")
Back to Journals » Drug Design, Development and Therapy » Volume 9
Postsynaptic density protein 95-regulated NR2B tyrosine phosphorylation and interactions of Fyn with NR2B in levodopa-induced dyskinesia rat models
Received 7 October 2014
Accepted for publication 11 November 2014
Published 19 December 2014 Volume 2015:9 Pages 199—206
DOI https://doi.org/10.2147/DDDT.S75495
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Shu-Feng Zhou
Maowen Ba,1,* Min Kong,2,* Guozhao Ma3
1Department of Neurology, Yuhuangding Hospital, Yantai City, Shandong, People’s Republic of China; 2Department of Neurology, Yantaishan Hospital, Yantai City, Shandong, People’s Republic of China; 3Department of Neurology, Shandong Provincial Qianfoshan Hospital, Shandong University, Shandong, People’s Republic of China
*These authors contributed equally to this work
Context: Abnormality in interactions between N-methyl-d-aspartate (NMDA) receptor and its signaling molecules occurs in the lesioned striatum in Parkinson’s disease (PD) and levodopa-induced dyskinesia (LID). It was reported that Fyn-mediated NR2B tyrosine phosphorylation, can enhance NMDA receptor function. Postsynaptic density protein 95 (PSD-95), one of the synapse-associated proteins, regulates interactions between receptor and downstream-signaling molecules. In light of the relationship between PSD-95, NR2B, and Fyn kinases, does PSD-95 contribute to the overactivity of NMDA receptor function induced by dopaminergic treatment? To further prove the possibility, the effects of regulating the PSD-95 expression on the augmented NR2B tyrosine phosphorylation and on the interactions of Fyn and NR2B in LID rat models were evaluated.
Methods: In the present study, parkinsonian rat models were established by injecting 6-hydroxydopamine. Subsequently, valid PD rats were treated with levodopa (50 mg/kg/day with benserazide 12.5 mg/kg/day, twice daily) intraperitoneally for 22 days to create LID rat models. Then, the effect of pretreatment with an intrastriatal injection of the PSD-95mRNA antisense oligonucleotides (PSD-95 ASO) on the rotational response to levodopa challenge was assessed. The effects of pretreatment with an intrastriatal injection of PSD-95 ASO on the augmented NR2B tyrosine phosphorylation and interactions of Fyn with NR2B in the LID rat models were detected by immunoblotting and immunoprecipitation.
Results: Levodopa administration twice daily for 22 days to parkinsonian rats shortened the rotational duration and increased the peak turning responses. The altered rotational responses were attenuated by PSD-95 ASO pretreatment. Meanwhile, PSD-95 ASO pretreatment decreased the level of PSD-95 protein expression and reduced both the augmented NR2B tyrosine phosphorylation and interactions of Fyn with NR2B triggered during the levodopa administration in the lesioned striatum of PD rats. However, the protein levels of Fyn and NR2B showed no difference under the above conditions.
Conclusion: The data demonstrate that the inhibition of PSD-95 protein expression suppressed the interactions of Fyn with NR2B and NR2B tyrosine phosphorylation and subsequently downregulated NMDA receptor overactivation, thus providing benefit for the therapy of LID. Therefore, PSD-95 is important for overactivity of NMDA receptor function due to facilitating NR2B tyrosine phosphorylation dependent on Fyn kinase by regulating interactions of Fyn with NR2B under the pathological conditions of LID.
Keywords: PSD-95 ASO, NMDA, rotational response
Introduction
Postsynaptic density protein 95 (PSD-95) is one member of the membrane-associated guanylate kinase family. PSD-95 can anchor and cluster postsynaptic glutamate receptors and play a critical role in synaptic plasticity and neural development.1–3 Recent research shows that the motor response alterations related to chronic levodopa treatment involve the aberrant alterations of glutamatergic postsynaptic density due to the pulsatile dopaminergic receptors activating on striatal neural cells.4–7 Abnormality in interactions between N-methyl-D-aspartate (NMDA) glutamate receptor and its signaling molecules occur in the lesioned striatum in Parkinson’s disease (PD) and levodopa-induced dyskinesia (LID).8 PSD-95 regulates interactions between receptor and downstream-signaling molecules,9,10 but the mechanisms are not fully understood under the pathological circumstances of LID.
Via the SH3, GK, and PDZ domains, PSD-95 can bind to NR2 subunits and numerous signal molecules, including the Src family kinases, such as Src and Fyn,10 which mediate the regulation of NMDA receptor functions and the activation of downstream-specific signaling cascades.11 It has been reported that Fyn-mediated NR2B tyrosine phosphorylation, can enhance NMDA receptor response10 and that PSD-95 coexpression can raise NR2B tyrosine phosphorylation by Fyn kinases. This finding suggests that PSD-95 is related to the upregulation of NMDA receptor function under normal circumstances.
