")
Back to Journals » Drug Design, Development and Therapy » Volume 8
Efficacy of short novel antimicrobial and anti-inflammatory peptides in a mouse model of methicillin-resistant Staphylococcus aureus (MRSA) skin infection
Received 2 August 2014
Accepted for publication 16 September 2014
Published 17 October 2014 Volume 2014:8 Pages 1979—1983
DOI https://doi.org/10.2147/DDDT.S72129
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Shu-Feng Zhou
Mohamed F Mohamed, Mohamed N Seleem
Department of Comparative Pathobiology, College of Veterinary Medicine, Purdue University, West Lafayette, IN, USA
Abstract: The therapeutic efficacy of two novel short antimicrobial and anti-inflammatory peptides (RR and RRIKA) was evaluated in a mouse model of staphylococcal skin infection. RR (2%) and RRIKA (2%) significantly reduced the bacterial counts and the levels of proinflammatory cytokines, tumor necrosis factor (TNF)-α, and interleukin (IL)-6, in methicillin-resistant Staphylococcus aureus USA 300-0114 skin lesions. Furthermore, the combined therapy of RRIKA (1%) and lysostaphin (0.5%) had significantly higher antistaphylococcal and anti-inflammatory activity compared to monotherapy. This study supports the potential use of these peptides for topical treatment of methicillin-resistant Staphylococcus aureus skin infections.
Keywords: antimicrobial peptides, MRSA, lysostaphin, skin infection, mice
Introduction
Bacterial infections, especially those caused by Staphylococcus aureus, are the most significant complication encountered in the management of wounds.1 Furthermore, multidrug-resistant S. aureus strains and their secreted toxins are responsible for interfering with the wound-healing process and creating portals of entry for systemic complications in affected patients.2 With the increasing incidence of staphylococcal resistance to topical antimicrobials, such as mupirocin and fusidic acid,3–5 there is a pressing need to develop novel antimicrobials and new approaches to circumvent this burgeoning problem. Recently, there has been increased interest in the development of antimicrobial peptides (AMPs) as novel therapeutics, due to their high potency, broad spectrum of activity, and reduced potential for resistance development.6,7 In addition to the potent bactericidal activity of AMPs, the recognized anti-inflammatory response of certain AMPs should be an advantage in the treatment of S. aureus skin infections.8
In a recent study, we described two novel short peptides – RR (WLRRIKAWLRR) and RRIKA (WLRRIKAWLRRIKA) – with potent bactericidal activity in vitro against multiple clinical isolates of methicillin-resistant S. aureus (MRSA).6 In particular, the peptides were active against the highly virulent MRSA USA 300-0114, a community-associated strain responsible for outbreaks of staphylococcal skin and soft-tissue infections in the US.9 Moreover, RR and RRIKA were superior in reducing adherent biofilms of both S. aureus and Staphylococcus epidermidis when compared to conventional antibiotics. Furthermore, both RR and RRIKA enhanced the antistaphylococcal activity of lysostaphin in vitro more than 1,000-fold.6 Although lysostaphin demonstrated potent efficacy against MRSA infections in different animal models,10–12 its therapeutic potential was hampered by the emergence of bacterial resistance.10,13 In light of our previous results, showing enhancement of the antimicrobial effectiveness of lysostaphin against MRSA when combined with AMPs in vitro,6 we moved forward with an in vivo experiment in a mouse model of MRSA skin infection.
Materials and methods
Bacterial isolate
We obtained the community-acquired MSRA strain NRS384 (MRSA USA 300-0114) isolated from a wound from a patient in Mississippi, USA. The strain is resistant to erythromycin and tetracycline; positive for mec (subtype IV); pvl+/−; multilocus sequence type 8−; eGenomic spa type 1, eGenomic spa repeats YHGFMBQBLO; Ridom spa type t008; agr grp I.
Peptides and antibacterial agents
RR and RRIKA were synthesized by GenScript (Piscataway, NJ, USA). Recombinant lysostaphin was purchased from Ambi Products (Lawrence, NY, USA). Linezolid and fusidic acid were purchased from Chem-Impex International (Wood Dale, IL, USA).
