")
Back to Journals » Drug Design, Development and Therapy » Volume 9
Zinc supplementation induces apoptosis and enhances antitumor efficacy of docetaxel in non-small-cell lung cancer
Authors Kocdor H , Ates H, Aydin S , Cehreli R, Soyarat F, Kemanli P, Harmanci D, Cengiz H , Kocdor M
Received 30 April 2015
Accepted for publication 10 June 2015
Published 27 July 2015 Volume 2015:9 Pages 3899—3909
DOI https://doi.org/10.2147/DDDT.S87662
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Wei Duan
Hilal Kocdor,1,2 Halil Ates,1 Suleyman Aydin,3 Ruksan Cehreli,1 Firat Soyarat,2 Pinar Kemanli,2 Duygu Harmanci,2 Hakan Cengiz,2 Mehmet Ali Kocdor4
1Institute of Oncology, Dokuz Eylul University, 2Department of Molecular Medicine, Institute of Health Sciences, Dokuz Eylul University, Izmir Turkey; 3Department of Biochemistry, Firat University School of Medicine, Elazig, 4Department of Surgery, School of Medicine, Dokuz Eylul University, Izmir, Turkey
Background: Exposure to exogenous zinc results in increased apoptosis, growth inhibition, and altered oxidative stress in cancer cells. Previous studies also suggested that zinc sensitizes some cancer cells to cytotoxic agents depending on the p53 status. Therefore, zinc supplementation may show anticancer efficacy solely and may increase docetaxel-induced cytotoxicity in non-small-cell lung cancer cells.
Methods: Here, we report the effects of several concentrations of zinc combined with docetaxel on p53-wild-type (A549) and p53-null (H1299) cells. We evaluated cellular viability, apoptosis, and cell cycle progression as well as oxidative stress parameters, including superoxide dismutase, glutathione peroxidase, and malondialdehyde levels.
Results: Zinc reduced the viability of A549 cells and increased the apoptotic response in both cell lines in a dose-dependent manner. Zinc also amplified the docetaxel effects and reduced its inhibitory concentration 50 (IC50) values. The superoxide dismutase levels increased in all treatment groups; however, glutathione peroxidase was slightly increased in the combination treatments. Zinc also caused malondialdehyde elevations at 50 µM and 100 µM.
Conclusion: Zinc has anticancer efficacy against non-small-cell lung cancer cells in the presence of functionally active p53 and enhances docetaxel efficacy in both p53-wild-type and p53-deficient cancer cells.
Keywords: lung cancer, zinc, docetaxel, A549, H1299
Introduction
Lung cancer is the most commonly diagnosed cancer. It is the leading cause of cancer-related deaths in males and the second cause of cancer-related deaths in females worldwide.1 Non-small-cell histology comprises ~85% of lung cancers, and 75% of patients are diagnosed at stages III and IV.2,3 At the advanced stages, taxane chemotherapy regimens are commonly used for the treatment of non-small-cell lung cancers (NSCLCs) as first-line options.4 However, long-term survival in patients with advanced NSCLC is <5% and toxic. Side effects such as febrile neutropenia, neuropathy, and hypersensitivity reactions are particularly high in taxane formulations. Furthermore, the therapeutic results are not satisfactory.4,5
Paclitaxel and its semisynthetic form, docetaxel, are isolated from the yew tree and primarily stabilize cytoplasmic microtubules via binding to the β-tubulin site, thereby causing cell cycle arrest at the G2/M phase and driving apoptosis.6 Several compounds have been tested to determine whether they increase taxane-induced anticancer efficacy. For example, the therapeutic efficacy of paclitaxel is restricted by the increasing frequency of chemotherapeutic resistance in NSCLC. Pulsing with both taxanes results in approximately fivefold higher resistant clones in NSCLC cells, such as A549 and H1299 cells.7 Several compounds, such as l-type calcium channel blockers, reverse docetaxel-induced multidrug resistance independent of ABCB1 expression in both cell lines.8 Additionally, the histone deacetylase inhibitor trichostatin A increases both cell cycle delay at the G2/M phase and apoptosis in docetaxel-treated A549 cells.9 In hormone-refractory prostate cancer (HRPC) models, taxol synergizes with several antioxidants in HRPC cells by inducing cell cycle arrest at the sub G1 phase, apoptosis, and caspase activity and decreasing Bcl-2 expression simultaneously.10 By contrast, TP53 mutations are the most frequent gene abnormalities leading to inactivation of p53, which effects paclitaxel sensitivity in NSCLC.11
A trace element, zinc, is essential for the wide range of physiological processes, including growth, development, and immune functions as well as the intracellular activities of ~300 enzymes and 2,000 transcription factors.12 It is also effective in decreasing oxidative stress and the generation of inflammatory cytokines such as TNF-α and IL-1β.13 Furthermore, the role of zinc in the development and progression of prostate cancer and its widespread antitumor efficacy have been shown in several malignancies.14–17 Intracellular zinc status is associated with prostate carcinogenesis. For example, zinc deficiency contributes to tumor progression and development in cultured HRPC cells,18 whereas increased levels of intracellular zinc decrease cancer cell proliferation and induce apoptosis.14,15 In a large case-control study, a direct association was found between zinc intake and prostate cancer risk.19 The vast majority of epithelial tumors are associated with reduced intratumoral or plasma zinc levels.20 Importantly, zinc deficiency reduces paclitaxel efficacy in cultured prostate cancer cells, whereas increased intracellular zinc concentrations sensitize prostate cancer cells to cytotoxic agents, including paclitaxel, via inhibition of NF-κB activation.21,22
Therefore, zinc supplementation may have growth inhibitory effects against NSCLC cells and may increase docetaxel efficacies. In this study, cellular viability, apoptosis, and cell cycle alterations as well as oxidative stress activities were tested in the NSCLC cell lines, A549, and H1299, when zinc was used alone and in combination with docetaxel.
