")
Back to Journals » Nature and Science of Sleep » Volume 12
Finger Twitches are More Frequent in REM Sleep Than in Non-REM Sleep
Authors Reiter AM , Roach GD , Sargent C , Lack L
Received 4 October 2019
Accepted for publication 17 December 2019
Published 23 January 2020 Volume 2020:12 Pages 49—56
DOI https://doi.org/10.2147/NSS.S233439
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Steven A Shea
Andrew M Reiter,1 Gregory D Roach,1 Charli Sargent,1 Leon Lack2
1Appleton Institute for Behavioural Science, Central Queensland University, Goodwood, SA 5034, Australia; 2College of Education, Psychology and Social Work, Flinders University of South Australia, Adelaide, SA 5001, Australia
Correspondence: Andrew M Reiter
Appleton Institute for Behavioural Science, Central Queensland University, PO Box 42, Goodwood, SA 5034, Australia
Tel +61 8 8378 4523
Email [email protected]
Introduction: Abnormal rapid eye movement (REM) sleep is often symptomatic of chronic disorders, however polysomnography, the gold standard method to measure REM sleep, is expensive and often impractical. Attempts to develop cost-effective ambulatory systems to measure REM sleep have had limited success. As elevated twitching is often observed during REM sleep in some distal muscles, the aim of this study was to assess the potential for a finger-mounted device to measure finger twitches, and thereby differentiate periods of REM and non-REM (NREM) sleep.
Methods: One night of sleep data was collected by polysomnography from each of 18 (3f, 15m) healthy adults aged 23.2 ± 3.3 (mean ± SD) years. Finger movement was detected using a piezo-electric limb sensor taped to the index finger of each participant. Finger twitch densities were calculated for each stage of sleep.
Results: Finger twitch density was greater in REM than in NREM sleep (p < 0.001). Each sleep stage had a unique finger twitch density, except for REM and stage N1 sleep which were similar. Finger twitch density was greater in late REM than in early REM sleep (p = 0.005), and there was a time–state interaction: the difference between finger twitch densities in REM and NREM sleep was greater in late sleep than in early sleep (p = 0.007).
Conclusion: Finger twitching is more frequent in REM sleep than in NREM sleep and becomes more distinguishable as sleep progresses. Finger twitches appear to be too infrequent to make definitive 30-second epoch determinations of sleep stage. However, an algorithm informed by measures of finger twitch density has the potential to detect periods of REM sleep and provide estimates of total REM sleep time and percentage.
Keywords: finger, twitch, REM sleep, actigraphy, wearable
Introduction
Since the discovery of rapid eye movements (REMs),1 sleep has been dichotomised as non-REM (NREM) sleep, which shows the restfulness normally associated with sleep, or REM sleep, characterised by REMs, skeletal muscle inhibition and cortical activation similar to wakefulness.2 REM sleep is suggested to be important for memory consolidation3 and development,4 and abnormal REM sleep patterns are symptomatic of chronic disorders e.g., depression5 and narcolepsy.6 The gold standard for objective measurement of REM sleep is polysomnography (PSG) which is expensive and has practical limitations.7,8 Several alternative techniques have been investigated to measure REM sleep,9,10 with limited success. In 2017, Fitbit11 introduced the capability to differentiate light, deep and REM sleep in wrist-worn actigraphy devices, but with a per-epoch accuracy of 69% compared with PSG,12 the search for an accurate, cost-effective ambulatory system to measure REM sleep continues.
The purpose of this study was to assess the potential for a simple finger-mounted device to measure finger twitches and thereby differentiate periods of REM and NREM sleep. Twitches are short-duration, involuntary muscle contractions, which when observed in the paws, tails, ears and whiskers of sleeping pets are often attributed to the acting out of dreams. Researchers have consistently reported elevated levels of twitching in the distal muscles of both animals and humans during REM sleep, and particularly late REM sleep.6,13–19 Studies of sleep twitches in adult humans have focused mainly on facial and chin muscles: Rivera‐García, Ramírez‐Salado, Corsi‐Cabrera, Calvo13 reported greater facial muscle contractions during late REM sleep than early REM sleep, and De Gennaro, Ferrara, Bertini15 reported the same temporal pattern for chin muscles. Similar differences in eye, throat, wrist and ankle fine muscle activity between REM sleep and NREM sleep were reported by Baldridge, Whitman, Kramer.20 To detect REM sleep, measurement of finger twitches would appear to be more practical and less invasive to sleep than the measurement of twitches in the face or chin. However, only one study of finger twitching during sleep in adult humans was identified: Stoyva19 examined finger activity using electrodes attached to the forearms of deaf and hearing participants during sleep, and reported that activity was greater in REM sleep than in NREM sleep, and greater in later REM sleep episodes than in the first REM sleep episode. The aim of the present study was, therefore, to determine whether finger twitching measured by a more practical and less invasive finger-mounted device would be greater in REM sleep than in NREM sleep (REM-NREM effect) and be greater in late REM sleep than in early REM sleep (time-of-night effect).
