")
Back to Journals » Cancer Management and Research » Volume 12
Expression Profiles of DNA Methylation and Demethylation Machinery Components in Pediatric Myelodysplastic Syndrome: Clinical Implications
Authors Lamim Lovatel V , de Souza Fernandez C, Ferreira Rodrigues E , de Cassia Tavares R , Sobral da Costa E , Abdelhay E , Coelho Soares Lima S , de Souza Fernandez T
Received 11 June 2019
Accepted for publication 18 November 2019
Published 23 January 2020 Volume 2020:12 Pages 543—556
DOI https://doi.org/10.2147/CMAR.S219026
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Antonella D'Anneo
Viviane Lamim Lovatel,1 Cecilia de Souza Fernandez,2 Eliane Ferreira Rodrigues,1 Rita de Cassia Tavares,3 Elaine Sobral da Costa,4 Eliana Abdelhay,5 Sheila Coelho Soares Lima,6 Teresa de Souza Fernandez1
1Cytogenetics Department, Bone Marrow Transplantation Center (CEMO), National Cancer Institute (INCA), Rio de Janeiro, RJ, Brazil; 2Mathematical and Statistical Institute, Federal Fluminense University (UFF), Niterói, RJ, Brazil; 3Outpatient Department, Bone Marrow Transplantation Center (CEMO), National Cancer Institute (INCA), Rio de Janeiro, RJ, Brazil; 4Pediatrics Department, Faculty of Medicine, Federal Rio de Janeiro University (UFRJ), Rio de Janeiro, RJ, Brazil; 5Stem Cell Department, Bone Marrow Transplantation Center (CEMO), National Cancer Institute (INCA), Rio de Janeiro, RJ, Brazil; 6Molecular Carcinogenesis Program, National Cancer Institute (INCA), Rio de Janeiro, RJ, Brazil
Correspondence: Teresa de Souza Fernandez
Instituto Nacional de Câncer (INCA), Centro de Transplante de Medula Óssea, Laboratório de Citogenética, Praça Cruz Vermelha N o 23, 6º Andar. Centro, Rio de Janeiro, RJ CEP: 20230-130, Brasil
Tel +55 21 3207-1701
Email [email protected]
Purpose: The aim of this study was to analyse the expression profiles of DNMT1, DNMT3A, DNMT3B (components of DNA methylation machinery), TET2 and APOBEC3B (components of DNA demethylation machinery) in pediatric MDS patients and investigate their associations with MDS subtypes, cytogenetics, evolution to acute myeloid leukemia (AML) and p15INK4B methylation level.
Patients and Methods: The expressions of DNMT1, DNMT3A, DNMT3B, TET2, and APOBEC3B were evaluated in 39 pediatric MDS patients by real-time quantitative PCR (qPCR). The quantification of p15INK4B methylation levels (MtL) was performed in 20 pediatric MDS patients by pyrosequencing. Mann–Whitney test was used to evaluate possible differences between the expression levels of selected in patients and donors, according to MDS subtypes, karyotypes, evolution to AML and p15INK4B MtL. The correlations between the expression levels of the different genes were assessed by Spearman rank correlation coefficient.
Results: We found that DNMTs expression levels were higher in pediatric MDS compared to donors [DNMT1 (p< 0.03), DNMT3A (p< 0.03), DNMT3B (p< 0.02)]. TET2 and APOBEC3B expression levels did not show a statistically significant difference between pediatric patients and donors. Considering MDS subtypes, patients at initial stage presented DNMT1 overexpression (p< 0.01), while DNMT3A (p< 0.02) and DNMT3B (p< 0.007) were overexpressed in advanced subtypes. TET2 and APOBEC3B expression did not differ in MDS subtypes. DNMT1 (p< 0.03), DNMT3B (p< 0.03), and APOBEC3B (p< 0.04) expression was higher in patients with normal karyotypes, while patients with abnormal karyotypes showed higher DNMT3A expression (p< 0.03). Karyotypes had no association with TET2 expression. DNMTs overexpression was observed in patients who showed disease evolution. A positive correlation was found between DNMTs expression and between APOBEC3B and DNMT3A/DNMT3B. However, TET2 expression was not correlated with DNMTs or APOBEC3B. p15INK4B MtL was higher in pediatric MDS patients compared with donors (p< 0.03) and its hypermethylation was associated with increased DNMT1 expression (p< 0.009).
Conclusion: Our results suggest that the overexpression of DNMTs and an imbalance between the expressions of the DNA methylation/demethylation machinery components play an important role in MDS development and evolution to AML. These results have clinical implications indicating the importance of DNMTs inhibitors for preventing or delaying the progression to leukemia in pediatric MDS patients.
Keywords: pediatric myelodysplastic syndrome, DNMTs, TET2, APOBEC3B
Introduction
Myelodysplastic syndrome (MDS) comprises a heterogeneous group of clonal hematopoietic stem cell diseases. MDS is characterised by ineffective hematopoiesis, presence of dysplasias, peripheral blood cytopenia and an increased risk of evolution to acute myeloid leukemia (AML).1,2 MDS has a higher incidence in adults and elderly individuals. By contrast, MDS is a rare disease in children, occurring in approximately 2–7% of the childhood hematological malignancies.3–6
Pediatric patients with MDS have some distinct genetic, epigenetic and clinical characteristics when compared to adult patients.4–6 Thus, pediatric MDS has a specific classification proposed by Hasle and colleagues in 20035 and revised in 2016.6 These patients can be classified as refractory cytopenia of childhood (RCC), refractory anemia with excess of blasts (RAEB) and refractory anemia with excess of blasts in transformation (RAEB-t).5,6 Recently, Locatelli and Strahm (2018) used for the advanced pediatric MDS the WHO classification of myeloid neoplasms. So, MDS with ≥2% of blasts in peripheral blood (PB) or 5-19% of blasts in bone marrow (BM) is classified as myelodysplastic syndrome with excess of blasts (MDS-EB).7 However, in pediatric MDS classification, the subtype MDS-EB in transformation (MDS-EB-t) is maintained. This subtype is characterized by 20-29% of blasts in the BM.6,7
Due to the heterogeneity and rarity of pediatric MDS, molecular alterations involved in initial and subsequent stages during its development and progression to AML are not well defined. Several studies in adult patients indicate the important role of epigenetic changes in the pathogenesis of MDS.6–9 This epigenetic nature can be explained as follows: (1) among the most frequently mutated genes in MDS, approximately 45% are responsible for the regulation of epigenetic mechanisms and (2) MDS is considered the most responsive disease to treatment with DNA methyltransferase inhibitors.9,10 However, there are few studies focused on the genes that act on the epigenetic mechanisms in pediatric patients with MDS, and these studies have shown that mutations in these genes are rare.11 Nevertheless, epigenetic mediators could be dysregulated by other mechanisms, such as transcriptional alterations and primarily involve components of the DNA methylation and demethylation machinery to result in aberrant methylation profiles.12
Aberrant promoter hypermethylation involving genes encoding cell adhesion molecules, cell cycle regulators and tumor suppressor genes have been associated with hematopoiesis dysregulation. In MDS, a high frequency of hypermethylation of p15INK4B, cadherin 1 (CDH1), death-associated protein kinase (DAPK) and suppressor of cytokine signalling (SOCS-1) has been observed.13 In pediatric MDS, p15INK4B methylation has been associated with disease pathogenesis and poor prognosis.14
DNA methylation is considered a guardian of hematopoietic stem cell fate because it acts to maintain the balance of these cells, their self-renewal capacity, and differentiation in specific hematopoietic cell populations.12 DNA methyltransferases (DNMTs) are enzymes responsible for catalysing the insertion of a methyl group on carbon 5 of a cytosine in the CpG context. DNMT1 is associated with the maintenance of DNA methylation patterns, while DNMT3A and DNMT3B mediate de novo methylation.15 By contrast, DNA demethylation can occur passively during replication through the inhibition of the methylation maintenance process or actively and independent of DNA replication.16 Active demethylation is initiated by two independent pathways. The first involves the progressive oxidation of 5-methylcytosine (5mC) and is catalysed by the ten-eleven-translocation (TET) family of enzymes; the second is driven by the apolipoprotein B mRNA editing enzyme (APOBECs) family, which deaminates 5mC and 5-hydroxylmethyl cytosine (5hmC).17–19 In both cases, mispairing takes place and the base excision repair machinery replaces the modified base by an unmethylated cytosine.18
The balance between the enzymes that act on DNA methylation and demethylation is essential for the maintenance of genomic stability and is referred to as the DNA methylation and demethylation machinery.18 It has been suggested that increased expression of de novo or maintenance DNMTs contributes to the development of leukemia by inducing aberrant hypermethylation of important regions in the genome.20 Up to date, to the best of our knowledge, there are no studies focusing on DNMTs expression in pediatric MDS, and only one study evaluated TET2 expression in pediatric patients.21 Even in adult patients, few studies have been performed to analyse the expression of DNA methylation and demethylation machinery components.17,22 Although the APOBEC family is an important component in the demethylation machinery18,19 and APOBEC3B has been described as a driving mutagenic agent during cancer development and progression,23 no studies involving the APOBEC family have been performed in MDS. Thus, the aim of this study was to analyse the expression of DNMT1, DNMT3A, DNMT3B (components of the DNA methylation machinery), TET2 and APOBEC3B (components of the DNA demethylation machinery) in pediatric patients with MDS and investigate their associations with MDS subtypes, cytogenetics, evolution to AML and the p15INK4B methylation levels of gene to verify the role of epigenetic alterations during pediatric MDS pathogenesis.
