko ND, Mochalova NB, Movsesyan SO. Serotonin and neuropeptide FMRFamide in the attachment organs of trematodes. Helminthologia. 2018;55(3):185–194. doi:10.2478/helm-2018-0022
51. Bai YF, Ma HT, Liu LN, et al. Activation of galanin receptor 1 inhibits locus coeruleus neurons via GIRK channels. Biochem Biophys Res Commun. 2018;503(1):79–85. doi:10.1016/j.bbrc.2018.05.181
52. Morais JS, Souza MM, Campanha TM, et al. Galanin subtype 1 and subtype 2 receptors mediate opposite anxiety-like effects in the rat dorsal raphe nucleus. Behav Brain Res. 2016;314:125–133. doi:10.1016/j.bbr.2016.08.007
53. Razani H, Diaz-Cabiale Z, Fuxe K, Ögren SO. Intraventricular galanin produces a time-dependent modulation of 5-HT1A receptors in the dorsal raphe of the rat. Neuroreport. 2000;11(18):3943–3948. doi:10.1097/00001756-200012180-00008
54. Lang R, Gundlach AL, Holmes FE, et al. Physiology, signaling, and pharmacology of galanin peptides and receptors: three decades of emerging diversity. Pharmacol Rev. 2015;67(1):118–175.
55. Fuxe K, Ögren SO, Jansson A, Cintra A, Härfstrand A, Agnati LF. Intraventricular injections of galanin reduces 5‐HT metabolism in the ventral limbic cortex, the hippocampal formation and the fronto‐parietal cortex of the male rat. Acta Physiol Scand. 1988;133(4):579–581. doi:10.1111/j.1748-1716.1988.tb08444.x
56. Hedlund PB, Galanin FK. 5‐HT1A Receptor interactions as an integrative mechanism in 5‐HT neurotransmission in the brain a. Ann N Y Acad Sci. 1996;780(1):193–212. doi:10.1111/j.1749-6632.1996.tb15124.x
57. Kehr J, Yoshitake T, Wang FH, et al. Galanin is a potent in vivo modulator of mesencephalic serotonergic neurotransmission. Neuropsychopharmacol. 2002;27(3):341–356. doi:10.1016/S0893-133X(02)00309-3
58. Misane I, Razani H, Jansson FH, Fuxe K, Ögren SO. Intraventricular galanin modulates a 5‐HT1A receptor‐mediated behavioural response in the rat. Eur J Neurosci. 1998;10(4):1230–1240. doi:10.1046/j.1460-9568.1998.00132.x
59. Razani H, Dı́az-Cabiale Z, Misane I, Wang FH, Fuxe K, Ögren SO. Prolonged effects of intraventricular galanin on a 5-hydroxytryptamine1A receptor mediated function in the rat. Neurosci Lett. 2001;299(1–2):145–149. doi:10.1016/S0304-3940(00)01788-2
60. Parker EM, Izzarelli DG, Nowak HP, et al. Cloning and characterization of the rat GALR1 galanin receptor from Rin14B insulinoma cells. Mol Brain Res. 1995;34(2):179–189. doi:10.1016/0169-328X(95)00159-P
61. Bartfai T, Lu X, Badie-Mahdavi H, et al. Galmic, a nonpeptide galanin receptor agonist, affects behaviors in seizure, pain, and forced-swim tests. Proc Natl Acad Sci. 2004;101(28):10470–10475. doi:10.1073/pnas.0403802101
62. Kuteeva E, Hökfelt T, Wardi T, Ogren SO. Galanin, galanin receptor subtypes and depression-like behaviour. Cell Mol Life Sci CMLS. 2008;65(12):1854–1863. doi:10.1007/s00018-008-8160-9
63. Hedlund P, Von Euler G, Fuxe K. Activation of 5-hydroxytryptamine1A receptors increases the affinity of galanin receptors in di-and telencephalic areas of the rat. Brain Res. 1991;560(1–2):251–259. doi:10.1016/0006-8993(91)91240-2
64. Borroto-Escuela DO, Narvaez M, Marcellino D, et al. Galanin receptor-1 modulates 5-hydroxtryptamine-1A signaling via heterodimerization. Biochem Biophys Res Commun. 2010;393(4):767–772. doi:10.1016/j.bbrc.2010.02.078
65. Mennicken F, Hoffert C, Pelletier M, Ahmad S, O’Donnell D. Restricted distribution of galanin receptor 3 (GalR3) mRNA in the adult rat central nervous system. J Chem Neuroanat. 2002;24(4):257–268. doi:10.1016/S0891-0618(02)00068-6
66. Yoshitake S, Kuteeva E, Hökfelt T, et al. Correlation between the effects of local and intracerebroventricular infusions of galanin on 5‐HT release studied by microdialysis, and distribution of galanin and galanin receptors in prefrontal cortex, ventral hippocampus, amygdala, hypothalamus, and striatum of awake rats. Synapse. 2014;68(5):179–193. doi:10.1002/syn.21730
67. HöKFELT TO, ZQ XU, SHI TJ, Holmberg K, Zhang X. Galanin in ascending systems: focus on coexistence with 5‐hydroxytryptamine and noradrenaline a. Ann N Y Acad Sci. 1998;863(1):252–263. doi:10.1111/j.1749-6632.1998.tb10700.x
