")
Back to Journals » Drug Design, Development and Therapy » Volume 9
The pan-inhibitor of Aurora kinases danusertib induces apoptosis and autophagy and suppresses epithelial-to-mesenchymal transition in human breast cancer cells
Authors Li J, Yang Y, Liu Q, Zhou Z, Pan S, He Z, Zhang X, Yang T, Pan S, Duan W, He S, Chen X, Qiu J, Zhou S
Received 16 September 2014
Accepted for publication 22 October 2014
Published 17 February 2015 Volume 2015:9 Pages 1027—1062
DOI https://doi.org/10.2147/DDDT.S74412
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Min Li
Jin-Ping Li,1,2 Yin-Xue Yang,3 Qi-Lun Liu,1 Zhi-Wei Zhou,2,4 Shu-Ting Pan,2,5 Zhi-Xu He,4 Xueji Zhang,6 Tianxin Yang,7 Si-Yuan Pan,8 Wei Duan,9 Shu-Ming He,10 Xiao-Wu Chen,11 Jia-Xuan Qiu,5 Shu-Feng Zhou2,4
1Department of Surgical Oncology, General Hospital of Ningxia Medical University, Yinchuan, Ningxia, People’s Republic of China; 2Department of Pharmaceutical Sciences, College of Pharmacy, University of South Florida, Tampa, FL, USA; 3Department of Colorectal Surgery, General Hospital of Ningxia Medical University, Yinchuan, Ningxia, People’s Republic of China; 4Guizhou Provincial Key Laboratory for Regenerative Medicine, Stem Cell and Tissue Engineering Research Center and Sino-US Joint Laboratory for Medical Sciences, Guiyang Medical University, Guiyang, Guizhou, People’s Republic of China; 5Department of Oral and Maxillofacial Surgery, The First Affiliated Hospital of Nanchang University, Nanchang, Jiangxi, People’s Republic of China; 6Research Center for Bioengineering and Sensing Technology, University of Science and Technology Beijing, Beijing, People’s Republic of China; 7Department of Internal Medicine, University of Utah and Salt Lake Veterans Affairs Medical Center, Salt Lake City, UT, USA; 8Department of Pharmacology, School of Chinese Materia Medica, Beijing University of Chinese Medicine, Beijing, People’s Republic of China; 9School of Medicine, Deakin University, Waurn Ponds, VIC, Australia; 10Department of Obstetrics and Gynecology, Xiaolan People’s Hospital affiliated to Southern Medical University, Zhongshan, Guangdong, People’s Republic of China; 11Department of General Surgery, The First People’s Hospital of Shunde affiliated to Southern Medical University, Shunde, Foshan, Guangdong, People’s Republic of China
Abstract: Danusertib (Danu) is a pan-inhibitor of Aurora kinases and a third-generation breakpoint cluster region-Abelson murine leukemia viral oncogene homolog 1 (Bcr-Abl) tyrosine kinase inhibitor, but its antitumor effect and underlying mechanisms in the treatment of human breast cancer remain elusive. This study aimed to investigate the effects of Danu on the growth, apoptosis, autophagy, and epithelial-to-mesenchymal transition (EMT) and the molecular mechanisms in human breast cancer MCF7 and MDA-MB-231 cells. The results demonstrated that Danu remarkably inhibited cell proliferation, induced apoptosis and autophagy, and suppressed EMT in both breast cancer cell lines. Danu arrested MCF7 and MDA-MB-231 cells in G2/M phase, accompanied by the downregulation of cyclin-dependent kinase 1 and cyclin B1 and upregulation of p21 Waf1/Cip1, p27 Kip1, and p53. Danu significantly decreased the expression of B-cell lymphoma-extra-large (Bcl-xl) and B-cell lymphoma 2 (Bcl-2), but increased the expression of Bcl-2-associated X protein (Bax) and p53-upregulated modulator of apoptosis (PUMA), and promoted the cleavage of caspases 3 and 9. Furthermore, Danu significantly increased the expression levels of the membrane-bound microtubule-associated protein 1A/1B-light chain 3 (LC3-II) and beclin 1 in breast cancer cells, two markers for autophagy. Danu induced the activation of p38 mitogen-activated protein kinase (MAPK) and extracellular signal-regulated kinases 1 and 2 (Erk1/2) and inhibited the activation of protein kinase B (Akt)/mammalian target of rapamycin (mTOR) signaling pathways in breast cancer cells. Treatment with wortmannin (a phosphatidylinositol 3-kinase inhibitor) markedly inhibited Danu-induced activation of p38 MAPK and conversion of cytosolic LC3-I to membrane-bound LC3-II. Pharmacological inhibition and small interfering RNA-mediated knockdown of p38 MAPK suppressed Akt activation, resulting in LC3-II accumulation and enhanced autophagy. Pharmacological inhibition and small interfering RNA-mediated knockdown of Erk1/2 also remarkably increased the level of LC3-II in MCF7 cells. Moreover, Danu inhibited EMT in both MCF7 and MDA-MB-231 cells with upregulated E-cadherin and zona occludens protein 1 (ZO-1) but downregulated N-cadherin, zinc finger E-box-binding homeobox 1 (TCF8/ZEB1), snail, slug, vimentin, and ß-catenin. Notably, Danu showed lower cytotoxicity toward normal breast epithelial MCF10A cells. These findings indicate that Danu promotes cellular apoptosis and autophagy but inhibits EMT in human breast cancer cells via modulation of p38 MAPK/Erk1/2/Akt/mTOR signaling pathways. Danu may represent a promising anticancer agent for breast cancer treatment. More studies are warranted to fully delineate the underlying mechanisms, efficacy, and safety of Danu in breast cancer therapy.
Keywords: Danusertib, breast cancer, cell cycle, apoptosis, autophagy, EMT
Introduction
Breast cancer is a major health issue worldwide and has been considered as the second most common cancer and the most common cause of cancer-related death in women worldwide, with 521,000 deaths in 2012.1,2 It is placing a heavy burden on individuals, families, and society.1,2 In the US, breast cancer is the most common cancer and the second most common cause of cancer-related death among women.3 There were 220,097 women and 2,078 men who were diagnosed with breast cancer, and there were 40,931 women and 443 men who died from breast cancer in 2011.4 Similarly, breast cancer is the most common and the leading cause of cancer-related death in women in the People’s Republic of China. In 2008, there were about 169,000 new cases of breast cancer, and 44,900 deaths resulted from breast cancer among women in the People’s Republic of China.2 The primary modalities for breast cancer therapy include surgery, radiation therapy, chemotherapy, hormone therapy, immunotherapy, and targeted therapy, which are applied alone or more often in combination.5 These breast cancer therapies aim to remove tumor tissue/burden or induce cancer cell death involving necrosis, senescence, autophagy, and mitotic catastrophe.6 However, currently used chemotherapy for breast cancer is often accompanied with side effects and drug resistance, resulting in therapeutic failure in breast cancer patients. Thus, there is an urgent need to identify and develop new agents with reduced side effects and improved efficacy for breast cancer treatment.
Mitosis is a critical step in the proliferation of cells involving several checkpoint systems controlling key steps of the process. Abnormalities in mitosis have been implicated in the initiation, development, and progression of cancer. Drugs that are capable of managing the abnormal progression of mitosis are currently being tested in cancer therapy.7 Aurora kinases, a class of centrosomal serine/threonine kinase involved in cell division, play a pivotal role in chromosome segregation during the process of cell division through the formation of bipolar spindles. There are three types of Aurora kinases in mammals, namely Aurora A, B, and C (AURKA/B/C). They are structurally and functionally different, and have different intracellular locations. The human AURKA is located on chromosome segment 20q13, which is often amplified and/or overexpressed in several human epithelial malignancies, including colon carcinoma, lymphoma, gastrointestinal adenocarcinomas, breast cancer, and bladder cancer.8–12 AURKB, known as the chromosomal passenger protein, is essential for accurate chromosome segregation and cytokinesis.13 Aberrant expression of AURKA and AURKB has been implicated in the initiation, development, and progression of a wide range of malignancies,14–17 which renders AURKA and AURKB to be potential therapeutic targets for cancer treatment through inhibiting their activities and/or expression.
Currently, there are a number of Aurora kinase inhibitors in different stages of preclinical and clinical development. Danusertib (Danu) is a pan-inhibitor of Aurora kinases and a third-generation breakpoint cluster region-Abelson murine leukemia viral oncogene homolog 1 (Bcr-Abl) tyrosine kinase inhibitor. It potently inhibits the activities of AURKA/B/C, with the half maximal inhibitory concentration (IC50) value of 13, 79, and 61 nM, respectively.18 Recently, Danu has been studied in Phase I and II trials, showing great therapeutic potential in the treatment of a wide range of cancers, including both advanced solid tumors and leukemias.18,19 The clinical anticancer activity of Danu has largely been consistent with its cytostatic effects. The best tumor response was stable disease which was observed in about 23.7% of patients with advanced or metastatic solid tumors.20 However, the effect and underlying mechanisms of Danu in breast cancer treatment have not yet been determined. In the present study, we investigated the effects of Danu on the proliferation, cell cycle distribution, apoptosis, autophagy, and epithelial-to-mesenchymal transition (EMT) in breast cancer cells, and explored the possible mechanisms responsible for the anticancer effect of Danu in breast cancer cells.
Materials and methods
Chemicals and reagents
Danu, previously known as PHA-739358 (N-[5-[(2R)-2-methoxy-2-phenylacetyl]-4,6-dihydro-1H-pyrrolo[3,4-c]pyrazol-3-yl]-4-(4-methylpiperazin-1-yl)benzamide) (Figure 1A), RNase A, propidium iodide (PI), thiazolyl blue tetrazolium bromide (MTT), Dulbecco’s phosphate-buffered saline (D-PBS), protease inhibitor and phosphatase inhibitor cocktails, 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid (HEPES), ethylenediaminetetraacetic acid (EDTA), and fetal bovine serum (FBS) were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). Dulbecco’s Modified Eagle’s Medium (DMEM)/F12 (1:1) and Roswell Park Memorial Institute (RPMI) 1640 medium were purchased from Corning Life Sciences Inc. (Manassas, VA, USA). Lipofectamine 2000, 6-diamidino-2-phenylindole (DAPI), and Opti-minimal essential medium (MEM) were bought from Thermo Fisher Scientific Inc. (Waltham, MA, USA). The annexin V:phycoerythrin (PE) apoptosis kit was purchased from BD Biosciences Inc. (San Jose, CA, USA). Cyto-ID® Autophagy detection kit was obtained from Enzo Life Sciences Inc. (Farmingdale, NY, USA). The polyvinylidene difluoride (PVDF) membrane was bought from EMD Millipore Inc. (Billerica, MA, USA). Western blotting substrate and Pierce™ bicinchoninic acid (BCA) protein assay kit were bought from Thermo Scientific Inc. (Hudson, NH, USA). Bafilomycin A1 (a known inhibitor of the late phase of autophagy, which prevents the maturation of autophagic vacuoles by inhibiting fusion between autophagosomes and lysosomes), wortmannin (WM, an irreversible and selective phosphatidylinositol 3-kinase [PI3K] inhibitor and a blocker of autophagosome formation), and SB202190 (an autophagy inducer and specific p38 mitogen-activated protein kinase [MAPK] inhibitor) were purchased from Invivogen Inc. (San Diego, CA, USA). U0126 (a specific extracellular signal-regulated kinases 1 and 2 [Erk1/2] inhibitor) was purchased from Santa Cruz Biotechnology Inc. (Dallas, TX, USA). The negative control small interfering RNA (siRNA), p38 MAPK siRNA, and p44/42 MAPK (Erk1/2) siRNA, and primary antibodies against human p53, p21 Wafl/Cipl, p27 Kipl, cyclin B1, cyclin dependent kinase 1 (CDK1)/cell division cycle protein 2 (CDC2), CDK2, the p53-upregulated modulator of apoptosis (PUMA), B-cell lymphoma 2 (Bcl-2), Bcl-2-like protein 4/Bcl-2-associated X protein (Bax), B-cell lymphoma extra-large (Bcl-xl), cleaved caspase 3, cleaved caspase 9, microtubule-associated protein 1A/1B-light chain 3 (LC3-I), LC3-II, p38 MAPK, phosphorylated (p) p38 MAPK at Thr180/Tyr182, Erk1/2, p-Erk1/2 at Thr202/Tyr204, protein kinase B (Akt), p-Akt at Ser473, mammalian target of rapamycin (mTOR), p-mTOR at Ser2448, beclin 1, and EMT antibody sampler kit (No 9782) were all purchased from Cell Signaling Technology Inc. (Beverly, MA, USA). The EMT antibody sampler kit contains primary antibodies to N-cadherin, E-cadherin, zona occludens protein 1 (ZO-1), vimentin, slug, snail, zinc finger E-box-binding homeobox 1 (TCF8/ZEB1), and β-catenin. The antibody against human β-actin was obtained from Santa Cruz Biotechnology Inc.
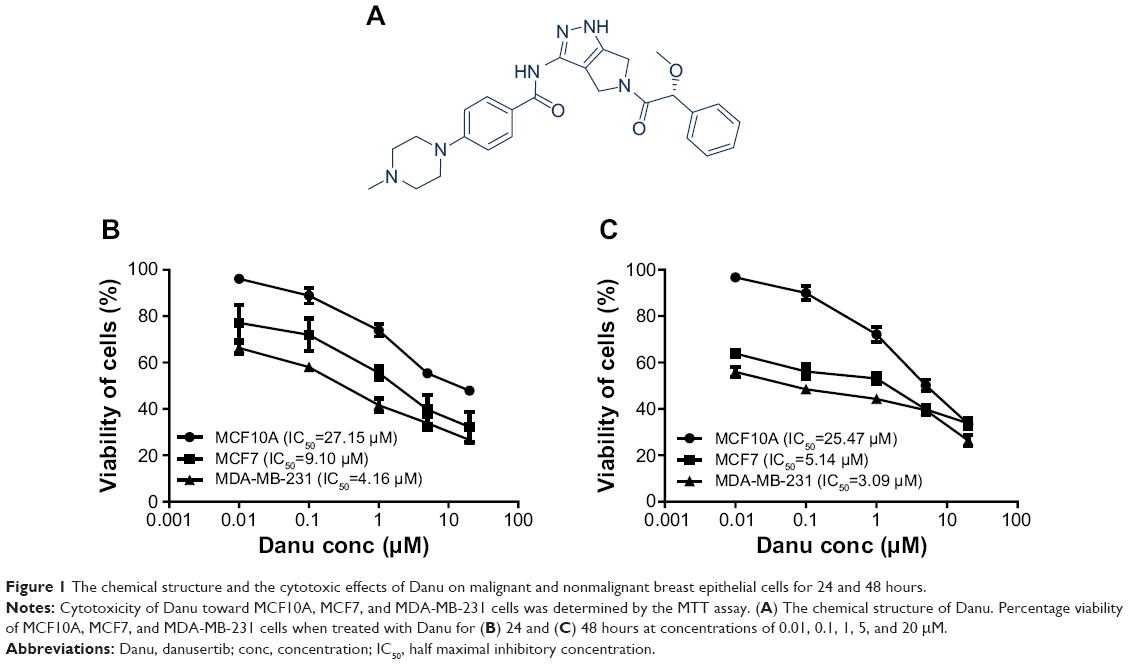 | Figure 1 The chemical structure and the cytotoxic effects of Danu on malignant and nonmalignant breast epithelial cells for 24 and 48 hours. |
Cell lines and cell culture
MCF7 and MDA-MB-231 cells are the epithelial breast cancer cell lines, and MCF10A is the normal breast epithelial cell line. MCF-10A cells are often used as a normal control in breast cancer studies. These cells are derived from the mammary tissue of a cystic fibrosis patient and show normal mammary epithelial cell morphology. These cells do not have mutations in the p53 gene but show homozygous loss of the p16/p15 locus and do not form tumors in nude mice or colonies in semi-solid low melting agarose. MCF7, MDA-MB-231, and MCF10A cells were all obtained from American Type Culture Collection (Manassas, VA, USA). MCF7 and MDA-MB-231 cells were maintained in RPMI 1640 medium supplemented with heat-inactivated 10% FBS and 1% penicillin/streptomycin. MCF10A cells were grown in DMEM/F12 (1:1) supplemented with 5% horse serum, 10 μg/mL insulin, 100 ng/mL cholera enterotoxin, 0.5 mg/mL hydrocortisone, and 20 ng/mL epidermal growth factor. All cells were maintained in a 5% CO2/95% air-humidified incubator at 37°C. Danu was dissolved in dimethyl sulfoxide (DMSO) with a stock concentration of 25 mM and freshly diluted to the predetermined concentrations with culture medium, with the final concentration of DMSO at 0.05% (v/v). The control cells received the vehicle only.
Cell viability assay
The effect of Danu on cell viability was examined using the MTT assay. Briefly, cells were seeded in 96-well culture plates at a density of 8×103 cells/well. After cells were attached, the cells were treated with Danu at different concentrations ranging from 0.01 μM to 20 μM. The concentration of DMSO was at 0.05% (v/v). The control cells received the vehicle only. After 24- or 48-hour incubation, 10 μL MTT (5 mg/L) was added to each well and cultured for a further 4 hours. Then, solution was removed and 150 μL DMSO was added to dissolve the crystal. The absorbance was measured at a wavelength of 490 nm using a Synergy™ H4 Hybrid microplate reader (BioTek Inc., Winooski, VT, USA). The half maximal inhibitory concentration (IC50) values were determined using the relative viability over Danu concentration curve.
Cell cycle distribution analysis
The effect of Danu on cell cycle distribution was examined using PI as a DNA stain to determine DNA content by flow cytometry, as described previously.21 Briefly, MCF10A, MCF7, and MDA-MB-231 cells were treated with Danu at concentrations of 0.01, 0.1, and 0.5 μM for 24 hours. In separate experiments, MCF10A, MCF7, and MDA-MB-231 cells were treated with 0.5 μM Danu for 4, 8, 12, 24, 48, and 72 hours. Following the Danu treatment, cells were detached, washed by D-PBS, centrifuged, and fixed in 70% ethanol at −20°C overnight. Then, the cells were resuspended in 1 mL of PBS containing 1 mg/mL RNase A and 50 μg/mL PI. Cells were incubated in the dark for 30 minutes at room temperature. The cells were subjected to cell cycle analysis using a flow cytometer (Becton Dickinson Immunocytometry Systems, San Jose, CA, USA). The flow cytometer collected 10,000 events for analysis.
Quantification of cellular apoptosis
Apoptotic cells were quantitated using the annexin V:PE apoptosis detection after cells were treated with Danu at concentrations of 0.01, 0.1, and 0.5 μM for 24 hours, as described previously.21 Briefly, cells were trypsinized and washed twice with cold PBS, and then resuspended in 1× binding buffer with 2.5 μL of annexin V:PE and 2.5 μL of 7-amino-actinomycin D (7-AAD, used as a nucleic acid dye) at 1×105 cells/mL in a total volume of 100 μL. The cells were gently mixed and incubated in the dark for 15 minutes at room temperature. A quota of 1× binding buffer (400 μL) was then added to a clean test tube and the number of apoptotic cells was quantified using a flow cytometer within 1 hour. Cells that stain positive for annexin V:PE and negative for 7-AAD are undergoing apoptosis; cells that stain positive for both annexin V:PE and 7-AAD are either in the endstage of apoptosis, are undergoing necrosis, or are already dead; cells that stain negative for both annexin V:PE and 7-AAD are alive and not undergoing apoptosis.
Quantification of cellular autophagy
MCF10A, MCF7, and MDA-MB-231 cells were treated with fresh medium alone, control vehicle alone (0.05% DMSO, v/v), or Danu (0.01, 0.1, and 0.5 μM) for 24 hours at 37°C. In separate experiments, MCF10A, MCF7, and MDA-MB-231 cells were treated with 0.5 μM Danu for 4, 8, 12, 24, 48, and 72 hours. To investigate the mechanisms for Danu-induced autophagy, cells were pretreated with 10 μM WM (a PI3K inhibitor and autophagy blocker), 10 μM bafilomycin A1 (an autophagy inhibitor), 10 μM SB202190 (a selective inhibitor of p38 MAPK), or 10 μM U0126 (a selective inhibitor of Erk1/2) for 30 minutes in MCF7 and MDA-MB-231 cells, then co-treated with 0.5 μM Danu for a further 24 hours. Groups of cells treated with each of these compounds alone were also included. All inhibitors were dissolved in DMSO at a final concentration of 0.05% (v/v). Cells were resuspended in 250 μL of phenol red-free culture medium (Thermo Fisher Scientific Inc.; No: 1294895) containing 5% FBS, and 250 μL of the diluted Cyto-ID® Green stain solution was added to each sample and mixed well. Cyto-ID® Green stain is a cationic amphiphilic tracer that selectively labels autophagic vacuoles in cells. Cells were incubated for 30 minutes at 37°C in the dark, collected by centrifugation, washed with 1× assay buffer, and resuspended in 500 μL fresh 1× assay buffer. Cells were subjected to flow cytometric analysis using the green (FL1) channel. The flow cytometer collected 10,000 events.
RNA interference
siRNA for silencing gene expression was performed by transfection of RNA oligonucleotides with lipofectamine 2000 according to the manufacturer’s instructions. MCF7 cells were transfected with the negative control siRNA, p38 MAPK siRNA, or Erk1/2 siRNA in Opti-MEM, using lipofectamine 2000. After incubation for 4 hours, the Opti-MEM™ medium was replaced with complete medium containing 10% FBS. The cells were then incubated for an additional 48 hours and treated with Danu. The cell samples were collected for further analysis.