Materials and methods
Cells
NSCLC cells, A549 (p53 wild-type), and H1299 (lymph node metastasis, p53 null-type) were obtained from the American Type Culture Collection. The A549 cells were cultured in DMEM/Ham’s F12K medium that containing 10% FBS, 1% penicillin/streptomycin, 1% l-glutamine, and 1% amphotericin B. The H1299 cells were cultured in RPMI-1640 medium containing 10% FBS, 1% penicillin/streptomycin, 1% l-glutamine, and 1% amphotericin B at 37°C in a 5% CO2 incubator.
Docetaxel/zinc therapy, cell viability analysis
Cell viability was determined using the CCK-8 (Cell Counting Kit-8; Sigma-Aldrich, St Louis, MO, USA) assay in accordance with the manufacturer’s instructions. Briefly, when the cells reached 80% confluency, the medium was replaced with fresh medium without zinc or docetaxel. Then, the cells were transferred to 96-well plates at a density of 10,000 cells/well. Docetaxel (0.001–100 μM) and zinc sulfate (10–1,000 μM) were added to the cell lines at different concentrations in each of the eight wells. The CCK-8 reagents were added to each well at the indicated time after the treatments. At the end of 24 hours and 48 hours, the absorbance was measured at a test wavelength of 450 nm and a reference wavelength of 650 nm using a microplate reader (Thermo Electron Corporation, Multiskan Ascent). Each experiment was repeated three times. The average inhibitory concentration 50 values (IC50 values) were determined at 24-hour and 48-hour treatment periods. The 24-hour doses were selected due to the lack of significant differences between the 24-hour and 48-hour doses.
Flow cytometry analyses
Apoptosis analysis
The AnnexinV-FITC Apoptosis Detection Kit I (BD Pharmingen) was used. The cells were plated in six-well plates at a density of 50×104 cells/well, and the total volume was brought to 3.2 mL with fresh media. After the initial 72-hour incubation, at which point the cells were ~80% confluent, the cells were treated with the previously established IC50 concentrations of docetaxel, zinc and combinations thereof in fresh medium for an additional 24 hours. The cells were harvested by trypsinization and collected by centrifugation at 1,500 rpm for 5 minutes. Following removal of the supernatant, the cells were resuspended in 400 μL binding buffer and stained with 5 μL annexin-V and 5 μL propidium iodide (PI, 50 μg/mL) at room temperature for 20 minutes in the dark. After adding 400 μL binding buffer to each tube, the tubes were placed in a double-laser flow cytometry device. The percentage apoptosis, necrosis, and viable cells were evaluated by using a flow cytometer. A Beckman CoulterEpicsXL-MCL was used for analysis of the H1299 cells, and a Becton Dickinson (BD) FACSCanto II cytometer was used for analysis of the A549 cells.
Cell cycle analysis
The Cycletest Plus DNA Reagent kit (BD Pharmingen) was used to determine the distribution of cells between cell-cycle phases. The cells were seeded in six-well plates at a density of 50×104 cells/well, and their volume was adjusted to 3.2 mL with media. After an initial incubation for 72 hours, at which point the cells were ~80% confluent, the cells were treated with previously defined IC50 doses of zinc and docetaxel and then incubated for 24 hours. Then, the medium was removed, and 500 μL trypsin was added to the cells, which were transferred into a tube. A volume of 1 mL fresh medium was added to each tube to inactivate the trypsin. The suspension was transferred to 17×100 mm tubes and centrifuged at 1,500 rpm for 5 minutes. The supernatant was removed, and the cells were resuspended by adding 1 mL buffer solution (sucrose-citrate buffer containing dimethyl sulfoxide) to the pellet (wash procedure). The wash was repeated two times more. The following steps were performed on ice. After removal of the buffer, 250 μL solution A (citrate buffer containing trypsin) was added to the pellet in each tube and incubated at room temperature for 10 minutes. Then, 200 μL solution B (citrate buffer containing trypsin inhibitor and RNAse) was added to each tube and incubated at room temperature for 10 minutes. Finally, 200 μL solution C (PI-staining solution) at 4°C was added to each tube and reincubated on ice for 10 minutes. Three hours later, the cells were analyzed using double laser-flow cytometry devices. A Beckman CoulterEpicsXL-MCL was used for analysis of the H1299 cells, and a BD FACSCanto II cytometer was used for analysis of the A549 cells. The cells in G0/G1, S (synthesis), and G2/M (mitosis) phases were evaluated using the ModFitLT3.2 software program.
High-performance liquid chromatography analysis of free malondialdehyde in the cells
The cell lysates were prepared and kept at −20°C before the antioxidant enzyme and malondialdehyde (MDA) analyses. For standard preparation, the MDA standard (T-9889) was purchased from Sigma-Aldrich. A 10 mM stock standard solution was prepared by dissolving the MDA in distilled water, and this was then diluted to working concentrations ranging from 1 μM to 50 μM. For sample preparation, the protein concentrations were measured using a BCA protein assay kit (Pierce BCA Protein Assay Kit 23225; Thermo Fisher Scientific, Waltham, MA, USA). The samples were lyophilized with a Flexi-Dry uP Freeze Dryer (FTS Systems, Stone Ridge, NY) because of the low protein concentrations.
For MDA measurement, 100 μL distilled water was added to a 40 μL volume of all cell samples. Then, 20 μL of 2.8 mmol/L BHT, 40 μL of 8.1% SDS, 600 μL of 8 g/L TBA:200 mL/L acetic acid (1:1) were added, and the pH was adjusted with 2 M NaOH (pH 3.5). All samples were placed in a water bath at 95°C for an hour. Then, the samples were placed on ice. A volume of 200 μL distilled water and 1,000 μL butanol:pyridine (15:1) were added, and all the samples were vortexed for 1 minute. After 3 minutes, the organic phases were transferred to new Eppendorf tubes and centrifuged at 10,000 rpm for 10 minutes. The samples were then placed in an autosampler vial for high-performance liquid chromatography (HPLC) analysis. The sample injection volume was 10 μL. As previously described,23 the analysis was performed using a HPLC set Prominence LC 20 (Shimadzu, Kyoto, Japan) equipped with Sil-20AC autosampler, LC-20AD pump, and RF-10Axl fluorescence detector. A MN Nucleosil 4.6 mm ×250 mm C18 column was used for analysis. Phosphate buffer (0.01 M in 30% methanol, pH 7.0) was used as the mobile phase. The flow rate of the mobile phase was 0.8 mL/min. MDA was detected by fluorescence (excitation 536 nm; emission 549 nm).