Materials and Methods
Participants
Participants were 18 (3 females, 15 males) young, healthy adults aged between 18 and 30 years (M = 23.22 years, SD = 3.26 years). Screening exclusion criteria included smoking, excessive caffeine consumption, excessive alcohol consumption, physical and mental disorders, irregular sleep patterns and shiftwork or transmeridian travel in the prior month. The Central Queensland University Human Research Ethics Committee approved the study and all participants provided written informed consent. The study was carried out in accordance with the principles of the Declaration of Helsinki.
Materials
Data were collected at the Appleton Institute for Behavioural Science, Central Queensland University, Adelaide. Each participant was accommodated within their own bedroom and bathroom in a sound-attenuated, windowless sleep laboratory. Sleep was monitored by standard PSG, using the Grael PSG/EEG Systems (Compumedics, Melbourne, Australia) and a montage of GrassTM gold-cup electrode leads (Astro-Med, Inc., West Warwick, USA). The montage included two electroencephalography (EEG) channels (C3-M2, C4-M1), right and left electrooculography (EOG), and three channels of chin electromyography (EMG). A standard sleep study limb movement sensor was used to detect finger movement. Pilot tests demonstrated that a limb sensor (Compumedics Part Number: 7012-0009-01) taped to the index finger could reliably detect small finger movements and could be readily interfaced to the PSG system.
Design
An observational study design was used. Consistent with the way REM density is used to identify REM sleep episodes,21,22 finger twitch density (in twitches per minute) was the dependent variable for the present study. The first independent variable was sleep stage: wakefulness, REM sleep, stage N1, stage N2 and stage N3. For the purposes of some analyses, the sleep stages were rationalised to three states: wakefulness, REM sleep and NREM sleep. The second independent variable was time: each sleep opportunity was divided into thirds (early sleep, middle sleep and late sleep).
Procedure
PSG electrodes were connected to each participant and a finger sensor was taped to the participants’ distal index finger joint, using adhesive electrode tape. Sleep and finger twitches were measured simultaneously by PSG during nine-hour sleep opportunities (23:00–08:00). All epochs of PSG sleep data were scored blind to finger movement data using Profusion Compumedics software (version 3.0) by a trained sleep technologist according to the AASM manual for scoring sleep.23
Several methods for scoring sleep twitches have been reported. Stoyva19 identified only discrete bursts of finger activity (rather than sustained background tonus) above 20μV on the EMG and scored each burst by its duration in seconds, with any fraction of a second counted as one full second, and ignored all activity associated with awakenings and within 5 s before or after gross body movements. Rivera‐García, Ramírez‐Salado, Corsi‐Cabrera, Calvo13 defined any waveforms exceeding baseline by 500% as facial muscle contractions, which were categorised as phasic (1–100ms) or sustained (>100ms). The phasic and sustained events were summed to calculate the relevant twitch densities. As the present study used a piezo-electric sensor rather than EMG to detect movement, a study-specific scoring system was devised based on some of these principles. Events were counted if they were noticeably different (at least 200% greater in amplitude) from the waveform immediately preceding it. All events were marked, irrespective of duration, as they were to be subsequently categorised by duration. A period of at least 3 s of stable waveform at reduced amplitude was needed before a new finger event was marked. All finger movements were manually scored by the primary researcher blind to the other PSG channels, and independently of the sleep technologist who scored the sleep. Each 30-second epoch was reviewed, and each valid finger event was marked. All finger events were assigned to the epoch in which the event commenced. For each participant, the first ten epochs (5 mins) after lights out were ignored due to high levels of settling movement, and the remaining 1070 epochs (8 hrs and 55 mins) were reviewed.
Results
The key sleep variables for the twitch measurement period are presented in Table 1. A total of 19,260 epochs of sleep were reviewed, in which a total of 3314 finger events were marked. A simple duration-based classification system for finger events was applied, resulting in the frequency distribution presented in Figure 1. For finger events shorter than 5 s, half-second categories were used; for finger events with durations between five and 10 s, one-second categories were used. Finger events with durations longer than 10 s were allocated to a single category.
