")
Back to Journals » Drug Design, Development and Therapy » Volume 14
Cyanidin-3-O-Glucoside Improves Colonic Motility During Severe Acute Pancreatitis by Inhibiting the H2S-Regulated AMPK/mTOR Pathway
Received 1 April 2020
Accepted for publication 22 July 2020
Published 19 August 2020 Volume 2020:14 Pages 3385—3391
DOI https://doi.org/10.2147/DDDT.S256450
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Wei Lian, Wensheng Chen
Department of Gastroenterology, Southwest Hospital of Army Medical University, Chongqing, People’s Republic of China
Correspondence: Wensheng Chen
Department of Gastroenterology, Southwest Hospital of Army Medical University, Gaotanyanzheng Street, Shapingba District, Chongqing 400038, People’s Republic of China
Email [email protected]
Background: Cyanidin-3-O-glucoside (C3G) is an important anthocyanin that can modulate digestive system functioning. Inflammation associated with severe acute pancreatitis (SAP) induces H2S production, which impairs the gastrointestinal (GI) system. We investigated the effects of C3G in attenuating SAP-associated colonic motility loss by examining the H2S level and activity of AMP-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) pathway.
Methods: A rat model of SAP was induced using sodium taurocholate, and the effect of C3G on colonic mobility, H2S production, and the inflammatory response was investigated. AMPK/mTOR pathway changes were detected to assess the pathways by which H2S influences colonic mobility in SAP-model rats. The mechanism underlying H2S function was further examined by subjecting colonic muscle cells (CMCs) to C3G, SAP plasma and an AMPK activator.
Results: Administering C3G improved colonic motility but suppressed the inflammatory response and H2S production in the SAP-model rats, which was associated with inhibiting the AMPK/mTOR pathway. Furthermore, activating the AMPK/mTOR pathway in CMCs promoted inflammation but suppressed Ca2+ levels, even after administering C3G.
Conclusion: Administering C3G may improve SAP-associated colonic mobility by inhibiting the H2S-mediated AMPK/mTOR pathway.
Keywords: AMP-activated protein kinase, AMPK, cyanidin-3-O-glucoside, colonic motility, severe acute pancreatitis, hydrogen sulfide
Introduction
Acute pancreatitis (AP) is an inflammatory process that occurs in an otherwise healthy pancreas and exhibits wide clinical variation.1 Although most cases of AP are mild, ~20% of patients with AP develop a severe form of the disease characterized by organ dysfunction,1 known as severe acute pancreatitis (SAP).2 Most patients with SAP experience a diverse range of severe symptoms,3 but current management strategies overlook the key role of intestinal function during SAP development. Previous studies have demonstrated that bacterial infection and intestinal organ sepsis are important factors in SAP development;4,5 thus, increasing research is being conducted regarding novel therapies to limit colonic injuries.6–9
Previous studies have reported that H2S is produced predominantly by cystathionine-γ-lyase (CSE) and other kinases in the transsulfuration pathway. Moreover, H2S initiates distinct biological responses in the human body,10–12 including the gastrointestinal (GI) tract, where H2S is produced by both GI tissues and gut bacterial flora.13,14 H2S production was also reported to inhibit GI motility in a fish model.13,14 Substantial H2S is produced during SAP attacks, where it inhibits inflammation in the GI system during SAP progression.15,16 Therefore, H2S produced during SAP attacks is hypothesized to be associated with impaired intestinal mobility, and modulating H2S production may be a novel strategy for managing SAP-associated colonic motility loss in clinical settings.
Previous studies investigating the events associated with colonic motility loss have furthered the development of novel therapeutic approaches. For example, dietotherapy is attracting increased attention for its efficacy in improving digestive system functioning with few adverse effects.17,18 Moreover, anthocyanins, which belong to the flavonoid family, are widely distributed in vegetables and other foods that are part of the human diet19 and may carry health benefits owing to their antioxidant and anti-inflammatory properties.20 Anthocyanins also affect the intestinal system; thus, studies have focused on their potential to modulate and improve the microflora in the GI tract.21 However, no previous studies examining the interaction between anthocyanins and the GI system have assessed the effect of anthocyanins on intestinal motility. As one of the most abundant natural anthocyanins, cyanidin-3-O-glucoside (C3G) contributes to modulating numerous biological processes, particularly those involved in immunoregulation.22–24 C3G exerts its functions in the GI tract via multiple mechanisms; therefore, the present study was conducted to investigate the protective effect of C3G against SAP-induced colonic motility loss by focusing on the effect of C3G on H2S and its downstream pathways.