Superoxide dismutase and glutathione peroxidase activities of the cells
The superoxide dismutase (SOD) activity of the cells was determined using a RANSOD kit (Randox Labs, Crumlin, UK). Lyophilized cells and standard solutions were used to assay for SOD. The absorbance was measured at 505 nm on a microplate reader (Bio-Tek Instruments, Inc., USA).
The glutathione peroxidase (GPx) activity of the cells was determined using a RANSEL kit (Randox Labs, Crumlin, UK) and a previously described method.24 Briefly, the lyophilized cells and standard solutions were used to assay for GPx. The absorbance was measured at 340 nm on a microplate reader (Bio-Tek Instruments, Inc.). In this method, GPx oxidizes glutathione via cumene hydroperoxidase. In the presence of GR and NADPH, oxidized glutathione is immediately converted to the reduced form, with concomitant oxidation of NADPH to NADP+. A 40-μL sample was added to the primary reagent (200 μL) and cumene (80 μL) into a 96-well plate. The plate was read after 1 minute, then again after 2 minutes for the final absorbance (after reaction). The GPx activity of these samples was calculated according to GPx manual included with the RANSEL kit.
Statistics
The triplicate data were analyzed using the SigmaStat 3.5 (Systat Inc., Santa Cruz, CA, USA) software program. For comparison of the groups, ANOVA was used. Significance was evaluated with Holm–Sidak test, and P<0.05 was accepted as statistically significant.
Results
Growth inhibitory effects of zinc and docetaxel on NSCLC cells
First, we determined the IC50 concentrations for zinc and docetaxel on A549 and H1299 cells using a CC8 kit. To investigate the effects of exogenous zinc on cellular viability, apoptosis, and cell cycle alterations, both cell types were treated with various zinc concentrations (50 μM, 100 μM, and IC50 level). For the combinations, the groups were treated with three different zinc concentrations and the same docetaxel dose (IC50). However, to determine the efficacy of zinc pretreatment on IC50 changes caused by docetaxel, NSCLC cells were treated with 50 μM and 100 μM of zinc. Then, cellular viability and apoptosis were analyzed after 24 hours.