Materials and Methods
Patients and Controls
Bone marrow (BM) cells were obtained from 39 pediatric patients with primary MDS between 2007 and 2017. These patients included 23 boys (59%) and 16 girls (41%). The mean age of the patients was 7 years (ranging from 1 to 18 years). Patients were diagnosed at the National Cancer Institute (INCA) and Martagão Gesteira Institute of Pediatrics (IPPMG, UFRJ). The diagnosis and classification were made according to the criteria proposed by WHO,1 but the subtype MDS-EB-t was retained in pediatric classification of MDS.6,7 Twenty-seven patients (69%) were classified as RCC, seven (18%) as MDS-EB and five (13%) as MDS-EB-t (Table 1). None of these patients had been previously treated for malignancy. Bone marrow cells were also obtained from 13 healthy pediatric bone marrow transplantation donors as controls, including eight boys (61.5%) and five girls (38.5%). The mean age of the healthy pediatric donors was 10 years (ranging from 4 to 18 years). The bone marrow samples were collected from the Bone Marrow Transplantation Center (CEMO) at the National Cancer Institute (INCA), Rio de Janeiro, Brazil and Pediatric and Puericulture Martagão Gesteira Institute (IPPMG), Federal University of Rio de Janeiro (UFRJ), Rio de Janeiro, Brazil. This study was reviewed and approved by Ethics Committees of the National Cancer Institute and IPPMG-UFRJ, and was conducted in accordance with the Declaration of Helsinki.
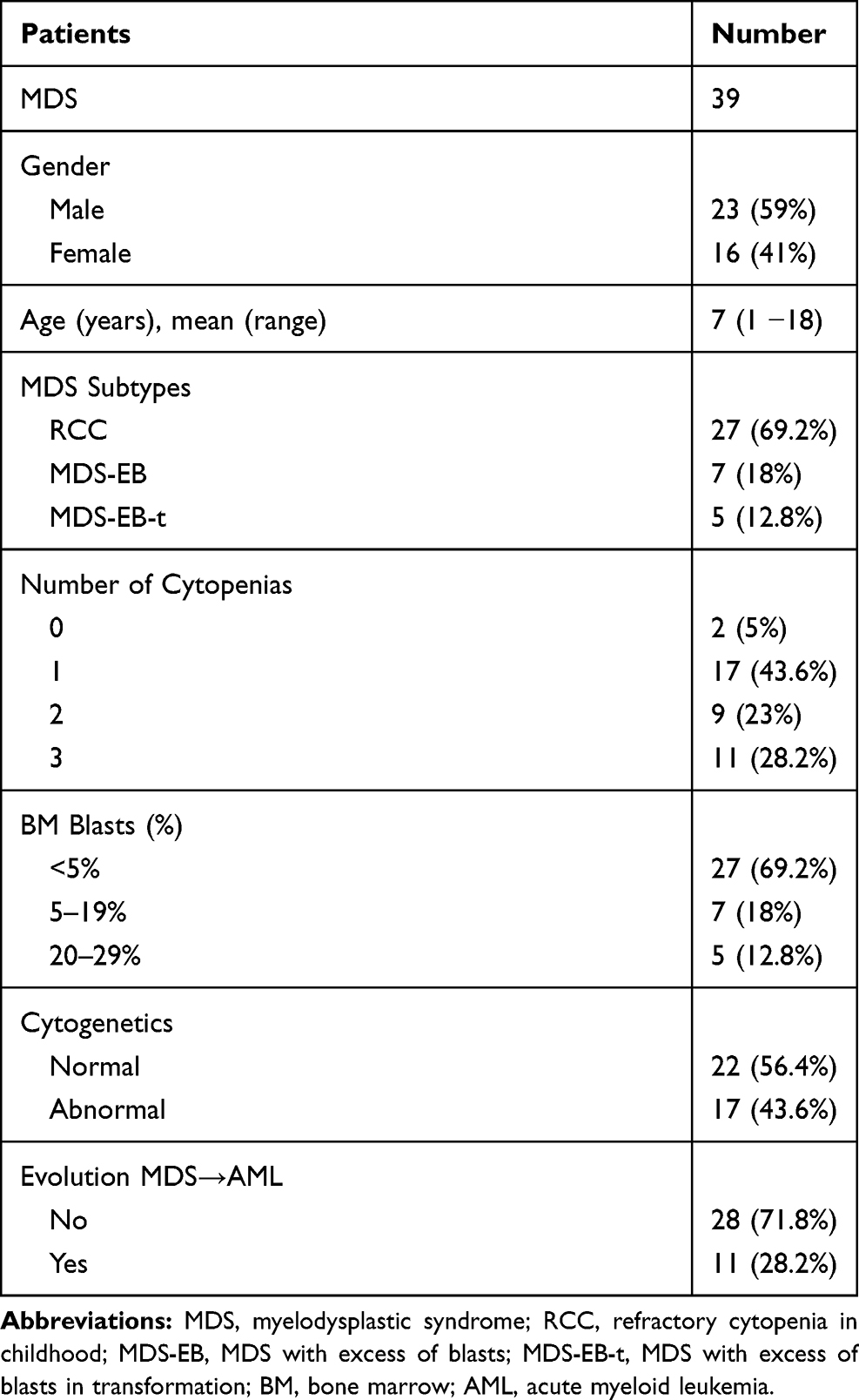 |
Table 1 Patients Characteristics |
Analysis of DNMTs, TET2, and APOBEC3B Expression in Pediatric MDS
Total RNA was extracted from bone marrow cells with TRIzol reagent (Life Technologies, USA) according to the manufacturer’s protocol and stored at −70°C. Two micrograms of total RNA were digested with DNAse amplification grade I (Life Technologies, USA) to remove any genomic DNA contaminant. The RNA was reverse transcribed into complementary DNA (cDNA) with a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, USA) according to the manufacturer’s instructions. Quantitative PCR reactions were performed in 10 µL containing 1x SsoFast EvaGreen Supermix (BIO-RAD, USA), 1.8 µM of each primer and 1 µL of cDNA (diluted 10-fold). Reactions were performed in a Rotor-Gene 6000 thermocycler (Qiagen, Germany) with an initial denaturation at 95°C for 30 s, followed by 40 cycles of 5 s at 95°C and 5 s at 60°C. Β-actin mRNA levels were used as a reference for normalisation. The following primers sequences were used for quantitative PCR: (1) DNMT1, 5ʹ-AAC TCC AAG ACC CAC CCT CC-3ʹ (forward) and 5ʹ-CAG ACT CGT TGG CAT CAA AGA T-3ʹ (reverse); (2) DNMT3A, 5ʹ-GGC TCT TCT TTG AGT TTG ACT TCT ACC-3ʹ (forward) and 5ʹ-GAT GTC CCT GTC ACT AAC-3ʹ (reverse); (3) DNMT3B, 5ʹ-ATC AGG ATG GGA AGG AGT TT-3ʹ (forward) and 5ʹ-TCG GAG AAC TTG CCA TCG CC-3ʹ (reverse); (4) TET2, 5ʹ-GCT GGG ACT ACT GCT GCT GCA CT-3ʹ (forward) and 5ʹ-ACG CAA GCC AGG CTA AAC A-3ʹ (reverse); (5) APOBEC3B, 5ʹ-ACC CAT CCT CTA TGG TCG GA-3ʹ (forward) and 5ʹ-GCT TGA AAT ACA CCT GGC CTC-3ʹ (reverse) and (6) Β-actin, 5ʹ-TGG AAC GGT GAA GGT GAC A-3ʹ (forward) and 5ʹ-ATG TGC AAT CAA AGT CCT CGG C-3ʹ (reverse). The relative expression of all genes was calculated using the ∆∆CT method with the median levels of healthy controls as a normaliser.24
Analysis of p15INK4B Promoter Methylation Levels in Pediatric MDS
The quantification of p15INK4B promoter methylation levels (MtL) was performed in BM cells DNA samples from 20 pediatric patients with MDS and four healthy pediatric donors according to Kim and colleagues in 2013.14 Bisulfite-converted DNA was sequenced using a pyrosequencing system (PSQTM 96MA, Qiagen, Germany). PCR reactions were performed with 0.2 mM/μL of each primer (forward and reverse), 1X PCR Buffer, 0.2 mM/μL of dNTPs, 1.5 mM/μL of MgCl2, 1 u/μL Platinum TaqTM (Invitrogen, CA, USA), and 100 ng of bisulfite-modified DNA in a final volume of 50 μL. PCR cycling conditions consisted of an initial denaturation at 95ºC for 10 mins, followed by 50 cycles of denaturation at 95ºC for 20 s, annealing at 57ºC for 20 s and elongation at 72ºC for 20 s, and a final elongation for 5 mins. PCR quality was confirmed on 2% agarose gels with ethidium bromide staining. Hot-start PCR was performed with HotStarTaq Master Mix kit (Qiagen 203445) and pyrosequencing was performed in accordance with the manufacturer’s protocol (Qiagen). The target CpGs were evaluated by converting the resulting pyrograms into numerical values for peak heights and calculating the average of all CpG sites analyzed at p15INK4B promoter. We used the mean and standard deviation (SD) [mean (0.84) + 2 SD (1.17) = cut-off (3.18)] of the donors for determining hypermethylated samples.14
Statistical Analyses
Comparisons of the DNMTs, TET2, and APOBEC3B expression between patients and donors, MDS subtypes, karyotypes and evolution from MDS to AML were performed using the Mann–Whitney U-test. Correlations between the expression levels of each pair of genes considered in this study were evaluated by the Spearman rank correlation coefficient. The analyses of p15INK4B MtL and its association with the expression of DNA methylation and demethylation machinery components were performed using the Mann–Whitney U-test. Statistical analyses were performed using GraphPad Prism (Graphpad Software, USA). A p-value < 0.05 was considered statistically significant in all analyses.
Results
Analysis of DNMTs, TET2, and APOBEC3B Expression in Pediatric Patients with MDS
The analysis of DNMTs expression levels in pediatric patients with MDS showed a higher expression compared with the donors [DNMT1 (p<0.03), DNMT3A (p<0.03), and DNMT3B (p<0.02)] (Figure 1A–C). Among all DNMTs, DNMT3B showed the highest expression level compared with DNMT1 and DNMT3A. TET2 and APOBEC3B expression levels did not show a statistically significant difference between pediatric patients and the donors (Figure 1D and E).