Western blotting analysis
The expression levels of cellular proteins of interest were determined using Western blotting assays. MCF10A, MCF7, and MDA-MB-231 cells were washed with pre-chilled PBS after 24-hour treatment with Danu at 0.01, 0.1, and 0.5 μM, lysed with the radioimmunoprecipitation assay (RIPA) buffer containing the protease inhibitor and phosphatase inhibitor cocktails, and centrifuged at 3,000× g for 10 minutes at 4°C. Protein concentrations were measured using a Pierce™ BCA protein assay kit. Subsequently, equal amounts of protein samples (30 μg) were resolved by 4× sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) sample loading buffer and electrophoresed on 7%–12% SDS-PAGE mini-gel after thermal denaturation at 95°C for 5 minutes. Then, proteins were transferred onto methanol-activated PVDF membrane at 400 mA for 2 hours at 4°C. Membranes were blocked with 5% skim milk for 1 hour at room temperature, probed with the primary antibody overnight at 4°C, and then blotted with respective secondary anti-mouse or anti-rabbit antibody. Visualization was performed using Bio-Rad ChemiDoc™ XRS (Hercules, CA, USA) with enhanced-chemiluminescence substrate, and the blots were analyzed using Image Lab 3.0 (Bio-Rad). The matching densitometric value of the internal control β-actin was used to normalize the protein expression level.
Statistical analysis
Data are expressed as the mean ± standard deviation (SD). Comparisons of multiple groups were performed using one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison procedure. Values of P<0.05 were considered statistically significant. Assays were performed at least three times independently.
Results
Danu shows significantly higher cytotoxicity toward breast cancer cells than normal breast epithelial cells
In order to evaluate the cytotoxicity of Danu, we used the MTT assay to measure the inhibitory effect of Danu on the proliferation of malignant MCF-7 and MDA-MB-231 cells, and nonmalignant MCF10A cells. The concentration-dependent inhibitory effects of Danu on the growth of MCF10A, MCF7, and MDA-MB-231 cells are shown in Figure 1B and C. MCF10A, MCF7, and MDA-MB-231 cells were exposed to Danu for 24 or 48 hours at different concentrations. When MCF10A cells were treated with Danu at 0.01, 0.1, 1, 5, and 20 μM for 24 hours, the percentage of cellular viability over the control cells (100%) was 96.1%, 88.9%, 73.9%, 55.4%, and 47.9%, respectively; the IC50 value of 24-hour Danu treatment was 27.15 μM. The percentage of live MCF7 cells over the control cells (100%) was 77.2%, 71.9%, 55.5%, 39.7%, and 32.2%, respectively. The IC50 value of 24-hour treatment was 9.10 μM. The percentage of live MDA-MB-231 cells over the control cells (100%) was 66.3%, 58%, 41.6%, 33.9%, and 26.6%, respectively; the IC50 value of 24-hour treatment was 4.16 μM (Figure 1B). When MCF10A cells were treated with Danu at 0.01, 0.1, 1.0, 5.0, and 20 μM for 48 hours, the percentage of cellular viability over the control cells (100%) was 96.6%, 89.9%, 72.1%, 50.1%, and 33.6%, respectively; the IC50 value of 48-hour treatment was 25.47 μM. The percentage of live MCF7 cells over the control cells (100%) was 63.9%, 56.1%, 53.1%, 39.8%, and 33.8%, respectively; the IC50 value of 48-hour treatment was 5.14 μM. The percentage of live MDA-MB-231 cells over the control cells (100%) was 55.9%, 48.5%, 44.3%, 39.5%, and 26.4%, respectively; the IC50 value of 48-hour treatment was 3.09 μM (Figure 1C). These results indicate that Danu induces concentration-dependent inhibitory effects on the growth of MCF10A, MCF7, and MDA-MB-231 cells, but that the cytotoxic effect of Danu is much lower in normal MCF10A cells than that in both malignant cell lines.
Danu induces G2/M arrest in breast cancer cells
Since we have observed an inhibitory effect on cell growth, we further assessed the effect of Danu on the cell cycle distribution of MCF10A, MCF7, and MDA-MB-231 cells by flow cytometry. Treatment of MCF10A, MCF7, and MDA-MB-231 cells with Danu arrested cells in G2/M phase in a concentration- and time-dependent manner. At basal level, the percentage of MCF10A, MCF7, and MDA-MB-231 cells in G2/M phase was 28.3%, 21.4%, and 25.9%, respectively. As shown in Figure 2A and B, there was a significant increase in the cell number in G2/M phase in MCF10A, MCF7, and MDA-MB-231 cells after incubation with Danu at 0.5 μM for 24 hours. The percentage of cells arrested in G2/M phase was 70.8%, 69.1%, and 80.6% in MCF10A, MCF7, and MDA-MB-231 cells, respectively. This gave an increase of 1.5-, 2.2-, and 2.1-fold, respectively, compared to the control cells treated with DMSO only (P<0.001 by one-way ANOVA; Figure 2B). The increase in magnitude of the number of malignant breast cells in G2/M phase was 1.4- and 1.5-fold higher than that in normal breast epithelial cells. However, a lower concentration of Danu (0.01 and 0.1 μM) did not significantly affect the number of MCF10A, MCF7, and MDA-MB-231 cells (P>0.05 by one-way ANOVA; Figure 2B). Treating MCF10A, MCF7, and MDA-MB-231 cells with Danu at 0.5 μM also significantly affected the number of cells in G1 phase. The number of cells in G1 phase was significantly reduced by 60.5%, 63.3%, and 77.3% in MCF10A, MCF7, and MDA-MB-231 cells treated with Danu at 0.5 μM, respectively (P<0.001 by one-way ANOVA; Figure 2B).
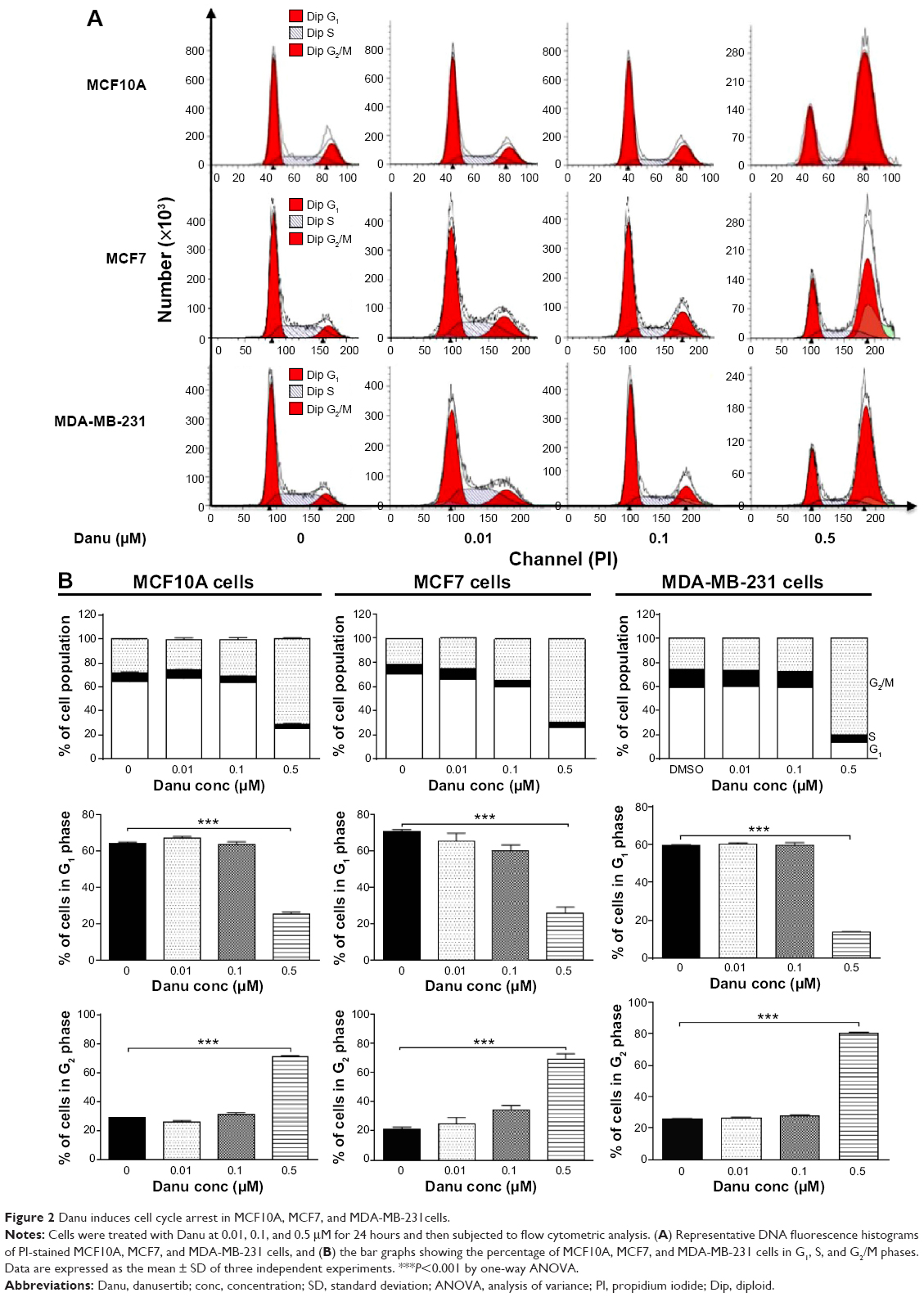 | Figure 2 Danu induces cell cycle arrest in MCF10A, MCF7, and MDA-MB-231cells. |
We further conducted separate experiments to evaluate the effect of Danu treatment at 0.5 μM on the cell cycle distribution of MCF10A, MCF7, and MDA-MB-231 cells over 72 hours. Compared to the control cells, the percentage of MCF10A cells in G2/M phase was increased from 9.6% at basal level to 14.8%, 17.5%, 21.4%, 21.7%, 73.8%, and 78.4% after 4-, 8-, 12-, 24-, 48-, and 72-hour treatment with 0.5 μM Danu, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 3A and B). There was a marked increase in the percentage of MCF10A cells in G2/M phase when treated with Danu for 12 to 72 hours (P<0.01 or 0.001 by one-way ANOVA; Figure 3A and B). The percentage of MCF7 cells in G2/M phase was increased from 11.1% at basal level to 15.2%, 22.5%, 33.4%, and 55.6% after 4, 8, 12, and 24 hours of treatment with 0.5 μM Danu, and declined to 39.9% and 22.7% after 48 and 72-hour treatment of Danu, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 3A and B). There was a remarkable increase in the percentage of MCF7 cells in G2/M phase after they were treated with Danu for 8 to 72 hours (P<0.01 or 0.001 by one-way ANOVA; Figure 3A and B). The percentage of MDA-MB-231 cells in G2/M phase was increased from 11.4% at basal level to 23.3%, 26.5%, 31.9%, and 70.4% after 4-, 8-, 12-, and 24-hour treatment with 0.5 μM Danu, and declined to 38.3% and 24.3% after 48- and 72-hour treatment of 0.5 μM Danu, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 3A and B). In addition, there was a marked increase in the percentage of MCF7 cells in G2/M phase when cells were treated with Danu for 4 to 72 hours (P<0.01 or 0.001 by one-way ANOVA; Figure 3A and B). These data demonstrate a concentration- and time-dependent G2/M arrest in breast cancer cells by Danu.
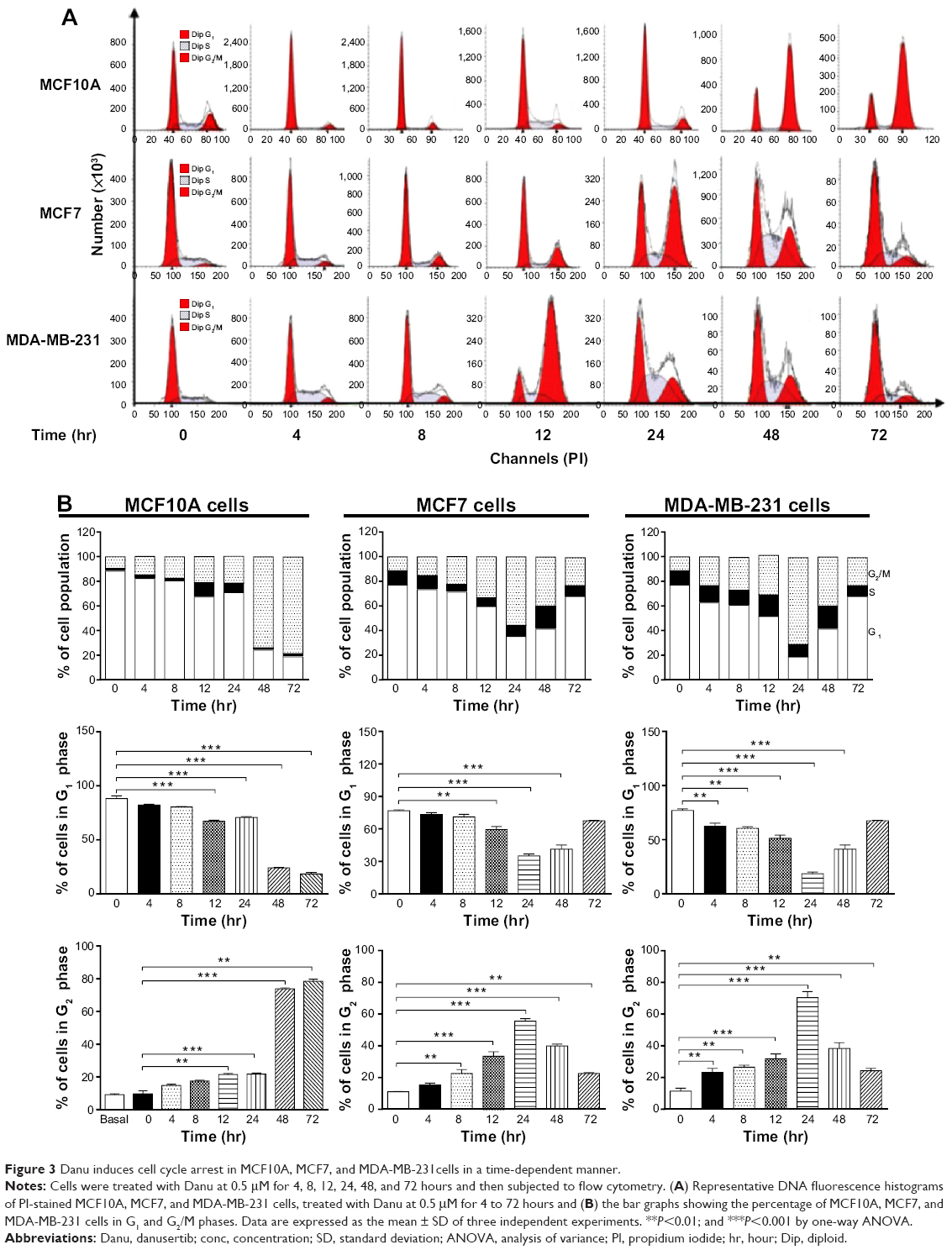 | Figure 3 Danu induces cell cycle arrest in MCF10A, MCF7, and MDA-MB-231cells in a time-dependent manner. |
Danu regulates the expression of CDK1/CDC2, CDK2, cyclin B1, p21 Waf1/Cip1, p27 Kip1, and p53 in MCF10A, MCF7, and MDA-MB-231cells
To explore the mechanism for Danu-induced cell cycle arrest, we examined the effect of Danu treatment on the expression levels of CDK1/CDC2, CDK2, cyclin B1, p21 Waf1/Cip1, p27 Kip1, and p53 in MCF10A, MCF7, and MDA-MB-231 cells using Western blotting assay. The expression levels of CDK1/CDC2, CDK2, and cyclin B1 were significantly decreased, whereas the expression levels of p21 Waf1/Cip1, p27 Kip1, and p53 were significantly increased in MCF10A, MCF7, and MDA-MB-231cells treated with Danu (Figure 4A and B). The expression of cyclin B1 was significantly suppressed in MCF10A cells with the treatment of Danu at 0.01, 0.1, and 0.5 μM for 24 hours. In comparison to the control cells, the expression level of cyclin B1 was decreased by 21%, 27%, and 65% when treated with 0.01, 0.1, and 0.5 μM Danu for 24 hours, respectively. There was a 37.5%, 40.6%, and 67.2% reduction in the expression level of CDK1/CDC2 in MCF10A cells incubated with 0.01, 0.1, and 0.5 μM Danu for 24 hours, respectively. Treatment of MCF10A cells with Danu at 0.01, 0.1, and 0.5 μM for 24 hours significantly increased the level of p27 Kip1 and p53 by 3.0-, 3.0-, and 7.5-fold and 5.1-, 6.6-, and 5.4-fold, respectively. However, there was no significant change in the expression levels of CDK2 and p21 Waf1/Cip1, when MCF10A cells were treated with 0.01, 0.1, and 0.5 μM Danu for 24 hours (P>0.05, by one-way ANOVA; Figure 4A and B). The expression level of cyclin B1 was significantly decreased in MCF7 cells with the treatment of Danu at 0.1 and 0.5 μM for 24 hours. In comparison to the control cells, the expression level of cyclin B1 was decreased 30.6% and 29.2% when treated with 0.1 and 0.5 μM Danu for 24 hours, respectively. There was a 9.6%, 22.5%, and 38.5% reduction in the expression level of CDK1/CDC2 in MCF7 cells incubated with 0.01, 0.1, and 0.5 μM Danu for 24 hours, respectively. The expression level of CDK2 was decreased by 16.3% and 35.9% when treated with 0.1 and 0.5 μM Danu for 24 hours, respectively. Treatment of MCF7 cells with Danu at 0.1 and 0.5 μM for 24 hours significantly increased the level of p21 Waf1/Cip1 and p27 Kip1 by 1.7- and 3.2-fold, and 1.9- and 2.5-fold, respectively. The expression level of p53 was increased 1.3-, 1.4-, and 1.8-fold when treated with 0.01, 0.1, and 0.5 μM Danu for 24 hours in MCF7 cells, respectively (P<0.01 or P<0.001 by one-way ANOVA; Figure 4A and B). Similarly, treatment of MDA-MB-231 cells with Danu at 0.01, 0.1, and 0.5 μM for 24 hours significantly decreased by 24.4%, 39.8%, and 46.3% in the level of CDK1/CDC2 and 17.9%, 16.1%, and 30.4% in the level of CDK2, respectively. There was a 12.8%, 55.3%, and 53.2% reduction in the expression level of cyclin B1 in MDA-MB-231 cells incubated with 0.01, 0.1, and 0.5 μM Danu for 24 hours, respectively. Treatment of MDA-MB-231 cells with Danu at 0.01, 0.1, and 0.5 μM for 24 hours significantly increased the level of p21 Waf1/Cip1 and p27 Kip1 by 1.4-, 2.4-, and 3.6-fold, and 3.5-, 3.6-, and 10.1-fold, respectively. The expression level of p53 was increased 1.4-, 1.5-, and 2.4-fold when treated with 0.01, 0.1, and 0.5 μM Danu for 24 hours in MDA-MB-231 cells, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 4A and B).