To evaluate the study’s hypothesis, rats were injected with sodium taurocholate to induce SAP, then C3G was administered. The effects of C3G on colonic motility, H2S production, and inflammatory cytokine levels were detected to identify the exact effect of C3G on SAP-induced colonic motility loss. Moreover, the changing pattern of H2S-mediated AMP-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) signaling25 was detected to elucidate the mechanism by which C3G restores colonic mobility. To validate the in vivo assay results, colonic muscle cells (CMCs) were isolated and subjected to plasma isolated from SAP-model rats, then C3G and the AMPK inhibitor, MK-3903, were used to elucidate the interaction between C3G and the H2S-mediated AMPK/mTOR pathway.
Materials and Methods
Chemicals and Antibodies
C3G (purity >98%; cat. no. HY-N0640) was purchased from MedChemExpress. Antibodies against CSE (cat. no. 12,217-1-AP) were purchased from ProteinTech Group, Inc. Antibodies against total (t)-AMPK (cat. no. ab32047), phosphorylated (p)-AMPK (cat. no. ab23875), total (t)-mTOR (cat. no. ab2732), p-mTOR (cat. no. ab109268) and GAPDH (cat. no. ab181602) were purchased from Abcam. Secondary antibodies (cat. no. A0277, goat anti-rabbit; cat. no. A0208, goat anti-rabbit; cat. no. A0216, goat anti-murine) were provided by Beyotime Institute of Biotechnology. MK-3903 (an AMPK activator) was purchased from Sigma-Aldrich (Merck KGaA). The AMPK activator, MK-3903 (cat. no. HY-107,988), was purchased from MedChemExpress.
SAP-Model Rats and C3G Administration
Adult male Wistar rats weighing 200–250 g were housed per routine protocols and grouped into the sham, SAP or SAP + C3G groups, with n=10 rats per group. The sham group was induced with SAP without injecting the corresponding agents. The SAP group rats were anesthetized with 50 mg/kg body weight pentobarbital sodium, subjected to a laparotomy, then injected with 5% sodium taurocholate into the pancreatic and bile ducts using a microinjection pump (1 mL/kg, 0.1 mL/min) for 10 min. The incision was then sutured (the overall survival rate of the SAP-model rats was ~60% over a 24-hour period; Figure S1). The SAP + C3G group rats were gavaged with 100 mg/kg body weight C3G and injected with sodium taurocholate. Before the subsequent assays, all rats were injected with 10 mL normal saline and fasted for 24 h. All animal experiments were conducted in accordance with the Institutional Animal Ethics Committee and Animal Care Guidelines for the Care and Use of Laboratory Animals of Southwest Hospital of Army Medical University (Ref no. A-20,170,505) and the Guide for the Care and Use of Laboratory Animals (1985), NIH, Bethesda, MD, USA, or the European Guidelines on Laboratory Animal Care.
Colonic Motility Measurements
Colonic motility was assessed by measuring fecal pellet output numbers before and after SAP induction26 and detected 1 h prior to model induction and 4, 8, 12, 16, 20, and 24 h after model induction. Upon completing the measurements, the rats were euthanized with an overdose of pentobarbital sodium (200 mg/kg).
Measurement of Serum H2S Levels
Serum H2S levels were measured according to a previous study.27 Aliquots (75 μL) of sera were mixed with 100 μL distilled water and 300 μL 10% trichloroacetic acid. The reaction was stopped with 150 μL of 1% zinc acetate. N,N-dimethyl-p-phenylenediamine sulfate (20 μM) in 7.2 M HCl and FeCl3 (30 μM; 133 μL) in 1.2 M HCl were then added to the mixture and incubated for 15 min. The absorbance at 670 nm was measured, and the H2S concentration was calculated.