The overactivity of NMDA receptor function results in the expression of LID. Recent evidence also shows that the basis for the effect is that chronic dopaminergic treatment leads to the persistent augment of striatal NMDA receptor response by enhanced NR2B tyrosine phosphorylation dependent on Fyn kinase.12–14 In light of the above relationship between PSD-95, NR2B, and Fyn kinases, does PSD-95 contribute to the overactivity of NMDA receptor function induced by dopaminergic treatment? To further prove the possibility, the effects of regulating the PSD-95 expression on the augmented NR2B tyrosine phosphorylation and on the interactions of Fyn and NR2B in LID rat models were evaluated.
Subjects and methods
Subjects
Sixty female Sprague Dawley rats (weight range, 190–260 g) were used in the study. The experiments were in accordance with the Animals Research: Reporting In Vivo Experiments (ARRIVE) guidelines and the National Institutes of Health care and use of laboratory animals guidelines (NIH publication number 80-23). All protocols involving animals were approved by the Institutional Ethics Committee of Shanghai Jiaotong University School of Medicine (Shanghai City, People’s Republic of China; number of approval 2012-02-25). All surgery was performed under pentobarbital anesthesia, and an effort was made to minimize the number of rats used and their suffering.
Reagents
Mouse monoclonal anti-PSD-95 and rabbit polyclonal anti-NR2B antibody were obtained from Sigma-Aldrich Co. (St Louis, MO, USA). Mouse monoclonal anti-phosphotyrosine and mouse monoclonal anti-Fyn antibodies were ordered from Santa Cruz Biotechnology Inc. (Dallas, TX, USA). Horseradish peroxidase-conjugated goat anti-mouse IgG secondary antibody and horseradish peroxidase-conjugated goat anti-rabbit IgG secondary antibody were ordered from Vector Laboratory (Burlingame, CA, USA). The sequences of PSD-95 missense (5′-GAATGGGTCACCTCC-3′) and antisense (5′-GAATGGGTCACCTCC-3′) oligonucleotides were synthesized by Shanghai Sangon (Shanghai City, People’s Republic of China).
Parkinsonian rat model creation and rotational screening
The rats were anesthetized with pentobarbital (50 mg/kg body weight, intraperitoneally [ip]). The rats were injected with 8 μg 6-hydroxydopamine ([6-OHDA] a concentration of 2 μg/μL, dissolved in 0.9% physiological saline containing 0.02% ascorbic acid) (Sigma-Aldrich Co.); The coordinate of the right medial forebrain bundle was in line with a rat brain atlas (bregma: 4.5 mm; lateral: 0.9 mm; dura: 7.5 mm).15 The stereotactic apparatus with the incisor bar was from Narishige (Tokyo, Japan). Three weeks after 6-OHDA lesion, the rats showing apomorphine-induced rotational response (>100 total turns) were considered successful parkinsonian rat models and were chosen for the next study.16 The control rats received sham-lesion (saline injection) surgery.
Drug treatment and behavioral observations
We set up five groups (eight rats per group), including control rats, PD rats (hemi-6-OHDA lesion), PD rats with chronic levodopa treatment, and PD rats with chronic levodopa treatment plus the PSD-95mRNA antisense oligonucleotides (PSD-95 ASO) or scrambled missense oligonucleotides (PSD-95 MSO) (10 nmol/5 μL per intrastriatal injection). The control rats and PD rats were treated ip with physiological saline twice daily for 22 days.
Rotational responses to acute levodopa challenge (50 mg/kg/day plus 12.5 mg/kg/day benserazide ip, twice daily) (Sigma-Aldrich Co.) in PD rats treated with levodopa for 22 days were assessed at the beginning (treatment day 1) and end (treatment day 22) of levodopa therapy. During the levodopa treatment period, PD rats developed increasingly severe contralateral rotation and/or abnormal involuntary movements affecting cranial, trunk, and limb muscles on the side of the body contralateral to the lesion, as described previously.17 Motor response changes that develop in parkinsonian rats during twice-daily levodopa treatment have features that resemble those appearing in similarly treated parkinsonian patients.16–18 These include shortened rotational response duration and increased peak turning, resembling human wearing-off fluctuations and dyskinesia.17–19 We used the rotational response because it is a well-established paradigm for studying the wearing-off and dyskinesia response16–25 without interference from abnormal involuntary movements that are known to disrupt and even replace the rat’s motor repertoire.16 The rotational duration was measured as the time between the first 5-minute interval when turning exceeded 20% of the peak rate and the first interval when turning fell below 20% of the peak rate. The peak turning intensity was measured as the peak number of contralateral turns in any 5-minute interval. All of the rats that sustained chronic levodopa treatment had developed dyskinesia by the end of the treatment period.