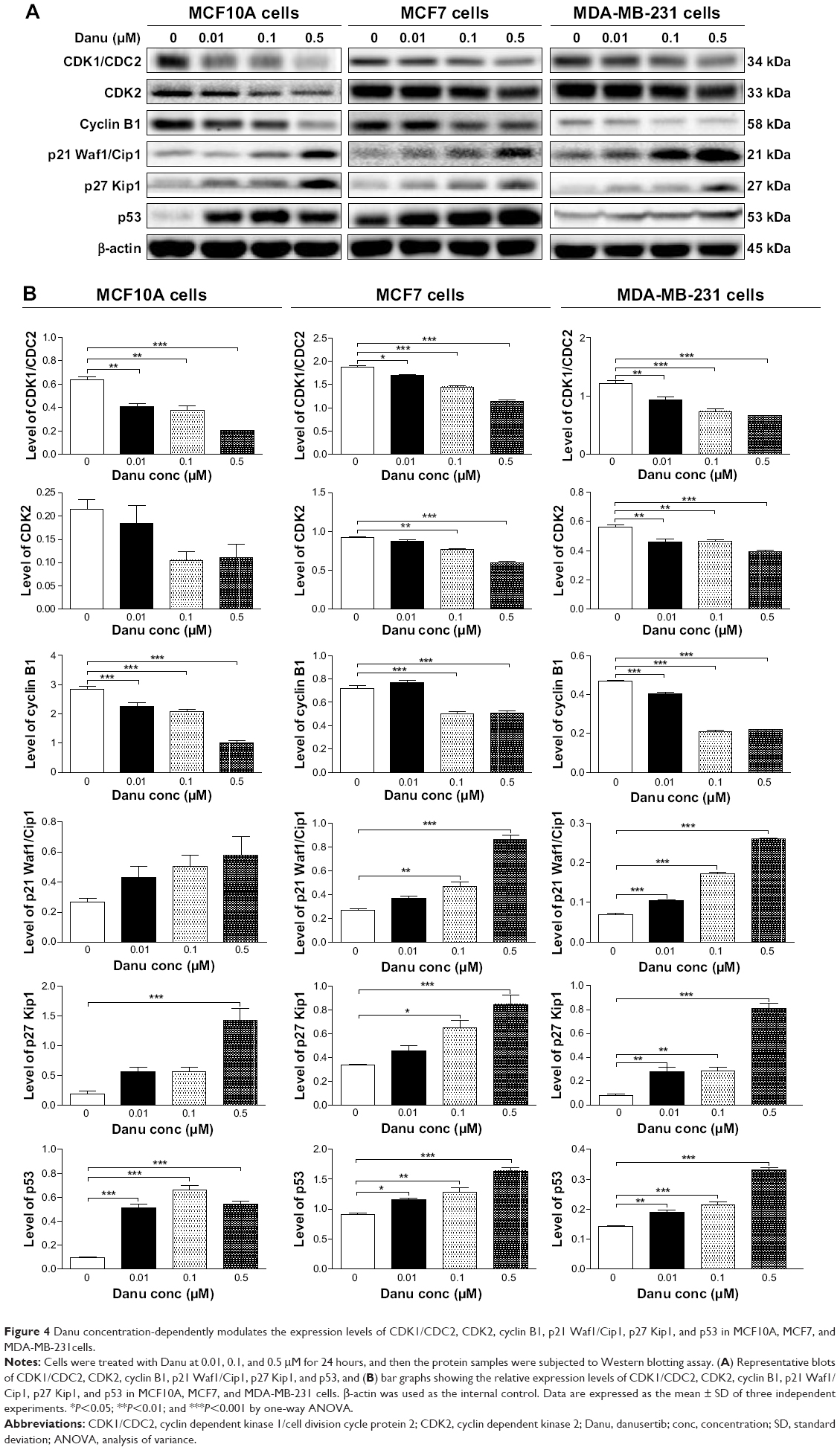 | Figure 4 Danu concentration-dependently modulates the expression levels of CDK1/CDC2, CDK2, cyclin B1, p21 Waf1/Cip1, p27 Kip1, and p53 in MCF10A, MCF7, and MDA-MB-231cells. |
To further investigate the molecular mechanism for Danu-induced cell cycle arrest, we conducted separate experiments to evaluate the expression levels of CDK1/CDC2, cyclin B1, p21 Waf1/Cip1, and p53 in MCF10A, MCF7, and MDA-MB-231 cells with the treatment of Danu over 72 hours using Western blotting assay. Treatment of MCF10A cells with Danu at 0.5 μM for 4, 8, 12, 24, 48, and 72 hours significantly decreased the level of cyclin B1 by 16.2%, 26.6%, 48.7%, 90.9%, 91.1%, and 96.2%, respectively. There was a 23.1%, 8.3%, 16.9%, 15.3%, and 15.4% reduction in the expression level of CDK1/CDC2 in MCF10A cells incubated with 0.5 μM Danu for 4, 8, 12, 24, and 72 hours, respectively. Treatment of MCF10A cells with Danu at 0.5 μM for 8, 24, and 48 hours significantly increased the level of p21 Waf1/Cip1 by 1.6-, 4.1-, and 1.9-fold, respectively. The expression level of p53 was increased 2.2-, 2.4-, 5.9-, 3.9-, and 4.1-fold when treated with 0.5 μM Danu for 8, 12, 24, 48, and 72 hours in MCF10A cells, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 5A and B). Incubation of MCF7 cells with Danu at 0.5 μM for 4, 8, 24, 48, and 72 hours significantly decreased the level of CDK1/CDC2 by 28.4%, 19.0%, 19.1%, 46.9%, and 64.0%, respectively. There was a 29.7%, 38.0%, and 38.2% reduction in the expression level of cyclin B1 in MCF7 cells incubated with 0.5 μM Danu for 8, 24, and 72 hours, respectively. Treatment of MCF7 cells with Danu at 0.5 μM for 8, 24, and 72 hours significantly increased the level of p21 Waf1/Cip1 by 3.2-, 4.4-, and 10-fold, respectively. The expression level of p53 was increased 1.7-, 1.7-, and 2.3-fold when treated with 0.5 μM Danu for 12, 24, and 72 hours in MCF7 cells, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 5A and B).
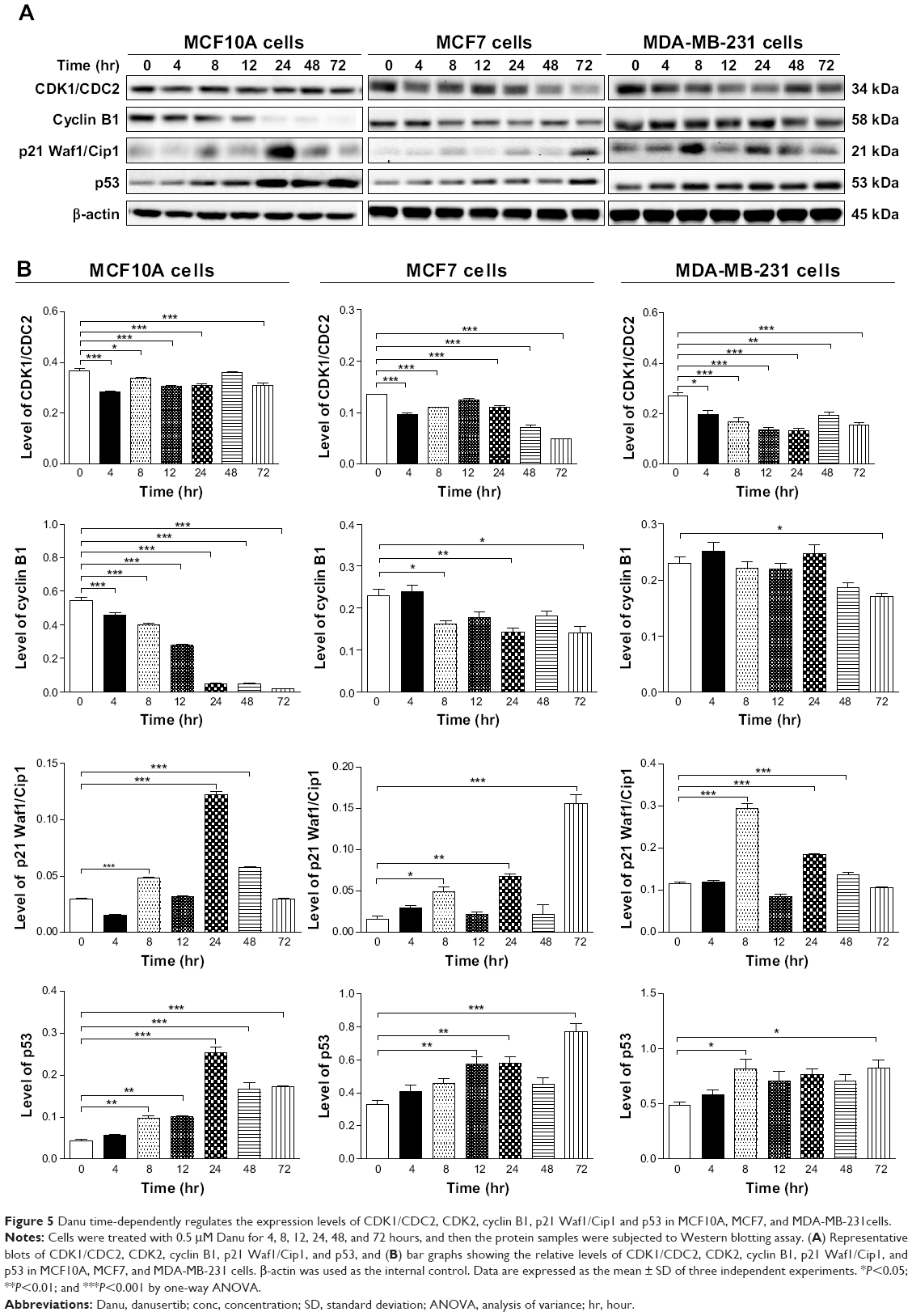 | Figure 5 Danu time-dependently regulates the expression levels of CDK1/CDC2, CDK2, cyclin B1, p21 Waf1/Cip1 and p53 in MCF10A, MCF7, and MDA-MB-231cells. |
Treatment of MDA-MB-231 cells with Danu at 0.5 μM for 4, 8, 12, 24, 48, and 72 hours significantly decreased the level of CDK1/CDC2 by 26.6%, 37.5%, 48.8%, 50.8%, 28.6%, and 42.8%, respectively. There was a 26% reduction in the expression level of cyclin B1 in MDA-MB-231 cells incubated with 0.5 μM Danu for 72 hours. Treatment of MDA-MB-231 cells with Danu at 0.5 μM for 8, 24, and 48 hours significantly increased the expression level of p21 Waf1/Cip1 by 2.6-, 1.6-, and 1.2-fold, respectively. The expression level of p53 was increased 1.6- and 1.7-fold when treated with 0.5 μM Danu for 8 and 72 hours in MDA-MB-231 cells, respectively (P<0.01 or 0.001 by one-way ANOVA; Figure 5A and B).
Danu induces the apoptosis of MCF10A, MCF7, and MDA-MB-231 cells via activation of mitochondrial pathway
To examine the apoptosis-inducing effect of Danu in MCF10A, MCF7, and MDA-MB-231 cells, the number of apoptotic cells was first quantified using a flow cytometric analysis; the results are shown in Figure 6A and B. The number of apoptotic cells was 12.7%, 16.7%, and 15.8% in MCF10A, MCF7, and MDA-MB-231 cells treated with the control vehicle only (0.05% DMSO, v/v), respectively. In MCF10A cells treated with Danu at 0.1 and 0.5 μM for 24 hours, the total percentages of apoptotic cells (early plus late apoptosis) were 20.1% and 36.6%, respectively, with a 1.6- and 2.9-fold increase compared to the control cells. When MCF7 cells were treated with Danu at 0.01, 0.1, and 0.5 μM for 24 hours, the total percentages of apoptotic cells were increased 1.4-, 2.0-, and 2.6-fold compared to the control cells, respectively. Similarly, MDA-MB-231 cells were treated with Danu at 0.1 and 0.5 μM for 24 hours, the total percentages of apoptotic cells were increased 2.2- and 3.0-fold compared to the control cells, respectively (P<0.05 by one-way ANOVA; Figure 6A and B).
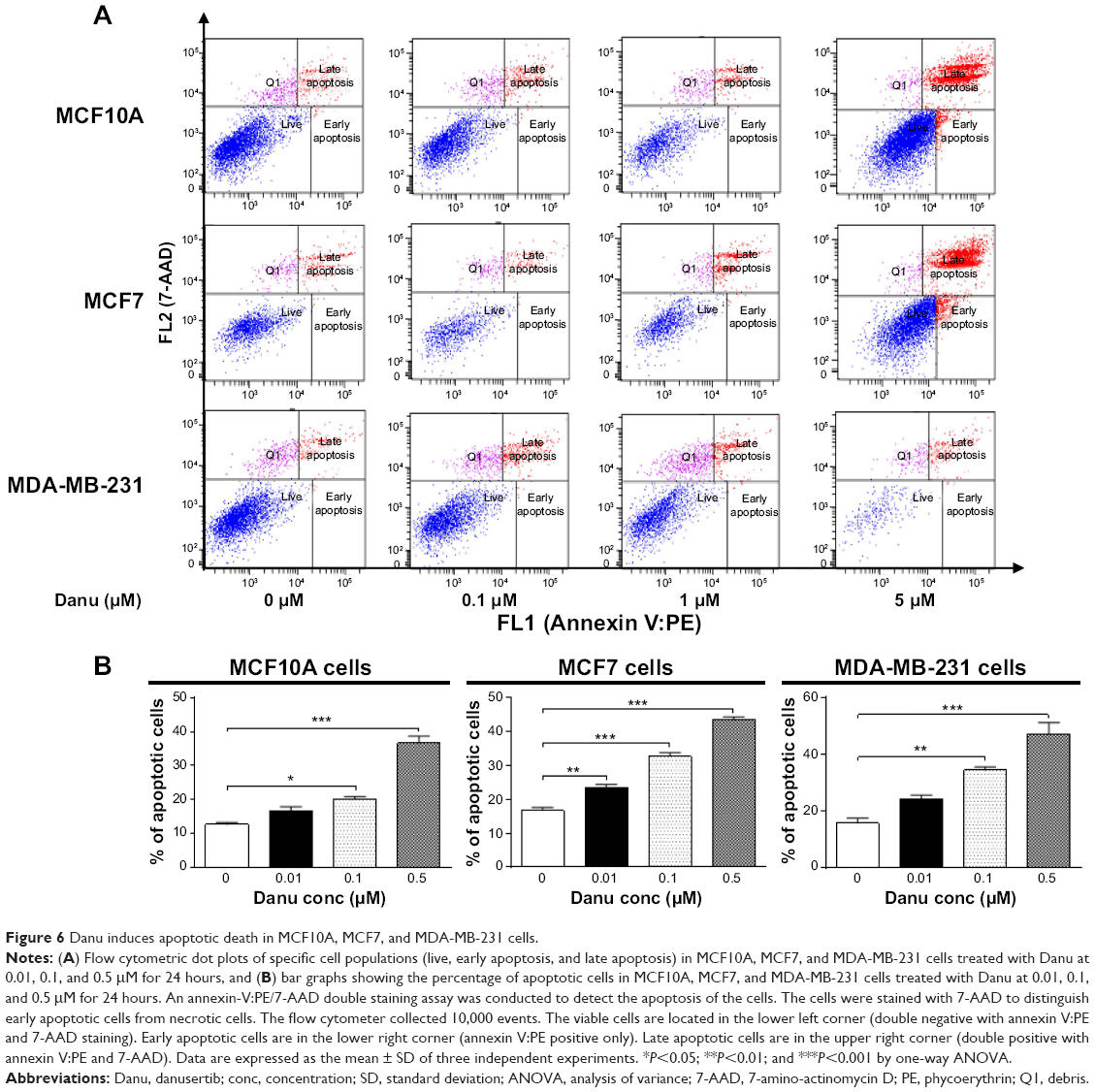 | Figure 6 Danu induces apoptotic death in MCF10A, MCF7, and MDA-MB-231 cells. |
To investigate the mechanism responsible for Danu-induced apoptosis in MCF10A, MCF7, and MDA-MB-231 cells, we evaluated the levels of Bcl-xl, Bcl-2, Bax, PUMA, cleaved caspase 3, and cleaved caspase 9 in the above three cell lines treated with Danu at 0.01, 0.1, and 0.5 μM for 24 hours. We first examined the effects of Danu treatment on the expression levels of the pro-apoptotic protein Bax and the anti-apoptotic proteins Bcl-2 and Bcl-xl. The expression level of Bax was concentration-dependently increased in three cell lines (P<0.05 by one-way ANOVA; Figure 7A and B). Incubation of MCF10A cells with 0.01 and 0.5 μM Danu for 24 hours significantly increased Bax expression 2.0-fold; treatment of MCF7 cells with 0.01 and 0.5 μM Danu for 24 hours resulted in an 1.2- and 1.4-fold increase in the expression of Bax, respectively; treatment of MDA-MB-231 cells with 0.01, 0.1, and 0.5 μM Danu for 24 hours remarkably increased the expression of Bax 2.0-fold. In contrast, the expression level of Bcl-2 was decreased by 46.4%, 58.0%, and 47.6% in MCF10A cells, 50.0%, 60.0%, and 55.0% in MCF7 cells, and 26.0%, 39.0%, and 50.0% in MDA-MB-231 cells when treated with Danu at 0.01, 0.1, and 0.5 μM, respectively. In addition, the expression level of Bcl-xl was significantly suppressed after treatment with Danu at 0.01, 0.1, and 0.5 μM in MCF7 and MDA-MB-231 cells cell lines. Compared to the control cells, the expression level of Bcl-xl was reduced by 21.4%, 42.9%, and 65.7% in MCF7 cells and 30.7%, 42%, and 37.2% in MDA-MB-231 cells when treated with Danu at 0.01, 0.1, and 0.5 μM for 24 hours, respectively (P<0.01 by one-way ANOVA; Figure 7A and B). However, there was no significant alteration in the expression level of Bcl-xl, when MCF10A cells were incubated with 0.01, 0.1, and 0.5 μM Danu for 24 hours (P>0.05 by one-way ANOVA; Figure 7A and B).
Additionally, the effect of Danu on the expression of PUMA was examined due to its important role in the regulation of anti-apoptotic proteins. Incubation of MCF10A, MCF7, and MDA-MB-231 cells with Danu increased the expression level of PUMA in a concentration-dependent manner. Treatment of cells with 0.01, 0.1, and 0.5 μM Danu for 24 hours significantly increased 1.4-fold in the expression level of PUMA in MCF7 and MDA-MB-231 cells (P<0.01 by one-way ANOVA; Figure 7A and B). In MCF10A cells, the expression level of PUMA increased 3.0- and 7.4-fold when treated with Danu at 0.1 and 0.5 μM for 24 hours (P<0.05 by one-way ANOVA; Figure 7A and B). Subsequently, we observed a significant increase in the activation of caspases 9 and 3 in MCF10A, MCF7, and MDA-MB-231 cells. Incubation of MCF10A with Danu at 0.1 and 0.5 μM for 24 hours significantly increased the level of cleaved caspase 3 and cleaved caspase 9 by 2.3- and 3.2-fold, respectively (P<0.05 by one-way ANOVA; Figure 7A and B). Similarly, treatment of MCF7 and MDA-MB-231 cells with 0.01, 0.1, and 0.5 μM Danu for 24 hours significantly increased the expression level of cleaved caspase 9 by 1.6-, 2.4-, and 3.2-fold, respectively, in both cell lines (P<0.001 by one-way ANOVA; Figure 7A and B). The level of cleaved caspase 3 was also increased 1.5- and 1.6-fold when MCF7 and MDA-MB-231 cells were treated with 0.1 and 0.5 μM Danu for 24 hours, respectively (P<0.01 by one-way ANOVA; Figure 7A and B). These results indicate that Danu induces a remarkable activation of caspases 9 and 3, and eventually leads to apoptotic death of MCF10A, MCF7, and MDA-MB-231 cells.
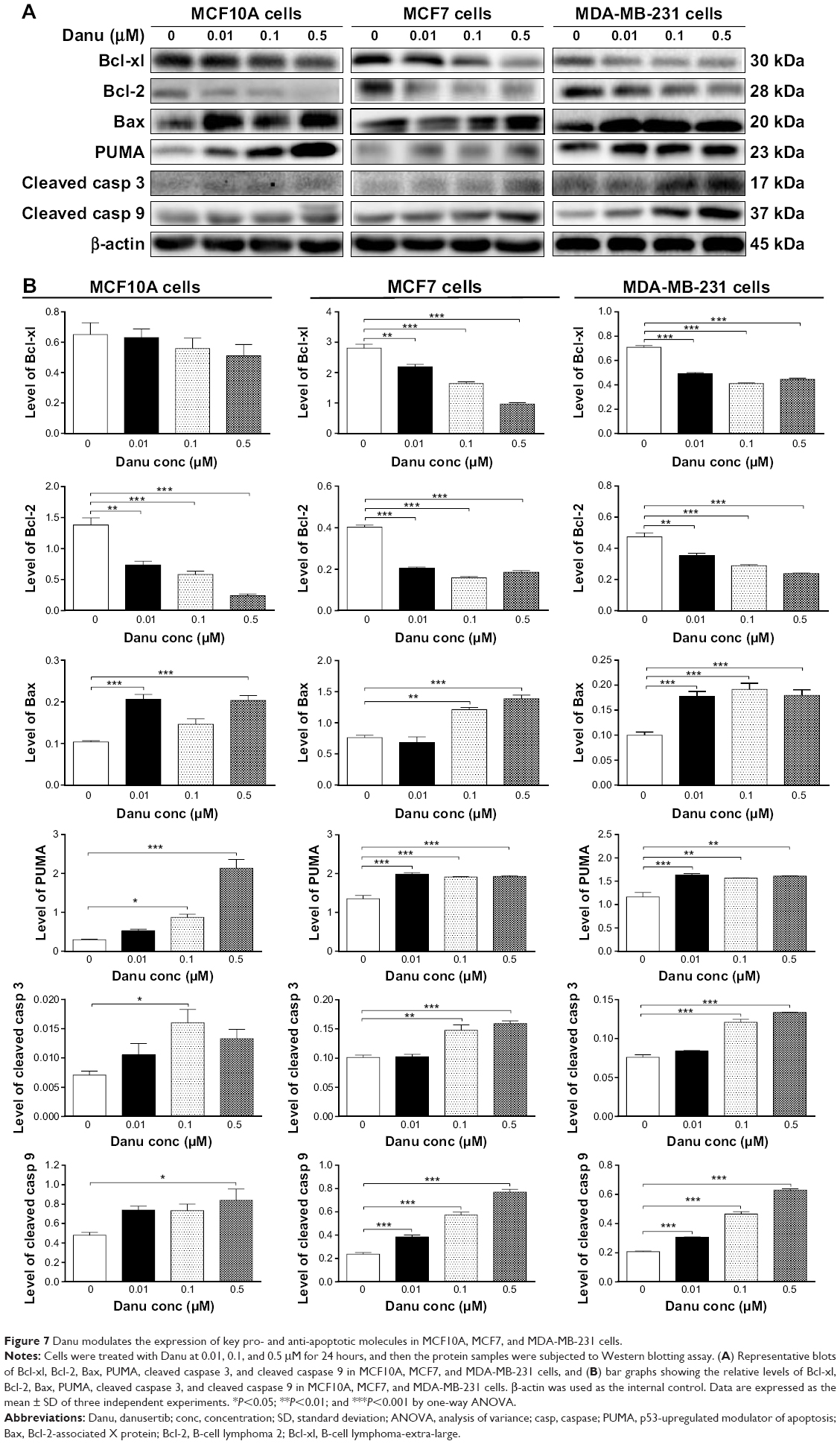 | Figure 7 Danu modulates the expression of key pro- and anti-apoptotic molecules in MCF10A, MCF7, and MDA-MB-231 cells. |
Danu induces the autophagy of MCF10A, MCF7, and MDA-MB-231 cells
Autophagy is a type II programmed cell death and a cytoprotective mechanism against extracellular stress. To determine the effect of Danu on autophagy, we first determined autophagic MCF10A, MCF7, and MDA-MB-231 cells using flow cytometric analysis. Danu treatment of MCF10A, MCF7, and MDA-MB-231 cells induced remarkable autophagy (Figures 8 and 9). In MCF10A cells, the percentage of autophagic cells at basal level was 0.56%; and incubation of the cells with Danu at 0.5 μM significantly increased the autophagy 10.9-fold (P<0.001 by one-way ANOVA; Figure 8A and B). Treatment of MCF10A cells with 0.5 μM Danu for 24, 48, and 72 hours increased the autophagy 5.0-, 5.2-, and 9.5-fold, respectively (P<0.05 by one-way ANOVA; Figure 9A and B). In MCF7 cells, the percentage of autophagic cells at basal level was 3.6%; incubation of the cells with Danu at 0.1 and 0.5 μM significantly increased the autophagy 6.4- and 6.0-fold, respectively (P<0.01 by one-way ANOVA; Figure 8A and B). Incubation of MCF7 cells with 0.5 μM Danu for 24, 48, and 72 hours increased the autophagy 9.2-, 23.6-, and 50.6-fold, respectively (P<0.01 by one-way ANOVA; Figure 9A and B). Similarly, in MDA-MB-231 cells, the percentage of autophagic cells at basal level was 1.3%; incubation of the cells with Danu at 0.5 μM significantly increased the autophagy 17.5-fold (P<0.001 by one-way ANOVA; Figure 8A and B). Exposure of MDA-MB-231 cells to 0.5 μM Danu for 24, 48, and 72 hours increased the autophagy 6.2-, 14.1-, and 11.9-fold, respectively (P<0.01 by one-way ANOVA; Figure 9A and B).