Detecting the Inflammatory Response
After the colonic motility measurements, the rats were euthanized with an overdose of pentobarbital sodium (200 mg/kg body weight), and plasma and colonic muscle tissue samples were collected. Tumor necrosis factor (TNF)-α (cat. no. H052) and interleukin (IL)-6 (cat. no. H007) production in the samples was detected using enzyme-linked immunosorbent assay (ELISA) kits (Nanjing Jiancheng Bio-engineering Institute Co., Ltd.) per the manufacturer’s instructions.
Western Blotting
Extracted protein was subjected to routine sodium dodecyl sulfate-polyacrylamide gel electrophoresis, then primary antibodies against CSE (1:500), t-AMPK (1:2000), p-AMPK (1:1000), t-mTOR (1:2000), p-mTOR (1:1000), and GAPDH (1:1000) were incubated on polyvinylidene difluoride membranes at 4°C overnight. After incubation with secondary horseradish peroxidase-conjugated IgG antibodies (1:5000), the relative protein expression levels were calculated using Gel-Pro-Analyzer (Media Cybernetics, Inc.).
Cell Preparation and Administration
CMCs were separated from the mucous membrane of each rat’s proximal colon and cultured in solution containing 0.15% collagenase II, 0.1% trypsin inhibitor, and 0.25% fetal bovine serum at 37°C. CMCs were identified via immunofluorescence detection of calponin and α-SMA (Figure S2), then treated with different combinations of 90 mM H2S solution,28 10 μg/mL C3G and 5 μmol/l MK-3903 for 24 h. Inflammatory responses in the CMCs were detected as described above.
Ca2+ Assay
CMCs (1×106) were subjected to repeated freezing and thawing to release the intracellular components. The suspension was then centrifuged at 3000 rpm for 20 min to collect the supernatant. The Ca2+ concentration was detected using an ELISA kit (Shanghai Keshun Biotechnology Co., Ltd.) per the manufacturer’s instructions.
Statistical Analysis
Data are presented as the mean ± standard deviation. One-way analysis of variance followed by Duncan’s post hoc multiple comparisons test were conducted. Statistical analyses were conducted, and graphs were created using GraphPad Prism 6 (GraphPad Prism Software, Inc.). P<0.05 was considered statistically significant.
Results
Administering C3G Improved Colonic Motility in SAP-Model Rats
The effect of C3G on colonic motility in the rats was assessed by measuring the fecal pellet output among the rats. Before inducing SAP, fecal pellet output numbers did not differ between the groups (sham vs SAP: P=0.993; sham vs SAP + C3G: P=0.991; SAP vs SAP + C3G: P=0.968; Figure 1). After inducing SAP, symptoms were induced via sodium taurocholate, and significant differences were detected between the sham and SAP groups after 4 h (P=0.04). C3G administration significantly alleviated colonic motility loss after 14 h (14 h: P=0.000; 24 h: P=0.000; Figure 1), indicating that C3G administration improved the colonic motility during SAP progression.
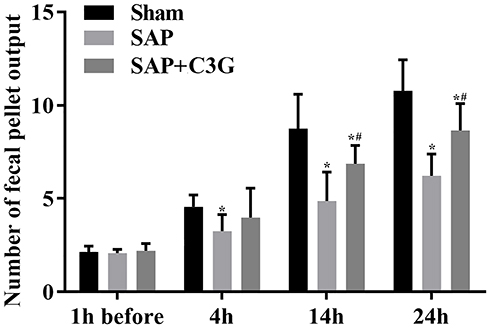 |
Figure 1 C3G administration improved colonic motility in SAP-model rats. *P<0.05 vs sham group; #P<0.05 vs SAP group. Abbreviations: C3G, cyanidin-3-O-glucoside; SAP, severe acute pancreatitis. |
C3G Suppressed the Inflammatory Response and H2S Levels in SAP-Model Rats
Inducing SAP increased the levels of H2S (P=0.001; Figure 2A), TNF-α (P=0.000) and IL-6 (P=0.000; Figure 2B and C). H2S suppressed the inflammatory response; however, its anti-inflammatory effect was offset by its negative effect on colonic motility. Thus, C3G administration inhibited both H2S and inflammatory cytokine production (H2S: P=0.048; TNF-α: P=0.006; IL-6: P=0.004; Figure 2).