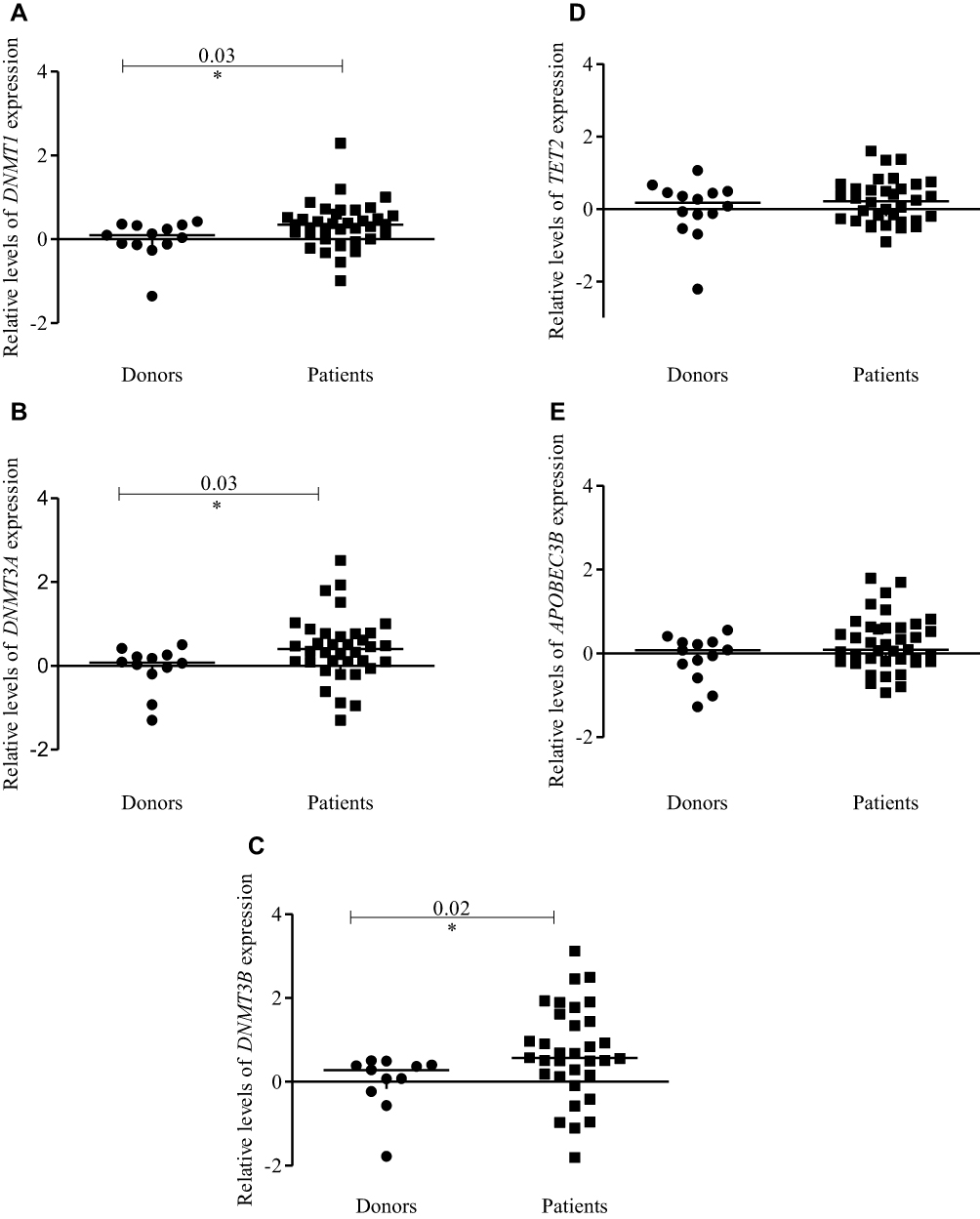 |
Figure 1 Expression profiles of DNMTs, TET2, and APOBEC3B in pediatric patients with MDS and healthy pediatric donors. Dot-plots (log10 scale on the Y-axis) show the expressions for (A) DNMT1, (B) DNMT3A, (C) DNMT3B, (D) TET2 and (E) APOBEC3B. Notes: *Indicates a significant difference (p<0.05) calculated by Mann–Whitney U-test. Abbreviations: DNMTs, DNA methyltransferases; TET, ten-eleven-translocation; APOBEC, apolipoprotein B mRNA editing enzyme; MDS, myelodysplastic syndrome. |
We observed a linear correlation between the expression levels of DNMT1 and DNMT3A (r= 0.52; p<0.001); DNMT1 and DNMT3B (r= 0.42; p<0.01); DNMT3A and DNMT3B (r= 0.68; p<0.0001); DNMT3A and APOBEC3B (r=0.52; p<0.001); and DNMT3B and APOBEC3B (r=0.71; p<0.0001) (Figure 2A, B, E, G and I, respectively). The expression levels of DNMT1 and APOBEC3B were not significantly correlated (r=0.22; p=0.18). The same profile was observed for TET2 expression and DNMTs or APOBEC3B expression (Figure 2C, D, F, H and J, respectively).
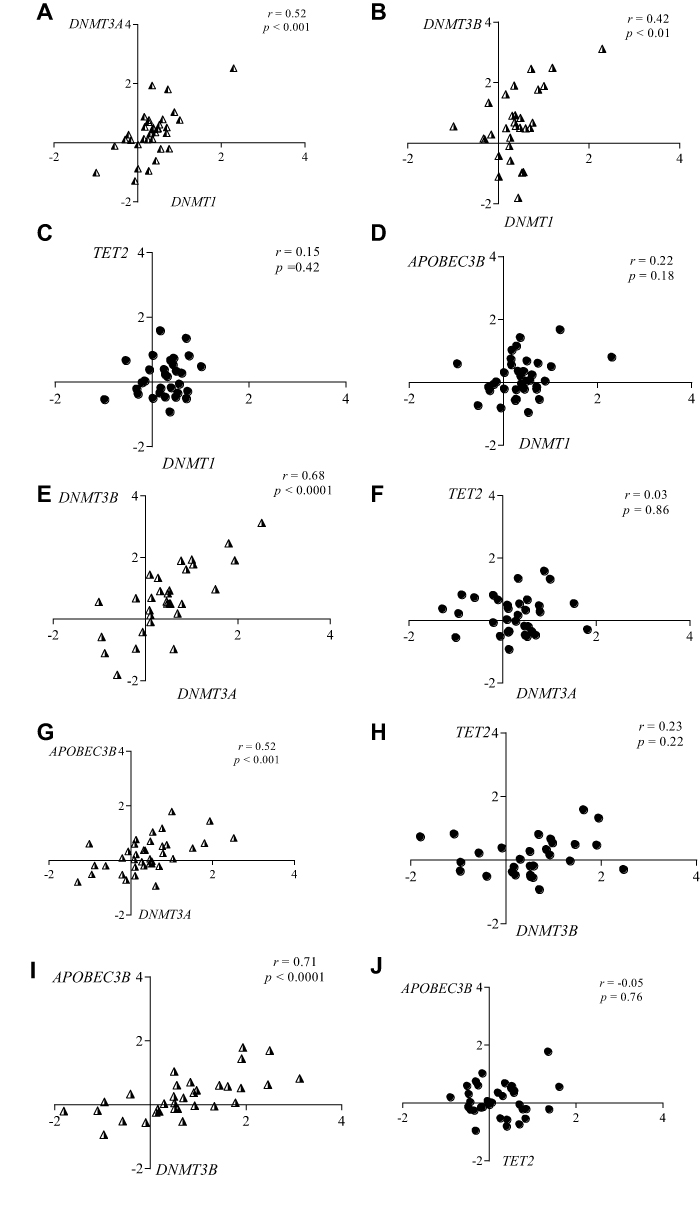 |
Figure 2 Linear correlation between the expression levels of DNMTs, TET2 and APOBEC3B in pediatric patients with MDS. Correlation plots (log10 scale on the X and Y-axis) between the expressions of (A) DNMT1 and DNMT3A; (B) DNMT1 and DNMT3B; (C) DNMT1 and TET2; (D) DNMT1 and APOBEC3B; (E) DNMT3A and DNMT3B; (F) DNMT3A and TET2; (G) DNMT3A and APOBEC3B; (H) DNMT3B and TET2; (I) DNMT3B and APOBEC3B and (J) TET2 and APOBEC3B. Notes: p-Value calculated by Spearman rank correlation coefficient (r). Abbreviations: DNMTs, DNA methyltransferases; TET, ten-eleven-translocation; APOBEC, apolipoprotein B mRNA editing enzyme; MDS, myelodysplastic syndrome. |