Oligonucleotide sequences were subjected to a Basic Local Alignment Search Tool (BLAST) search on the National Center for Biotechnology Information BLAST server using the GenBank database. PSD-95 ASO are specific for rat PSD-95 mRNA. The effect of pretreatment with the PSD-95 ASO or PSD-95 MSO (10 nmol/5 μL per intrastriatal injection) on the rotational response to levodopa challenge was assessed 3 days after withdrawal of chronic levodopa treatment. The dose (10 nmol) of PSD-95 ASO or MSO was chosen for the study according to previous research as that,26 which could effectively downregulate the expression of PSD-95. In our preliminary experiment, we tried different doses (5 nmol and 10 nmol) of PSD-95 ASO or MSO. The data indicated that the 10 nmol dose of PSD-95 ASO had obvious effects on rotational behavioral alterations. PSD-95 ASO or PSD-95 MSO was infused into the striatum ipsilateral to the lesioned nigrostriatal pathway every 24 hours for 3 days under pentobarbital anesthesia by means of a syringe at coordinates (bregma +0.5 mm; lateral 2.5 mm; dura 4.6 mm). Following recovery, rats were returned to their home cage. Each was placed in the rotometer and given a single challenge dose of levodopa after 12 hours.
Protein extract
Protein extract and Western blot were carried out according to our methods described in a previous paper.25 After sacrifice, rat striatum was dissected and frozen at −80°C for immunoblotting. Striatal tissue was immersed in ice-cold TEVP buffer (10 mM Tris-HCl, 5 mM NaF, 1 mM Na3VO4, 1 mM ethylenediaminetetraacetic acid, 1 mM ethylene glycol tetraacetic acid, pH 7.4). The tissue was homogenized by sonication. The homogenate was centrifuged at 800× g for 10 minutes at 4°C. The supernatant was again centrifuged at 10,000× g at 4°C for 30 minutes. After centrifugation, the pellet was used as total striatal protein homogenate (membrane and cytoplasmic compartments). The total striatal protein homogenate pellet was suspended in ice-cold sample buffer. Protein concentrations were detected with a bicinchoninic acid (BCA) assay kit (Pierce, Rockford, IL, USA).
Immunoblotting
A 15 μg denatured protein sample from the group of control, PD, chronic levodopa treatment rats, and levodopa treatment + PSD-95 ASO or PSD-95 MSO (n=5) was loaded onto a 7.5% sodium dodecyl sulfate polyacrylamide electrophoresis gel. After electrophoresis, the proteins were electrotransferred onto PVDF membrane by way of 250 mA current for 90 minutes at 4°C. The membrane was blocked with 5% skimmed milk for 2 hours and then incubated at 4°C overnight with the indicated antibodies, including mouse monoclonal anti-PSD-95 antibody (1:500), rabbit polyclonal anti-NR2B antibody (1:1,000), mouse monoclonal anti-Fyn antibody (1:1,000), or beta-actin (1:1,000; Abcam plc, Cambridge, UK). The membrane was washed in 1× Tris-buffered saline with 0.1% Tween 20 and incubated with horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG secondary antibody diluted 1:2,000 for 1 hour. Blotting protein was detected by chemiluminescent working solution (Cell Signaling Technology, Boston, USA) and exposed to Kodak BioMax® MS-1 films. Quantitative analysis of immunoblotting was performed by calculating the relative density of the immunoreactive bands after acquisition of the blot image with a Nikon charge-coupled device video camera module and analysis with NIH Image software (v 1.34). The band density is expressed as the mean percent of sham-lesioned striatum for every immunoblot. All of these experiments for biochemical assessments were carried out five times and were repeatable on striatal extracts from five rats. Omission of the primary antibodies served as negative controls. The omission of the primary antibodies against PSD-95, NR2B, or Fyn in immunoblotting procedures provided no visible immunoreactivity in immunoblotting.
Immunoprecipitation
After fourfold dilution with immunoprecipitation buffer containing 50 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) (pH 7.1), 1% Triton X-100, 10% glycerol, 0.5% Nonidet P-40, 1.5 mM MgCl , 1 mM ZnCl, 150 mM NaCl, and inhibitors of phosphatase and protease, a 400 μg protein sample was preincubated with 25 μL protein A Sepharose CL-4B (Amersham, Bucks, UK) for 1 hour at 4°C to remove protein nonspecifically adhered to protein A beads. After centrifugation, the supernatant was incubated with 2 μg antibody overnight at 4°C. Twenty-five microliters of protein A Sepharose CL-4B was added, and the incubation continued for 2 hours at 4°C. The immune complex was isolated by centrifugation and the pellet was washed three times with immunoprecipitation buffer. Bound proteins were eluted by boiling for 5 minutes in Laemmli sample buffer. Then, the sample was centrifuged and supernatant was used for protein analysis by immunoblotting as described above.