Mouse infection
Female BALB/c mice (6–8 weeks old) were obtained from Harlan Laboratories (Indianapolis, IN, USA). All procedures were approved by the Purdue University Animal Care and Use Committee (1207000676). The murine model of MRSA skin infection has been described before.14,15 Eight groups of mice (n=5) were inoculated with 40 μL of MRSA USA 300 (3×107 CFU) intradermally. Forty-eight hours after infection and formation of an open wound, six 6 groups were treated topically with either 2% fusidic acid, 2% RR, 2% RRIKA, 1% RRIKA, 0.5% lysostaphin, or 1% RRIKA plus 0.5% lysostaphin formulated in 20 mg petroleum jelly. One group received vehicle only (petroleum jelly), and the last group was treated orally with linezolid (25 mg/kg). All groups were treated twice a day for 3 days. Twenty-four hours after the last treatment, mice were humanely killed, the area around the wound lightly swabbed with 70% ethanol, and the wound (around 1 cm2) excised for bacterial counting after homogenization in 1 mL tryptic soy broth.
Cytokine detection
Enzyme-linked immunosorbent assay (ELISA) development kits for detection of cytokines were purchased from R&D Systems (Minneapolis, MN, USA). Homogenized skin lesions after bacterial counting were centrifuged at 15,000 rpm for 10 minutes. The supernatants were removed and kept at -20°C until analyzed. The supernatants were examined for cytokine production: tumor-necrosis factor (TNF)-α, interleukin (IL)-6, and IL-1β using ELISA as described before.16 Cytokine levels were expressed as percentage change relative to negative control.
Statistical analyses
Data are presented as means ± standard deviation. Statistical analyses were performed using GraphPad Prism 6.0 (GraphPad Software, La Jolla, CA, USA). P-values were calculated by the two-tailed Student’s t-test. P-values <0.05 were considered significant.
Results and discussion
As shown in Figure 1, all treatments significantly reduced the mean bacterial counts compared with the control group (P≤0.01). Among groups treated with monotherapy, the group treated with 2% RRIKA had the highest reduction in CFU (2.08±0.20 log10), followed by 2% fusidic acid (1.94±0.36 log10), 2% RR (1.83±0.30 log10), and linezolid (1.74±0.53 log10). Groups treated with 0.5% lysostaphin or 1% RRIKA achieved a significant reduction of 1.79±0.56 log10 and 1.08±0.36 log10, respectively, when compared with the control group (P≤0.01) (Figure 1). The combination therapy (0.5% lysostaphin with 1% RRIKA) was significantly more effective than treatment with RRIKA or lysostaphin alone, and achieved statistically significant bacterial reduction of 2.65±0.44 log10 compared to the negative control (P≤0.01). The bacterial reduction in the combined therapy was also statistically significant compared to monotherapy (P<0.05) or control antibiotics (P<0.05). Previously, RR and RRIKA showed potent immunomodulatory activities in vitro through inhibition of mitogen-activated protein kinase (MK2).17,18 In this study, topical treatment with RRIKA and RR significantly reduced TNFα and IL-6 production in MRSA skin lesions. As shown in Figure 2A and B, 2% RR and 2% RRIKA reduced the TNFα level by 49% and 56%, respectively, while the IL-6 level was reduced by 29% and 60%, respectively. Treatment with 0.5% lysostaphin and 2% fusidic acid caused 39% and 29% reduction of the TNFα level, respectively, and 13% and 25% reduction of the IL-6 level, respectively, which is in agreement with previous findings.19,20 There was a synergistic anti-inflammatory response observed when 0.5% lysostaphin was combined with 1% RRIKA. The combined therapy significantly reduced TNFα and IL-6 levels by 62% and 67%, respectively. On the other hand, none of the treatments interfered with IL-1β production, which is necessary for S. aureus clearance in cutaneous infections (Figure 2C).21–22