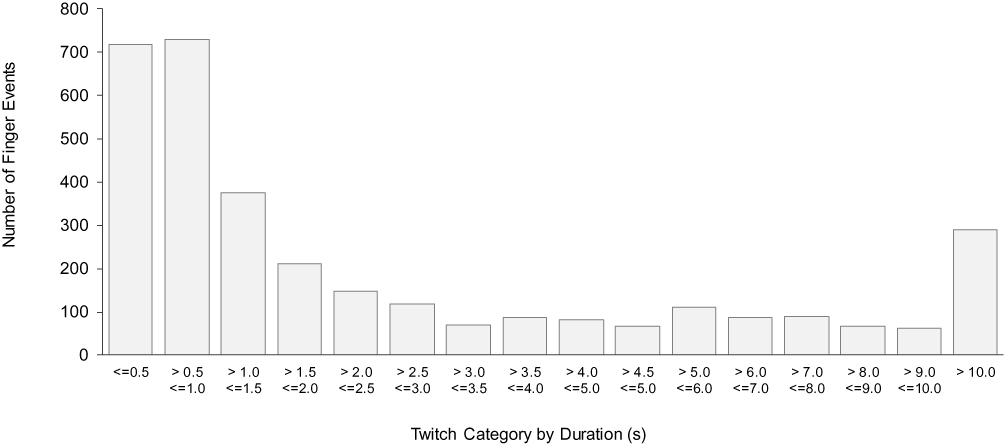 |
Figure 1 Frequency distribution of all finger movement events by duration category. |
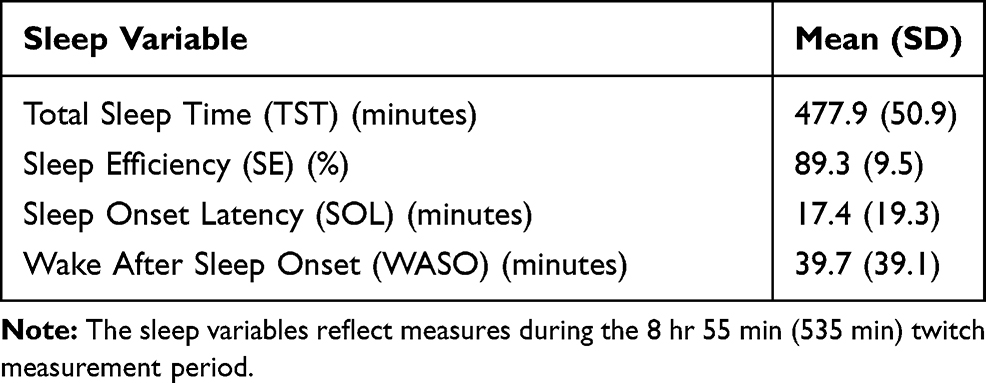 |
Table 1 Mean (SD) Sleep Variables for the Twitch Measurement Period |
The frequency distribution revealed most finger events were of short duration, with 44% of events less than or equal to 1 s in duration, and 70% of events less than or equal to 3 s in duration. As sleep disturbances greater than 3 s in duration are generally scored as arousals, finger events with duration less than or equal to 3 s were designated as finger twitches for the present study. This definition meant that longer events, such as those associated with body movements, were not counted as twitches. Under this definition, 2302 finger events were classified as twitches. Each finger twitch was matched with its sleep stage for the corresponding epoch and twitch densities were calculated by dividing the number of finger twitches in a particular stage by the corresponding duration in that stage in minutes. The key descriptive statistics for wakefulness, NREM sleep, REM sleep and the twitch measurement period are presented in Table 2. There was considerable inter-individual variability in the overall twitch densities (range: 0.07 to 0.72 twitches per minute) and in the twitch densities in wakefulness (range: 0.15 to 2.33 twitches per minute), NREM sleep (range: 0.04 to 0.73 twitches per minute) and REM sleep (range: 0.05 to 0.83 twitches per minute). Overall twitch density was correlated with twitch density in REM sleep (r = 0.79, p < 0.001) and NREM sleep (r = 0.84, p < 0.001), but not with twitch density during wakefulness (r = 0.32, p = 0.19). Twitch density in NREM sleep and REM sleep were also correlated (r = 0.66, p = 0.003).