The IC50 values of zinc on A549 and H1299 cells were 287.1 μM and 458.2 μM, respectively, whereas the docetaxel concentrations were 20.44 μM and 33.15 μM, respectively (Figure 1). The docetaxel IC50 values for A549 cells were reduced to 8.16 μM and 6.6 μM levels after exposure to 50 μM and 100 μM of zinc, respectively, whereas these values were reduced to 12.16 μM and 14.69 μM in H1299 cells. Zinc significantly reduced the cellular viability of A549 cells in a dose-dependent fashion when combined with docetaxel and used alone (P<0.001). There was significant suppression of viability in combination groups. However, zinc supplementation did not modify H1299 cell viability at all concentrations, although significant suppression was observed for the combination groups. For apoptotic induction, zinc significantly increased the effects of docetaxel on both cell lines (Figure 2).
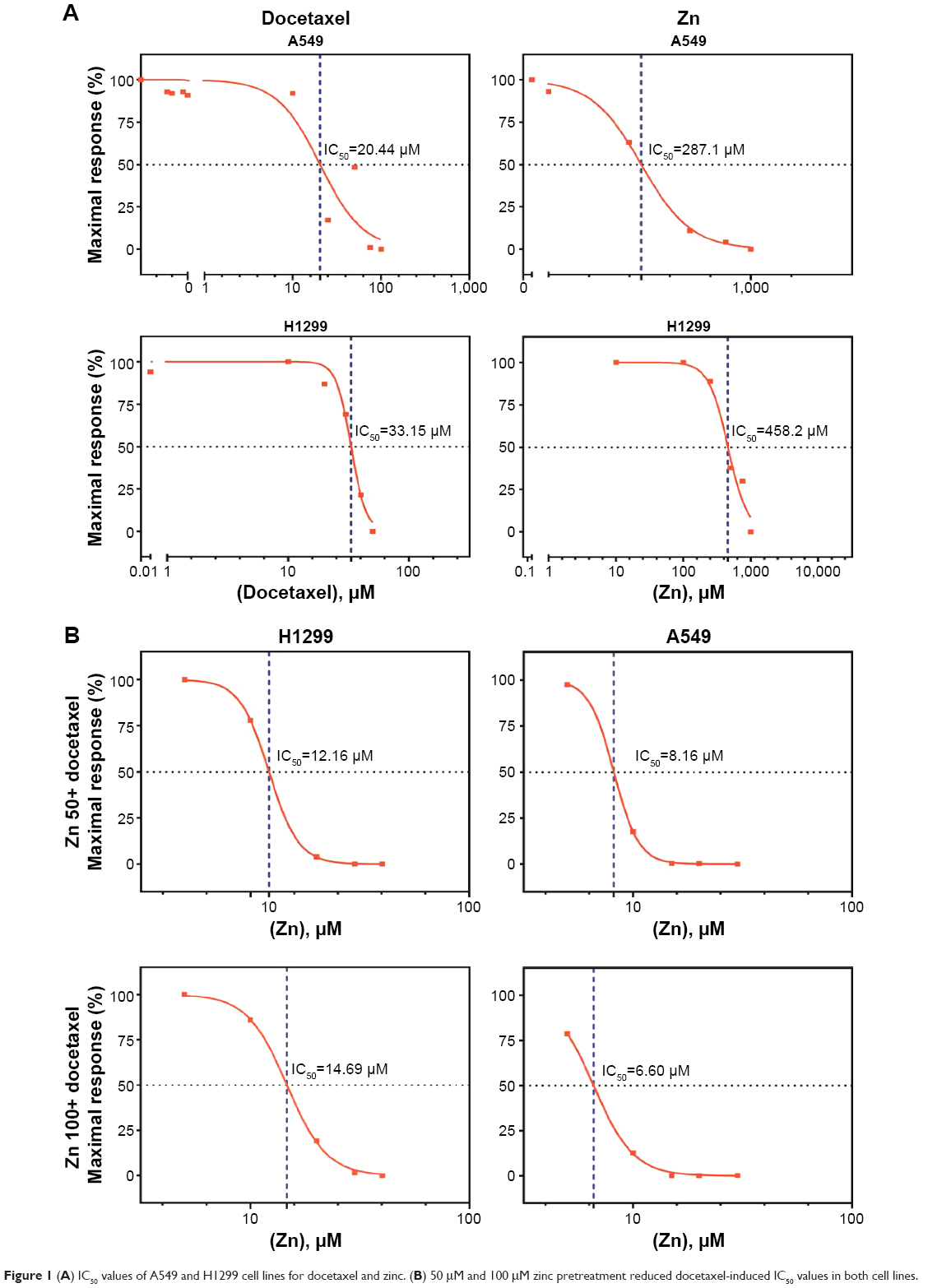 | Figure 1 (A) IC50 values of A549 and H1299 cell lines for docetaxel and zinc. (B) 50 μM and 100 μM zinc pretreatment reduced docetaxel-induced IC50 values in both cell lines. |
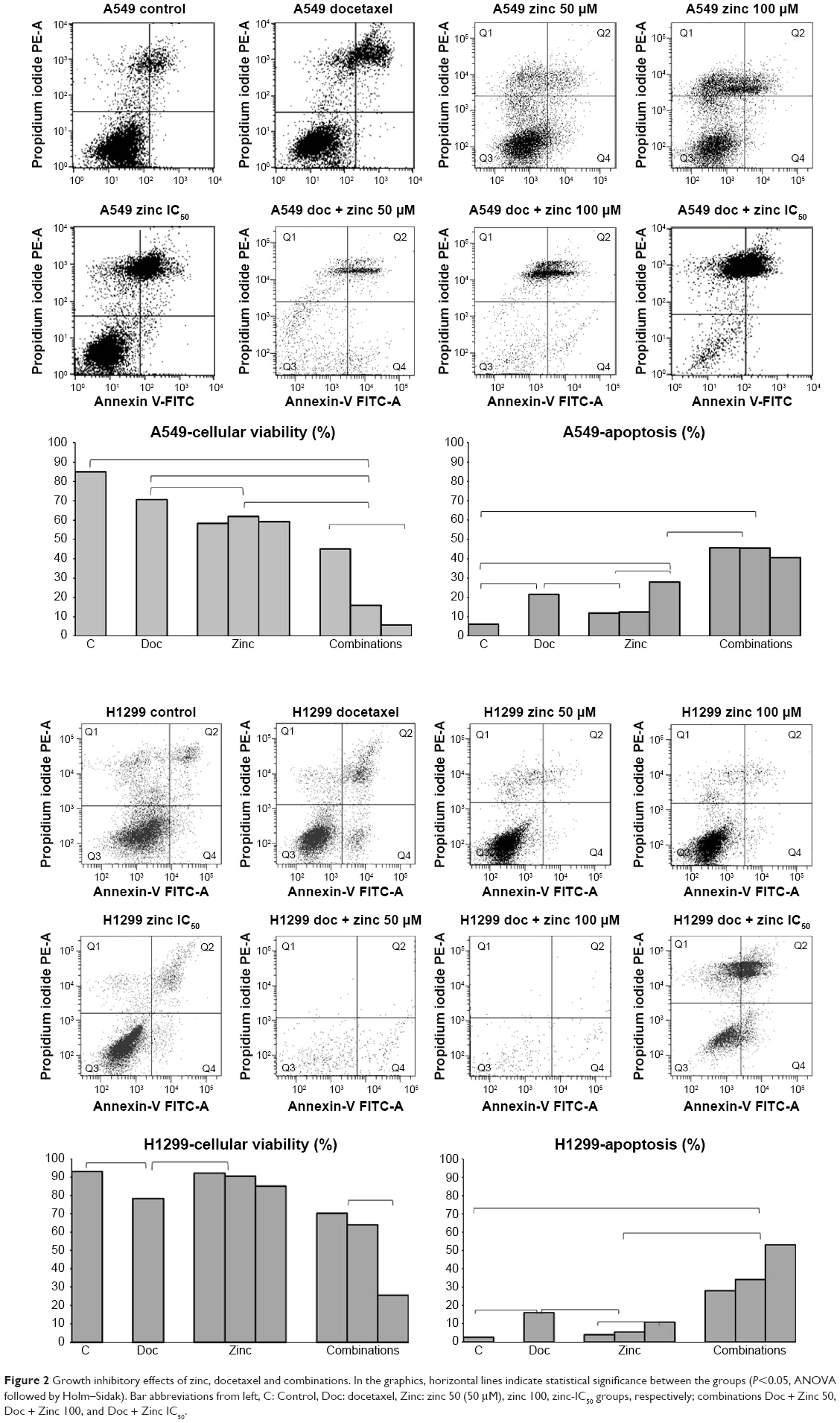 | Figure 2 Growth inhibitory effects of zinc, docetaxel and combinations. In the graphics, horizontal lines indicate statistical significance between the groups (P<0.05, ANOVA followed by Holm–Sidak). Bar abbreviations from left, C: Control, Doc: docetaxel, Zinc: zinc 50 (50 μM), zinc 100, zinc-IC50 groups, respectively; combinations Doc + Zinc 50, Doc + Zinc 100, and Doc + Zinc IC50. |
Cell cycle analysis
As shown in Figure 3, the increase of the G0/G1 phase cell population was accompanied by a concomitant S-phase decrease in A549 cells treated with zinc and combinations in a dose-dependent manner. However, zinc and combinations reduced the G0/G1 population and increased S-phase accumulation in H1299 cells. As expected, G2/M phase accumulation significantly increased after docetaxel treatment in both cell lines. H1299 cells showed significantly increased apoptotic/necrotic cell population in the combination groups.