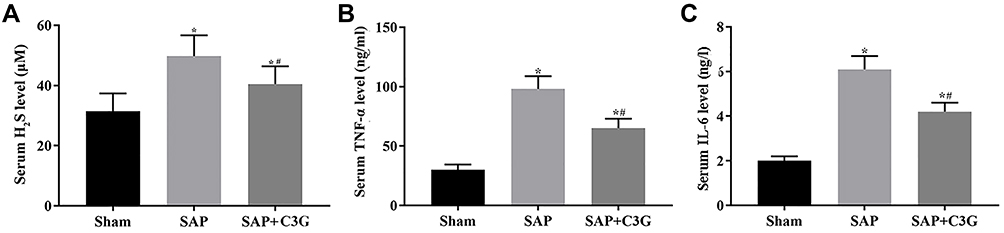 |
Figure 2 C3G administration suppressed H2S production and cytokine levels in SAP-model rats. Serum (A) H2S, (B) TNF-α and (C) IL-6 levels. *P<0.05 vs sham group; #P<0.05 vs SAP group. Abbreviations: C3G, cyanidin-3-O-glucoside; SAP, severe acute pancreatitis; TNF, tumor necrosis factor; IL, interleukin. |
Administering C3G Inhibited CSE Levels and Deactivated the AMPK/mTOR Signaling Pathway in SAP-Model Rats
To identify the mechanism by which SAP improved colonic motility, AMPK/mTOR pathway activity was detected. SAP increased the CSE expression (P=0.000) and activated the AMPK/mTOR pathway by increasing p-AMPK levels (P=0.000) and the p-AMPK/t-AMPK ratio (P=0.000) and suppressing p-mTOR levels (P=0.000) and the p-mTOR/t-mTOR ratio (P=0.000; Figure 3). Moreover, C3G reversed the expression patterns of these indicators (CSE: P=0.001; p-AMPK: P=0.014; p-mTOR: P=0.035; p-AMPK/t-AMPK ratio: P=0.003; p-mTOR/t-mTOR ratio: P=0.018); thus, the effect of C3G on colonic motility was associated with inhibiting the H2S/AMPK/mTOR pathway.
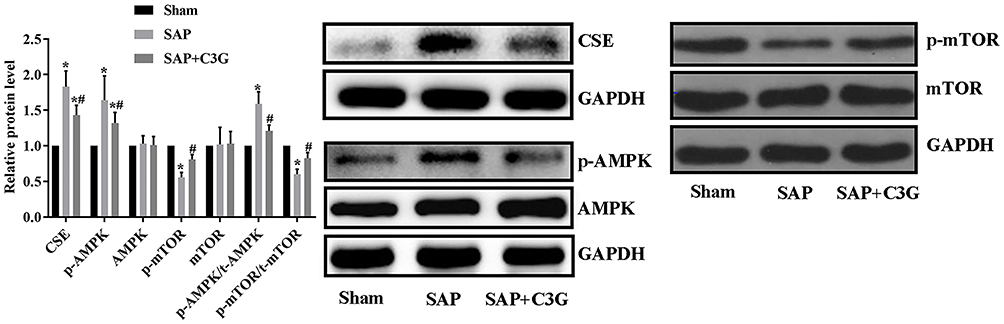 |
Figure 3 C3G administration suppressed CSE expression and deactivated the AMPK/mTOR pathway. *P<0.05 vs sham group; #P<0.05 vs SAP group. Abbreviations: C3G, cyanidin-3-O-glucoside; SAP, severe acute pancreatitis; CSE, cystathionine-γ-lyase. |
Activation of AMPK/mTOR Blocked the Protective Effect of C3G Against SAP Plasma-Induced CMC Impairments
To further assess the pathway underlying the effects of C3G, CMCs were isolated and subjected to H2S solution, C3G, and an AMPK activator in different combinations. H2S administration increased CSE expression (P=0.000) and activated AMPK/mTOR signaling in CMCs (p-AMPK: P=0.000; p-mTOR: P=0.002; p-AMPK/t-AMPK: P=0.000; p-mTOR/t-mTOR: P=0.011; Figure 4A), which also induced cytokine production (TNF-α: P=0.000; IL-6: P=0.000; Figure 4B and C) and suppressed Ca2+ accumulation (P=0.002; Figure 4D) in CMCs. C3G administration suppressed