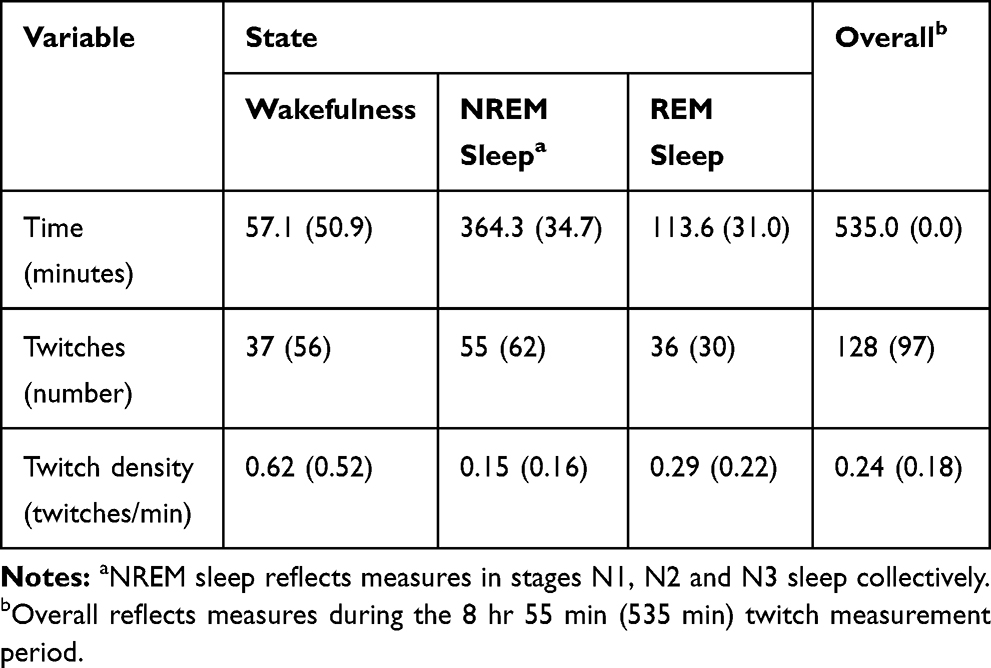 |
Table 2 Mean (SD) Times, Twitches and Twitch Densities During Wakefulness, NREM Sleep, REM Sleep and the Twitch Measurement Period |
Examination of the density frequency distributions indicated most were positively skewed, and Kolmogorov–Smirnov tests confirmed many were also non-normal. All twitch densities were therefore scaled and transformed [log(X + 1)] prior to analysis, consistent with the approach of De Gennaro, Ferrara, Bertini.15 All descriptive statistics and effect sizes presented in this section reflect untransformed densities, whereas the inferential statistics reflect the results of tests on transformed densities.
REM-NREM Effect
A one-way repeated measures ANOVA was performed on transformed twitch densities for wakefulness, NREM sleep and REM sleep. Mauchly’s test indicated the assumption of sphericity was violated, χ2(2) = 6.70, p = 0.035, so degrees of freedom were corrected using the Greenhouse-Geisser estimates of sphericity (ε = 0.75). The ANOVA revealed a significant omnibus effect, F(1.49, 25.33) = 29.15, p < 0.001. A post hoc pairwise comparison showed that REM sleep twitch density was greater than NREM sleep twitch density (p < 0.001) and the effect size was medium-large (d = 0.73), suggesting an overall REM-NREM effect. The mean densities provided in Table 2 and the one-way ANOVA results suggested wakefulness twitch density may also be greater than REM sleep twitch density, and a post hoc pairwise comparison revealed wakefulness twitch density was greater than REM sleep twitch density (p = 0.007); the effect size was large (d = 0.83). In summary, the mean twitch densities during wakefulness, NREM sleep and REM sleep were distinguishable, and the corresponding ratio was approximately 4:1:2.
Finger twitch density in REM sleep was also compared to finger twitch density in each NREM sleep stage. The descriptive statistics for NREM sleep stages N1, N2 and N3 are presented in Table 3. A one-way repeated measures ANOVA was performed on transformed twitch densities in wakefulness, REM sleep, stage N1, stage N2 and stage N3. Mauchly’s test indicated the assumption of sphericity was violated, χ2(9) = 19.49, p = 0.022, so degrees of freedom were corrected using the Greenhouse-Geisser estimates of sphericity (ε = 0.64). The ANOVA revealed a significant omnibus effect, F(2.54, 43.23) = 37.67, p < 0.001. Post hoc pairwise comparisons revealed no difference between REM sleep twitch density and stage N1 twitch density (p = 1.00). REM sleep twitch density was greater than stage N2 twitch density (p = 0.001); the effect size was medium-large (d = 0.64). REM sleep twitch density was also greater than stage N3 twitch density (p < 0.001); the effect size was large (d = 1.15).