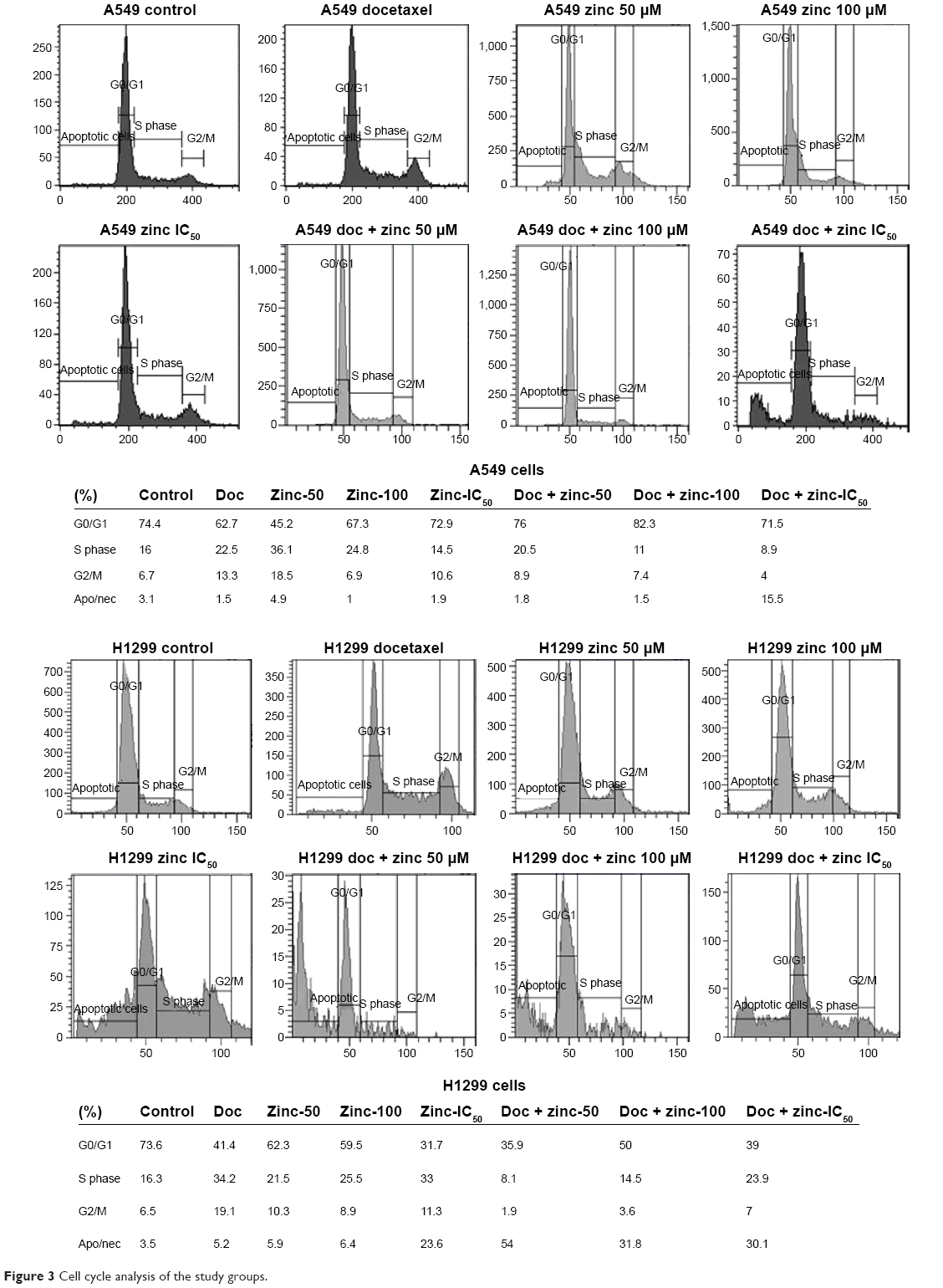 | Figure 3 Cell cycle analysis of the study groups. |
Antioxidant enzyme status and MDA alterations
To assess the importance of oxidative stress, the SOD activity, GPx activity, and MDA levels were evaluated in NSCLC cells. In both cell lines, the SOD values significantly increased all treatment groups compared to the control, but the highest values were observed in the cells treated with 100 μM zinc. GPx was not elevated when both the A549 and H1299 cells were treated with 50 μM and 100 μM doses, but were slightly increased in the docetaxel and combination groups (Table 1). The maximum MDA response was observed when the A549 cells were exposed to 50 μM of zinc and combinations, except the IC50 doses, whereas the maximum MDA response occurred when the H1299 cells were exposed to 100 μM of zinc and docetaxel (P<0.001, ANOVA followed by Holm–Sidak test).