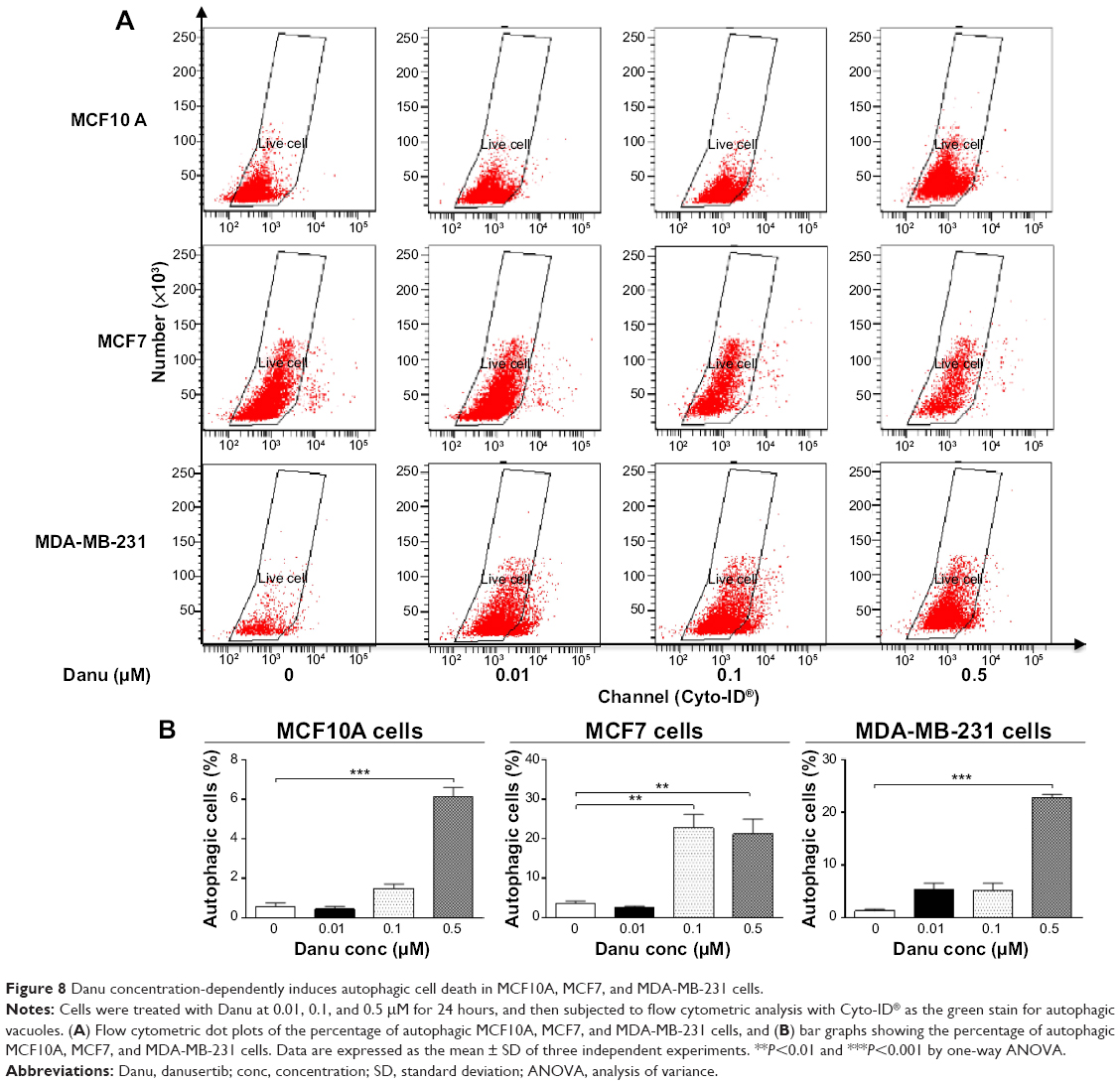 | Figure 8 Danu concentration-dependently induces autophagic cell death in MCF10A, MCF7, and MDA-MB-231 cells. |
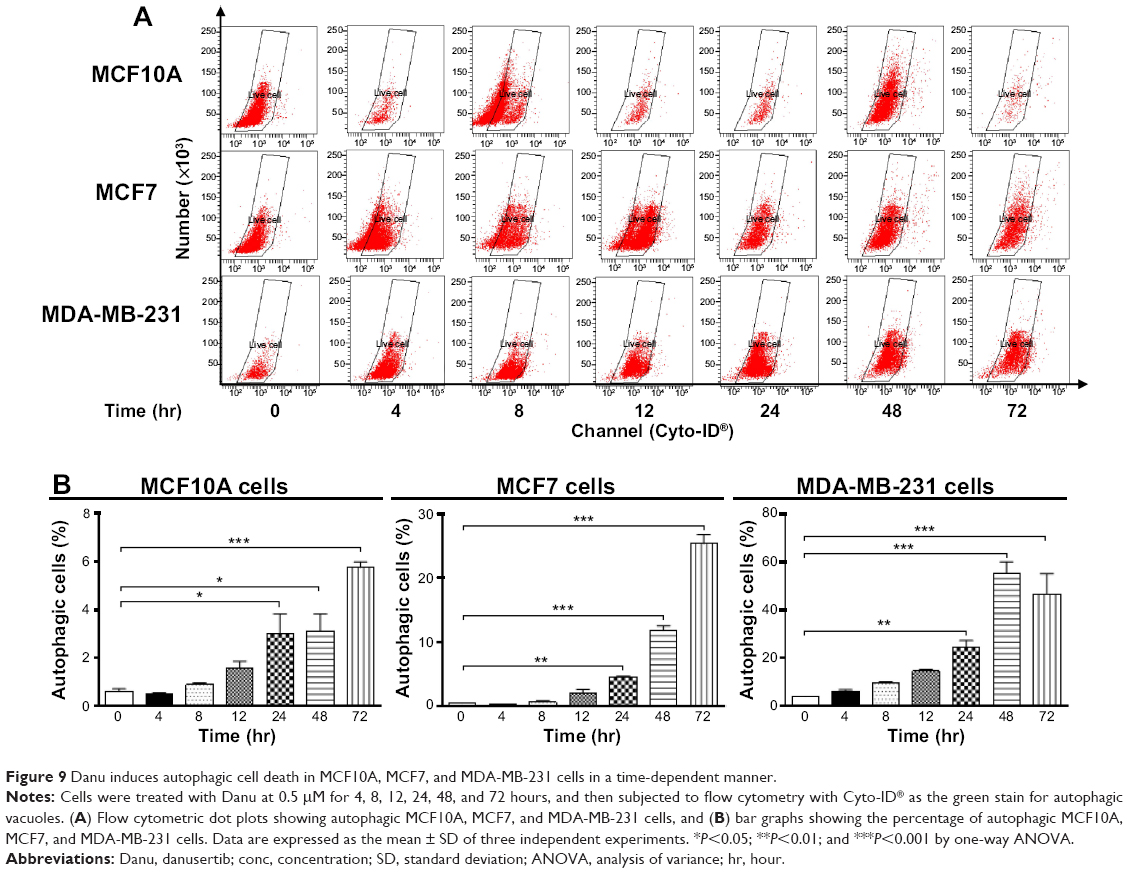 | Figure 9 Danu induces autophagic cell death in MCF10A, MCF7, and MDA-MB-231 cells in a time-dependent manner. |
Next, we examined the effect of Danu on the expression levels of beclin 1, LC3-I, and LC3-II. Beclin 1 plays an pivotal role in the process of autophagy through the formation of a complex with vacuolar protein sorting 34 (Vps34, also called class III PI3K) and recruitment of other Atgs that are critical for subsequent autophagosome formation.22 Upon autophagy initiation, LC3 is cleaved at the C-terminus by Atg4 to form the cytosolic LC3-I.23 LC3-I is consequently proteolytically cleaved and lipidated by Atg3 and Atg7 to form LC3-II, which localizes to the autophagosome membrane. Treatment of MCF7 cells with Danu at 0.01, 0.1, and 0.5 μM for 24 hours significantly increased the expression level of beclin 1 1.5-, 1.4-, and 1.5-fold, respectively, compared to the control cells (P<0.001 by one-way ANOVA; Figure 10A and B). There was a 1.2- and 1.4-fold increase in the level of beclin 1 in MDA-MB-231 cells treated with 0.1 and 0.5 μM Danu for 24 hours, respectively (P<0.01 by one-way ANOVA; Figure 10A and B). However, incubation of MCF10A cells with Danu at 0.01, 0.1, and 0.5 μM for 24 hours did not significantly alter the expression level of beclin 1 (P>0.05 by one-way ANOVA; Figure 10A and B). After 24-hour treatment with Danu at 0.01, 0.1, and 0.5 μM, our Western blotting analysis revealed two clear bands of LC3-I and -II in both MCF7 and MDA-MB-231 cells. In both MCF7 and MDA-MB-231 cells, there was a concentration-dependent increase in the expression of LC3-II (P<0.001 by one-way ANOVA; Figure 10A and B). Compared to the control cells, there was a 1.6-fold increase in the LC3-II level in MCF7 cells treated with Danu at 0.5 μM for 24 hours (P<0.001 by one-way ANOVA; Figure 10A and B). In MDA-MB-231 cells, treatment with 0.1 and 0.5 μM Danu resulted in a 2.0- and 3.2-fold increase in the expression of LC3-II, respectively, compared to the control cells (P<0.01 by one-way ANOVA; Figure 10A and B). In addition, treatment of MCF7 and MDA-MB-231 cells with Danu decreased the expression of LC3-I, although expression was not significantly different. In MCF10A cells, the relative expression level of LC3-II was negligible (0.02–0.03) in the control cells treated with the control vehicle only (0.05% DMSO, v/v) and treated with Danu at 0.01 and 0.1 μM (Figure 10A and B).
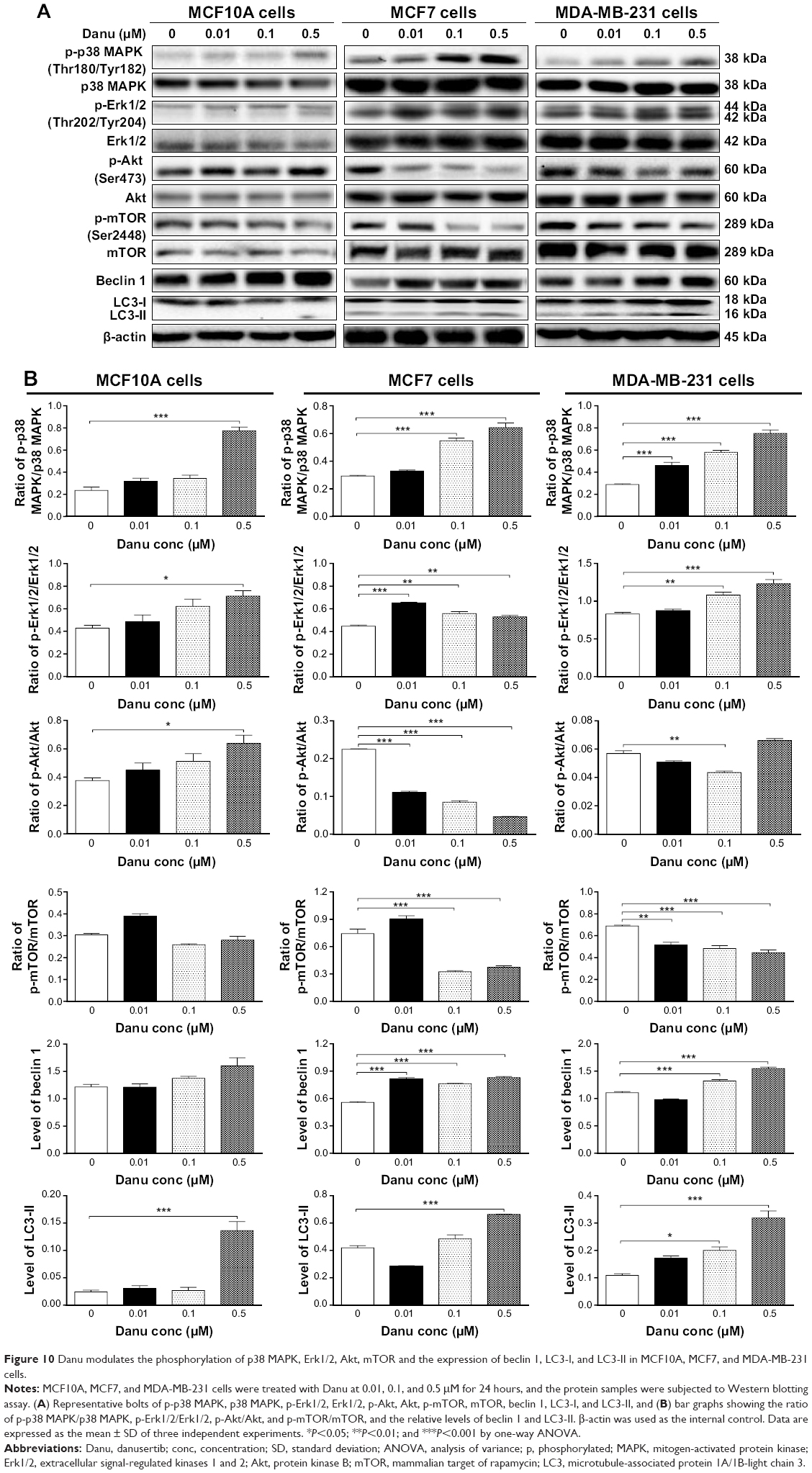 | Figure 10 Danu modulates the phosphorylation of p38 MAPK, Erk1/2, Akt, mTOR and the expression of beclin 1, LC3-I, and LC3-II in MCF10A, MCF7, and MDA-MB-231 cells. |
To further elucidate the effect of Danu on autophagy in three breast cell lines, we examined the effect of Danu on the expression levels of beclin 1 and LC3-II over 72 hours. Treatment of MCF7 cells with Danu at 0.5 μM for 8, 12, 24, and 72 hours significantly increased the expression of beclin 1 by 1.5-, 1.6-, 1.5-, and 2.5-fold, respectively, compared to the control cells (P<0.01 by one-way ANOVA; Figures 11B and 12B). There was a 1.2-, 1.4-, and 1.6-fold increase of beclin 1 in MDA-MB-231 cells treated with 0.5 μM Danu for 12, 24, and 48 hours, respectively (P<0.001 by one-way ANOVA; Figures 11C and 12C). However, incubation of MCF10A cells with Danu at 0.5 μM over 72 hours did not significantly alter the expression level of beclin 1 (P>0.05 by one-way ANOVA; Figures 11A and 12A). In both MCF7 and MDA-MB-231 cells, there was a marked increase in the expression of LC3-II. Compared to the control cells, there was a 2.1-, 2.1, and 2.6-fold increase in the LC3-II level in MCF7 cells treated with Danu at 0.5 μM for 4, 12, and 48 hours, respectively (P<0.01 by one-way ANOVA; Figures 11B and 12B). In MDA-MB-231 cells, 0.5 μM Danu over 72 hours resulted in a 1.5- and 2.5-fold increase in the expression of LC3-II, compared to the control cells (P<0.001 by one-way ANOVA; Figures 11C and 12C). In MCF10A cells, the relative expression level of LC3-II was also negligible (0.02–0.05) when cells were treated with Danu at 0.5 μM for 4, 8, 12, and 24 hours (Figures 11A and 12A). These results indicate that Danu induces autophagy in breast cancer cells, and MCF10A cells are less sensitive to Danu-induced autophagy than malignant MCF7 and MDA-MB-231 cells.
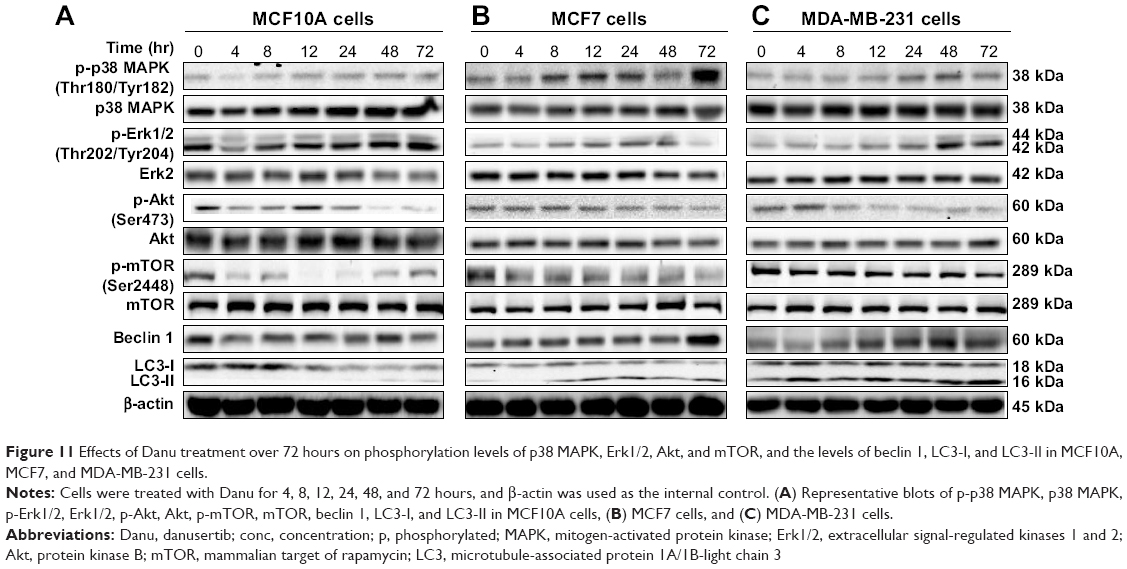 | Figure 11 Effects of Danu treatment over 72 hours on phosphorylation levels of p38 MAPK, Erk1/2, Akt, and mTOR, and the levels of beclin 1, LC3-I, and LC3-II in MCF10A, MCF7, and MDA-MB-231 cells. |
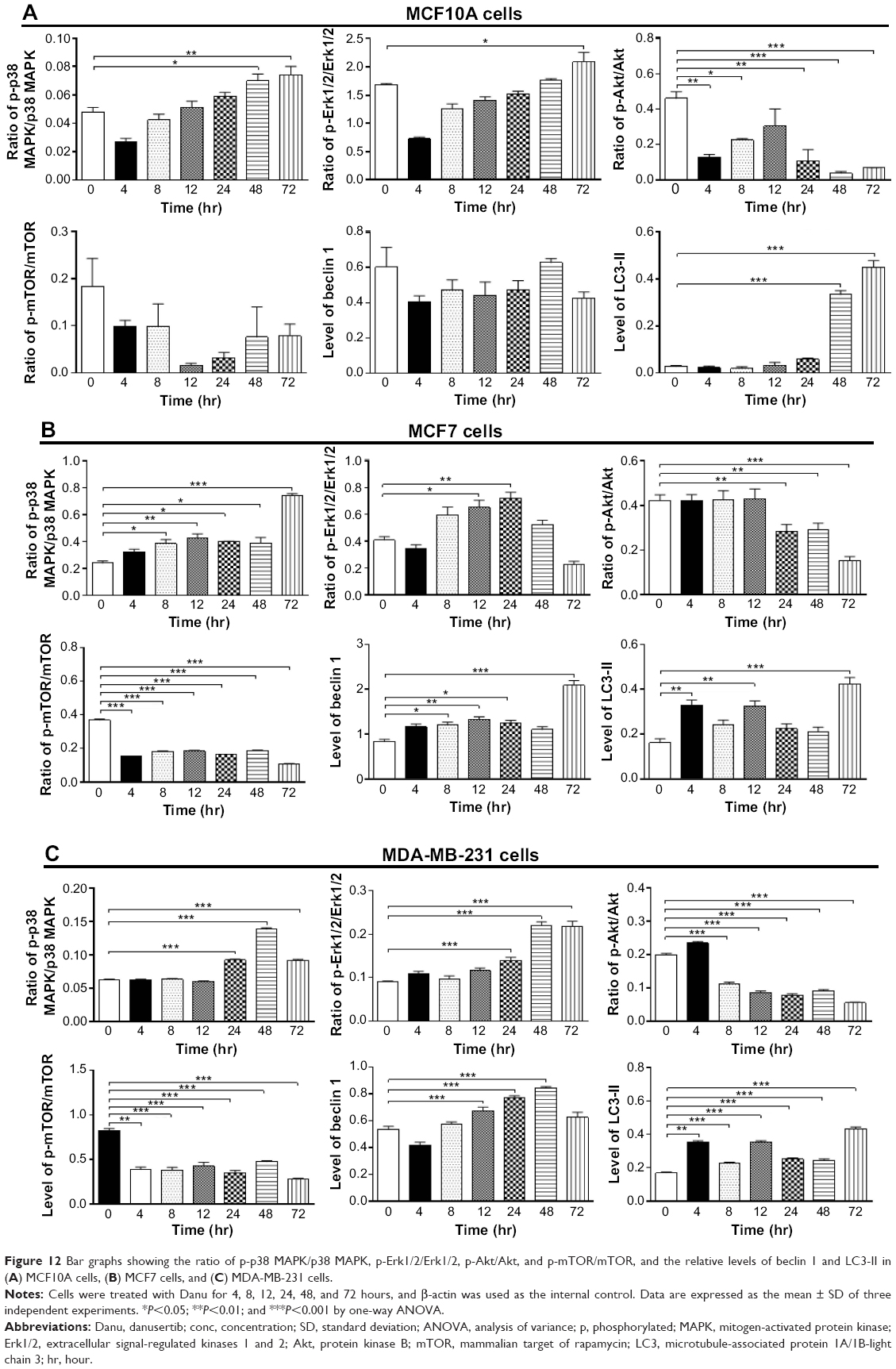 | Figure 12 Bar graphs showing the ratio of p-p38 MAPK/p38 MAPK, p-Erk1/2/Erk1/2, p-Akt/Akt, and p-mTOR/mTOR, and the relative levels of beclin 1 and LC3-II in (A) MCF10A cells, (B) MCF7 cells, and (C) MDA-MB-231 cells. |
Danu induces the activation of p38 MAPK and Erk1/2
MAPK family members, including c-Jun N-terminal kinase (JNK), Erk1/2, and p38 MAPK, have been reported to be involved in autophagy. To investigate whether the p38 MAPK and Erk1/2 signaling pathways were involved in Danu-induced autophagy, Western blotting assay was performed to detect the activated state of associated proteins. As shown in Figures 10, 11, and 12, Danu remarkably inhibited the phosphorylation of p38 MAPK; on the contrary, Danu induced the phosphorylation of Erk1/2 in three breast cell lines.