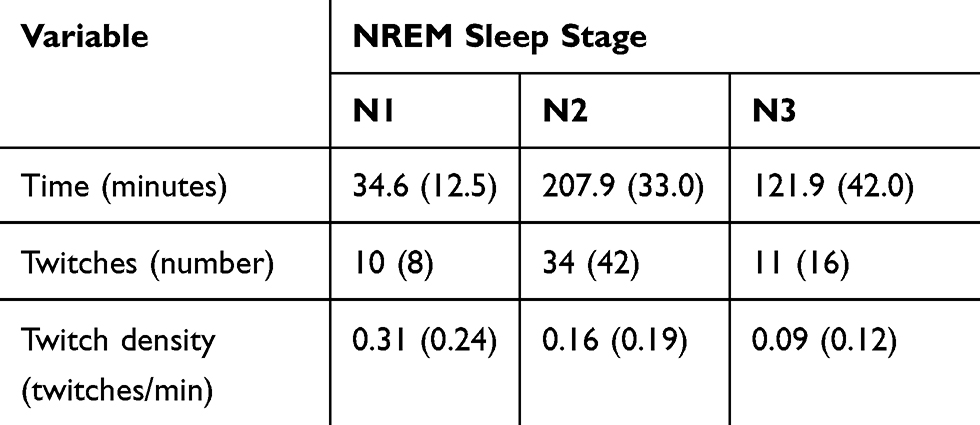 |
Table 3 Mean (SD) Times, Twitches and Twitch Densities During NREM Sleep Stages N1, N2 and N3 |
Post hoc pairwise comparisons revealed stage N1 twitch density was greater than stage N2 twitch density (p < 0.001); the effect size was large (d = 0.72). Stage N2 twitch density was greater than stage N3 twitch density (p < 0.001); the effect size was medium (d = 0.44). In summary, four of the five sleep stages had distinguishable twitch densities as illustrated in Figure 2. Wakefulness twitch density was greater than REM sleep twitch density and stage N1 twitch density, which were similar. REM sleep and stage N1 twitch densities were greater than stage N2 twitch density, which in turn was greater than stage N3 sleep twitch density.
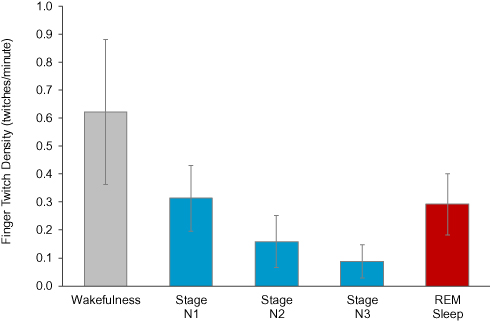 |
Figure 2 Mean twitch densities in wakefulness, NREM sleep stages N1, N2 and N3, and REM sleep. Error bars represent the 95% confidence intervals of the means. |
Time-of-Night Effect
Each sleep opportunity was divided approximately into thirds: early sleep, middle sleep and late sleep. Early sleep represents 2h:55m of sleep opportunity, whereas middle and late sleep represent 3h:00m of sleep opportunity. Each third was likely to include two sleep cycles and at least one episode of REM sleep. The mean twitch densities for each third of sleep for each state are presented in Table 4.
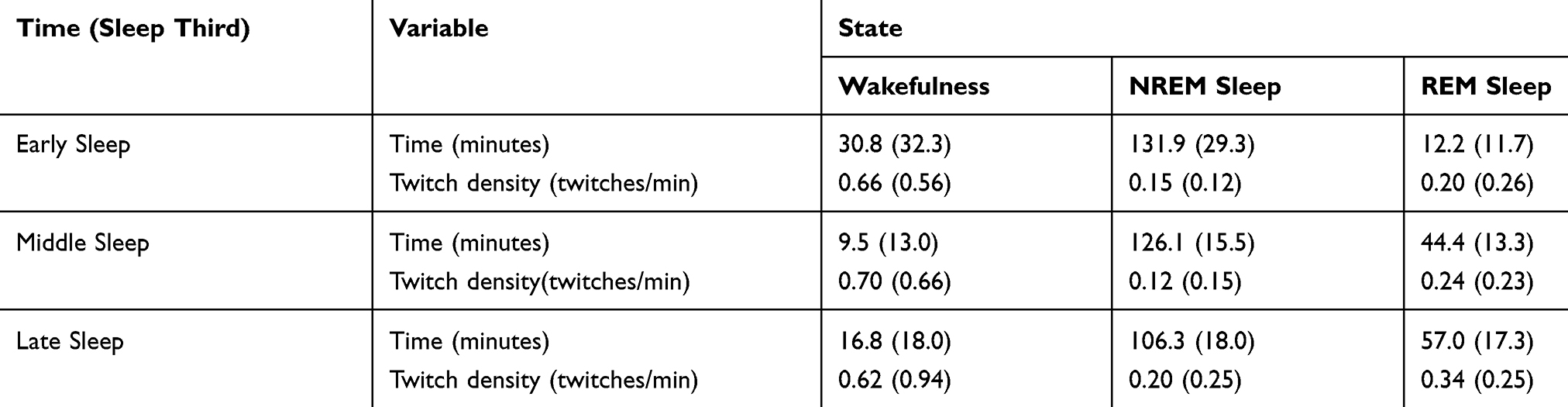 |
Table 4 Mean (SD) Times, Twitch Densities (Twitches/Minute) as Function of Time and State |
A 3 (time) x 3 (state) repeated measures factorial ANOVA was performed on transformed twitch densities. Mauchly’s test indicated the assumption of sphericity was not violated for the main effect of time (χ2(2) = 2.05, p = 0.359), but was violated for the main effect of state (χ2(2) = 7.60, p = 0.022) and the interaction (χ2(9) = 26.47, p = 0.002). Degrees of freedom were therefore corrected using Greenhouse-Geisser estimates of sphericity for the main effect of state (ε = 0.72) and the interaction (ε = 0.67). The main effect of time was not significant, F(2,34) = 0.76, p = 0.475, indicating twitch densities collapsed across all states were not different between each third of sleep. The main effect of state was significant, F(1.45,24.62) = 10.18, p = 0.002, confirming twitch densities collapsed across time were different for wakefulness, NREM sleep and REM sleep. A post hoc pairwise comparison revealed REM sleep twitch density in late sleep was greater than REM sleep twitch density in early sleep (p = 0.005); the effect size was medium (d = 0.55), providing support for the time-of-night effect. Pairwise comparisons revealed no difference between wakefulness twitch density in early sleep and wakefulness twitch density in late sleep (p = 0.329), or between NREM sleep twitch density in early sleep and NREM sleep twitch density in late sleep (p = 1.000).