CSE expression (P=0.000) and AMPK/mTOR pathway activity (p-AMPK: P=0.000; p-mTOR: P=0.041; p-AMPK/t-AMPK: P=0.002; p-mTOR/t-mTOR: P=0.016), which contributed to inhibiting the inflammatory response (TNF-α: P=0.008; IL-6: P=0.000) and restored Ca2+ accumulation (P=0.011). However, the AMPK activator impaired the protective effect of C3G against SAP plasma, which reactivated the AMPK/mTOR pathway by increasing p-AMPK (P=0.001) and p-AMPK/t-AMPK (P=0.004) levels but suppressing p-mTOR (P=0.043) and p-mTOR/t-mTOR (P=0.024) levels, which also contributed to increasing the CSE level (P=0.029; Figure 4A). These changes in CSE and AMPK/mTOR signaling increased TNF-α (P=0.041) and IL-6 (P=0.023; Figure 4B and C) production and decreased Ca2+ production (P=0.025; Figure 4D). Thus, the protective effect of C3G against SAP-induced colonic motility loss depended on H2S-mediated AMPK/mTOR pathway inhibition.
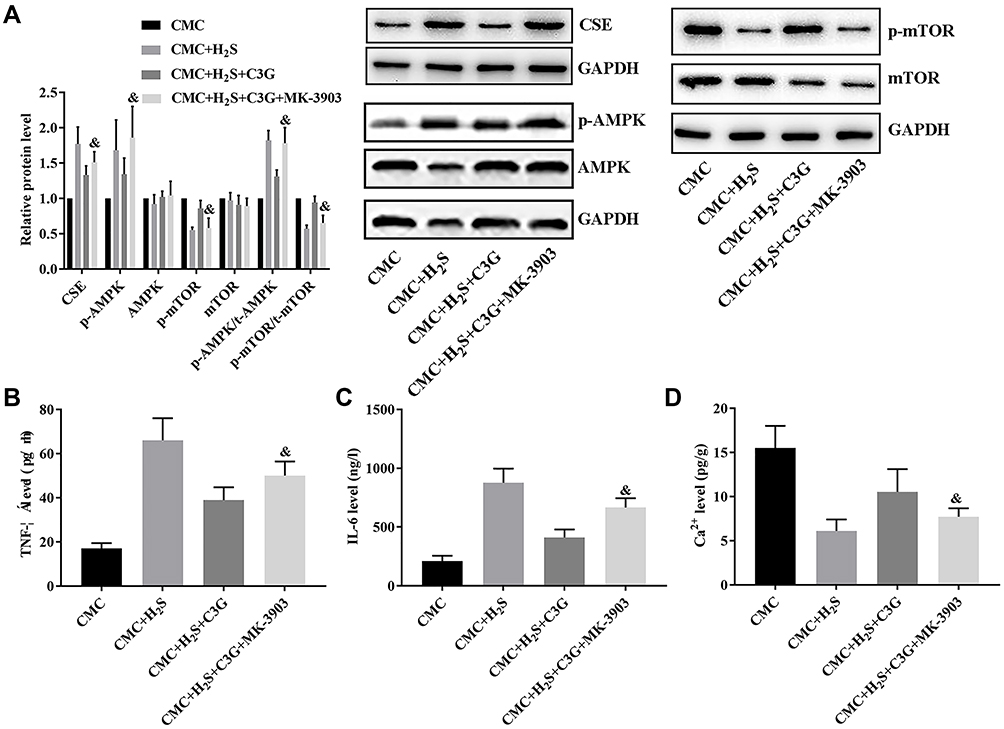 |
Figure 4 AMPK activation offset the effect of C3G on H2S-treated CMCs. (A) CSE levels and AMPK activity, (B) TNF-α levels, (C) serum IL-6 levels, (D) Ca2+ accumulation. &P<0.05 vs CMC + H2S + C3G group. |
Discussion
H2S is the third member of gasotransmitter family synthesized endogenously via the transsulfuration pathway, which is an important mechanism for providing cells with cysteine.29,30 Being increasingly recognized as a functionally relevant mediator of a number of physiological functions, deficiencies in the H2S production can cause a chronic inflammatory response by inducing pro-inflammatory molecule production, thus resulting in development of various diseases.29 Regarding the protective effects on GI system, H2Scan decrease production of TNF-α and leukocytes.15 However, the Being increasingly recognized as a functionally relevant mediator of a number of physiological functions. Tamizhselvi et al31 revealed that H2S induced inflammation in AP rats. Therefore, the functions and related mechanisms of H2S in GI diseases should be assessed.