First, we examined the effect of Danu on the phosphorylation of p38 MAPK at Thr180/Tyr182 in the three cell lines. Exposure of MCF10A, MCF7, and MDA-MB-231 cells to 0.01, 0.1, and 0.5 μM Danu for 24 hours increased the phosphorylation level of p38 MAPK at Thr180/Tyr182; however, incubation of three cell lines with Danu did not significantly affect the expression of total p38 MAPK. The ratio of p-p38 MAPK at Thr180/Tyr182 over total p38 MAPK was concentration-dependently increased by Danu in both cell lines compared to the control cells. In MCF7 cells, the p-p38 MAPK/p38 MAPK ratio was increased from 0.29 at basal level to 0.32 and 0.64, when MCF7 cells were treated with Danu at 0.1 and 0.5 μM, respectively (P<0.001 by one-way ANOVA; Figure 10A and B). In MDA-MB-231 cells, 0.01, 0.1, and 0.5 μM Danu significantly decreased the ratio of p-p38 MAPK over p38 MAPK from 0.29 at basal level to 0.46, 0.58, and 0.75, respectively (P<0.001 by one-way ANOVA; Figure 10A and B). In MCF10A cells, only 0.5 μM Danu increased the ratio of p-p38 MAPK over total p38 MAPK. Treatment of MCF10A, MCF7, and MDA-MB-231 cells with Danu at 0.01, 0.1, and 0.5 μM for 24 hours increased the phosphorylation level of Erk1/2 at Thr202/Tyr204, but incubation of these three cell lines with Danu did not significantly affect the expression of total Erk1/2. The ratio of p-Erk1/2 over total Erk1/2 was concentration-dependently increased by Danu in all three cell lines compared to the control cells. In MCF7 cells, the p-Erk1/2:Erk1/2 ratio was increased from 0.45 at basal level to 0.65, 0.55, and 0.53, when MCF7 cells were treated with Danu at 0.01, 0.1, and 0.5 μM, respectively (P<0.001 by one-way ANOVA; Figure 10A and B). In MDA-MB-231 cells, 0.1 and 0.5 μM Danu significantly increased the ratio of p-Erk1/2 over total Erk1/2 from 0.83 at basal level to 1.08 and 1.23, respectively (P<0.001 by one-way ANOVA; Figure 10A and B). In MCF10A cells, only 0.5 μM Danu increased the ratio of p-Erk1/2 over total Erk1/2 from 0.42 at basal level to 0.71 (P<0.05 by one-way ANOVA; Figure 10A and B).
We further investigated the activation of MAPK by Danu in breast epithelial cell lines over 72 hours. A time-course study of p38 MAPK and Erk1/2 phosphorylation in MCF-7 and MDA-MB-231 cells showed that the activation of p38 MAPK and Erk1/2 was increased with drug treatment time. In MCF7 cells, the p-p38 MAPK/p38 MAPK ratio was increased from 0.26 at basal level to 0.30, 0.42, 0.46, 0.27, 0.4, and 0.76, when MCF7 cells were treated with Danu at 0.5 μM for 4, 8, 12, 24, 48, and 72 hours, respectively (P<0.001 by one-way ANOVA; Figures 11B and 12B). In MDA-MB-231 cells, treatment with 0.5 μM Danu for 24, 48, and 72 hours significantly increased the ratio of p-p38 MAPK over total p38 MAPK from 0.06 at basal level to 0.09, 0.14, and 0.09, respectively (P<0.001 by one-way ANOVA; Figures 11C and 12C). In MCF10A cells, treatment with 0.5 μM Danu for 48 and 72 hours increased the ratio of p-p38 MAPK over total p38 MAPK from 0.05 at basal level to 0.07 (P<0.05 by one-way ANOVA; Figures 11A and 12A). Treatment of MCF10A, MCF7, and MDA-MB-231 cells with Danu at 0.5 μM over 72 hours increased the activation of Erk1/2 at Thr202/Tyr204. In MCF7 cells, the p-Erk1/2:Erk1/2 ratio was increased from 0.41 at basal level to 0.65 and 0.72, when MCF7 cells were treated with 0.5 μM Danu for 12 and 24 hours, respectively (P<0.05 by one-way ANOVA; Figures 11B and 12B). In MDA-MB-231 cells, treatment with 0.5 μM Danu significantly increased the ratio of p-Erk1/2 over total Erk1/2 from 0.09 at basal level to 0.14, 0.22, and 0.22, respectively (P<0.001 by one-way ANOVA; Figures 11C and 12C). In MCF10A cells, only 0.5 μM Danu for 72 hours increased the ratio of p-Erk1/2 over total Erk1/2 from 1.69 at basal level to 2.09 (P<0.05 by one-way ANOVA; Figures 11A and 12A). These observations indicate the involvement of p38 MAPK and Erk1/2 in Danu-induced autophagy.
Effects of p38 MAPK and Erk1/2 activation on Danu-induced autophagy
Recent studies have demonstrated the involvement of p38 MAPK and Erk1/2 signaling pathways in autophagy. In our study, treatment with Danu for more than 24 hours resulted in the sustained activation of p38 MAPK and Erk1/2, and significantly increased expression of LC3-II and beclin 1. Thus, to further investigate the roles of p38 MAPK and Erk1/2 in Danu-induced autophagy, we used the autophagy inhibitor WM, an inhibitor for the sequestration step of autophagosomes, and bafilomycin A1, an inhibitor for the maturation step of autophagosomes. First, we examined the effects of autophagy inhibitors on Danu-induced autophagy in MCF7 and MDA-MB-231 cells using flow cytometry. In MCF7 cells, WM at 10 μM decreased Danu-induced autophagy by 31.3%; however, bafilomycin A1 at 100 nM increased Danu-induced autophagy 1.2-fold, compared to the control cells treated with Danu alone (P<0.05 by one-way ANOVA; Figure 13A and B). In MDA-MB-231 cells, WM at 10 μM also decreased Danu-induced autophagy by 45.4%; however, bafilomycin A1 at 100 nM increased Danu-induced autophagy 1.3-fold, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 13A and B).
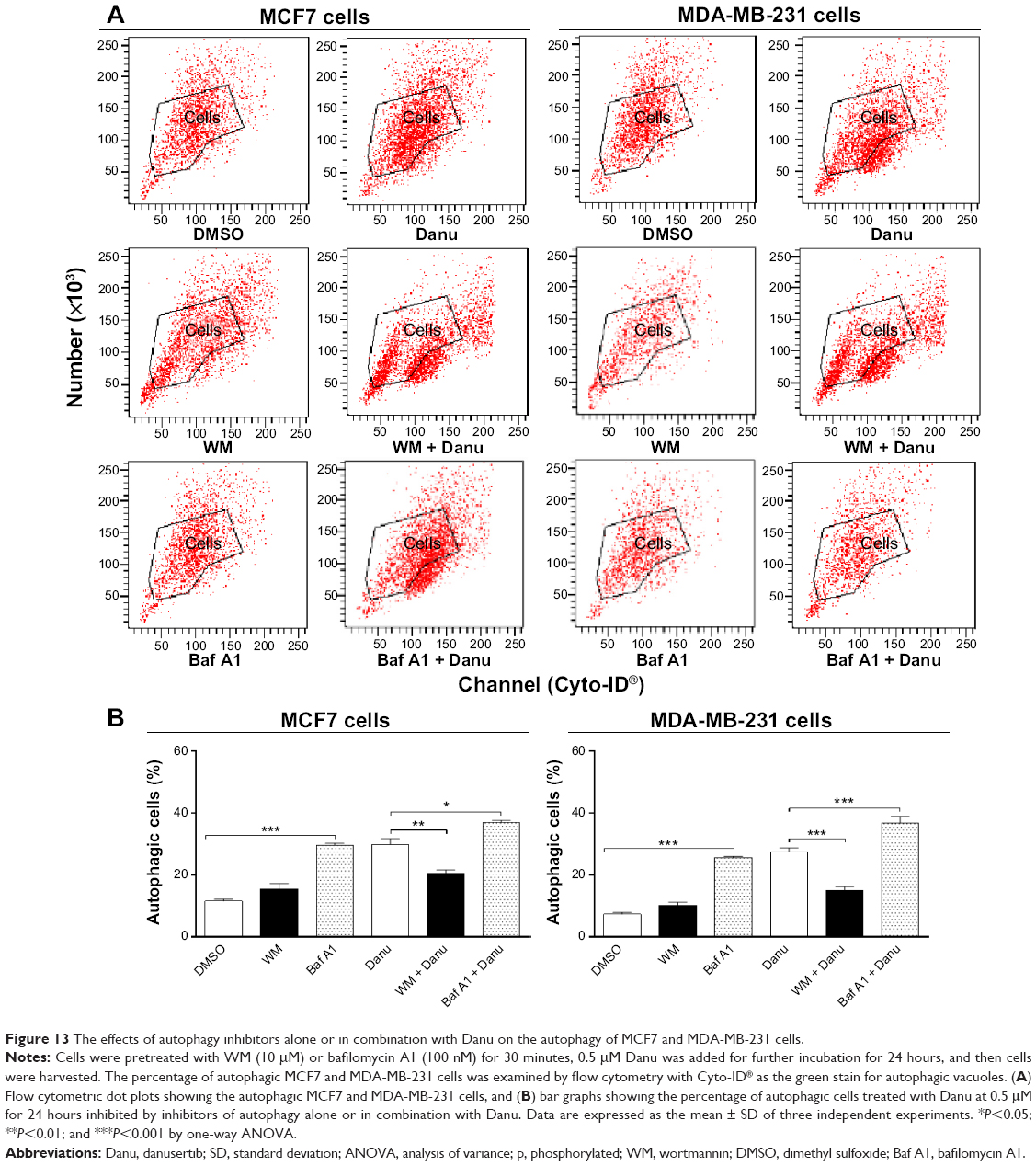 | Figure 13 The effects of autophagy inhibitors alone or in combination with Danu on the autophagy of MCF7 and MDA-MB-231 cells. |
Furthermore, we examined the expression levels of beclin 1 and LC3-II when MCF7 and MDA-MB-231 cells were treated with Danu alone or in combination with autophagy inhibitors, by Western blotting assay. Treatment of cells with 0.5 μM Danu in combination with 10 μM WM or 100 nM bafilomycin A1 decreased the expression level of beclin 1 by 83.12% and 66.9% in MCF7 cells, respectively, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 14A and B). However, in MDA-MB-231 cells, incubation of cells with 0.5 μM Danu in combination with 10 μM WM or 100 nM bafilomycin A1 did not significantly change the level of beclin 1 (P>0.05 by one-way ANOVA; Figure 14A and B). Treating cells with 0.5 μM Danu in combination with 10 μM WM decreased the level of LC3-II by 96.2% in MCF7 cells (P<0.01 by one-way ANOVA; Figure 14A and B). In MDA-MB-231 cells, treatment of cells with 0.5 μM Danu in combination with 10 μM WM decreased the level of LC3-II by 25.0%, but there was no significant difference compared to the control cells treated with Danu alone (P>0.05 by one-way ANOVA; Figure 14A and B). Danu (0.5 μM) combined with 100 nM bafilomycin A1 increased the expression level of LC3-II in MCF7 cells and MDA-MB-231 cells by 3.9- and 12.2-fold, respectively, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 14A and B).
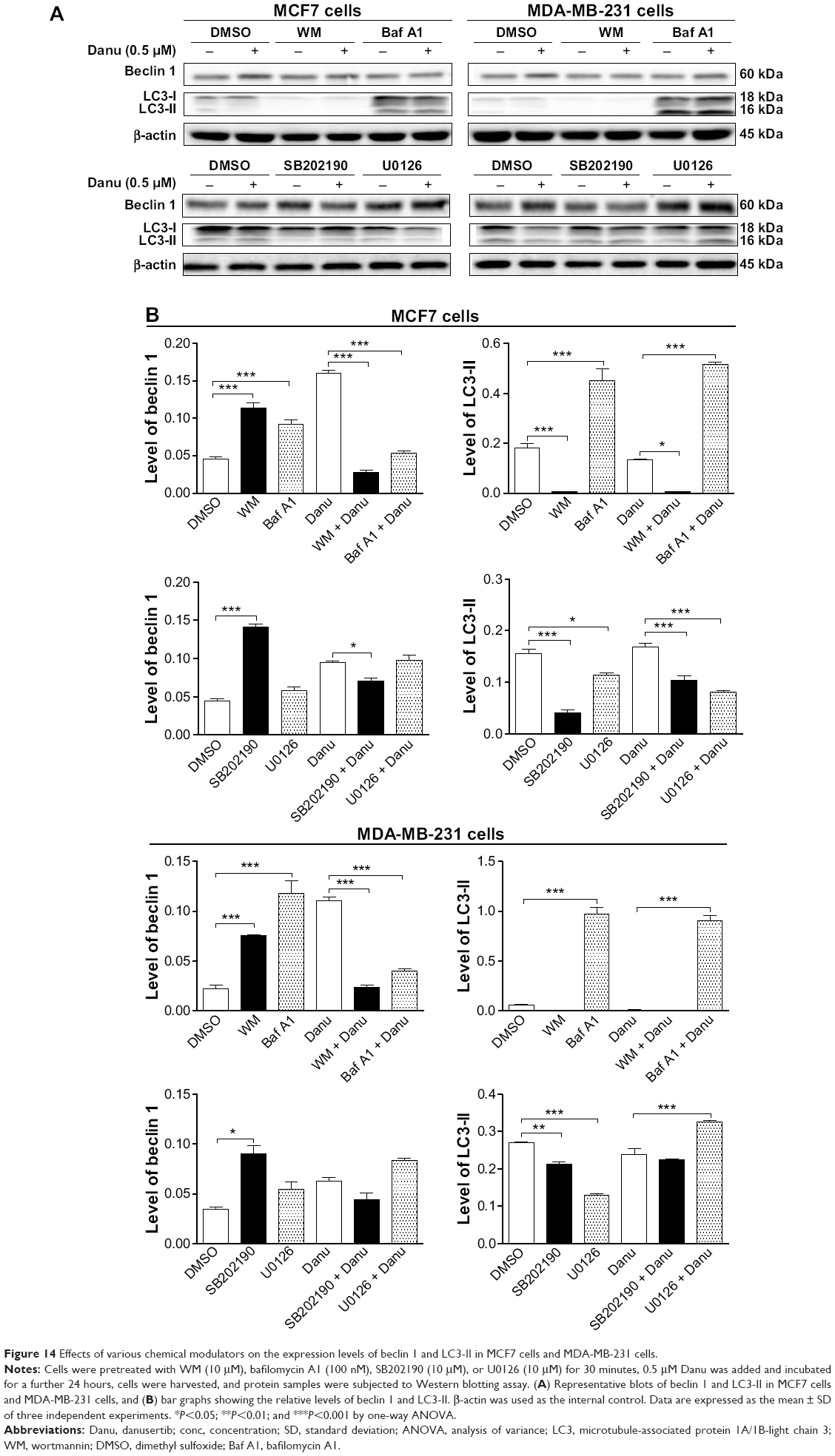 | Figure 14 Effects of various chemical modulators on the expression levels of beclin 1 and LC3-II in MCF7 cells and MDA-MB-231 cells. |
In addition, we examined the activation of p38 MAPK at Thr180/Tyr182 and Erk1/2 at Thr202/Tyr204 when cells were treated with WM and bafilomycin A1 prior to Danu treatment. In MCF7 and MDA-MB-231 cells, pretreatment with WM prior to Danu markedly attenuated the phosphorylation of p38 MAPK and Erk1/2 compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 14A and B). Moreover, pretreatment with bafilomycin A1 prior to Danu decreased the ratio of p-p38 MAPK over total p38 MAPK, and p-Erk1/2 over total Erk1/2 in MCF7 cells, compared to the control cells treated with Danu alone (P<0.05 by one-way ANOVA; Figure 15A and B). In MDA-MB-231 cells, there was no significant difference (P>0.05 by one-way ANOVA; Figure 15A and B). Taken together, these results show that pretreatment with WM inhibits Danu-induced autophagy, but pretreatment with bafilomycin A1 markedly increases Danu-induced autophagy. These observations suggest that p38 MAPK and Erk1/2 regulate autophagosome formation at the sequestration step.