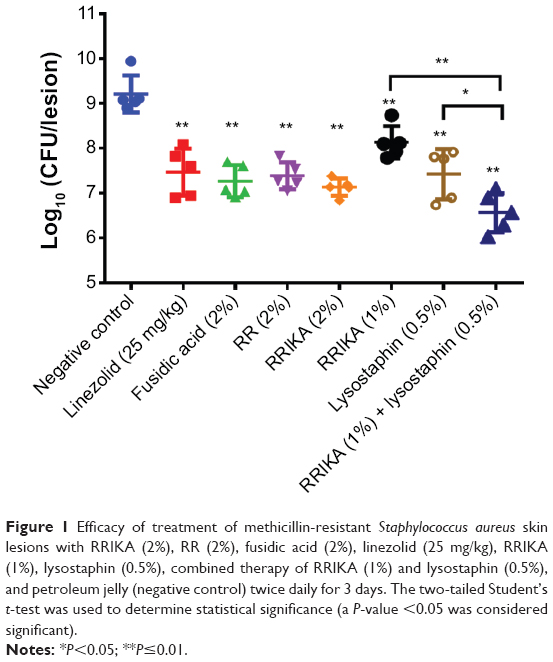 | Figure 1 Efficacy of treatment of methicillin-resistant Staphylococcus aureus skin lesions with RRIKA (2%), RR (2%), fusidic acid (2%), linezolid (25 mg/kg), RRIKA (1%), lysostaphin (0.5%), combined therapy of RRIKA (1%) and lysostaphin (0.5%), and petroleum jelly (negative control) twice daily for 3 days. The two-tailed Student’s t-test was used to determine statistical significance (a P-value <0.05 was considered significant). |
 | Figure 2 The effect of peptides on cytokines TNFα (A), IL-6 (B), and IL-1β (C) production in methicillin-resistant Staphylococcus aureus skin lesions. Tissue homogenate supernatants were examined for cytokine production using the enzyme-linked immunosorbent assay. Cytokine levels were expressed as percentage change relative to negative control. Data are presented as means ± standard deviation from duplicates consisting of four mice per group. Statistical analysis was calculated by the two-tailed Student’s t-test. P-values <0.05 were considered significant (*). |
In this study, topical application of RR and RRIKA was shown to be very effective in reducing the bacterial load in MRSA skin lesions. Moreover, peptides reduced the release of TNFα and IL-6, which might benefit the healing of infected wounds.23–27 In addition, the combination of RRIKA with lysostaphin was significantly more effective in the treatment of MRSA skin lesions than treatment with either peptide alone. This combination therapy is also expected to overcome some of the limitations associated with lysostaphin monotherapy through hindering the emergence of bacterial resistance and lowering the required therapeutic dose.
In conclusion, our findings in MRSA skin lesions should significantly impact and inform efforts to use a combination of anti-inflammatory and AMP therapies as novel topical treatment options for multidrug-resistant pathogens.
Acknowledgments
Mohamed F Mohamed is supported by a scholarship from the Egyptian Cultural and Educational Bureau (ECEB) in Washington, DC, USA. The bacterial strain used in this study was provided by the Network on Antimicrobial Resistance in Staphylococcus aureus (NARSA) for distribution by BEI Resources, NIAID, NIH: Staphylococcus aureus, strain USA300-0114, NR-46070.
Disclosure
The authors report no conflicts of interest in this work.
References
Sen CK, Gordillo GM, Roy S, et al. Human skin wounds: a major and snowballing threat to public health and the economy. Wound Repair Regen. 2009;17(6):763–771. | ||
Guo S, Dipietro LA. Factors affecting wound healing. J Dent Res. 2010;89(3):219–229. | ||
Dobie D, Gray J. Fusidic acid resistance in Staphylococcus aureus. Arch Dis Child. 2004;89(1):74–77. | ||
Kresken M, Hafner D, Schmitz FJ, Wichelhaus TA. Prevalence of mupirocin resistance in clinical isolates of Staphylococcus aureus and Staphylococcus epidermidis: results of the Antimicrobial Resistance Surveillance Study of the Paul-Ehrlich-Society for Chemotherapy, 2001. Int J Antimicrob Agents. 2004;23(6):577–581. | ||
Chambers HF, Deleo FR. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol. 2009;7(9):629–641. | ||
Mohamed MF, Hamed MI, Panitch A, Seleem MN. Targeting methicillin-resistant Staphylococcus aureus with short salt-resistant synthetic peptides. Antimicrob Agents Chemother. 2014;58(7):4113–4122. | ||
Hancock RE, Sahl HG. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotechnol. 2006;24(12):1551–1557. | ||
Montgomery CP, Daniels MD, Zhao F, Spellberg B, Chong AS, Daum RS. Local inflammation exacerbates the severity of Staphylococcus aureus skin infection. PloS One. 2013;8(7):e69508. | ||