68. Seutin V, Verbanck P, Massotte L, Dresse A. Galanin decreases the activity of locus coeruleus neurons in vitro. Eur J Pharmacol. 1989;164(2):373–376. doi:10.1016/0014-2999(89)90481-0
69. Hökfelt T, Broberger C, Diez M, et al. NPY, two peptides with multiple putative roles in the nervous system. Horm Metab Res. 1999;31(5):330–334. doi:10.1055/s-2007-978748
70. Ma X, Tong YG, Schmidt R, et al. Effects of galanin receptor agonists on locus coeruleus neurons. Brain Res. 2001;919(1):169–174. doi:10.1016/S0006-8993(01)03033-5
71. Pieribone VA, Xu ZQ, Zhang X, Grillner S, Bartfai T, Hökfelt T. Galanin induces a hyperpolarization of norepinephrine-containing locus coeruleus neurons in the brainstem slice. Neurosci. 1995;64(4):861–874. doi:10.1016/0306-4522(94)00450-J
72. Millón C, Flores-Burgess A, Narváez M, et al. The neuropeptides galanin and galanin (1–15) in depression-like behaviours. Neuropeptides. 2017;64:39–45. doi:10.1016/j.npep.2017.01.004
73. Borroto-Escuela DO, Narvaez M, Di Palma M, et al. Preferential activation by galanin 1–15 fragment of the GalR1 protomer of a GalR1–GalR2 heteroreceptor complex. Biochem Biophys Res Commun. 2014;452(3):347–353. doi:10.1016/j.bbrc.2014.08.061
74. Díaz‐Cabiale Z, Parrado C, Rivera A, et al. (NPY) interactions in central cardiovascular control: involvement of the NPY Y1 receptor subtype. Eur J Neurosci. 2006;24(2):499–508. doi:10.1111/j.1460-9568.2006.04937.x
75. Fuxe K, Marcellino D, Rivera A, et al. Receptor–receptor interactions within receptor mosaics. Impact on neuropsychopharmacology. Brain Res Rev. 2008;58(2):415–452. doi:10.1016/j.brainresrev.2007.11.007
76. Branchek TA, Smith KE, Gerald C, Walker MW. Galanin receptor subtypes. Trends Pharmacol Sci. 2000;21(3):109–117. doi:10.1016/S0165-6147(00)01446-2
77. Narváez M, Borroto-Escuela DO, Millón C, et al. Galanin receptor 2-neuropeptide Y Y1 receptor interactions in the dentate gyrus are related with antidepressant-like effects. Brain Struct Funct. 2016;221(8):4129–4139. doi:10.1007/s00429-015-1153-1
78. Narváez M, Borroto-Escuela DO, Santín L, et al. A novel integrative mechanism in anxiolytic behavior induced by galanin 2/neuropeptide Y Y1 receptor interactions on medial paracapsular intercalated amygdala in rats. Front Cell Neurosci. 2018;1(12):119. doi:10.3389/fncel.2018.00119
79. Díaz-Cabiale Z, Flores-Burgess A, Parrado C, et al. Galanin receptor/neuropeptide y receptor interactions in the central nervous system. Curr Protein Pept Sci. 2014;15(7):666–672. doi:10.2174/1389203715666140901111709
80. Kummer W. Galanin-and neuropeptide Y-like immunoreactivities coexist in paravertebral sympathetic neurones of the cat. Neurosci Lett. 1987;78(2):127–131. doi:10.1016/0304-3940(87)90620-3
81. Flores-Burgess A, Millón C, Gago B, et al. (1–15) enhancement of the behavioral effects of fluoxetine in the forced swimming test gives a new therapeutic strategy against depression. Neuropharmacol. 2017;118:233–241. doi:10.1016/j.neuropharm.2017.03.010
82. Borroto-Escuela DO, Li X, Tarakanov AO, et al. Existence of brain 5-HT1A–5-HT2A isoreceptor complexes with antagonistic allosteric receptor–receptor interactions regulating 5-HT1A receptor recognition. ACS Omega. 2017;2(8):4779–4789. doi:10.1021/acsomega.7b00629
83. Borroto-Escuela DO, Narváez M, Ambrogini P, et al. Receptor–receptor interactions in multiple 5-HT1A heteroreceptor complexes in raphe-hippocampal 5-HT transmission and their relevance for depression and its treatment. Mol. 2018;23(6):1341. doi:10.3390/molecules23061341
84. Flores-Burgess A, Millón C, Gago B, et al. (1–15)-fluoxetine interaction in the novel object recognition test. Involvement of 5-HT1A receptors in the prefrontal cortex of the rats. Neuropharmacol. 2019;155:104–112. doi:10.1016/j.neuropharm.2019.05.023
85. de Souza MM, Silote GP, Herbst LS, Funck VR, Joca SRL, Beijamini V. The antidepressant-like effect of galanin in the dorsal raphe nucleus of rats involves GAL(2) receptors. Neurosci Lett. 2018;681:26–30. doi:10.1016/j.neulet.2018.05.029.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.