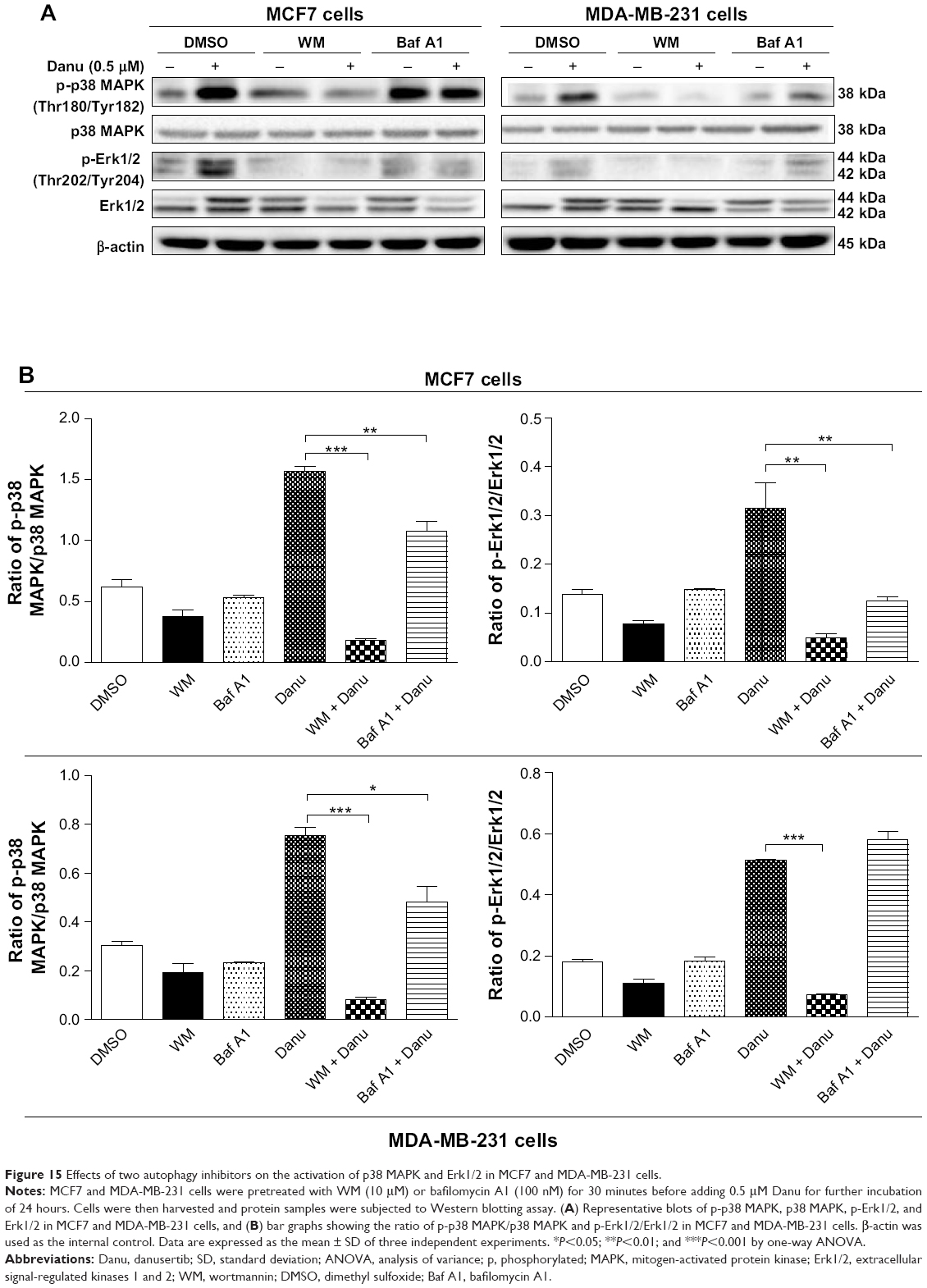 | Figure 15 Effects of two autophagy inhibitors on the activation of p38 MAPK and Erk1/2 in MCF7 and MDA-MB-231 cells. |
p38 MAPK regulates Danu-induced autophagy via Akt activation
We further dissected the roles of p38 MAPK and Erk1/2 in Danu-induced autophagy. Cells were pretreated with SB202190 (a p38 MAPK inhibitor) or U0126 (an Erk1/2 inhibitor) for 30 minutes, and then treated with Danu for a further 24 hours. First, we examined the effects of SB202190 and U0126 on Danu-induced autophagy in MCF7 and MDA-MB-231 cells using flow cytometry. In MCF7 cells and MDA-MB-231 cells, 10 μM SB202190 decreased Danu-induced autophagy; however, there was no significant difference compared to the control cells treated with Danu alone (P>0.05 by one-way ANOVA; Figure 16A and B). Ten μM U0126 significantly decreased Danu-induced autophagy by 32.7% and 32.4% in MCF7 and MDA-MB-231 cells, respectively, compared to the control cells treated with Danu alone (P<0.01 by one-way ANOVA; Figure 16A and B). Next, we examined the expression levels of beclin 1 and LC3-II when MCF7 and MDA-MB-231 cells were treated with Danu alone or in combination with SB202190 or U0126, by Western blotting assay. Treatment of cells with 0.5 μM Danu in combination with 10 μM SB202190 remarkably decreased the expression level of beclin 1 by 25.5% in MCF7 cells (P<0.05 by one-way ANOVA; Figure 16A and B); however, there was no significant change in MDA-MB-231 cells, compared to the control cells treated with Danu alone (P>0.05 by one-way ANOVA; Figure 16A and B). Treating cells with 0.5 μM Danu plus 10 μM U0126 did not significantly change the expression level of beclin 1 in MCF7 and MDA-MB-231 cells (P>0.05 by one-way ANOVA; Figure 16A and B). Moreover, incubation of cells with 0.5 μM Danu plus 10 μM SB202190 significantly decreased the level of LC3-II by 38.7% in MCF7 cells (P<0.01 by one-way ANOVA; Figure 16A and B), there was no significant change in MDA-MB-231 cells, compared to the control cells treated with Danu alone (P>0.05 by one-way ANOVA; Figure 16A and B). Treatment of cells with 0.5 μM Danu plus 10 μM U0126 decreased the level of LC3-II by 52.4% but increased the level of LC3-II 1.4-fold in MCF7 cells, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 16A and B).
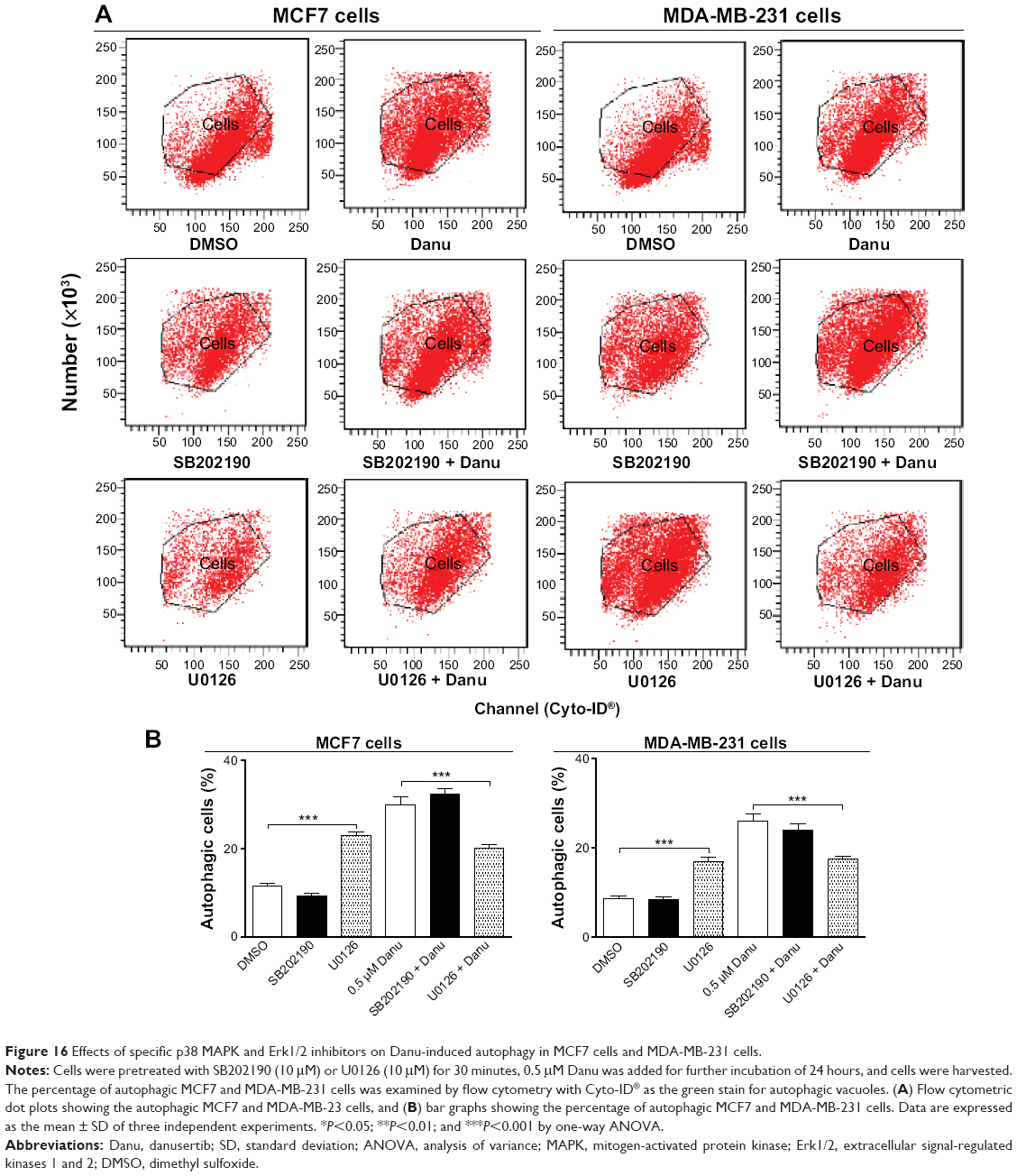 | Figure 16 Effects of specific p38 MAPK and Erk1/2 inhibitors on Danu-induced autophagy in MCF7 cells and MDA-MB-231 cells. |
Furthermore, SB202190 completely blocked p38 MAPK activation, and U0126 completely blocked Erk1/2 activation. Treatment of cells with 0.5 μM Danu plus 10 μM SB202190 significantly decreased the ratio of p-p38 MAPK over total p38 MAPK by 92.2% in MCF7 cells (P<0.001 by one-way ANOVA; Figure 17A and B), and there was a 52.9% decrease in the ratio of p-p38 MAPK over total p38 MAPK in MDA-MB-231 cells, compared to the control cells treated with Danu alone (P<0.01 by one-way ANOVA; Figure 17A and B). Treating cells with 0.5 μM Danu plus 10 μM U0126 significantly increased the ratio of p-p38 MAPK/p38 MAPK 2.2-fold in MCF7 cells (P<0.001 by one-way ANOVA; Figure 17A and B), and there was a 52.9% decrease in the ratio of p-p38 MAPK over p38 MAPK in MDA-MB-231 cells, compared to the control cells treated with Danu alone (P<0.01 by one-way ANOVA; Figure 17A and B). Moreover, treatment of cells with 0.5 μM Danu plus 10 μM SB202190 increased the ratio of p-Erk1/2 over total Erk1/2 2.6-fold in both MCF7 and MDA-MB-231 cells, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 17A and B). Incubation of cells with 0.5 μM Danu in combination with 10 μM U0126 significantly decreased the ratio of p-Erk1/2 over total Erk1/2 81.6% and 69.7% in MCF7 cells and MDA-MB-231 cells, respectively, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 17A and B). Treatment of cells with 10 μM SB202190 alone significantly decreased the ratio of p-Akt at Ser473 over Akt 96.4% and 91.1% in MCF7 cells and MDA-MB-231 cells, respectively, compared to the control cells treated with DMSO only (P<0.001 by one-way ANOVA; Figure 17A and B); exposure of cells to 10 μM U0126 significantly decreased the p-Akt/Akt ratio 85.4% and 75.5% in MCF7 cells and MDA-MB-231 cells, respectively, compared to the control cells treated with DMSO only (P<0.001 by one-way ANOVA; Figure 17A and B). However, there was no significant change in the p-Akt/Akt ratio when MCF7 cells and MDA-MB-231 cells were treated with 0.5 μM Danu plus 10 μM SB202190, and there was also no significant change when MCF7 cells were treated with 0.5 μM Danu plus 10 μM U0126, compared to the control cells treated with Danu alone (P>0.05 by one-way ANOVA; Figure 17A and B). However, treatment of cells with 0.5 μM Danu combined with 10 μM U0126 significantly increased the ratio of p-Akt over total Akt in MDA-MB-231 cells by 2.1-fold, compared to the control cells treated with Danu alone (P<0.001 by one-way ANOVA; Figure 17A and B).
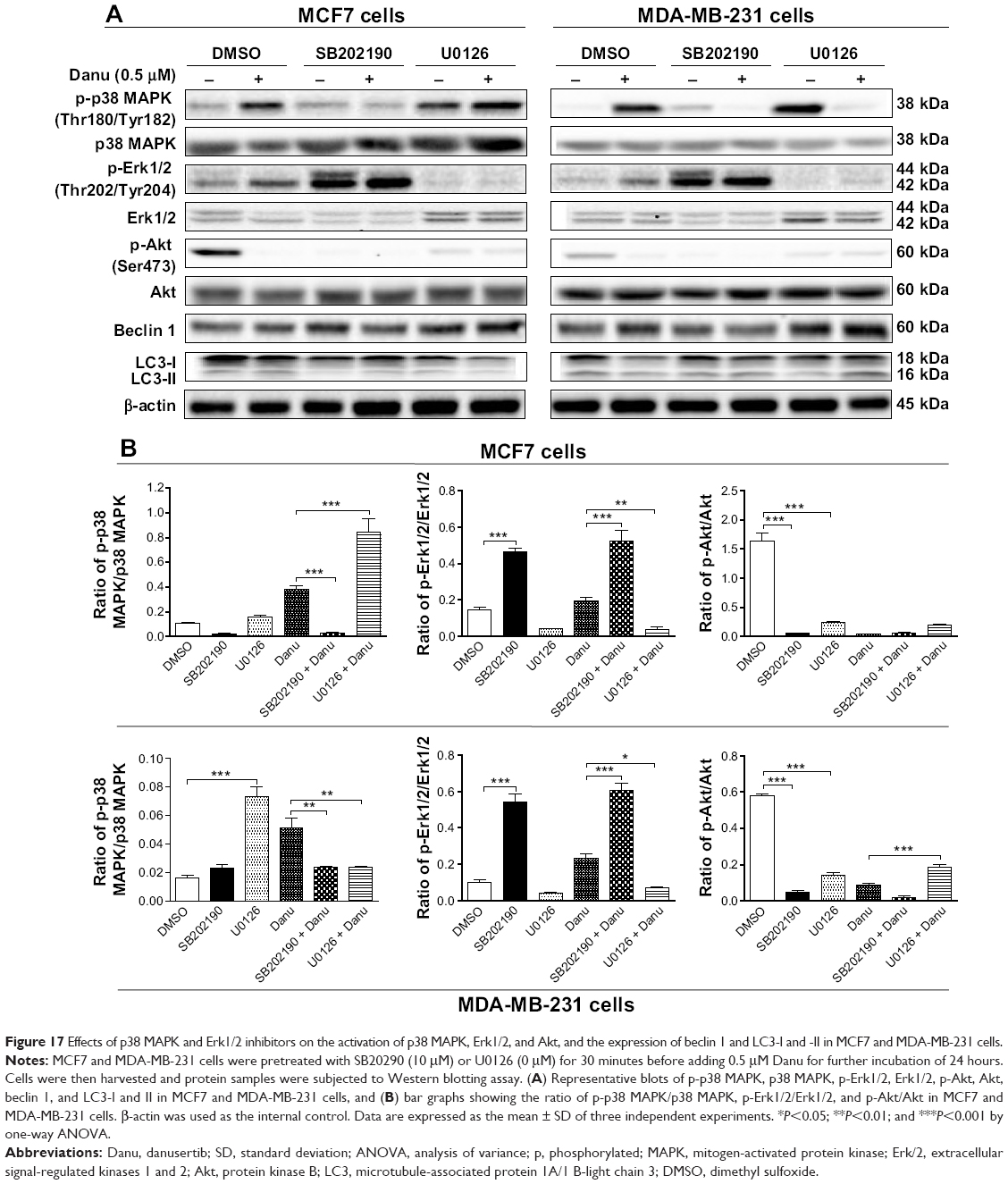 | Figure 17 Effects of p38 MAPK and Erk1/2 inhibitors on the activation of p38 MAPK, Erk1/2, and Akt, and the expression of beclin 1 and LC3-I and -II in MCF7 and MDA-MB-231 cells. |
To further confirm the roles of p38 MAPK and Erk1/2 in Danu-induced autophagy, p38 MAPK- and Erk1/2-specific siRNAs were used to knock down the gene expression of p38 MAPK or Erk1/2. Transfection of MCF-7 cells with p38 MAPK siRNA downregulated the level of Danu-induced phosphorylation of p38 MAPK, and increased LC3-II level compared with parental or nonspecific siRNA-transfected control cells. Compared to MCF-7 cells transfected with control siRNA, transfecting p38 MAPK siRNA significantly decreased by 50% in the p-p38 MAPK/p38 MAPK ratio, and transfecting Erk1/2 siRNA significantly decreased by 85.6% in the ratio of p-Erk1/2 over total Erk1/2 (P<0.001 by one-way ANOVA; Figure 18A and B). After transfection of MCF-7 cells with p38 MAPK siRNA, we found that the ratio of p-Akt over total Akt and the level of LC3-II were increased 2.2- and 3.6-fold compared to transfecting control siRNA, respectively (P<0.001 by one-way ANOVA; Figure 18A and B); however, there were no significant changes in the p-Erk1/2:Erk1/2 ratio and the level of beclin 1 compared to transfecting control siRNA (P>0.05 by one-way ANOVA; Figure 18A and B). After transfection of MCF-7 cells with Erk1/2 siRNA, we observed that the level of LC3-II was increased 1.5-fold compared to the cells transfected with the control siRNA (P<0.001 by one-way ANOVA; Figure 18A and B); however, there were no significant changes in the ratios of p-p38 MAPK over total p38 MAPK, and p-Akt over total Akt, and in the level of beclin 1 compared to the cells transfected with the control siRNA (P>0.05 by one-way ANOVA; Figure 18A and B).
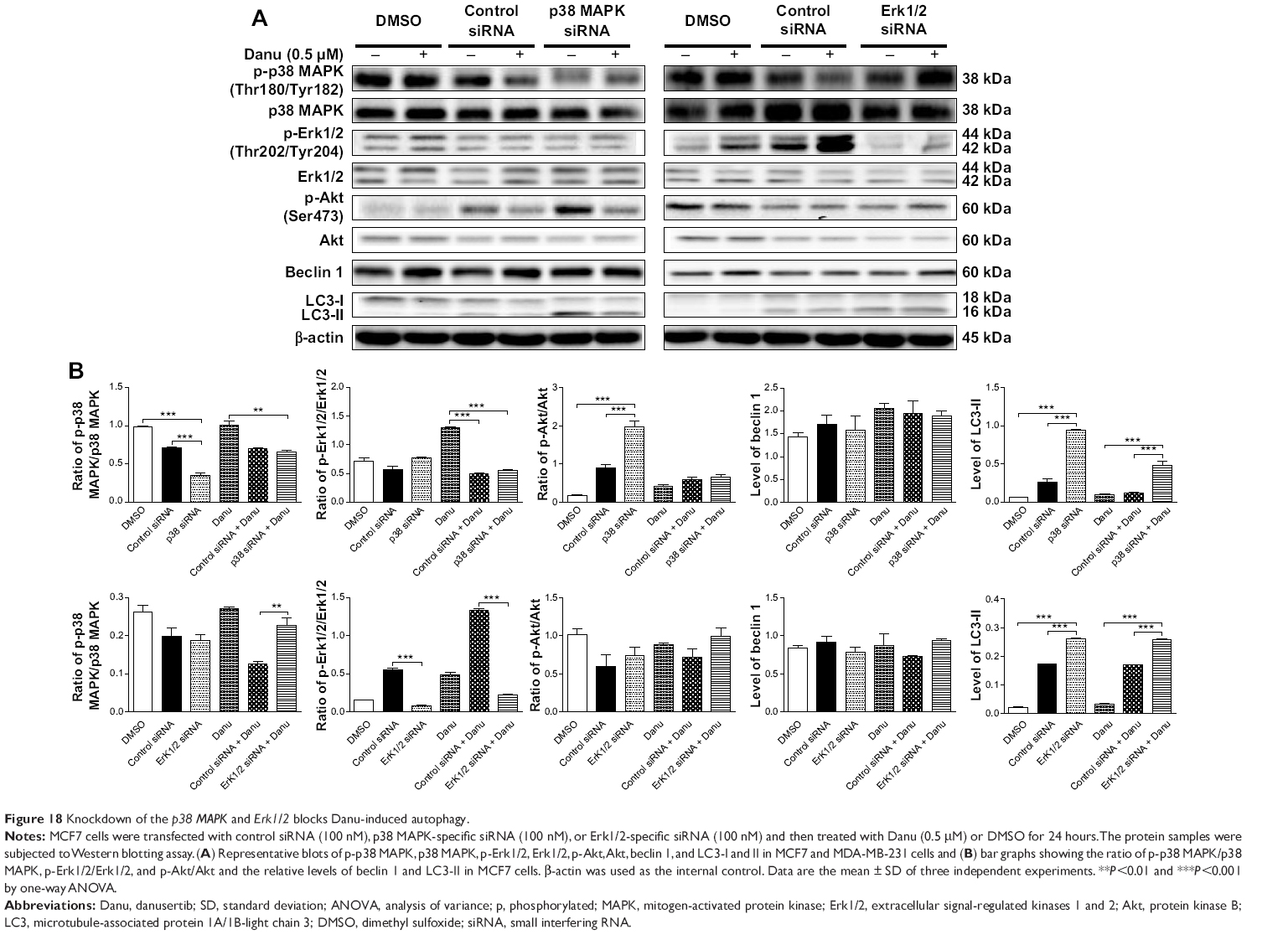 | Figure 18 Knockdown of the p38 MAPK and Erk1/2 blocks Danu-induced autophagy. |
Furthermore, after transfection of MCF-7 cells with p38 MAPK siRNA and 0.5 μM Danu treatment, we found that the level of LC3-II was markedly increased 3.9-fold compared to Danu treated siRNA transfection cells (P<0.001 by one-way ANOVA; Figure 18A and B); however, there were no significant changes in the ratios of p-p38 MAPK over total p38 MAPK, p-Erk1/2 over total Erk1/2, and p-Akt over total Akt, and no significant change in the level of beclin 1 compared to transfecting control siRNA with 0.5 μM Danu (P>0.05 by one-way ANOVA; Figure 18A and B). After transfection of MCF-7 cells with Erk1/2 siRNA transfection cells treated with 0.5 μM Danu, we observed that the ratio of p-p38 MAPK over total p38 MAPK and the level of LC3-II were increased 1.8- and 1.5-fold, respectively, compared to transfecting control siRNA with 0.5 μM Danu (P<0.001 by one-way ANOVA; Figure 18A and B); however, there were no significant changes in the ratio of p-Akt over total Akt and the level of beclin 1, compared to transfecting control siRNA (P>0.05 by one-way ANOVA; Figure 18A and B); in addition, the ratio of p-Erk1/2 over total Erk1/2 was decreased by 83.5% compared to transfecting control siRNA (P<0.001 by one-way ANOVA; Figure 18A and B). These results have further confirmed that p38 MAPK and Erk1/2 signaling pathways are involved in Danu-induced autophagy.