King MD, Humphrey BJ, Wang YF, Kourbatova EV, Ray SM, Blumberg HM. Emergence of community-acquired methicillin-resistant Staphylococcus aureus USA 300 clone as the predominant cause of skin and soft-tissue infections. Ann Intern Med. 2006;144(5):309–317. | ||
Kumar JK. Lysostaphin: an antistaphylococcal agent. Appl Microbiol Biotechnol. 2008;80(4):555–561. | ||
Desbois AP, Gemmell CG, Coote PJ. In vivo efficacy of the antimicrobial peptide ranalexin in combination with the endopeptidase lysostaphin against wound and systemic meticillin-resistant Staphylococcus aureus (MRSA) infections. Int J Antimicrob Agents. 2010;35(6):559–565. | ||
Kokai-Kun JF, Walsh SM, Chanturiya T, Mond JJ. Lysostaphin cream eradicates Staphylococcus aureus nasal colonization in a cotton rat model. Antimicrob Agents Chemother. 2003;47(5):1589–1597. | ||
Kusuma C, Jadanova A, Chanturiya T, Kokai-Kun JF. Lysostaphin-resistant variants of Staphylococcus aureus demonstrate reduced fitness in vitro and in vivo. Antimicrob Agents Chemother. 2007;51(2):475–482. | ||
Cho JS, Zussman J, Donegan NP, et al. Noninvasive in vivo imaging to evaluate immune responses and antimicrobial therapy against Staphylococcus aureus and USA300 MRSA skin infections. J Invest Dermatol. 2011;131(4):907–915. | ||
Cho JS, Pietras EM, Garcia NC, et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J Clin Invest. 2010;120(5):1762–1773. | ||
De Vry CG, Valdez M, Lazarov M, et al. Topical application of a novel immunomodulatory peptide, RDP58, reduces skin inflammation in the phorbol ester-induced dermatitis model. J Invest Dermatol. 2005;125(3):473–481. | ||
Ward B, Seal BL, Brophy CM, Panitch A. Design of a bioactive cell-penetrating peptide: when a transduction domain does more than transduce. J Pept Sci. 2009;15(10):668–674. | ||
Lopes LB, Brophy CM, Flynn CR, et al. A novel cell permeant peptide inhibitor of MAPKAP kinase II inhibits intimal hyperplasia in a human saphenous vein organ culture model. J Vasc Surg. 2010;52(6):1596–1607. | ||
Schmelcher M, Powell AM, Becker SC, Camp MJ, Donovan DM. Chimeric phage lysins act synergistically with lysostaphin to kill mastitis-causing Staphylococcus aureus in murine mammary glands. Appl Environ Microbiol. 2012;78(7):2297–2305. | ||
Rubin BK, Tamaoki J. Antibiotics as Anti-inflammatory and Immunomodulatory Agents. Basel: Birkhäuser; 2005. | ||
Miller LS, Cho JS. Immunity against Staphylococcus aureus cutaneous infections. Nat Rev Immunol. 2011;11(8):505–518. | ||
Cho JS, Guo Y, Ramos RI, et al. Neutrophil-derived IL-1β is sufficient for abscess formation in immunity against Staphylococcus aureus in mice. PloS Pathog. 2012;8(11):e1003047. | ||
Jialal I, Miguelino E, Griffen SC, Devaraj S. Concomitant reduction of low-density lipoprotein-cholesterol and biomarkers of inflammation with low-dose simvastatin therapy in patients with type 1 diabetes. J Clin Endocrinol Metab. 2007;92(8):3136–3140. | ||
Wallace HJ, Stacey MC. Levels of tumor necrosis factor-alpha (TNF-alpha) and soluble TNF receptors in chronic venous leg ulcers – correlations to healing status. J Invest Dermatol. 1998;110(3):292–296. | ||
Cowin AJ, Hatzirodos N, Rigden J, Fitridge R, Belford DA. Etanercept decreases tumor necrosis factor-alpha activity in chronic wound fluid. Wound Repair Regen. 2006;14(4):421–426. | ||
Donath MY. Targeting inflammation in the treatment of type 2 diabetes: time to start. Nat Rev Drug Discov. 2014;13(6):465–476. | ||
Khanna S, Biswas S, Shang Y, et al. Macrophage dysfunction impairs resolution of inflammation in the wounds of diabetic mice. PloS One. 2010;5(3):e9539. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.