Associations of DNMTs, TET2, and APOBEC3B Genes Expression with the MDS Subtypes, Cytogenetics and Disease Evolution
When considering MDS subtypes, we observed a higher DNMT1 expression (p<0.01) at initial stage (RCC) (Figure 3A) and a higher expression of DNMT3A (p< 0.02) (Figure 3B) and DNMT3B (p<0.007) (Figure 3C) in advanced stages (MDS-EB/MDS-EB-t) compared to controls. TET2 and APOBEC3B expression did not significantly differ between MDS subtypes and controls (Figure 3D and E).
 |
Figure 3 Expression profiles of DNMTs, TET2, and APOBEC3B according to the pediatric MDS subtypes and healthy pediatric donors. Dot-plots (log10 scale on the Y-axis) show the expression of (A) DNMT1, (B) DNMT3A, (C) DNMT3B, (D) TET2 and (E) APOBEC3B in donors, RCC (initial MDS stage), MDS-EB/MDS-EB-t (advanced MDS stages). Notes: *Indicates a significant difference (p<0.05) and **Indicates a significant difference (p<0.01) calculated by Mann–Whitney U-test. Abbreviations: DNMTs, DNA methyltransferases; TET, ten-eleven-translocation; APOBEC, apolipoprotein B mRNA editing enzyme; MDS, myelodysplastic syndrome; RCC, refractory cytopenia of childhood; MDS-EB, MDS with excess of blasts; MDS-EB-t, MDS with excess of blasts in transformation. |
In relation of cytogenetics (Table S1), we found that DNMT1 (p<0.03), DNMT3B (p<0.03) and APOBEC3B (p<0.04) expression levels were higher in patients with normal karyotypes (Figure 4A, C and E), while patients with abnormal karyotypes showed a higher DNMT3A expression (p < 0.03) relative to donors (Figure 4B). Karyotypes had no association with TET2 expression (Figure 4D).