Danu inhibits EMT in MCF7 and MDA-MB-231 cells
EMT is a critical process involving the initiation, growth, invasion, and metastasis of cancer.24 Herein, we examined the effect of Danu treatment on EMT-associated markers in MCF7 and MDA-MB-231 cells using the Western blotting assay. As shown in Figure 19A and B, incubation of MCF-7 cells with Danu resulted in a remarkable increase in the expression level of E-cadherin and a decrease in the expression level of N-cadherin. There was a 1.3-fold increase in the expression of E-cadherin when treated with 0.5 μM Danu for 24 hours, whereas 0.01, 0.1, and 0.5 μM Danu suppressed the expression level of N-cadherin 41.1%, 58.9%, and 56.1%, respectively (P<0.001 by one-way ANOVA; Figure 19A and B). Consequently, with increasing concentrations of PLB, an increased ratio of E-cadherin over N-cadherin was observed. The E-cadherin/N-cadherin ratio was increased from 0.37 at basal level to 0.77 and 1.13, when MCF7 cells were treated with Danu at 0.1 and 0.5 μM Danu for 24 hours, respectively (P<0.001 by one-way ANOVA; Figure 19A and B). In MDA-MB-231 cells, there was a 1.5-fold increase in the expression of E-cadherin when cells were treated with 0.5 μM Danu for 24 hours. Meanwhile, 0.1 and 0.5 μM Danu decreased the expression level of N-cadherin by 32.3% and 25.8%, respectively. The E-cadherin/N-cadherin ratio was increased from 1.5 to 2.8 and 3.0, when MDA-MB-231 cells were treated with Danu at 0.1 and 0.5 μM for 24 hours, respectively (P<0.01 by one-way ANOVA; Figure 19A and B).
In order to further examine the effect of Danu on EMT in MCF7 and MDA-MB-231 cells, we tested the expression levels of several key regulators of E-cadherin. Snail and slug (both zinc finger transcriptional factors), together with TCF8/ZEB1, are suppressors of E-cadherin in EMT.26 In addition, snail blocks the cell cycle and confers resistance to cell death, and slug protects damaged cells from apoptosis by repressing p53-induced transcription of the pro-apoptotic Bcl-2 family protein PUMA. Danu significantly reduced the expression level of snail and slug in both cell lines. In MCF7 cells, 24-hour Danu treatment at 0.01, 0.1, and 0.5 μM significantly suppressed the expression level of snail by 35.9%, 50.5%, and 76.6%, and inhibited the expression of slug by 13.9%, 45.2%, and 72.3%, respectively (P<0.05 by one-way ANOVA; Figure 19A and B). In MDA-MB-231 cells, treatment with 0.01, 0.1, and 0.5 μM Danu suppressed the expression level of snail by 19.1%, 48.3%, and 44.8%, respectively. Treatment of cells with 0.1 and 0.5 μM Danu for 24 hours reduced expression level of slug by 27.9% and 35.1%, respectively (P<0.01 by one-way ANOVA; Figure 19A and B). Furthermore, Danu induced a marked reduction in the expression level of TCF8/ZEB1 in MCF7 and MDA-MB-231 cells. In MCF7 cells, 0.1 and 0.5 μM Danu significantly suppressed the expression level of TCF8/ZEB1 by 14% and 29%, respectively. Similarly, there was a 31.6%, 52.6%, and 42.1% reduction in the expression of TCF8/ZEB1 in MDA-MB-231 cells treated with 0.01, 0.1, and 0.5 μM Danu, respectively (P<0.05 by one-way ANOVA; Figure 19A and B). Vimentin is a type III intermediate filament protein that is expressed in mesenchymal cells. β-catenin can act as an integral component of a protein complex in adherens junctions, which help cells maintain epithelial layers, and β-catenin participates in the Wnt signaling pathway as a downstream target.26 Treatment of cells with 0.01, 0.1, and 0.5 μM Danu significantly suppressed the expression level of vimentin by 25.0%, 40.0%, and 55.0%, respectively, in MCF7 cells. Danu at 0.1 and 0.5 μM reduced vimentin level by 28.4% and 49.0%, respectively, in MDA-MB-231 cells (P<0.01 by one-way ANOVA; Figure 19A and B). There was a significant reduction in the expression level of β-catenin in both cell lines treated with Danu. Treatment of cells with Danu at 0.01, 0.1, and 0.5 μM decreased the expression level of β-catenin by 35.6%, 40.1%, and 43.9% in MCF7 cells, respectively, and 0.1 and 0.5 μM Danu reduced β-catenin expression by 22.4% and 25.8% in MDA-MB-231 cells, respectively (P<0.01 by one-way ANOVA; Figure 19A and B). Finally, the expression of ZO-1 was examined in MCF7 and MDA-MB-231 cells exposed to Danu. Treatment of MCF7 cells with 0.1 and 0.5 μM Danu for 24 hours increased the expression of ZO-1 1.5- and 1.3-fold, respectively, and treatment with 0.01, 0.1, and 0.5 μM Danu resulted in a 1.4-, 1.6-, and 1.9-fold increase in the expression level of ZO-1 in MDA-MB-231 cells, respectively (P<0.05 by one-way ANOVA; Figure 19A and B).
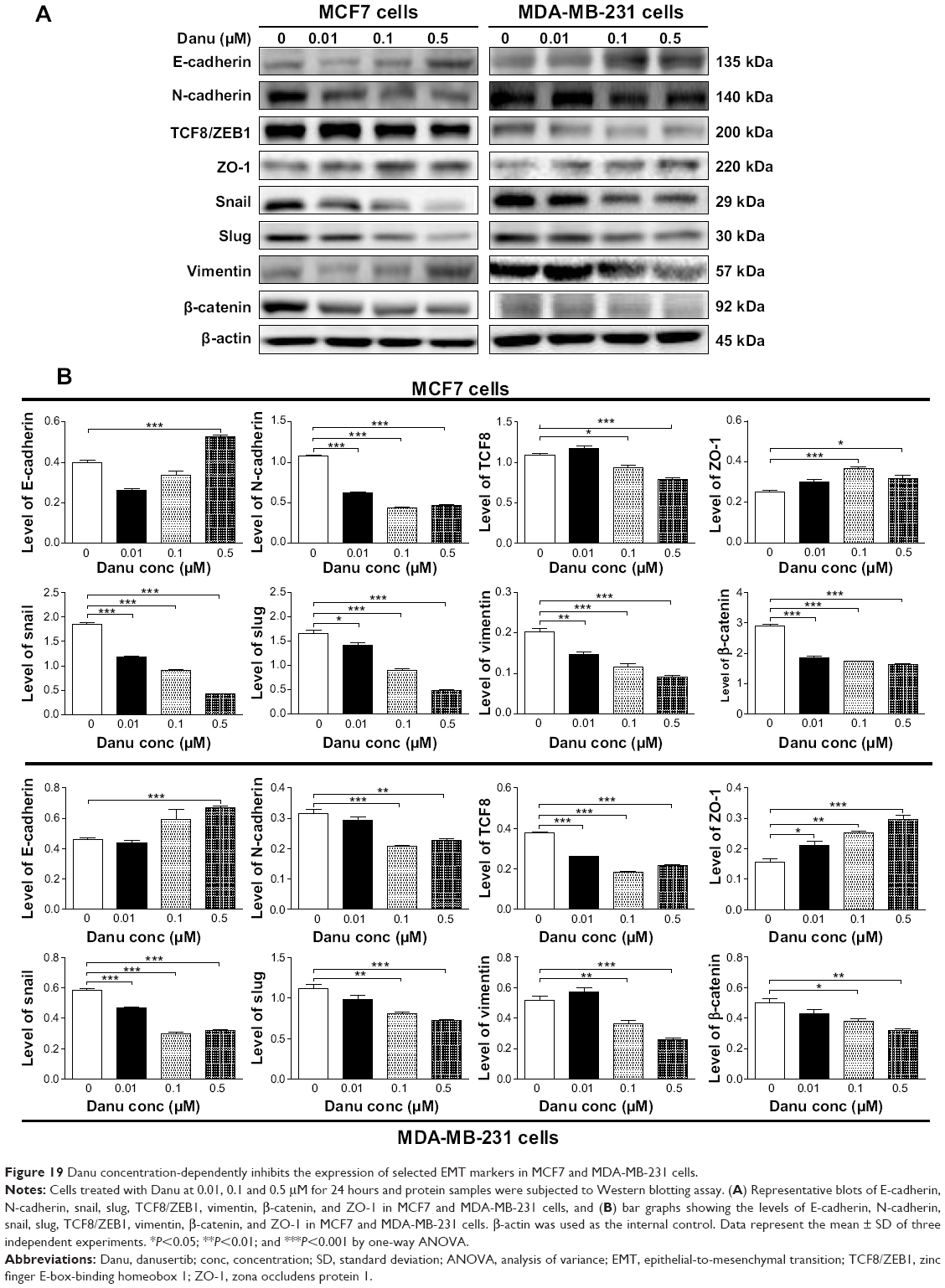 | Figure 19 Danu concentration-dependently inhibits the expression of selected EMT markers in MCF7 and MDA-MB-231 cells. |
Furthermore, we examined the effect of Danu on the expression of selected EMT markers in MCF7 and MDA-MB-231 cells over 72 hours. There was a significantly inhibitory effect of Danu on EMT in both cells. In comparison to the control cells, treatment of MCF7 cells with 0.5 μM Danu for 4, 8, 12, 24, 48, and 72 hours increased the expression of E-cadherin by 1.7-, 2.2-, 3.2-, 2.8-, 2.6-, and 2.0-fold, respectively, while the expression of N-cadherin was decreased by 40.2% and 44.1% after 48 and 72 hours, respectively, which in turn led to a significant increase in the ratio of E-cadherin over N-cadherin (P<0.05 by one-way ANOVA; Figure 20A and B). The expression of snail was significantly decreased by 41.3% with the 0.5 μM Danu treatment for 72 hours, and the expression of slug was reduced by 37.5%, 44.0%, 54.2%, and 68.9% with the 0.5 μM Danu treatment for 12, 24, 48, and 72 hours, respectively (P<0.05 by one-way ANOVA; Figure 20A and B). The expression of vimentin was significantly decreased 29.1%, 39.9%, 36.8%, and 38.4% with the 0.5 μM Danu treatment for 4, 24, 48, and 72 hours, respectively. Moreover, the expression of β-catenin was reduced by 60.0%, 71.8%, and 59.7% with the 0.5 μM Danu treatment for 24, 48, and 72 hours, respectively (P<0.05 by one-way ANOVA; Figure 20A and B). The expression of TCF8/ZEB1 was significantly decreased by 22.6%, 23.5%, 45.5%, and 81.7% with the 0.5 μM Danu treatment for 4, 24, 48, and 72 hours, respectively. However, the expression of ZO-1 was increased by 1.6- and 2.4-fold with the 0.5 μM Danu treatment for 48 and 72 hours, respectively (P<0.05 by one-way ANOVA; Figure 20A and B). In MDA-MB-231 cells, incubation with 0.5 μM Danu for 12 and 72 hours led to a 1.6- and 1.5-fold increase in the expression of E-cadherin, respectively; and incubation of 0.5 μM Danu for 4, 24, 48, and 72 hours resulted in a 40.8%, 50.5%, 43.0%, and 61.8% reduction in the expression of N-cadherin compared to the control cells. Consequently, it led to an increase in the ratio of E-cadherin over N-cadherin. The expression of snail was decreased by 25.2%, 72.3%, and 84.3%, respectively, with the 0.5 μM Danu treatment for 24, 48, and 72 hours (P<0.05; by one-way ANOVA Figure 20A and B). However, the expression of slug did not significantly change with the 0.5 μM Danu treatment over 72 hours (P>0.05 by one-way ANOVA; Figure 20A and B). The expression of vimentin was significantly decreased by 42.6%, 65.7%, and 62.8% with the 0.5 μM Danu treatment for 4, 8, and 12 hours, respectively. The expression of β-catenin was reduced by 41.4%, 47.2%, and 59.3% with the 0.5 μM Danu treatment for 12, 24, and 72 hours, respectively (P<0.05 by one-way ANOVA; Figure 20A and B). The expression of TCF8/ZEB1 was significantly decreased by 70.4%, 74.8%, 74.9%, and 86.0% with the 0.5 μM Danu treatment for 8, 24, 48, and 72 hours, respectively. The expression of ZO-1 was increased by 2.9- and 2.3-fold with the 0.5 μM Danu treatment for 48 and 72 hours, respectively (P<0.05 by one-way ANOVA; Figure 20A and B). Taken together, Danu exhibits a potent inhibitory effect on EMT in MCF7 and MDA-MB-231 cells.
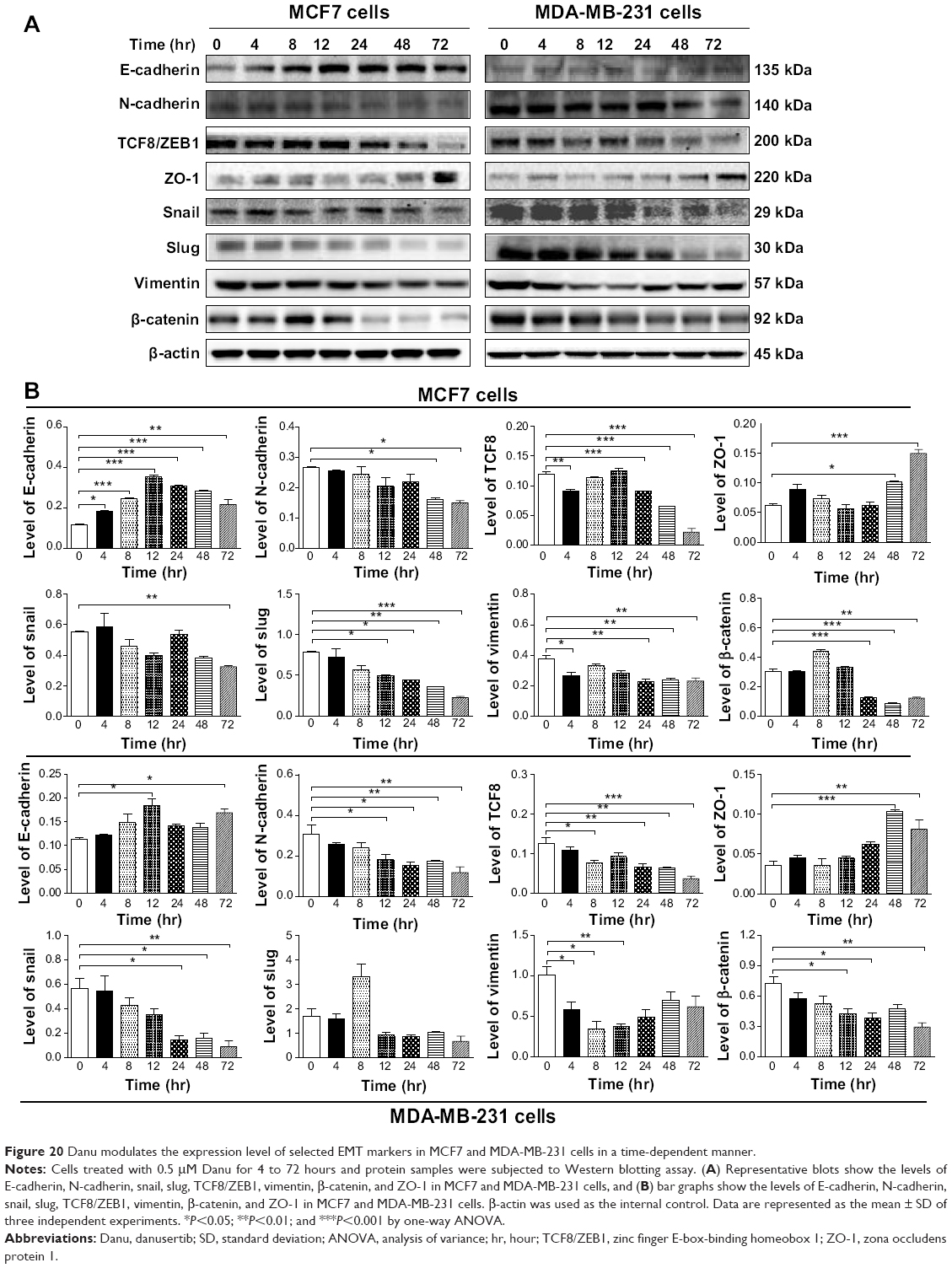 | Figure 20 Danu modulates the expression level of selected EMT markers in MCF7 and MDA-MB-231 cells in a time-dependent manner. |
Discussion
Several Aurora kinase inhibitors have been tested in breast cancer preclinically in the last couple of years, indicating Aurora kinases are promising therapeutic targets for breast cancer. In this study, we investigated the anti-proliferative and anti-invasive effects of Danu on nonmalignant and malignant breast cell lines. The Aurora family comprises three related kinases (AURKA/B/C) that share the highest degree of sequence homology in their catalytic domains.27,28 Overexpression of Aurora kinases has been demonstrated in a variety of human malignancies including that of breast cancer, and the overexpression has been shown to induce chromosomal instability, resulting in aneuploidy.29,30 Notably, aneuploidy is the salient feature of most cancer cells and is closely correlated with mammalian cell transformation.31 Recently, there are a number of Aurora kinase inhibitors that have been discovered and are under preclinical or clinical development at different stages. Danu, a small adenosine triphosphate competitive molecule that inhibits AURKA/B/C, has entered Phase II clinical trials for patients with solid tumors.19 Danu has strong activity on AURKB, as determined by inhibition of its substrate histone H3 in position Ser10 and also on AURKA, as determined by inhibition of autophosphorylation in position Thr288.32,33 It has been shown that Danu inhibits the proliferation of hepatocellular carcinoma and pancreatic cancer cells via inducing cell division failure and resulting in polyploidy.34,35 In the present study, our results show that Danu inhibits proliferation of cells and induces G2/M arrest, apoptosis, and autophagy in nonmalignant and malignant breast cell lines via p38 MAPK/Erk1/2–Akt/mTOR mediated signaling pathways. Additionally, Danu also inhibits EMT in malignant breast cell lines.
Cell cycle control is a dominant regulatory mechanism in cell growth, involving a number of functional proteins and related signaling pathways, including CDK1 and 2, cyclin B1, p21 Wafl/Cip1, and p27 Kipl. Targeting the cell cycle has been proposed to be a promising strategy for cancer treatment, and many cytotoxic agents and/or DNA-damaging agents have been developed which are able to arrest the cell cycle at the G0/G1, S, or G2/M phase, and then induce cell apoptosis.36 G2/M transition is controlled by CDC2, also known as CDK1, which partners with cyclin B1. CDC family proteins and cyclin complexes play a critical role in the regulation of G2/M transition. G2/M arrest is closely regulated via the activation and deactivation of CDC family proteins and cyclin complexes. In the present study, we found that Danu concentration-dependently arrested MCF10A, MCF7, and MDA-MB-231 cells in the G2/M phase. We further explored the effect of Danu on the key regulators in cell cycle checkpoints including CDC2/CDK1, CDK2, and cyclin B1 in all cells. The CDC2–cyclin B1 complex is pivotal in regulating the G2/M phase transition and mitosis. We observed a significant decrease in the expression level of cyclin B1 and CDC2 in MCF10A, MCF7, and MDA-MB-231 cells treated with Danu, providing an explanation for the effect of Danu on G2/M phase arrest in MCF10A, MCF7, and MDA-MB-231 cells.
Furthermore, cell cycle progression is also tightly regulated by the relative balance between the cellular concentrations of cyclin-dependent kinase inhibitors, such as the Cip/Kip family. The Cip/Kip family, including p21 Waf1/Cip1 and p27 Kipl, binds to cyclin–CDK complexes, preventing the kinase activation and blocking the progression of the cell cycle at the G2/M phases.37 Moreover, it has been reported that p21 Waf1/Cip1, a cyclin-dependent kinase inhibitor regulated by p53, can bind to the CDK1/CDC2–cyclin B1 complex, thereby inducing cell cycle arrest. We observed that the expression of p53, p27 Kip1, and p21 Waf1/Cip1 was concentration- and time-dependently increased in MCF10A, MCF7, and MDA-MB-231 cells treated with Danu, which probably contributes to the inhibitory effect of Danu on cell proliferation, and the inducing effect on cell cycle arrest in nonmalignant and malignant breast cells. These results indicate that upregulation of p53, p21 Waf1/Cip1, and p27 Kip1 expression, and the suppression of CDK2 and cyclin B1 by Danu may result in G2/M phase arrest in human breast cells.
Apoptosis not only plays a crucial role in tissue development and homeostasis but is also involved in a wide range of pathologies.38 Apoptotic cells have morphologically defined nuclear features of chromatin condensation and DNA fragmentation.39 The apoptotic process occurs via two distinct signaling pathways, known as the tumor necrosis factor/Fas-receptor, and mitochondria-mediated pathways, with the involvement of the caspase family, including caspases 3, 6, 7, 8, and 9.40,41 In the present study, we observed a concentration-dependent apoptosis induced by Danu in MCF10A, MCF7, and MDA-MB-231 cells. Anti-apoptotic members of Bcl-2 can be inhibited by post-translational modification and/or by increased expression of PUMA, which is an essential regulator of p53-mediated cell apoptosis.42 In our study, we found that the expression level of cleaved caspase 9 was significantly increased after Danu treatment, which subsequently activated caspase 3. Activated caspase 3, in turn, induced apoptosis with a decrease in Bcl-2 level. Moreover, we noted a concentration-dependent increase in the expression level of PUMA and Bax in MCF10A, MCF7, and MDA-MB-231 cells. These results indicate that Danu induces mitochondria-dependent apoptosis in nonmalignant and malignant breast cells.