Statistical analysis
Data, expressed as mean ± standard error of the mean (SEM), were analyzed by analysis of variance (ANOVA) tests; statistical significance was defined as P<0.05. Statistical tests were calculated using SPSS software (v 13.0).
Results
PSD-95 ASO effects on motor response alterations induced by levodopa treatment
In our previous study,25 chronic levodopa treatment shortened the rotational duration and increased the peak turning responses. In the present study, PSD-95 ASO (10 nmol/5 μL) administration into the striatum ipsilateral to the 6-OHDA lesion 3 days after withdrawal of chronic levodopa treatment prolonged the shortened rotational duration and produced a 38.5%±5.1% reduction of peak turning in response to an acute levodopa challenge. Levodopa-treated rats given a PSD-95 MSO intrastriated infusion showed no evidence of change in their rotational duration and peak turning in response to levodopa (Figure 1).
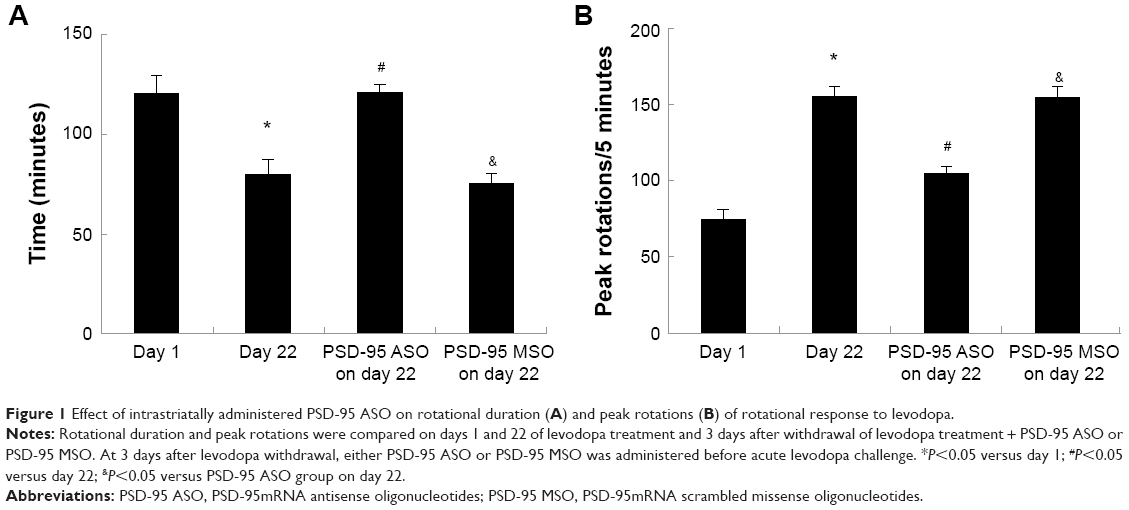 | Figure 1 Effect of intrastriatally administered PSD-95 ASO on rotational duration (A) and peak rotations (B) of rotational response to levodopa. |
Effect of PSD-95 ASO on the expression of PSD-95 in the lesioned striatum
Immunoblotting analysis of striatal total protein extracts indicated the change of PSD-95 expression. The abundance of PSD-95 in striatal total protein extracts was reduced by 6-OHDA lesion. There were 40.7%±6.5% reductions of PSD-95 in 6-OHDA-lesioned rats (P<0.05, compared with control group). Chronic levodopa treatment increased PSD-95 expression significantly. The abundance of PSD-95 on levodopa treatment day 22 increased to 150.2%±9.7% of control (P<0.05, compared with 6-OHDA lesion group). Treatment of rats with PSD-95 ASO every 24 hours for 3 days reduced the PSD-95 protein level by 91.8%±6.7%, while the same dosage of PSD-95 MSO did not affect the PSD-95 expression (Figure 2).