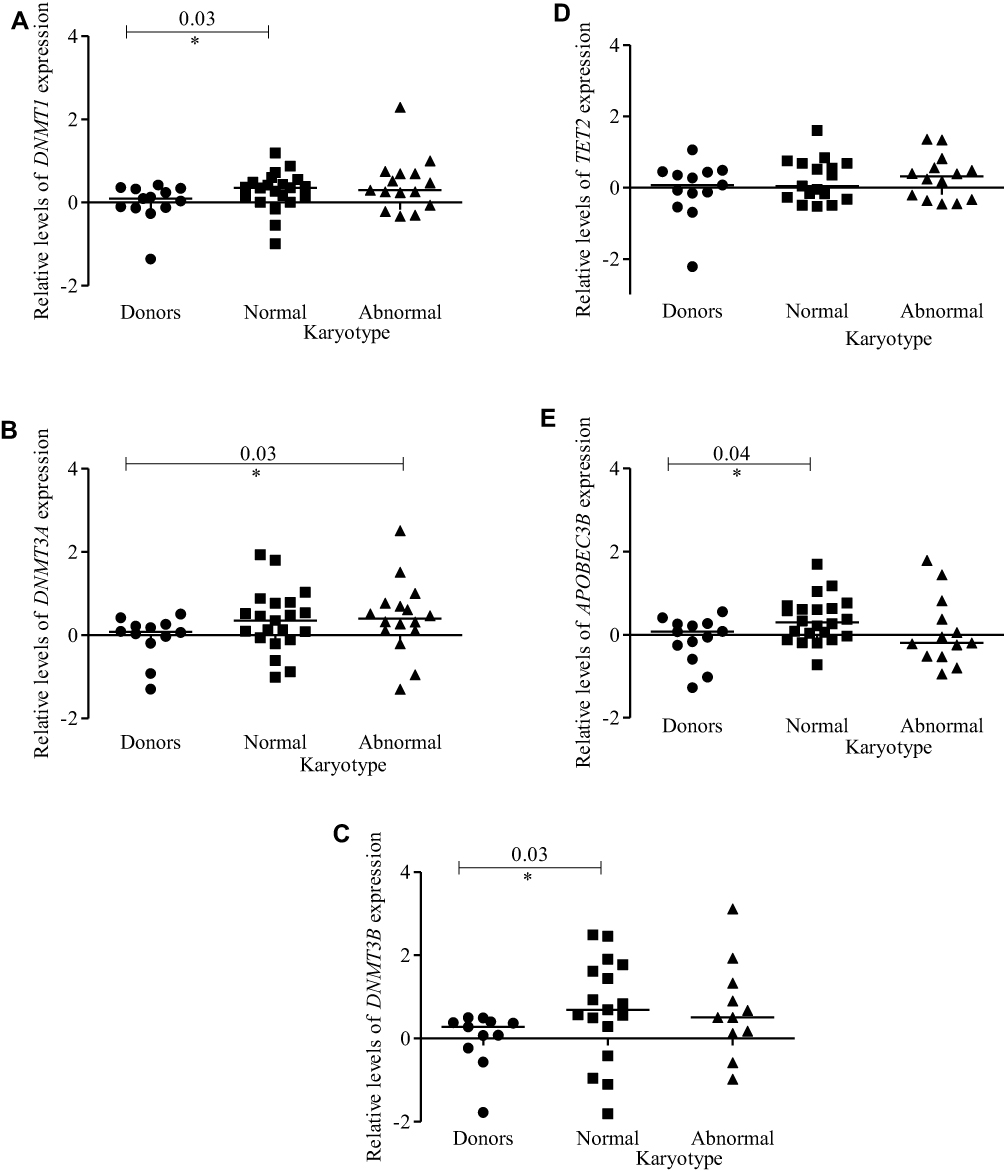 |
Figure 4 Expression profiles of DNMTs, TET2, and APOBEC3B according to cytogenetics (normal and abnormal karyotypes). Dot-plots (log10 scale on the Y-axis) show the expressions of (A) DNMT1, (B) DNMT3A, (C) DNMT3B, (D) TET2 and (E) APOBEC3B in donors and patients. Notes: *Indicates a significant difference (p< 0.05) calculated by Mann–Whitney U-test. Abbreviations: DNMTs, DNA methyltransferases; TET, ten-eleven-translocation; APOBEC, apolipoprotein B mRNA editing enzyme; MDS, myelodysplastic syndrome. |
We also analysed the association between DNMTs, TET2 and APOBEC3B expression and the evolution from MDS to AML. The evolution of disease was observed in 28% of the patients (11/39). Patients who progressed to AML had higher expression levels of DNMT1, DNMT3A, and DNMT3B when compared to donors (p<0.04, p<0.02 and p<0.005, respectively) (Figure 5A–C). TET2 and APOBEC3B expression did not differ between patients who showed evolution from MDS to AML and controls (Figure 5D and E). Table 2 contains a summary of all analyses performed comparing DNMTs, TET2 and APOBEC3B expression between patients and donors, and according to MDS subtypes, karyotypes and evolution from MDS to AML.