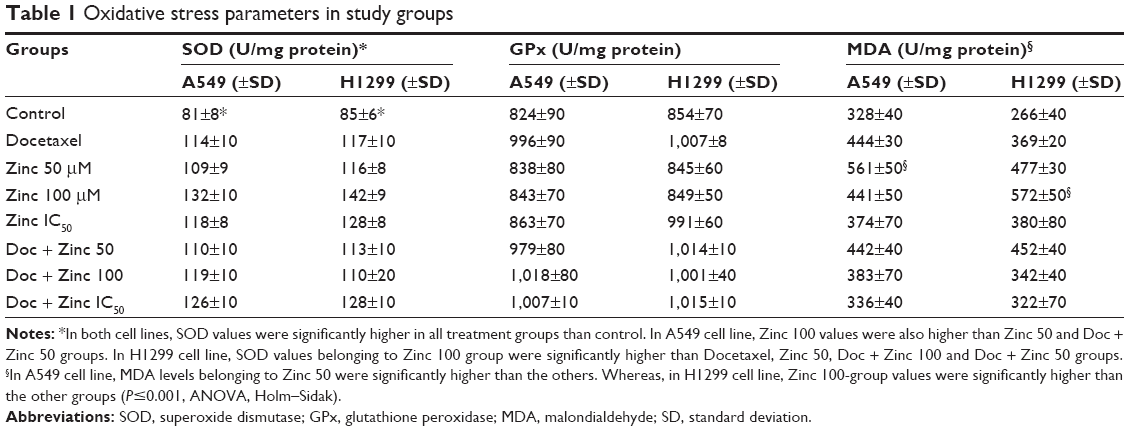 | Table 1 Oxidative stress parameters in study groups |
Discussion
Zinc is reported to induce apoptosis and cytotoxicity in different tumor types including prostate, ovarian, hepatoma, pancreatic, and breast cancers,14–17,25,26 but it is less studied in lung cancers. Here, we found that zinc exhibited growth inhibitory and apoptotic effects in a dose-dependent manner, up to the IC50 concentrations for cultured lung cancer cells. Importantly, these effects were significantly increased when combined with docetaxel. Apoptotic cell contents were higher for the combination than individual treatments and control.
The exact mechanisms underlying this synergism and zinc-induced apoptosis are unclear, but zinc homeostasis and p53 functions appear to be key factors for subsequent physiological processes including DNA repair, cell cycle regulation, and response to oxidative stress.27 Both exposures to excessive zinc and zinc depletion activate the apoptosis, and also, the zinc-induced apoptosis are commonly associated with the oxidative stress. For example, intracellular zinc deficiency can enhance DNA damage due to oxidative stress and also blocks critical cellular signals that drive DNA repair and apoptosis,27 such that, zinc deficiency results in increased sensitivity to oxidative stress, increased DNA damage, and accelerated carcinogenesis.28 The DNA repair functions of p53 are commonly lost, and redox sensitive transcription factors, such as AP-1 and NF-κB show reduced nuclear binding as a result of low cellular zinc status.27 The zinc exposure causes intracellular acidosis, increased the reactive oxygen species (ROS) generation in several tumor cells,29,30 and stimulates the caspase-dependent and -independent apoptosis.30,31 Moreover, the zinc-induced ROS generation has been shown in the presence of functionally active p53 in breast cancer cells.29,30 By contrast, reactivation of mutant p53 increases the sensitivity of chemoresistant cancer cells to antineoplastic agents with zinc supplementation, which was demonstrated for glioblastoma and colorectal cancer cells.32 Zinc-related apoptosis is associated with decreased Bcl-2 and survivin expression in prostate cancer cells, and other studies found that survivin inhibition with deguelin results in enhanced efficacy of docetaxel in breast cancer cells.15,33 Therefore, based on these data, we hypothesize that the addition of zinc into docetaxel regimens is a reasonable option to increase docetaxel-induced apoptosis and cytotoxicity in NSCLC cells.
In the present study, NSCLC-derived H1299 (p53-null) and A549 (p53-wild-type) cells were used. The IC50 doses of docetaxel and zinc were higher for the p53-null H1299 cells than A549 cells (Figure 1). It is well known that the complete loss of p53 or the expressions of mutant p53 variants are commonly seen in NSCLC with 40–70% frequencies and are associated with increased resistance to chemotherapy and radiotherapy.11,34,35 Functional p53 status may influences docetaxel and zinc-induced cytotoxicity.11 For example, Liu et al36 demonstrated that docetaxel induces apoptotic cell death in C4-2 and LNCaP prostate cancer cell express wild-type p53 more than DU145 (mutant p53) and PC-3 (p53-null) cells. They also shown knockdown of p53 protein in C4-2 cells increases docetaxel resistance. The role of p53 on zinc-induced apoptosis has been shown in some similar studies.29,37 In our study, zinc showed cytotoxicity in p53-wild lung cancer cells but not in null cells at different supraphysiological concentrations. These results suggest that zinc-induced cytotoxicity and/or apoptotic induction are associated with the availability of functionally active p53 in NSCLC cells.
In our study, we observed increased MDA levels, which are an indirect indicator of oxidative stress and ROS production, using higher zinc concentrations (50 μM and 100 μM but not the IC50) in p53-wild-type A549 cells, whereas H1299 cells had higher MDA levels at 100 μM zinc concentrations. Based on our data, it is suggested that supraphysiological zinc supplementation induces ROS generation and apoptosis in NSCLC cells through p53 activation. In the presence of altered intracellular zinc concentrations (low or high), ROS stimulates p53 activation and/or p53 itself to stimulate ROS production, and finally, it may cause cell death. Previous studies demonstrated that when cells are exposed to several zinc chelators, mutant p53 forms start to accumulate, and these forms lose their DNA binding capacity. Finally, redox-active compounds cause DNA-strand breaks, whereas removal of the zinc chelators from the medium returns to cell “wild-type p53” form.38 In addition, exposure of MCF7 (p53-wild-type) cells to 10–25 μM zinc causes an increase in the expression of Bax, proapoptotic Bcl-2 and, PIG3 (p53-induced gene product) and also generation of ROS.