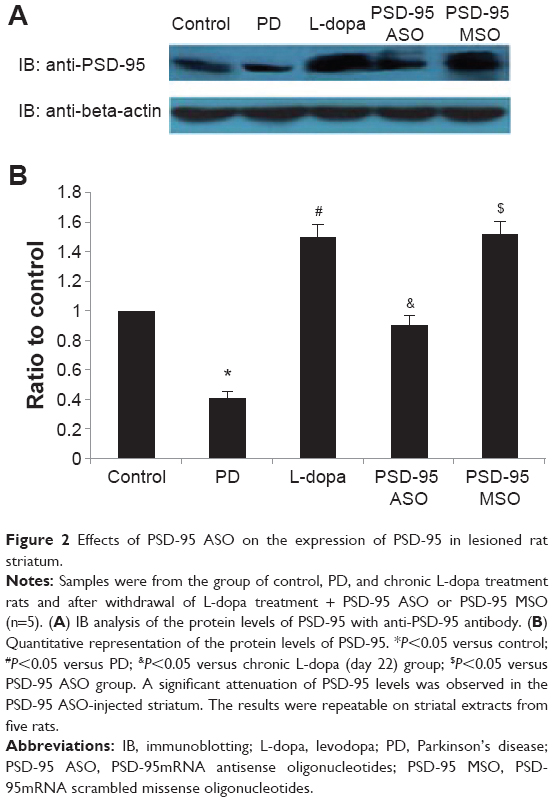 | Figure 2 Effects of PSD-95 ASO on the expression of PSD-95 in lesioned rat striatum. |
Effect of PSD-95 ASO on the augmented tyrosine phosphorylation of NR2B induced by chronic levodopa treatment
To detect the tyrosine phosphorylation level of NR2B, striatal samples (400 μg) were immunoprecipitated with rabbit polyclonal anti-NR2B antibody, then the immunoprecipitates were separated by 7.5% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a PVDF membrane. The membrane was blotted with mouse monoclonal anti-phosphotyrosine antibody. Chronic levodopa treatment increased the tyrosine phosphorylation of NR2B significantly. The abundance of tyrosine-phosphorylated NR2B on levodopa treatment day 22 increased to 1.6-fold that of control (P<0.05, compared with 6-OHDA lesion group). PSD-95 ASO pretreatment reduced the elevation from 1.6- to 1.3-fold relative to control, while the NR2B protein level was not affected. PSD-95 MSO pretreatment had no effect on either tyrosine-phosphorylated NR2B or NR2B protein level. The data indicate that PSD-95 participates in the chronic levodopa treatment-induced alteration in the tyrosine phosphorylation of NR2B (Figure 3).
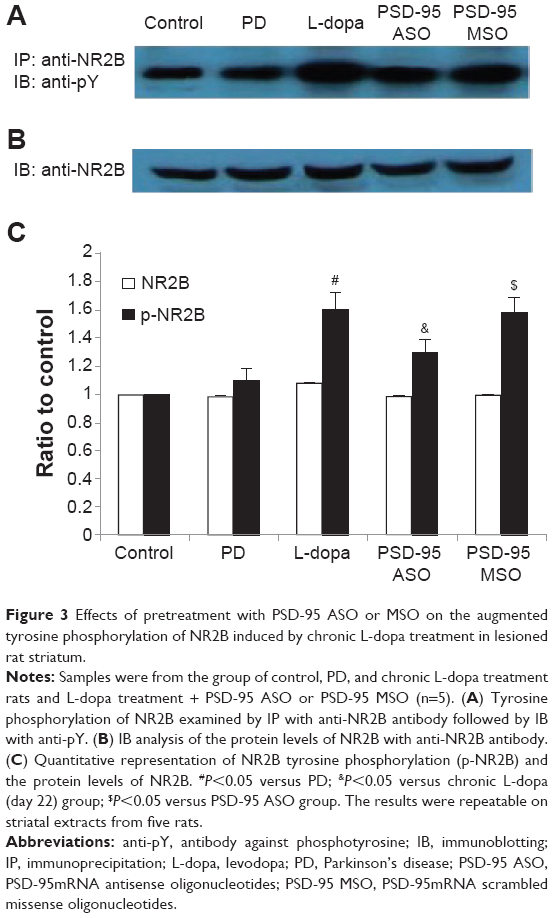 | Figure 3 Effects of pretreatment with PSD-95 ASO or MSO on the augmented tyrosine phosphorylation of NR2B induced by chronic L-dopa treatment in lesioned rat striatum. |
Effect of PSD-95 ASO on the altered interactions of Fyn with NR2B induced by chronic levodopa treatment
The interactions of Fyn with NR2B was detected under non-denatured conditions by co-immunoprecipitation. The co-immunoprecipitation was performed by immunoprecipitation of the striatal samples with antibody of mouse monoclonal anti-Fyn followed by probing the blots with the antibody against NR2B.
The interactions of Fyn with NR2B in the 6-OHDA-lesioned striatum was significantly reduced to 70.1%±5.3% (P<0.05, compared with control group). After chronic levodopa treatment, the interactions of Fyn with NR2B increased up to 1.8-fold relative to control (P<0.05, compared with the 6-OHDA lesion group). The elevation decreased to 1.4-fold relative to control in rats pretreated with PSD-95 ASO, but not in the PSD-95 MSO-treated groups. Immunoblotting analysis demonstrated that the Fyn expression had no differences in control, PD, levodopa treatment, and drug-pretreated rats (Figure 4).