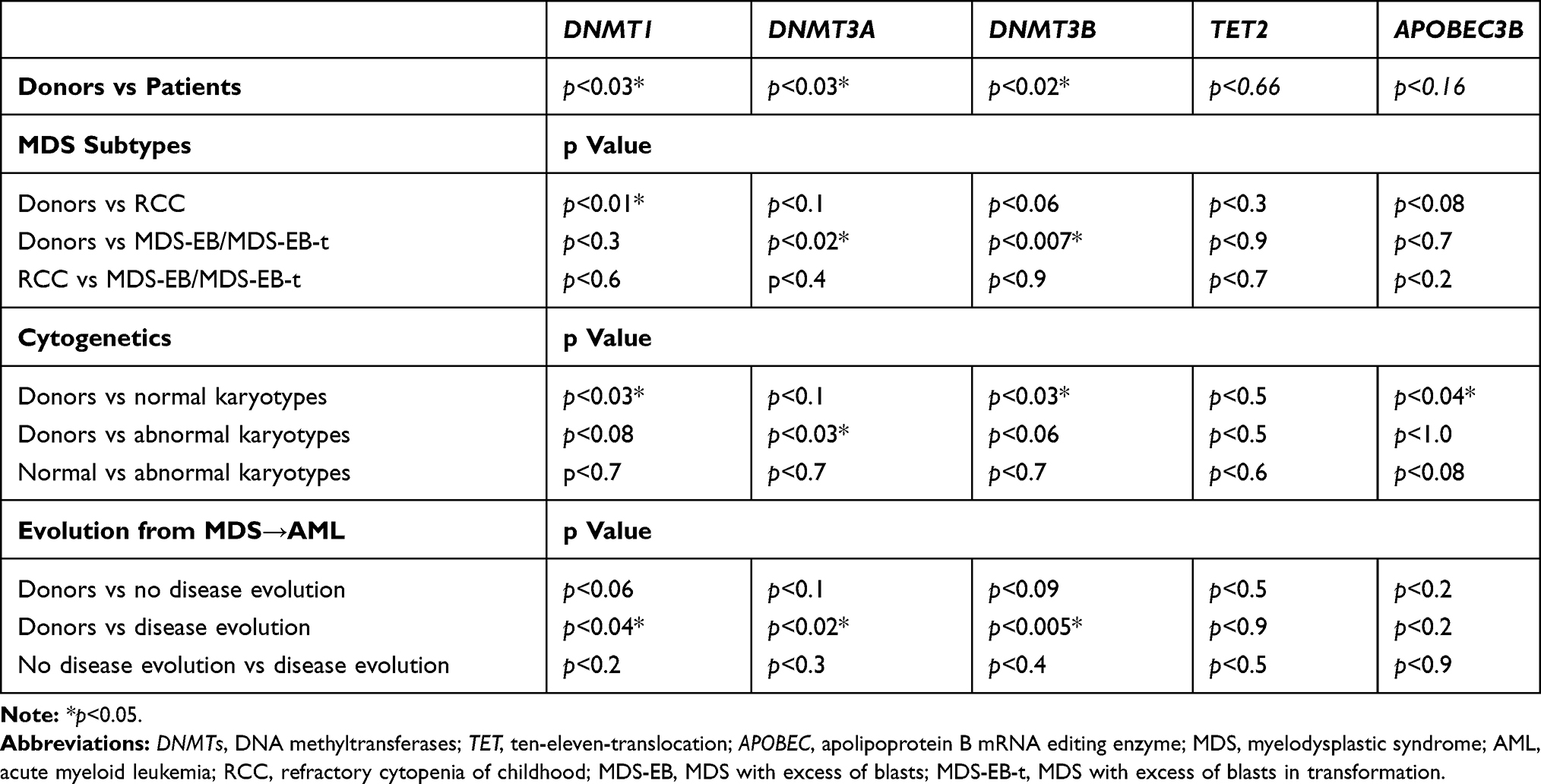 |
Table 2 Summary of All Analyses Performed Comparing DNMTs, TET2 and APOBEC3B Expressions Between Patients and Donors, and According to MDS Subtypes, Karyotypes, Evolution from MDS to AML |
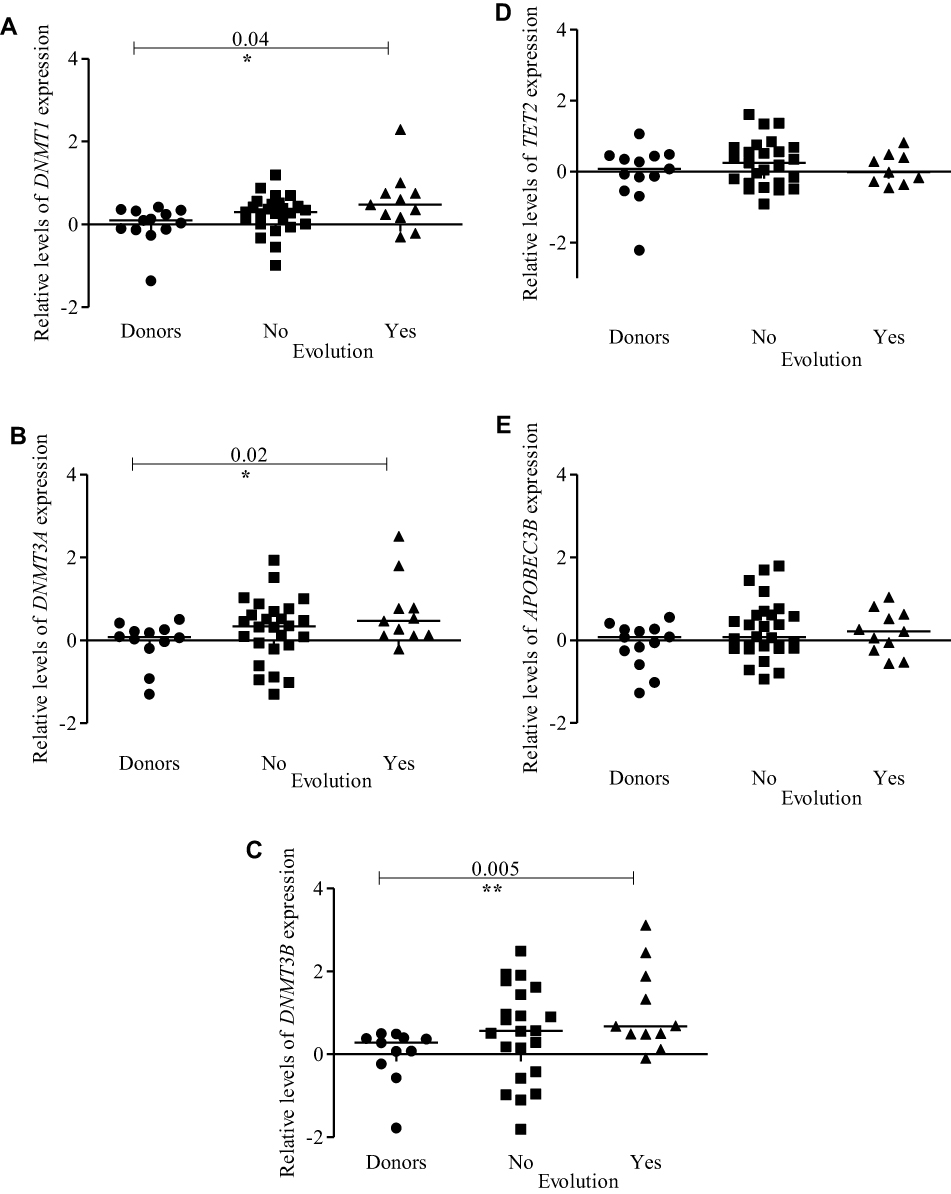 |
Figure 5 Expression profiles of DNMTs, TET2, and APOBEC3B according to evolution from MDS to AML. Dot-plots (log10 scale on the Y-axis) show the expressions of (A) DNMT1, (B) DNMT3A, (C) DNMT3B, (D) TET2, and (E) APOBEC3B in healthy donors, patients who did not show disease evolution and patients who showed disease evolution. Notes: *Indicates a significant difference (p<0.05); **Indicates a significant difference (p<0.01) calculated by Mann–Whitney U-test. Abbreviations: DNMTs, DNA methyltransferases; TET, ten-eleven-translocation; APOBEC, apolipoprotein B mRNA editing enzyme; MDS, myelodysplastic syndrome; AML, acute myeloid leukemia. |
Analysis of p15INK4B Promoter Methylation and Its Association with the Expression of DNA Methylation/Demethylation Machinery Components
The analysis of p15INK4B promoter MtL in 20 patients showed that three had no methylation (15%), 11 had methylation (55%) and six were hypermethylated (30%). When comparing the patients who presented methylation with the donors, we observed a statistically significant increase in the first group (p<0.03) (Figure 6A) and when subdividing MDS cases in methylated and hypermethylated, increased MtL were observed in the latter group (Figure 6B). Next, we compared DNMTs expression in the different groups defined by p15INK4B methylation status and showed increased DNMT1 expression in patients with methylation (p<0.02) and hypermethylation (p<0.009) when compared to donors (Figure 6C). For the other genes (DNMT3A, DNMT3B, TET2, and APOBEC3B) no statistically significant differences were observed (data not shown).