Consistent with previous studies,16,32,33 SAP symptoms initiated CSE synthesis and increased H2S levels. The enhanced release of H2S should be associated with a weakened inflammatory response, but it seemed that the anti-inflammation effects of H2S were blocked by its and suppressive effects on ed colonic motility during SAP progression. However, C3G administration suppressed the plasma cytokine levels and improved suppressed colonic motility in SAP rats. Therefore, an interaction was hypothesized to have occurred among C3G, H2S, and inflammation: H2S exerted an anti-inflammatory effect during SAP progression, but its positive effect was offset by its negative effect on colonic motility in SAP rats. C3G administration improved colonic motility by suppressing H2S production. In the meanwhile, the anti-inflammatory effects of C3G compensated for the lack in anti-inflammatory factors induced by the deficient production of H2S. Thus, applying C3G as a treatment agent for impairments associated with SAP not only improved the colonic motility loss but also contributed to the control of inflammatory response.
H2S regulates multiple pathways. In the present study, activity of the AMPK/mTOR pathway was detected to examine the signaling pathway mediating the effect of C3G. The results indicated that SAP and H2S solution induced AMPK/mTOR pathway activity both in vivo and in vitro, whereas C3G inhibited this activity. In addition, the CMCs were also treated with AMPK activator MK-3903. The set of the MK-3903 group was employed to validate that the effects of C3G were dependent on the inhibition of AMPK/mTOR pathway. Activation of the AMPK/mTOR pathway in CMCs impaired the protective effect of C3G against H2S, increased the cytokine production, and inhibited Ca2+ accumulation in CMCs, indicating suppressed motility potential in the cells. The results clearly demonstrated the inhibition of AMPK/mTOR pathway was indispensable for the protective effects of C3G on SAP-induced colonic motility loss. Thus, the changes in AMPK/mTOR pathway partially explained the hypothesis we proposed above: the interaction between C3G and H2S influenced the activation of AMPK signaling, which finally led to the improved colonic motility and suppressed inflammatory responses associated with SAP initiation.
In conclusion, the in vivo and in vitro assay results demonstrated that administering C3G increased colonic motility in rats by suppressing H2S production. Moreover, the effect of C3G depended on the H2S-mediated AMPK/mTOR pathway. H2S administration and AMPK activation impaired the motility potential of CMCs, even after C3G administration. However, the present study examined only the downstream pathways involved in the protective effect of C3G against SAP on the GI system. Therefore, further studies are required to improve our understanding of the mechanisms underlying C3G functions.
Data Sharing Statement
Data will be provided when required.
Ethics Approval
All animal experiments were conducted in accordance with the Institutional Animal Ethics Committee and Animal Care Guidelines for the Care and Use of Laboratory Animals of Southwest Hospital of Army Medical University (Ref No. A-20,170,505) and with the Guide for the Care and Use of Laboratory Animals (1985), NIH, Bethesda, MD, USA, or the European Guidelines on Laboratory Animal Care.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no competing interests for this work.
References
1. Bollen TL. Acute pancreatitis: international classification and nomenclature. Clin Radiol. 2016;71(2):121–133. doi:10.1016/j.crad.2015.09.013
2. Leppaniemi A, Tolonen M, Tarasconi A, et al. 2019 WSES guidelines for the management of severe acute pancreatitis. World J Emerg Surg. 2019;14:27.