REM sleep twitch density was compared between early and middle sleep, and middle and late sleep. Pairwise comparisons showed no difference between REM sleep twitch density in early sleep and REM sleep twitch density in middle sleep (p = 0.051), and no difference between REM sleep twitch density in middle sleep and REM sleep twitch density in late sleep (p = 0.071). Although the early-middle sleep and middle-late sleep pairwise comparisons were not significant, a linear trend analysis confirmed a significant linear increase in REM sleep twitch density from early to middle to late sleep, F(1,17) = 14.18, p = 0.002; the effect size was large, η2 = 0.46. This indicates that as the night progressed, REM sleep twitch density increased proportionately. There was no corresponding linear trend for wakefulness twitch density, F(1,17) = 2.85, p = 0.110, or for NREM sleep twitch density F(1,17) = 0.11, p = 0.740.
The ANOVA also revealed a significant interaction, F(2.67, 45.65) = 5.62, p = 0.001, and simple contrasts were performed to examine interactions of interest. The difference between REM sleep twitch density and NREM sleep twitch density was greater in late sleep than in early sleep, F(1,17) = 9.30, p = 0.007; the effect size for this interaction was large (η2 = 0.35), suggesting time moderated the difference between REM sleep twitch density and NREM sleep twitch density. The difference between REM sleep twitch density and NREM sleep twitch density was greater in middle sleep than in early sleep, F(1,17) = 12.28, p = 0.003; the effect size for this interaction was large (η2 = 0.42). The difference between REM sleep twitch density and NREM sleep twitch density was not different between middle sleep and late sleep, F(1,17) = 0.003, p = 0.959. These results suggest that the increase in difference between REM sleep twitch density and NREM sleep twitch density from early sleep to late sleep occurred mainly from early sleep to middle sleep, and that the twitch density difference was maintained from middle sleep to late sleep, as illustrated in Figure 3.
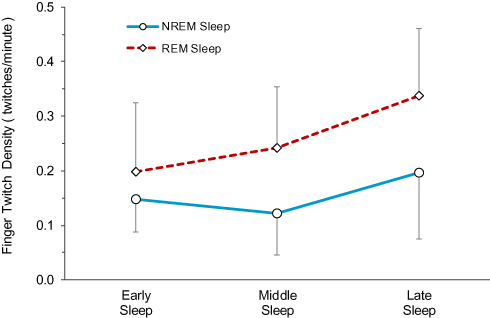 |
Figure 3 Mean twitch densities in REM sleep and NREM sleep in early, middle and late sleep. Error bars represent the 95% confidence intervals of the means. |
Discussion
The results of this study provide evidence that the REM-NREM and time-of-night effects for finger twitching can be measured using a finger-mounted device. The mean finger twitch densities during wakefulness, NREM sleep and REM sleep were 0.62, 0.15 and 0.29 twitches per minute. The mean finger twitch densities during wakefulness, NREM sleep and REM sleep were distinguishable from each other, but could not be readily compared with previous research, due to the different muscle groups studied, the range of methods used to measure twitching, and the variety of twitch scoring definitions. However, the 2:1 ratio of finger twitch density in REM sleep to finger twitch density in NREM sleep is consistent with the 2.3:1 ratio reported by Stoyva.19 Finger twitching in four of the five stages of sleep was distinguishable, but again there are limited reference studies. Stoyva19 did not report these comparisons, but Rivera‐García, Ramírez‐Salado, Corsi‐Cabrera, Calvo13 reported a similar hierarchy for facial muscle twitches.