Autophagy is a primary, morphologically distinctive mode of programmed cell death, which is an important process involving the engulfment and degradation of nonessential or abnormal cellular organelles and proteins in living cells.43 The role of autophagy in cancer cell death is still controversial. Genetic evidence indicates that autophagy can suppress tumor growth, but it also can support the survival of established tumors under stress conditions.44,45 However, increasing evidence shows that targeting autophagy is a promising strategy for the treatment of breast cancer,46–49 and induction of autophagy has been tested for a potential therapeutic strategy to treat cancer.50 Beclin 1 and LC3 are two specific markers of cell autophagy, and both of them are strongly involved in the autophagic process, especially in its early stages. Beclin 1, initially isolated as a Bcl-2-binding protein, was the first human protein is shown to be indispensable for autophagy. Beclin 1 and Vps34 (class III PI3K) form a complex in charge of autophagic nucleation in mammals. Beclin 1 promotes autophagy and inhibits proliferation of cancer cells by forming and activating the autophagy-promoting complex beclin 1–Vps34, and the suppression of beclin 1 expression impairs autophagy.51 LC3, associated with the formation of the autophagic vacuole, is currently considered as a specific molecular marker for autophagosomes in mammals. LC3 proteins can be divided into two forms: LC3-I (18 kDa) and LC3-II (16 kDa). The amount of LC3-II correlates with the extent of autophagosome formation, and it is an autophagosomal marker.52 In our study, the induction of autophagy was supported by the following observations. First, using flow cytometric analysis, we found Danu treatment of MCF10A, MCF7, and MDA-MB-231 cells induced remarkable autophagy in concentration- and time-dependent manners. Second, there was a striking increase in the expression level of beclin 1 and LC3-II in MCF10A, MCF7, and MDA-MB-231 cells treated with Danu in concentration- and time-dependent manners, as expressed in Western blotting. These results indicate that Danu can induce autophagy in breast cells, and nonmalignant cells are less sensitive to Danu-induced autophagy than malignant MCF7 and MDA-MB-231 cells. Furthermore, we also found that Danu significantly induced p38 MAPK and Erk1/2 activation; on the contrary, it downregulated the activation of Akt and mTOR.
MAPK family members, including JNK, Erk1/2, and p38 MAPK have been reported to be involved in autophagy.53 p38 MAPK is an important stress kinase involved in a number of cellular processes, including inflammation response, cell growth and differentiation, cell cycle progression, and cell death. In the present study, we have observed that Danu treatment significantly increased the phosphorylation of p38 MAPK and Erk1/2 in MCF10A, MCF7, and MDA-MB-231 cells in concentration- and time-dependent manners. With increasing concentrations or sustained time exposed to Danu, an increasing amount of phosphorylated p38 MAPK and Erk1/2, and highly accumulated LC3-II were observed. Thus, the results demonstrate that the activation of p38 MAPK and Erk1/2 is involved in Danu-induced autophagy. We further confirmed our findings via pharmacological inhibitors of autophagy. WM, an inhibitor for the sequestration step of autophagosome, completely blocked Danu-induced p38 MAPK and Erk1/2 activation in MCF7 and MDA-MB-231 cells. These observations suggest that p38 MAPK and Erk1/2 regulate autophagosome formation at the sequestration step. It is noteworthy that SB202190 (an inhibitor of p38 MAPK) downregulates Danu-induced activation of Erk1/2, resulting in a reduction in LC3-II conversion in MCF7 and MDA-MB-231 cells. On the contrary, knockdown of p38 MAPK using p38 MAPK siRNA inhibited Danu-induced phosphorylation of Erk1/2, and eventually caused an accumulation of LC3-II. On the other hand, U0126 (an inhibitor of Erk1/2) also inhibited Danu-induced activation of p-p38 MAPK expression level, and caused the reduction of LC3-II conversion in MCF7 cells. Knockdown of the Erk1/2 gene using Erk1/2 siRNA aided Danu-induced increases in p-p38 MAPK expression level, but that still led to an accumulation of LC3-II in MCF7 cells. These observations further confirmed that p38 MAPK and Erk1/2 played important roles in Danu-induced autophagy.
To understand the underlying mechanism for the autophagy-inducing effect of Danu, we further investigated the classic mTOR-signaling pathway. mTOR is an atypical serine/threonine kinase that is present in two distinct complexes: mTOR complex 1 (mTORC1) and mTOR complex 2 (mTORC2). mTOR plays a key role in cell growth, autophagic cell death, and homeostasis. mTOR is phosphorylated at Ser2448 via the PI3K/Akt signaling pathway, and is autophosphorylated at Ser248. Inhibition of mTOR promotes the dissociation of mTOR from the complex of Atg13 with ULK1 and ULK2. This releases ULK1/2 to activate FIP200, a protein critical for autophagosome formation and autophagy initiation; as an upstream effector of mTOR, Akt can activate mTOR via direct phosphorylation and inhibition of tuberous sclerosis complex 2 (TSC2), which is a negative regulator of mTOR.54 In the present study, Danu significantly reduced the phosphorylation of mTOR at Ser2448 and Akt at Ser473 in both MCF7 and MDA-MB-231 cells, in a concentration-dependent manner. With increasing concentrations of Danu, a decreased phosphorylation level of mTOR and Akt was observed. In addition, there was a time-dependent decrease in the phosphorylation level of mTOR and Akt in MCF7 and MDA-MB-231 cells treated with Danu. However, the total Akt and total mTOR were not significantly changed when MCF7 and MDA-MB-231 cells were treated with Danu. These results suggest that Danu causes a downregulation in the phosphorylation of Akt and mTOR, contributing to the autophagy-inducing effect of Danu in MCF7 and MDA-MB-231 cells.
We further provided evidence about the existence of cross-talk among the p38 MAPK, Erk1/2, and mTOR signaling pathways in the process of Danu-induced autophagy. The possible role of p38 MAPK in the regulation of autophagy has been reported, in that activation of p38 MAPK led to an inhibition of autophagy.55 However, a subsequent study presented an opposite finding that activation of p38 MAPK caused an induction of autophagy.56 Moreover, it has been reported that a novel cell type-specific role of p38 MAPK in the control of autophagy and cell death in colorectal cancer cells exists.57 However, the possible molecular mechanisms that link p38 MAPK to autophagy are not yet known. Our studies have demonstrated that Danu induced activation of p38 MAPK and Erk1/2, and decreased activation of Akt and mTOR. To confirm p38 MAPK and Erk1/2 induced autophagy via the Akt/mTOR signaling pathway, we silenced p38 MAPK and Erk1/2 using p38 MAPK siRNA and Erk1/2 siRNA in MCF7 cells and tested the alteration in the phosphorylation of Akt. We found that MCF7 cells transfected with p38 MAPK siRNA downregulated the level of Danu-induced p-p38 MAPK, but upregulated the activation of Akt, leading to LC3-II accumulation. However, transfection of MCF7 cells with Erk1/2 siRNA did not elicit a marked activation of Akt, but effectively promoted LC3 conversion. Furthermore, we observed the effect of p38 MAPK inhibitor SB202190 and Erk1/2 inhibitor U0126 on Danu-inhibited phosphorylation of Akt in MCF7 and MDA-MB-231 cells. SB202190 completely inhibited the activation of Akt, and U0126 also inhibited the activation of Akt in both cell lines. Taken together, these results demonstrate that p38 MAPK and Erk1/2 signaling pathways are involved in the autophagy-inducing effect of Danu in MCF7 and MDA-MB-231 cells.
EMT, a physiological process essential during embryonic development, is characterized by a loss of cell polarity and cell–cell adhesion, and gain active cell motility when epithelial cells convert to a mesenchymal cell phenotype.58 In the metastasis of cancer, the increased motility and invasive behavior of cancer cells underwent EMT, due to the loss of adhesion molecules such as E-cadherin, and the gain of mesenchymal proteins such as N-cadherin and vimentin.59 In the clinic, breast cancer patient mortality is mainly attributed to the spread of cancerous cells to areas outside the mammary gland and the inadequate strategies to effectively block progression to metastasis, in which EMT plays a critical role.24 In primary breast cancer cells, reduction or loss of expression of E-cadherin and β-catenin were observed.24 In the present study, Danu significantly increased the ratio of E-cadherin over N-cadherin, which would result in an EMT inhibition in breast cancer. Furthermore, Danu suppressed the expression of snail, slug, and TCF8/ZEB1 in MCF7 and MDA-MB-231 cells. Snail and slug are master regulators of EMT, downregulating E-cadherin by silencing gene expression. TCF8/ZEB1 functions as an activator of the EMT process through downregulation of epithelial genes. Moreover, Danu significantly increased the expression level of ZO-1, but decreased the expression level of vimentin and β-catenin. ZO-1 is required for tight junction formation and function involving the regulation of paracellular permeability, and the maintenance of cell polarity, blocking the movement of transmembrane proteins between the apical and the basolateral cell surfaces,60 whereas vimentin is a marker expressed in mesenchymal cells. β-catenin can act as an integral component of a protein complex in adherens junctions that helps cells maintain epithelial layers, and β-catenin participates in the Wnt signaling pathway as a downstream target.24 Similar data were obtained in a time-dependent manner in MCF7 and MDA-M-231 cells. These results indicated that Danu inhibited EMT in breast cancer cells.
Conclusion
In summary, the present study showed that Danu inhibited cell proliferation, induced G2/M arrest, activated the mitochondria-dependent apoptotic pathway, promoted autophagy, and suppressed EMT in MCF7 and MDA-MB-231 cells. Danu induced autophagy via p38 MAPK/Erk1/2–Akt/mTOR signaling pathways in MCF7 and MDA-MB-231 cells. These results indicate that Danu may represent a new anticancer drug that can kill breast cancer cells and prevent EMT. More studies are needed to fully elucidate the underlying mechanisms, efficacy, and safety of Danu in the treatment of human breast cancer.
Acknowledgments
The authors appreciate the financial support from the Startup Fund of the College of Pharmacy, University of South Florida, Tampa, FL, USA. Dr Zhi-Wei Zhou is a holder of a postdoctoral scholarship from the College of Pharmacy, University of South Florida, Tampa, FL, USA.
Disclosure
The authors report no conflicts of interest in this work.
References
Bray F, Jemal A, Grey N, Ferlay J, Forman D. Global cancer transitions according to the Human Development Index (2008–2030): a population-based study. Lancet Oncol. 2012;13(8):790–801. | ||
Ferlay J, Soerjomataram I, Ervik M, et al. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11. Lyon, France: IARC; 2013. | ||
Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64(1):9–29. | ||
USCSW Group. United States Cancer Statistics: 1999–2011. Incidence and Mortality Web-based Report. Atlanta GA: Department of Health and Human Services, Centers for Disease Control and Prevention, and National Cancer Institute; 2014. | ||
Morgan G, Ward R, Barton M. The contribution of cytotoxic chemotherapy to 5-year survival in adult malignancies. Clin Oncol (R Coll Radiol). 2004;16(8):549–560. | ||
Okada H, Mak TW. Pathways of apoptotic and non-apoptotic death in tumour cells. Nat Rev Cancer. 2004;4(8):592–603. | ||
Andrews PD, Knatko E, Moore WJ, Swedlow JR. Mitotic mechanics: the auroras come into view. Curr Opin Cell Biol. 2003;15(6):672683. | ||
Kaestner P, Stolz A, Bastians H. Determinants for the efficiency of anticancer drugs targeting either Aurora-A or Aurora-B kinases in human colon carcinoma cells. Mol Cancer Ther. 2009;8(7):2046–2056. | ||
Kelly KR, Shea TC, Goy A, et al. Phase I study of MLN8237 – investigational Aurora A kinase inhibitor–in relapsed/refractory multiple myeloma, non-Hodgkin lymphoma and chronic lymphocytic leukemia. Invest New Drugs. 2014;32(3):489–499. | ||
Sehdev V, Katsha A, Ecsedy J, Zaika A, Belkhiri A, El-Rifai W. The combination of alisertib, an investigational Aurora kinase A inhibitor, and docetaxel promotes cell death and reduces tumor growth in preclinical cell models of upper gastrointestinal adenocarcinomas. Cancer. 2013;119(4):904–914. | ||
Weier HU, Mao JH. Meta-analysis of Aurora Kinase A (AURKA) expression data reveals a significant correlation between increased AURKA expression and distant metastases in human ER-positive breast cancers. J Data Mining Genomics Proteomics. 2013;4(1):127. | ||
Zhou N, Singh K, Mir MC, et al. The investigational Aurora kinase A inhibitor MLN8237 induces defects in cell viability and cell-cycle progression in malignant bladder cancer cells in vitro and in vivo. Clin Cancer Res. 2013;19(7):1717–1728. | ||
Ruchaud S, Carmena M, Earnshaw WC. The chromosomal passenger complex: one for all and all for one. Cell. 2007;131(2):230–231. | ||
Lapenna S, Giordano A. Cell cycle kinases as therapeutic targets for cancer. Nat Rev Drug Discov. 2009;8(7):547–566. | ||
Lens SM, Voest EE, Medema RH. Shared and separate functions of polo-like kinases and aurora kinases in cancer. Nat Rev Cancer. 2010;10(12):825–841. | ||
Lok W, Klein RQ, Saif MW. Aurora kinase inhibitors as anti-cancer therapy. Anticancer Drugs. 2010;21(4):339–350. | ||
Portella G, Passaro C, Chieffi P. Aurora B: a new prognostic marker and therapeutic target in cancer. Curr Med Chem. 2011;18(4):482–496. | ||
Carpinelli P, Ceruti R, Giorgini ML, et al. PHA-739358, a potent inhibitor of Aurora kinases with a selective target inhibition profile relevant to cancer. Mol Cancer Ther. 2007;6(12 Pt 1):3158–3168. | ||
Meulenbeld HJ, Mathijssen RH, Verweij J, de Wit R, de Jonge MJ. Danusertib, an aurora kinase inhibitor. Expert Opin Investig Drugs. 2012;21(3):383–393. | ||
Steeghs N, Eskens FA, Gelderblom H, et al. Phase I pharmacokinetic and pharmacodynamic study of the aurora kinase inhibitor danusertib in patients with advanced or metastatic solid tumors. J Clin Oncol. 2009;27(30):5094–6101. | ||
Li YC, He SM, He ZX, et al. Plumbagin induces apoptotic and autophagic cell death through inhibition of the PI3K/Akt/mTOR pathway in human non-small cell lung cancer cells. Cancer Lett. 2014;344(2):239–259. | ||
Kang R, Zeh HJ, Lotze MT, Tang D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ. 2011;18(4):571–580. | ||
Kabeya Y, Mizushima N, Ueno T, et al. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000;19(21):5720–5728. | ||
Foroni C, Broggini M, Generali D, Damia G. Epithelial-mesenchymal transition and breast cancer: role, molecular mechanisms and clinical impact. Cancer Treat Rev. 2012;38(6):689–697. | ||
Adhikary A, Chakraborty S, Mazumdar M, et al. Inhibition of epithelial to mesenchymal transition by E-cadherin up-regulation via repression of slug transcription and inhibition of E-cadherin degradation: dual role of SMAR1 in breast cancer cells. J Biol Chem. 2014;289(37):25431–25444. | ||
Dubois-Marshall S, Thomas JS, Faratian D, Harrison DJ, Katz E. Two possible mechanisms of epithelial to mesenchymal transition in invasive ductal breast cancer. Clin Exp Metastasis. 2011;28(8):811–818. | ||
Fu J, Bian M, Jiang Q, Zhang C. Roles of Aurora kinases in mitosis and tumorigenesis. Mol Cancer Res. 2007;5(1):1–10. | ||
Ducat D, Zheng Y. Aurora kinases in spindle assembly and chromosome segregation. Exp Cell Res. 2004;301(1):60–67. | ||
Gautschi O1, Heighway J, Mack PC, Purnell PR, Lara PN Jr, Gandara DR. Aurora kinases as anticancer drug targets. Clin Cancer Res. 2008;14(6):1639–1648. | ||
Katayama H, Brinkley WR, Sen S. The Aurora kinases: role in cell transformation and tumorigenesis. Cancer Metastasis Rev. 2003;22(4):451–464. | ||
Li R, Yerganian G, Duesberg P, et al. Aneuploidy correlated 100% with chemical transformation of Chinese hamster cells. Proc Natl Acad Sci U S A. 1997;94(26):14506–14511. | ||
Bayliss R, Sardon T, Vernos I, Conti E. Structural basis of Aurora-A activation by TPX2 at the mitotic spindle. Mol Cell. 2003;12(4):851–862. | ||
Crosio C, Fimia GM, Loury R, et al. Mitotic phosphorylation of histone H3: spatio-temporal regulation by mammalian Aurora kinases. Mol Cell Biol. 2002;22(3):874–885. | ||
Benten D, Keller G, Quaas A, et al. Aurora kinase inhibitor PHA-739358 suppresses growth of hepatocellular carcinoma in vitro and in a xenograft mouse model. Neoplasia. 2009;11(9):934–944. | ||
Xie L1, Kassner M, Munoz RM, et al. Kinome-wide siRNA screening identifies molecular targets mediating the sensitivity of pancreatic cancer cells to Aurora kinase inhibitors. Biochem Pharmacol. 2012;83(4):452–461. | ||
Hengartner MO. The biochemistry of apoptosis. Nature. 2000;407(6805):770–776. | ||
Sancar A, Lindsey-Boltz LA, Unsal-Kaçmaz K, Linn S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu Rev Biochem. 2004;73:39–85. | ||
Galluzzi L, Bravo-San Pedro JM, Vitale I, et al. Essential versus accessory aspects of cell death: recommendations of the NCCD 2015. Cell Death Differ. 2015;22(1):58–73. | ||
Wyllie AH, Kerr JF, Currie AR. Cell death: the significance of apoptosis. Int Rev Cytol. 1980;68:251–306. | ||
Wolf BB, Green DR. Suicidal tendencies: apoptotic cell death by caspase family proteinases. J Biol Chem. 1999;274(29):20049–20052. | ||
McIlwain DR, Berger T, Mak TW. Caspase functions in cell death and disease. Cold Spring Harb Perspect Biol. 2013;5(4):a008656. | ||
Taylor RC, Cullen SP, Martin SJ. Apoptosis: controlled demolition at the cellular level. Nat Rev Mol Cell Biol. 2008;9(3):231–241. | ||
Klionsky DJ, Emr SD. Autophagy as a regulated pathway of cellular degradation. Science. 2000;290(5497):1717–1721. | ||
Denton D, Nicolson S, Kumar S. Cell death by autophagy: facts and apparent artefacts. Cell Death Differ. 2012;19(1):87–95. | ||
Yang S, Kimmelman. A critical role for autophagy in pancreatic cancer. Autophagy. 2011;7(8):912–913. | ||
Gajewska M, Sobolewska A, Kozlowski M, Motyl T. Role of autophagy in mammary gland development. J Physiol Pharmacol. 2008;59(Suppl 9):237–249. | ||
Cook KL, Shajahan AN, Clarke R. Autophagy and endocrine resistance in breast cancer. Expert Rev Anticancer Ther. 2011;11(8):1283–1294. | ||
Chen N, Karantza-Wadsworth V. Role and regulation of autophagy in cancer. Biochim Biophys Acta. 2009;1793(9):1516–1523. | ||
Maycotte P, Thorburn A. Targeting autophagy in breast cancer. World J Clin Oncol. 2014;5(3):224–240. | ||
Hsieh CC, Kuro-o M, Rosenblatt KP, Brobey R, Papaconstantinou J. The ASK1-Signalosome regulates p38 MAPK activity in response to levels of endogenous oxidative stress in the Klotho mouse models of aging. Aging (Albany NY). 2010;2(9):597–611. | ||
Wang W1, Fan H, Zhou Y, Duan P, Zhao G, Wu G. Knockdown of autophagy-related gene BECLIN 1 promotes cell growth and inhibits apoptosis in the A549 human lung cancer cell line. Mol Med Rep. 2013;7(5):1501–1505. | ||
Lee YJ, Ha YJ, Kang YN, et al. The autophagy-related marker LC3 can predict prognosis in human hepatocellular carcinoma. PLoS One. 2013;8(11):e81540. | ||
Sui X, Kong N, Ye L, et al. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014;344(2):174–179. | ||
Dunlop EA, Tee AR. mTOR and autophagy: a dynamic relationship governed by nutrients and energy. Semin Cell Dev Biol. 2014;36C:121–129. | ||
Brancho D, Tanaka N, Jaeschke A, et al. Mechanism of p38 MAP kinase activation in vivo. Genes Dev. 2003;17(16):1969–1978. | ||
Brancho D1, Ventura JJ, Jaeschke A, Doran B, Flavell RA, Davis RJ. Role of MLK3 in the regulation of mitogen-activated protein kinase signaling cascades. Mol Cell Biol. 2005;25(9):3670–3681. | ||
Comes F, Matrone A, Lastella P, et al. A novel cell type-specific role of p38α in the control of autophagy and cell death in colorectal cancer cells. Cell Death Differ. 2007;14(4):693–702. | ||
Guarino M, Rubino B, Ballabio G. The role of epithelial-mesenchymal transition in cancer pathology. Pathology. 2007;39(3):305–318. | ||
Beavon IR. The E-cadherin-catenin complex in tumour metastasis: structure, function and regulation. Eur J Cancer. 2000;36(13 Spec No):1607–1620. | ||
Shin K, Margolis B. ZOning out tight junctions. Cell. 2006;126(4):647–649. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.