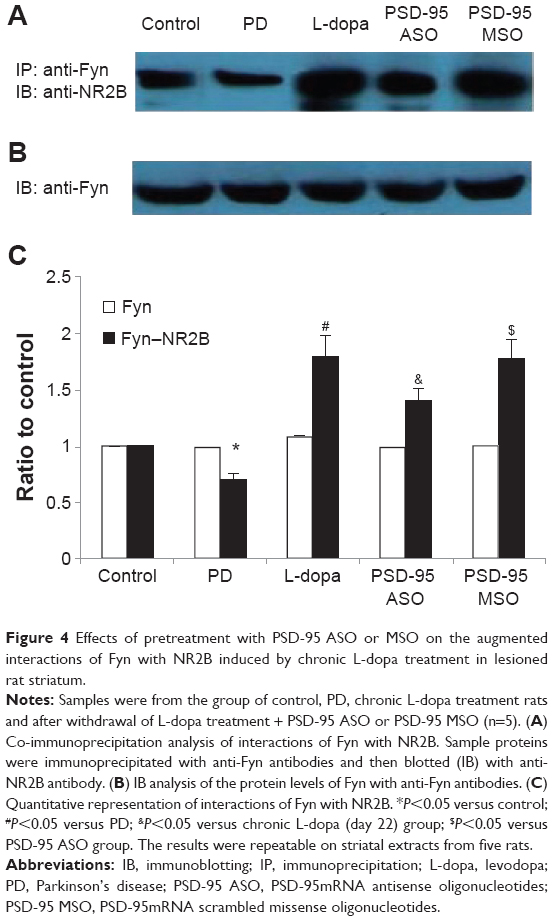 | Figure 4 Effects of pretreatment with PSD-95 ASO or MSO on the augmented interactions of Fyn with NR2B induced by chronic L-dopa treatment in lesioned rat striatum. |
Discussion
Our previous studies found that levodopa treatment administered twice daily for 22 days to 6-OHDA-lesioned rats shortened the rotational duration and increased the peak turning responses.25–27 The expression of motor response alterations persisted after withdrawal of levodopa treatment.28 The striatal synaptic plasticity changes modified motor behavior. Earlier study connected PSD-95 to various forms of behavioral and synaptic plasticity.2–4,8 PSD-95 can anchor and cluster postsynaptic glutamate receptors and play a critical role in synaptic plasticity and neural development.3,10,12 It is well known that synaptic plastic responses, such as long-term potentiation, are critical for behavioral alterations, and these are thought to be mechanisms for the consolidation of information storage in the brain. Long-term potentiation can be mediated by the modulation of PSD-95. For example, others utilizing antisense oligonucleotides, antisense-induced disruption of rat hippocampal PSD-95 reportedly impairs the consolidation of memory.2 In the present study, parkinsonism seems to be related to decreased PSD-95 protein levels. In contrast, LID can be related to increased PSD-95 protein levels. It has also been demonstrated that the expression and subcellular distribution of PSD-95 are altered in the striatum of levodopa-treated unilateral 6-OHDA-lesioned rats with increased expression and enrichment in the synaptic compartment.8 Thus, part of our results confirm previous findings. In addition, based on these findings, pretreatment with PSD-95mRNA antisense oligodeoxynucleotide s (PSD-95 ASO) not only significantly reduced the PSD-95 protein level but also attenuated the rotational response alterations, including peak rotations and response duration to levodopa. Porras et al have also shown that PSD-95 downregulation interferes with behavioral sensitization known as the abnormal involuntary movements to levodopa in 6-OHDA-lesioned rats.29 Thus, the present evidence suggests a close functional relationship between PSD-95 protein levels in the striatum and the expression of levodopa-induced rotational response alterations.
PSD-95 regulates interaction between receptor and downstream-signaling molecules. Given the many functions of PSD-95, in the present study, we define precisely what role the changes observed here might play in PD and LID.