As the night progressed, finger twitch density in REM sleep increased relative to finger twitch density in NREM sleep. The time-of-night effect for REM sleep finger twitching found in the present study is consistent with the temporal patterns reported in other REM sleep muscle twitching and eye movement studies. De Gennaro, Ferrara, Bertini15 reported a time-of-night effect for eye movements and chin muscle twitches, and Stoyva19 reported the effect for finger activity. No time-of-night effect was found for finger twitching in either wakefulness or NREM sleep in the present study. Stoyva19 noted there was no overall difference in the rate of finger activity in NREM sleep between early and late sleep, but that this varied between individuals. No reference to a time-of-night effect for twitching in periods of wakefulness during sleep was identified in any previous research. Analysis of the time–state interactions showed the REM-NREM effect was greater in late sleep than in early sleep. Most of the increase in the REM-NREM effect occurred from early to middle sleep, and this increase was then maintained through to late sleep.
From a theoretical perspective, evidence suggests rapid eye movements in REM sleep are stimulated by ponto-geniculo-occipital (PGO) spikes of electrical potential from within the pons and nearby areas of the midbrain.6 It appears that during REM sleep, the fingers exhibit elevated bursts of activity similar to eye movements, although the activities may not necessarily be equal in number or synchronised. As finger twitches may be stimulated by the same PGO spikes associated with eye movements during REM sleep, further investigation could be undertaken to assess the temporal relationships between finger twitches and eye movements. Further research is also recommended to determine whether the finger twitching effects found in this sample of young, healthy adults will be found in other samples from the general population, and in samples in which abnormalities of REM sleep are a clinical symptom.
A key challenge to the measurement of sleep by finger twitches will be the inter-individual variability found in the present study. Although the effects were large on average across the sample, they varied considerably between participants. A sleep detection algorithm informed by finger twitch measures would need to be personalised, as across the general population there is a wide range of healthy individual sleep architectures.7 Further research assessing relationships between finger twitching and key sleep parameters is therefore recommended to facilitate the development of an algorithm that will cater for the observed levels of inter-individual variability. A second challenge for algorithm developers will be discrimination of N1 and REM sleep, which had similar twitch densities; an algorithm would require sufficient sophistication to monitor surrounding sleep epochs to decide whether a particular epoch should be designated as N1 or REM sleep. The finger twitch data collected in the present study could be used as a training sample to develop algorithm parameters, then applied to a second independently collected validation sample to see how effectively it measures REM sleep.
There have been many attempts to develop inexpensive and portable sleep measurement systems, but only wrist-based actigraphy has achieved large-scale commercialisation. This study demonstrates that a simple finger-mounted device can detect differences in finger twitching between REM and NREM sleep, and it follows that an algorithm informed by finger twitch density has the potential to use the REM-NREM and time-of-night effects to estimate periods of REM sleep. Sleep is complex, and accurate temporal measurements are needed for clinical diagnoses and research purposes. Whether technology that incorporates finger twitch detection can improve sleep diagnostics sufficiently to be of clinical utility remains to be investigated.
Acknowledgments
This study was financially supported by grants from the Australian Research Council and the National Health and Medical Research Council. An abstract of this paper was presented as a poster at Sleep DownUnder 2018 and published as a poster abstract in a special issue of the Journal of Sleep Research: https://onlinelibrary.wiley.com/doi/10.1111/jsr.152_12766. An abstract of this paper was also presented as a poster at Shiftwork 2019 (24th International Symposium on Shiftwork & Working Time) and published as a poster abstract in a supplement of Sleep Science: http://sleepscience.org.br/details/589. An abstract of this paper was also presented as a poster at World Sleep 2019 and published as a poster abstract in a supplement of Sleep Medicine: https://www.sciencedirect.com/science/article/pii/S1389945719312511.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Aserinsky E, Kleitman N. Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science. 1953;118(3062):273–274. doi:10.1126/science.118.3062.273
2. Carskadon MA, Dement WC. Normal human sleep. In: Kryger M, Roth T, Dement WC, eds. Principles and Practice of Sleep Medicine. Elsevier; 2017:15–24.e13.