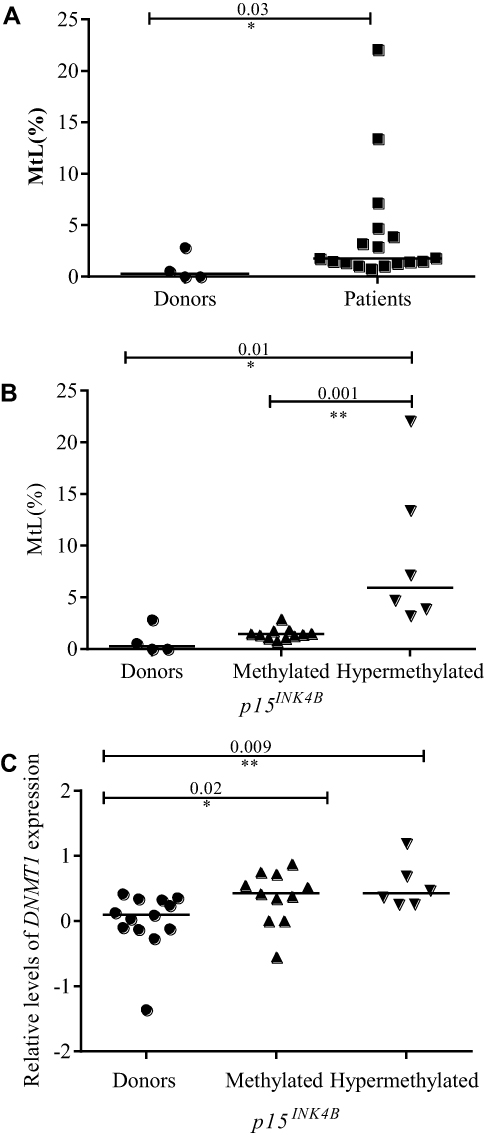 |
Figure 6 p15INK4B promoter methylation profile in pediatric MDS and its association with DNMT1 expression. (A) The comparison of p15INK4B methylation profile between patients (who showed any detectable methylation) and donors, (B) Methylation level of p15INK4B in pediatric MDS according to the methylation cut-off established by the standard deviation (SD) [mean (0.84) + 2 SD (1.17) = cut-off (3.18)] of the donors and (C) Dot-plots (log10 scale on the Y-axis) show the association between p15INK4B gene methylation profile and DNMT1 expression. Notes: *Indicates a significant difference (p<0.05); **Indicates a significant difference (p<0.01) calculated by Mann–Whitney U-test. Abbreviations: DNMT1, DNA methyltransferase 1; p15INK4B, Cyclin-dependent kinase 4 inhibitor B; MtL, quantification of methylation level. |
Discussion
In the present study, we described the expression levels of the DNA methylation and demethylation machinery components in pediatric patients with MDS. DNMT1, DNMT3A, and DNMT3B were overexpressed in pediatric MDS when compared with healthy pediatric donors. The initial stage of MDS showed a higher expression of DNMT1, while in advanced stages an increased expression was observed for DNMT3A and DNMT3B. DNMTs overexpression in MDS was previously demonstrated in adult patients.25,26 Langer et al observed DNMT1, DNMT3A and DNMT3B overexpression in initial stage and in advanced stage when compared to control group.25 Hopfer et al showed a higher expression of DNMT3A and DNMT3B in advanced stages relative to initial stage.26
DNMTs activity is essential for normal hematopoiesis as well as in the process of blood cell differentiation.20,26 DNMT1 has been shown to be important for HSC self-renewal, niche retention, and multilineage hematopoietic differentiation.27 DNMT1 overexpression has already been associated with aberrant methylation in tumour suppressor genes.12 DNMT3A controls the balance between self-renewal and differentiation in normal hematopoiesis28 and alterations in this enzyme result in HSC expansion, impaired differentiation and seems to be associated with more advanced MDS subtypes.26,28 DNMT3A and DNMT3B together play a role in the epigenetic repression of stem cell gene networks during HSC differentiation.29 Recently, it has been demonstrated that DNMT3B acts as a master regulator of transcription in the pediatric AML genome.30
As DNMT1 acts on maintenance methylation while DNMT3A and DNMT3B mediate de novo methylation,15 our results suggest that the three enzymes act in the development of the disease and mainly in its evolution to AML. However, it is noteworthy that the increased expression of DNMT3A and DNMT3B suggesting that new patterns of methylation occur, especially in patients with disease evolution. In fact, some studies have identified aberrant methylomes in adult patients with MDS, as well as hypermethylation of specific genes, particularly during the course of AML.10,12,17,26,31 In our results, we observed an association between p15INK4B promoter methylation and higher DNMT1 expression levels. In this way, it is possible that DNMT3A and DNMT3B produce new methylation patterns during the development of the MDS and that DNMT1 acts by maintaining these patterns.
Some studies demonstrated that there is a coordinate action of the DNMTs.32,33 In accordance, a significant correlation between the expression of these enzymes was observed in the present study, suggesting that there is a cooperative action of the DNMTs expression in pediatric MDS pathogenesis.
TET2 and APOBEC3B act as components of the DNA demethylation machinery and they have relevant roles in cells homeostasis. TET2 has been shown to regulate myeloid differentiation and clonal cell expansion during hematopoiesis. Its decreased expression has been previously reported in patients with MDS/AML, and it has also been associated with a poor prognosis.34,35 APOBEC3B, in addition to its potential role in DNA demethylation, has also been shown to be an important source of mutations in human cancer.23 Through its deamination activity, APOBEC3B can induce C to T and C to G transitions, leaving specific footprints in tumour DNA. Although this APOBEC-mediated mutational signature has been reported in several solid tumours, it is still poorly explored in the context of hematological malignancies.19,36 In adult MDS patients, C to T transitions are the most frequent point mutations.11,31,37 Furthermore, a more recent study with MDS pediatric patients showed that its mutational load is also characterised by C to T substitutions.19
In our study, TET2 and APOBEC3B expression levels did not differ between pediatric patients and donors nor according to MDS subtypes. On the other hand, APOBEC3B expression levels were higher in patients with normal karyotypes when compared with patients with abnormal karyotypes and with healthy donors. In pediatric MDS, most patients with RCC show normal karyotypes.38 However, some of these patients will manifest rapid disease evolution, suggesting the contribution of molecular alterations.9 Our results suggest a possible role of APOBEC3B at initial stage of disease.
We also analysed the correlation between the expression of DNMTs, TET2, and APOBEC3B in pediatric patients with MDS, showing that DNMTs and TET2 expression are not significantly correlated. However, it was possible to observe a correlation between DNMTs and APOBEC3B expression. Alterations in DNMTs also cause DNA imbalances and/or histone modifications, which may lead to chromatin remodelling, genomic instability, and gene expression pattern changes.39 So, our results reinforce the relevant role of DNMTs and also suggests the participation of APOBEC3B, possibly contributing to genomic instability in the pathogenesis of pediatric MDS.