The synergistic effects of zinc on docetaxel cytotoxicity may occur by inducing ROS generation and increasing oxidative stress. The combination of zinc and docetaxel induced apoptosis and cytotoxicity as well as MDA levels. Interestingly, zinc also reduced the IC50 values of docetaxel and this may related to intracellular action of ROS. Similar to other chemotherapeutic agents, taxanes induce ROS production, and high levels of ROS participate in paclitaxel cytotoxicity.39 For example, exposure of A549 cells to paclitaxel resulted in an intracellular increase of O2− and H2O2 levels. Then, the addition of N-acetylcysteine (NAC) and glutathione, which are H2O2 scavengers, induced a fourfold increase in the paclitaxel IC50 level.40 We also examined the antioxidant enzymes SOD and GPx in cell lysates and found significant increases in all treatment modalities compared to the control, with the highest SOD values in the combination group. These results may be related to the higher zinc and docetaxel concentrations. Zinc is an essential component of CuZnSOD, which is the first-line defense enzyme to avoid the harmful effects of ROS.27,41
In our study, we selected three different zinc concentrations, and zinc concentration was slightly supraphysiological (50 μM) along with higher concentrations (100 μM and IC50). Because, the zinc treatment at ~15 μM concentration is accepted as physiological level.42 Previous studies showed that zinc-induced apoptosis occurs from 50 μM and higher, but zinc-induced ROS generation is observed at 100 μM doses.14,17,25,43,44 Although, we have not counted intracellular free zinc concentrations after the drug and zinc exposures, in a published study, maximum intracellular zinc concentrations have been obtained at 50 μM ZnSO4 treatment, and finally decreased to initial level when the concentration of ZnSO4 exceeded 100 μM.17 In addition, the cell growth inhibition and apoptosis were evaluated 24 hours after the treatment. Apoptotic changes due to zinc treatments (100 ng/mL and 1,000 ng/mL) were remarkably seen in prostate cancer cells in an in vitro study.45 It has also been shown that, the zinc ions translocated into mitochondria 24 hours after the 150 μM zinc sulfate treatment and at the beginning of the 18 hours of treatment, cytochrome c started to be seen in the cytoplasm and reached to the peak level in approximately 36 hours of treatment.44
The cell cycle analysis revealed G0/G1 accumulation in A549 cell after zinc treatment in a dose-dependent manner. This results confirmed p53 activation in A549 cells. It is well known that activated p53 causes G1 arrest by inducing p21, followed by cyclin inhibition to induce DNA repair before cellular division,46 and the cell cycle analysis showed increased S phase and significantly higher apoptotic/necrotic cell death (~50%) in p53-deficient NSCLC cells.
The zinc induced-apoptosis and the synergism with docetaxel were speculated through ROS generation and p53 activation here. However, one of the major limitations of the present study was the lack of tests that showed the p53 functions or the p53-related protein alterations, as well as the apoptotic protein expressions after the docetaxel and the zinc applications. Another important limitation of this study was the absence of the counting of intracellular zinc and ROS content after the compound exposures.
Conclusion
In conclusion, the exposure to exogenous zinc results in remarkable growth inhibition, increased apoptosis in the NSCLC cells at 50 μM and 100 μM concentrations and zinc supplementation potentially sensitizes the NSCLC cells to the docetaxel treatment.
Acknowledgment
This work supported by Izmir Medical and Scientific Research Activities Support Association (Turkey).
Disclosure
The authors report no conflicts of interest in this work.
References
Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. | ||
Govindan R, Page N, Morgensztern D, et al. Changing epidemiology of small-cell lung cancer in the United States over the last 30 years: analysis of the surveillance, epidemiologic, and end results database. J Clin Oncol. 2006;24:4539–4544. | ||
Siegel RL, Miller KD, Jemal A. Cancer statistics 2015. CA Cancer J Clin. 2015;65:5–29. | ||
Socinski MA. Update on taxanes in the first-line treatment of advanced non-small-cell lung cancer. Curr Oncol. 2014;21:e691–e703. | ||
Chu Q, Vincent M, Logan D, Mackay JA, Evans WK; Lung Cancer Disease Site Group of Cancer Care Ontario’s Program in Evidence-based Care. Taxanes as first-line therapy for advanced non-small cell lung cancer: a systematic review and practice guideline. Lung Cancer. 2005;50:355–374. | ||
McGrogan BT, Gilmartin B, Carney DN, McCann A. Taxanes, microtubules and chemoresistant breast cancer. Biochim Biophys Acta. 2008;1785:96–132. | ||
Breen L, Murphy L, Keenan J, Clynes M. Development of taxane resistance in a panel of human lung cancer cell lines. Toxicol In Vitro. 2008;22:1234–1241. | ||
Chiu LY, Ko JL, Lee YJ, Yang TY, Tee YT, Sheu GT. L-type calcium channel blockers reverse docetaxel and vincristine-induced multidrug resistance independent of ABCB1 expression in human lung cancer cell lines. Toxicol Lett. 2010;192:408–418. | ||
Zhang Q-C, Jiang S-J, Zhang S, Ma X-B. Histone deacetylase inhibitör trichostatin A enhances anti-tumor effects of docetaxel or erlotinib in A 549 cell line. Asian Pac J Cancer Prev. 2012;13:3471–3476. | ||
Ping S-Y, Hour T-C, Lin S-R, Yu D-S. Taxol synergizes with antioxidants in inhibiting hormonal refractory prostate cancer cell growth. Urol Oncol. 2010;28:170–179. | ||
Li G, Zhao J, Peng X, Liang J, Deng X, Chen Y. Radiation/paclitaxel treatment of p53-abnormal non-small cell lung cancer xenograft tumor and associated mechanism. Cancer Biother Radiopharm. 2012;27:227–233. | ||