3. Rasch B, Born J. About sleep’s role in memory. Physiol Rev. 2013;93(2):681–766.
4. Blumberg MS, Coleman CM, Gerth AI, McMurray B. Spatiotemporal structure of REM sleep twitching reveals developmental origins of motor synergies. Curr Biol. 2013;23(21):2100–2109. doi:10.1016/j.cub.2013.08.055
5. Palagini L, Carmassi C, Conversano C, et al. Transdiagnostic factors across fibromyalgia and mental disorders: sleep disturbances may play a key role. A clinical review. Clin Exp Rheumatol. 2016;34(96):S00.
6. Siegel JM. REM sleep. Principles Pract Sleep Med. 2005;4:120–135.
7. Kelly JM, Strecker RE, Bianchi MT. Recent developments in home sleep-monitoring devices. ISRN Neurol. 2012;2012.
8. Marino M, Li Y, Rueschman MN, et al. Measuring sleep: accuracy, sensitivity, and specificity of wrist actigraphy compared to polysomnography. Sleep. 2013;36(11):1747–1755. doi:10.5665/sleep.3142
9. Hedner J, White DP, Malhotra A, et al. Sleep staging based on autonomic signals: a multi-center validation study. J Clin Sleep Med. 2011;7(3):301.
10. Herscovici S, Pe’er A, Papyan S, Lavie P. Detecting REM sleep from the finger: an automatic REM sleep algorithm based on peripheral arterial tone (PAT) and actigraphy. Physiol Meas. 2007;28(2):129–140. doi:10.1088/0967-3334/28/2/002
11. Fitbit. Start sleeping better with fitbit. 2017. Available from: https://www.fitbit.com/sleep-better.
12. Beattie Z, Pantelopoulos A, Ghoreyshi A, Oyang Y, Statan A, Heneghan C Estimation of sleep stages using cardiac and accelerometer data from a wrist-worn device.
13. Rivera‐García AP, Ramírez‐Salado I, Corsi‐Cabrera M, Calvo J. Facial muscle activation during sleep and its relation to the rapid eye movements of REM sleep. J Sleep Res. 2011;20(1pt1):82–91. doi:10.1111/j.1365-2869.2010.00853.x
14. Aserinsky E, Kleitman N. A motility cycle in sleeping infants as manifested by ocular and gross bodily activity. J Appl Physiol. 1955;8(1):11–18. doi:10.1152/jappl.1955.8.1.11
15. De Gennaro L, Ferrara M, Bertini M. Muscle twitch activity during REM sleep: effect of sleep deprivation and relation with rapid eye movement activity. Psychobiology. 2000;28(3):432–436.
16. Dement W. The occurrence of low voltage, fast, electroencephalogram patterns during behavioral sleep in the cat. Electroencephalogr Clin Neurophysiol. 1958;10(2):291–296. doi:10.1016/0013-4694(58)90037-3
17. Gardner R, Grossman WI, Roffwarg HP, Weiner H. The relationship of small limb movements during REM sleep to dreamed limb action. Psychosom Med. 1975;37(2):147–159. doi:10.1097/00006842-197503000-00005
18. Geisler P, Meier-Ewert K, Matsubayshi K. Rapid eye movements, muscle twitches and sawtooth waves in the sleep of narcoleptic patients and controls. Electroencephalogr Clin Neurophysiol. 1987;67(6):499–507. doi:10.1016/0013-4694(87)90051-4
19. Stoyva JM. Finger electromyographic activity during sleep: its relation to dreaming in deaf and normal subjects. J Abnorm Psychol. 1965;70(5):343–349. doi:10.1037/h0022511
20. Baldridge BJ, Whitman RM, Kramer M. The concurrence of fine muscle activity and rapid eye movements during sleep. Psychosom Med. 1965;27(1):19–26. doi:10.1097/00006842-196501000-00003
21. Berry R, Brooks R, Gamaldo C, et al. The Aasm Manual for the Scoring of Sleep and Associated Events, Version 2. Darien, IL: American Acadamy of Sleep Medicine; 2016.
22. Takahashi K, Atsumi Y. Precise measurement of individual rapid eye movements in REM sleep of humans. Sleep. 1997;20(9):743–752. doi:10.1093/sleep/20.9.743
23. Iber C, Ancoli-Israel S, Chesson A, Quan S. AASM Manual for Scoring Sleep. Am Assoc Sleep Med. 2007.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.