3. Portelli M, Jones CD. Severe acute pancreatitis: pathogenesis, diagnosis and surgical management. Hepatobiliary Pancreat Dis Int. 2017;16(2):155–159. doi:10.1016/S1499-3872(16)60163-7
4. Huang L, Jiang Y, Sun Z, Gao Z, Wang J, Zhang D. Autophagy strengthens intestinal mucosal barrier by attenuating oxidative stress in severe acute pancreatitis. Digest Dis Sci. 2018;63(4):910–919. doi:10.1007/s10620-018-4962-2
5. Wen W, Zheng H, Jiang Y, et al. Effect of intestinal epithelial autophagy on bacterial translocation in severe acute pancreatitis. Clin Res Hepatol Gas. 2017;41(6):703–710. doi:10.1016/j.clinre.2017.03.007
6. Piao X, Liu B, Guo L, Meng F, Gao L. Picroside II shows protective functions for severe acute pancreatitis in rats by preventing NF-kappaB-dependent autophagy. Oxid Med Cell Longev. 2017;7085709(10):21.
7. Xiong J, Wang K, Yuan C, et al. Luteolin protects mice from severe acute pancreatitis by exerting HO-1-mediated anti-inflammatory and antioxidant effects. Int J Mol Med. 2017;39(1):113–125. doi:10.3892/ijmm.2016.2809
8. Liu MW, Wei R, Su MX, Li H, Fang TW, Zhang W. Effects of panax notoginseng saponins on severe acute pancreatitis through the regulation of mTOR/Akt and caspase-3 signaling pathway by upregulating miR-181b expression in rats. BMC Complement Altern Med. 2018;18(1):018–2118. doi:10.1186/s12906-018-2118-8
9. Ma Z, Song G, Liu D, et al. N-acetylcysteine enhances the therapeutic efficacy of bone marrow-derived mesenchymal stem cell transplantation in rats with severe acute pancreatitis. Pancreatology. 2019;19(2):258–265. doi:10.1016/j.pan.2019.01.004
10. Xiao Q, Ying J, Xiang L, Zhang C. The biologic effect of hydrogen sulfide and its function in various diseases. Medicine. 2018;97(44):0000000000013065. doi:10.1097/MD.0000000000013065
11. Wu DD, Wang DY, Li HM, Guo JC, Duan SF, Ji XY. Hydrogen sulfide as a novel regulatory factor in liver health and disease. Oxid Med Cell Longev. 2019;20(3831713).
12. Wen YD, Wang H, Zhu YZ. The drug developments of hydrogen sulfide on cardiovascular disease. Oxid Med Cell Longev. 2018;2018:4010395. doi:10.1155/2018/4010395
13. Dombkowski RA, Naylor MG, Shoemaker E, et al. Hydrogen sulfide (H2S) and hypoxia inhibit salmonid gastrointestinal motility: evidence for H2S as an oxygen sensor. J Exp Biol. 2011;214(23):4030–4040. doi:10.1242/jeb.061473
14. Farrugia G, Szurszewski JH. Carbon monoxide, hydrogen sulfide, and nitric oxide as signaling molecules in the gastrointestinal tract. Gastroenterology. 2014;147(2):303–313. doi:10.1053/j.gastro.2014.04.041
15. Wallace JL, Caliendo G, Santagada V, Cirino G, Fiorucci S. Gastrointestinal safety and anti-inflammatory effects of a hydrogen sulfide-releasing diclofenac derivative in the rat. Gastroenterology. 2007;132(1):261–271. doi:10.1053/j.gastro.2006.11.042
16. Tamizhselvi R, Moore PM. Hydrogen sulfide acts as a mediator of inflammation in acute pancreatitis: in vitro studies using isolated mouse pancreatic acinar cells. J Cell Mol Med. 2007;11(2):12. doi:10.1111/j.1582-4934.2007.00024.x
17. Vomero ND, Colpo E. Nutritional care in peptic ulcer. Arq Bras Cir Dig. 2014;27(4):298–302. doi:10.1590/S0102-67202014000400017
18. Sandoval-Ramirez BA, Catalan U, Fernandez-Castillejo S, Rubio L, Macia A, Sola R. Anthocyanin tissue bioavailability in animals: possible implications for human health. A systematic review. J Agr Food Chem. 2018;66(44):11531–11543. doi:10.1021/acs.jafc.8b04014
19. Ming-Ming M, Yan L, Xiang-Yong L, et al. Cyanidin-3-O-glucoside ameliorates lipopolysaccharide-induced injury both in vivo and in vitro suppression of NF-κB and MAPK pathways. Inflammation. 2015;38(4):1669–1682. doi:10.1007/s10753-015-0144-y
20. Jung-Min K, Kyung-Mi K, En-Hee P, et al. Anthocyanins from black soybean inhibit Helicobacter pylori-induced inflammation in human gastric epithelial AGS cells. Microbiol Immunol. 2013;57(5):366–373. doi:10.1111/1348-0421.12049
21. Kalt W, Cassidy A, Howard LR, et al. Recent research on the health benefits of blueberries and their anthocyanins. Adv Nutr. 2019;22(5536953).