Prasad AS, Mukhtar H, Beck FW, et al. Dietery zinc and prostate cancer in the TRAMP Mouse model. J Med Food. 2010;13:70–76. | ||
Prasad AS, Beck FWJ, Snell DC, Kucuk O. Zinc in cancer prevention. Nutr Cancer. 2009;61:879–887. | ||
Yan M, Hardin K, Ho E. Differential response to zinc-induced apoptosis in benign prostate hyperplasia and prostate cancer cells. J Nutr Biochem. 2010;21:687–694. | ||
Ku JH, Seo SY, Kwak C, Kim HH. The role of survivin and Blc-2 in zinc induced apoptosis in prostate cancer cells. Urol Oncol. 2012;30:562–568. | ||
Franklin RB, Costello LC. Zinc as an anti-tumor agent in prostate cancer and in other cancers. Arch Biochem Biophys. 2007;463:211–217. | ||
Yuan N, Wang YH, Li KJ, et al. Effects of exogenous zinc on the cellular zinc distribution and cell cycle of A549 cells. Biosci Biotechnol Biochem. 2012;76(11):2014–2020. | ||
Golovine K, Uzzo RG, Makhov P, Crispen PL, Kunkle D, Kolenko VM. Depletion of intracellular zinc increases expression of tumorigenic cytokines VEGF, IL-6 and IL-8 in prostate cancer cells via NF-kB-dependent pathway. Prostate. 2008;68:1443–1449. | ||
Gallus S, Foschi R, Negri E, et al. Dietary zinc and prostate cancer risk: a case-control study from Italy. Eur Urol. 2007;52:1052–1057. | ||
Gumulec J, Masarik M, Adam V, Eckschlager T, Provaznik I, Kizek R. Serum and tissue zinc in epithelial malignancies: a meta-analysis. PLoS One. 2015;9:1–11. | ||
Killile AN, Downing Killilea DW. Zinc deficiency reduces paclitaxel efficacy in LNCaP prostate cancer cells. Cancer Lett. 2007;258:25870–25879. | ||
Uzzo RG, Leavis P, Hatch W, et al. Zinc inhibits nuclear factor-kB activation and sensitizes prostate cancer cells to cytotoxic agents. Clin Cancer Res. 2002;8:3579–3583. | ||
Hong YL, Yeh SL, Chang CY, Hu ML. Total plasma malondialdehyde levels in 16 Taiwanese college students determined by various thiobarbituric acid tests and an improved high-performance liquid chromatography-based method. Clin Biochem. 2000;33:619–624. | ||
Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med. 1967;70:158–169. | ||
Wang YH, Li KJ, Mao L, et al. Effects of exogenous zinc on cell cycle, apoptosis and viability of MDAMB231, HepG2 and 293 T cells. Biol Trace Elem Res. 2013;154:418–426. | ||
Costello LC, Franklin RB. Cytotoxic/tumor suppressor role of zinc fort he treatment of cancer: an enigma and an opportunity. Expert Rev Anticancer Ther. 2012;12:121–128. | ||
Ho E. Zinc deficiency, DNA damage and cancer risk. J Nutr Biochem. 2004;15:572–578. | ||
Song Y, Leonard SW, Traber MG, Ho E. Zinc deficiency affects DNA damage, oxidative stress, antioxidant defences and DNA repair in rats. J Nutr. 2009;139:1626–1631. | ||
Ostrakhovitch EA, Cherian MG. Role of p53 and reactive oxygen species in apoptotic response to copper and zinc in epithelial breast cancer cells. Apoptosis. 2005;10:111–121. | ||
Donadelli M, Dalla Pozza E, Scupoli MT, Costanzo C, Scarpa A, Palmieri M. Intracellular zinc increase inhibits p53 (−/−) pancreatic adenocarcinoma cell growth by ROS/AIF-mediated apoptosis. Biochim Biophys Acta. 2009;1793:273–280. | ||
Jayaraman AK, Jayaraman S. Increased level of exogenous zinc induces cytotoxicity and upregulates the expression of the ZnT-1 zinc transporter gene in pancreatic cancer cells. J Nutr Biochem. 2011;22:79–88. | ||
Cirone M, Garufi A, Di Renzo L, Granato M, Faggioni A, D’Orazi G. Zinc supplementation is required for cytotoxic and immunogenic effects of chemotherapy in chemoresistant p53-functionally deficient cells. Oncoimmunology. 2013;2:e26198. | ||
Ghanbari P, Mohseni M, Tabasinezhad M, et al. Inhibition of survivin restores to sensitivity of breast cancer cells to docetaxel and vinblastin. Appl Biochem Biotechnol. 2014;174:667–681. | ||
Victorsson K, DePetris L, Lewensohn R. The role of p53 treatment responses of lung cancer. Biochem Biophys Res Commun. 2005;331:868–880. | ||
Brown CJ, Lain S, Verma CS, Fersht AR, Lane DP. Awakening guardian angels: drugging the p53 pathway. Nat Rev Cancer. 2009;12:862–873. | ||
Liu C, Zhu Y, Lou W, et al. Functional p53 determines docetaxel sensitivity in prostate cancer cells. Prostate. 2013;73:418–427. | ||
Margalit O, Simon AJ, Yakubov E, et al. Zinc supplementation augments in vivo antitumor effect of chemotherapy by restoring p53 function. Int J Cancer. 2012;131:E562–E568. | ||
Hainaut P, Mann K. Zinc binding and redox control of p53 structure and function. Antioxid Redox Signal. 2001;3:611–623. | ||
Lyle PA, Mitsopoulos P, Suntres ZE. N-Acetylcysteine modulates the cytotoxic effects of paclitaxel. Chemotherapy. 2011;57:298–304. | ||
Alexandre J, Batteux F, Nicco C, et al. Accumulation of hydrogen peroxide is an early and crucial step for paclitaxel-induced cancer cell death both in vitro and in vivo. Int J Cancer. 2006;119:41–48. | ||
Chasapis CT, Loutsidou AC, Spiliopoulou CA, Stefanidou ME. Zinc and human health: an update. Arch Toxicol. 2012;86:521–534. | ||
Lin SF, Wei H, Maeder D, Franklin RB, Feng P. Profiling of zinc-altered gene expression in human prostate normal vs cancer cells: a time course study. J Nutr Biochem. 2009;20:1000–1012. | ||
Sankavaram K, Chong L, Bruno RS, Freake HC. Zinc status alters growth and oxidative stress responses in rat hepatoma cells. Nutr Cancer. 2014;66:104–116. | ||
Rudolf E, Rudolf K, Cervinka M. Zinc induced apoptosis in HEP-2 cancer cells: the role of oxidative stress and mitochondria. Biofactors. 2005;23:107–120. | ||
Liang J-Y, Liu Y-Y, Zou J, Franklin RB, Costello LC, Feng P. Inhibitory effect of zinc on human prostatic carcinoma cell growth. Prostate. 1999;40:200–207. | ||
Vermeulen K, Berneman ZN, Van Bockstaele DR. Cell cycle and apoptosis. Cell Prolif. 2003;36:165–175. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.