Abnormality in the assembly of receptor with other proteins might result in synaptic dysfunctional signaling. The interaction between receptor and scaffolding proteins allows the formation of subsynaptic rafts of multiple signaling and regulatory molecules at the postsynaptic density. These can be assembled to form an integrated processing unit.1,3 An appreciation of the functioning of a receptor cannot be made in isolation from its interactions with other components of the signalosome. For example, the altered levels of PSD-95 can result in enhanced NMDA receptor signaling, as described in both parkinsonism and LID, because of abnormal interaction between NMDA receptor and scaffolding proteins. As such, interaction subserves indirect interactions with regulatory proteins such as phosphatases and kinases.30,31 This can underlie abnormal NMDA receptor phosphorylation.6
The results of the present study show that chronic levodopa treatment elevates NR2B tyrosine phosphorylation levels and increases association between Fyn and NR2B, which contributes to overactivity of NMDA receptor function. Pretreatment with PSD-95mRNA antisense oligodeoxynucleotide (PSD-95 ASO) reduced the elevated NR2B tyrosine phosphorylation and interaction of Fyn with NR2B triggered during the levodopa administration in the 6-OHDA-lesioned striatum. The data demonstrate that PSD-95 is important for facilitating NR2B tyrosine phosphorylation by Fyn kinase. Functioning as a scaffold, PSD-95 brings Fyn kinases in the proximity of its substrate–NR2B subunits.10,11 Therefore, PSD-95 can control the upregulation of NMDA receptor function by Fyn kinase. The overactivation of NMDA receptors can increase their synaptic strength and striatal output changes in ways that characteristically affect motor responses to dopaminergic stimulation. The inhibition of PSD-95 protein expression suppressed the interactions of Fyn with NR2B and NR2B tyrosine phosphorylation and subsequently downregulated NMDA receptor overactivation, thus provides benefit for the therapy of LID.
In addition to tyrosine phosphorylation of NR2 subunits mediated by Fyn kinases, many of the PSD-95-associated downstream signaling modules of NMDA receptors have been implicated in PD and LID. Elucidating the roles and regulation mechanisms of these signaling pathways is important for further understanding the molecular mechanisms of PD and LID and may provide some practical therapeutic approaches for neurodegenerative diseases.
Conclusion
The results of this study provide evidence that PSD-95 is important for overactivation of NMDA receptor function due to facilitating NR2B tyrosine phosphorylation by Fyn kinase under the pathological conditions of LID, which is likely to be achieved by modulating interactions of Fyn with NR2B.
Acknowledgments
The present research was sponsored by the Chinese National Natural Science Foundation (number 81100954) and Shandong Provincial Natural Science Foundation (ZR2013HQ003).
Disclosure
The authors report no conflicts of interest in this work.
References
Dosemeci A, Makusky AJ, Jankowska-Stephens E, Yang X, Slotta DJ, Markey SP. Composition of the synaptic PSD-95 complex. Mol Cell Proteomics. 2007;6:1749–1760. | ||
Migaud M, Charlesworth P, Dempster M, et al. Enhanced long-term potentiation and impaired learning in mice with mutant postsynaptic density-95 protein. Nature. 1998;396:433–439. | ||
Xu W, Schlüter OM, Steiner P, Czervionke BL, Sabatini B, Malenka RC. Molecular dissociation of the role of PSD-95 in regulating synaptic strength and LTD. Neuron. 2008;57:248–262. | ||
Hurley MJ, Jackson MJ, Smith LA, Rose S, Jenner P. Immunoautoradiographic analysis of NMDA receptor subunits and associated postsynaptic density proteins in the brain of dyskinetic MPTP-treated common marmosets. Eur J Neurosci. 2005;21:3240–3250. | ||
Quintana A, Melon C, Kerkerian-Le Goff L, Salin P, Savasta M, Sgambato-Faure V. Forelimb dyskinesia mediated by high-frequency stimulation of the subthalamic nucleus is linked to rapid activation of the NR2B subunit of N-methyl-D-aspartate receptors. Eur J Neurosci. 2010;32:423–434. | ||
Oh JD, Chase TN. Glutamate-mediated striatal dysregulation and the pathogenesis of motor response complications in Parkinson’s disease. Amino Acids. 2002;23:133–139. | ||
Sgambato-Faure V, Cenci MA. Glutamatergic mechanisms in the dyskinesias induced by pharmacological dopamine replacement and deep brain stimulation for the treatment of Parkinson’s disease. Prog Neurobiol. 2012;96:69–86. | ||