22. Antonio S, Sirajudheen A, Raffaella C, et al. Cyanidin-3-O-glucoside counters the response to TNF-alpha of endothelial cells by activating Nrf2 pathway. Mol Nutr Food Res. 2013;57(11):1979–1987. doi:10.1002/mnfr.201300102
23. Wang Q, Xia M, Liu C, et al. Cyanidin-3-O-beta-glucoside inhibits iNOS and COX-2 expression by inducing liver X receptor alpha activation in THP-1 macrophages. Life Sci. 2008;83(5):176–184. doi:10.1016/j.lfs.2008.05.017
24. Speciale A, Canali R, Chirafisi J, Saija A, Virgili F, Cimino F. Cyanidin-3-O-glucoside protection against TNF-alpha-induced endothelial dysfunction: involvement of nuclear factor-kappaB signaling. J Agr Food Chem. 2010;58(22):12048–12054. doi:10.1021/jf1029515
25. Wang H, Zhong P, Sun L. Exogenous hydrogen sulfide mitigates NLRP3 inflammasome-mediated inflammation through promoting autophagy via the AMPK-mTOR pathway. Biol Open. 2019;8(7):bio043653. doi:10.1242/bio.043653
26. Murakami T, Kamada K, Mizushima K, et al. Changes in intestinal motility and gut microbiota composition in a rat stress model. Digestion. 2017;95(1):55–60. doi:10.1159/000452364
27. Kang K, Zhao M, Jiang H, Tan G, Pan S, Sun X. Role of hydrogen sulfide in hepatic ischemia-reperfusion-induced injury in rats. Liver Transpl. 2009;15(10):1306–1314. doi:10.1002/lt.21810
28. Tang G, Wu L, Liang W, Wang R. Direct stimulation of K ATP channels by exogenous and endogenous hydrogen sulfide in vascular smooth muscle cells. Mol Pharmacol. 2005;68(6):1757–1764. doi:10.1124/mol.105.017467
29. Rosado JO, Salvador M, Bonatto D. Importance of the trans-sulfuration pathway in cancer prevention and promotion. Mol Cell Biochem. 2007;301(1–2):1–12. doi:10.1007/s11010-006-9389-y
30. Tessari P, Cecchet D, Vettore M, Coracina A, Puricelli L, Kiwanuka E. Decreased homocysteine trans-sulfuration in hypertension with hyperhomocysteinemia: relationship with insulin resistance. J Clin Endocrinol Metab. 2018;103(1):56–63. doi:10.1210/jc.2017-01076
31. Tamizhselvi R, Koh YH, Sun J, Zhang H, Bhatia M. Hydrogen sulfide induces ICAM-1 expression and neutrophil adhesion to caerulein-treated pancreatic acinar cells through NF-kappaB and Src-family kinases pathway. Exp Cell Res. 2010;316(9):1625–1636. doi:10.1016/j.yexcr.2010.02.044
32. Hegde A, Bhatia M. Hydrogen sulfide in inflammation: friend or foe? Inflamm Allergy Drug Targets. 2011;10(2):118–122. doi:10.2174/187152811794776268
33. Whiteman M, Winyard PG. Hydrogen sulfide and inflammation: the good, the bad, the ugly and the promising. Expert Rev Clin Pharmacol. 2011;4(1):13–32. doi:10.1586/ecp.10.134
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.