Nash JE, Johnston TH, Collingridge GL, Garner CC, Brotchie JM. Subcellular redistribution of the synapse-associated proteins PSD-95 and SAP97 in animal models of Parkinson’s disease and L-DOPA-induced dyskinesia. FASEB J. 2005;19:583–585. | ||
Prybylowski K, Fu Z, Losi G, et al. Relationship between availability of NMDA receptor subunits and their expression at the synapse. J Neurosci. 2002;22:8902–8910. | ||
Losi G, Prybylowski K, Fu Z, Luo J, Wenthold RJ, Vicini S. PSD-95 regulates NMDA receptors in developing cerebellar granule neurons of the rat. J Physiol. 2003;548:21–29. | ||
Nakazawa T, Tezuka T, Yamamoto T. Regulation of NMDA receptor function by Fyn-mediated tyrosine phosphorylation. Nihon Shinkei Seishin Yakurigaku Zasshi. 2002;22:165–167. Japanese. | ||
Sheng M, Kim MJ. Postsynaptic signaling and plasticity mechanisms. Science. 2002;298:776–780. | ||
Johnson KA, Conn PJ, Niswender CM. Glutamate receptors as therapeutic targets for Parkinson’s disease. CNS Neurol Disord Drug Targets. 2009;8:475–491. | ||
Wessell RH, Ahmed SM, Menniti FS, Dunbar GL, Chase TN, Oh JD. NR2B selective NMDA receptor antagonist CP-101,606 prevents levodopa-induced motor response alterations in hemi-parkinsonian rats. Neuropharmacology. 2004;47:184–194. | ||
Paxions G, Watson C. The rat brain in stereotoxic coordinates. Sydney: Academic Press; 2007. | ||
Papa SM, Engber TM, Kask AM, Chase TN. Motor fluctuations in levodopa treated parkinsonian rats: relation to lesion extent and treatment duration. Brain Res. 1994;662:69–74. | ||
Lundblad M, Andersson M, Winkler C, Kirik D, Wierup N, Cenci MA. Pharmacological validation of behavioural measures of akinesia and dyskinesia in a rat model of Parkinson’s disease. Eur J Neurosci. 2002;15:120–132. | ||
Marin C, Aguilar E, Bonastre M, Tolosa E, Obeso JA. Early administration of entacapone prevents levodopa-induced motor fluctuations in hemiparkinsonian rats. Exp Neurol. 2005;192:184–193. | ||
Henry B, Crossman AR, Brotchie JM. Characterization of enhanced behavioral responses to L-DOPA following repeated administration in the 6-hydroxydopamine-lesioned rat model of Parkinson’s disease. Exp Neurol. 1998;151:334–342. | ||
Engber TM, Papa SM, Boldry RC, Chase TN. NMDA receptor blockade reverses motor response alterations induced by levodopa. Neuroreport. 1994;5:2586–2588. | ||
Marin C, Jimenez A, Bonastre M, Chase TN, Tolosa E. Non-NMDA receptor-mediated mechanisms are involved in levodopa-induced motor response alterations in Parkinsonian rats. Synapse. 2000;36:267–274. | ||
Tomiyama M, Kimura T, Maeda T, Kannari K, Matsunaga M, Baba M. A serotonin 5-HT1A receptor agonist prevents behavioral sensitization to L-DOPA in a rodent model of Parkinson’s disease. Neurosci Res. 2005;52:185–194. | ||
Marin C, Rodriguez-Oroz MC, Obeso JA. Motor complications in Parkinson’s disease and the clinical significance of rotational behavior in the rat: have we wasted our time? Exp Neurol. 2006;197:269–274. | ||
Lane EL, Cheetham SC, Jenner P. Does contraversive circling in the 6-OHDA-lesioned rat indicate an ability to induce motor complications as well as therapeutic effects in Parkinson’s disease? Exp Neurol. 2006; 197:284–290. | ||
Ba M, Kong M, Ma G, et al. Cellular and behavioral effects of 5-HT1A receptor agonist 8-OH-DPAT in a rat model of levodopa-induced motor complications. Brain Res. 2007;1127:177–184. | ||
Hou XY, Zhang GY, Zong YY. Suppression of postsynaptic density protein 95 expression attenuates increased tyrosine phosphorylation of NR2A subunits of N-methyl-D-aspartate receptors and interactions of Src and Fyn with NR2A after transient brain ischemia in rat hippocampus. Neurosci Lett. 2003;343:125–128. | ||
Ba M, Kong M, Yang H, et al. Changes in subcellular distribution and phosphorylation of GluR1 in lesioned striatum of 6-hydroxydopamine-lesioned and l-dopa-treated rats. Neurochem Res. 2006;31:1337–1347. | ||
Ba M, Kong M, Yu G, Sun X, Liu Z, Wang X. GluR1 phosphorylation and persistent expression of levodopa-induced motor response alterations in the Hemi-Parkinsonian rat. Neurochem Res. 2011;36:1135–1144. | ||
Porras G, Berthet A, Dehay B, et al. PSD-95 expression controls L-DOPA dyskinesia through dopamine D1 receptor trafficking. J Clin Invest. 2012;122:3977–3989. | ||
Nagura H, Ishikawa Y, Kobayashi K, et al. Impaired synaptic clustering of postsynaptic density proteins and altered signal transmission in hippocampal neurons, and disrupted learning behavior in PDZ1 and PDZ2 ligand binding-deficient PSD-95 knockin mice. Mol Brain. 2012;5:43. | ||
Colledge M, Dean RA, Scott GK, Langeberg LK, Huganir RL, Scott JD. Targeting of PKA to glutamate receptors through a MAGUK-AKAP complex. Neuron. 2000;27:107–119. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.