Taken together, our results suggest the presence of an imbalance between the DNA methylation and demethylation machinery components during pediatric MDS pathogenesis, with an overexpression of DNMT1, DNMT3A, and DNMT3B (Figure 7). Our results have clinical implications and indicate the importance of hypomethylating agents for preventing or delaying the progression to leukemia, especially for children undergoing hematopoietic stem cell transplantation (HSCT). Waespe et al demonstrated that azacytidine treatment prior to hematopoietic stem cell transplantation was well tolerated in pediatric patients with advanced MDS and was associated with superior event-free survival.40 Therefore, laboratory tests for detecting the level of DNMTs expression may aid the prediction of disease evolution and may select the patients with MDS for the use of DNA methyltransferases inhibitors.
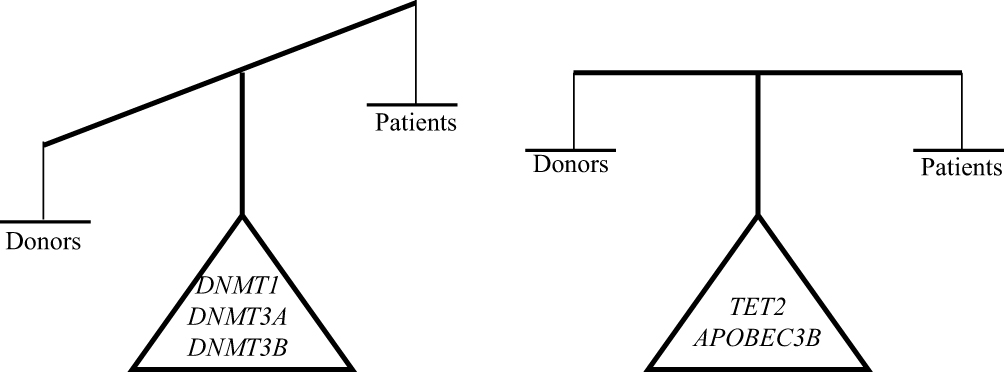 |
Figure 7 Imbalance of the DNA methylation and demethylation machinery components expression in pediatric MDS. Abbreviations: DNMTs, DNA methyltransferases; TET, ten-eleven-translocation; APOBEC, apolipoprotein B mRNA editing enzyme; MDS, myelodysplastic syndrome. |
Since this study was the first to address the alterations in DNA methylation and demethylation machinery components in pediatric patients with MDS, it is necessary to confirm our results in other cohorts to provide a better understanding of the aberrant expression of these genes in the pathogenesis of pediatric MDS.
Conclusion
Our results suggest that the overexpression of DNMTs and an imbalance between the expressions of the DNA methylation/demethylation machinery components play an important role in MDS development and evolution to AML. These results have clinical implications indicating the importance of DNMTs inhibitors for preventing or delaying the progression to leukemia in pediatric MDS patients.
Abbreviations
APOBEC, apolipoprotein B mRNA editing enzyme; AML, acute myeloid leukemia; BM, bone marrow; cDNA, complementary DNA; DNMTs, DNA methyltransferases; HSCs, hematopoietic stem cells; HSCT, hematopoietic stem cell transplantation; MDS, myelodysplastic syndrome; TET, ten-eleven-translocation; MDS-EB, MDS with excess of blasts; MDS-EB-t, MDS with excess of blasts in transformation; MtL, quantification of methylation level; RCC, refractory cytopenia of childhood; 5mC, 5-methylcytosine; 5hmC, 5-hydroxylmethyl cytosine; p15INK4B, Cyclin-dependent kinase 4 inhibitor B.
Data Sharing Statement
The data used to support the findings of this study are included within the article and the original data used to support the findings of this study are available from the corresponding author upon request.
Acknowledgement
This study was supported by the Brazilian Ministry of Health (National Institute of Cancer/INCA, Brazil) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Ethics Approval and Consent to Participate
This study was approved by local Ethic Committees, Ethic Committee of the Brazilian National Cancer Institute (CEP #62/10) and Pediatric and Puericulture Martagão Gesteira Institute (CEP 08926213.9.0000.5264). Informed consent was obtained from children’s parents.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Arber DA, Orazi A, Hasserjia R, et al. The 2016 revision to the World Health Organization (WHO) classification of myeloid neoplasms and acute leukemia. Blood. 2016;127:2391–2405. doi:10.1182/blood-2016-03-643544
2. Montalban-Bravo G, Garcia-Manero G. Myelodysplastic syndromes: 2018 update on diagnosis, risk-stratification and management. Am J Hematol. 2018;93:129–147. doi:10.1002/ajh.24930
3. Babushok DV, Bessler M, Olson TS. Genetic predisposition to myelodysplastic syndrome and acute myeloid leukemia in children and young adults. Leuk Lymphoma. 2016;57:520–536. doi:10.3109/10428194.2015.1115041
4. Glaubach T, Robinson LJ, Corey SJ. Pediatric myelodysplastic syndromes: they do exist! J Pediatr Hematol Oncol. 2014;36:1–7. doi:10.1097/MPH.0000000000000046
5. Hasle H, Niemeyer CM, Chessells JM, et al. A pediatric approach to the WHO classification of myelodysplastic and myeloproliferative diseases. Leukemia. 2003;17:277–282. doi:10.1038/sj.leu.2402765
6. Hasle H. Myelodysplastic and myeloproliferative disorders of childhood. Hematology Am Soc Hematol Educ Program. 2016;2016:598–604. doi:10.1182/asheducation-2016.1.598
7. Locattelli F, Strahm B. How I treat myelodysplastic syndromes of childhood. Blood. 2018;131:1406–1414. doi:10.1182/blood-2017-09-765214
8. Heuser M, Yun H, Thol F. Epigenetics in myelodysplastic syndromes. Semin Cancer Biol. 2018;51:170–179. doi:10.1016/j.semcancer.2017.07.009
9. Bejar R, Steensma DP. Recent developments in myelodysplastic syndromes. Blood. 2014;124:2793–2803. doi:10.1182/blood-2014-04-522136
10. Issa JP. The myelodysplastic syndrome as a prototypical epigenetic disease. Blood. 2013;121:3811–3817. doi:10.1182/blood-2013-02-451757
11. Schwartz JR, Jing M, Lamprecht T, et al. The genomic landscape of pediatric myelodysplastic syndromes. Nat Commun. 2017;8:1557. doi:10.1038/s41467-017-01590-5
12. Bröske AM, Vockentanz L, Kharazi S, et al. DNA methylation protects hematopoietic stem cell multipotency from myeloerythroid restriction. Nat Genet. 2009;41:1207–1215. doi:10.1038/ng.463
13. Fernandez TS, Mencalha A, Fernandez CS. Epigenetics in cancer: the myelodysplastic syndrome as a model to study epigenetic alterations as dignostic and prognostic biomarkers. In: Khan TK, editor. Biomarker.
14. Kim M, Kook H, Park HJ, et al. Quantitative comparison of CDKN2B methylation in pediatric and adult myelodysplastic syndromes. Acta Haematol. 2013;130:115–121. doi:10.1159/000347038
15. Li KK, Luo LF, Shen Y, Xu J, Chen Z, Chen SJ. DNA methyltransferases in hematologic malignancies. Semin Hematol. 2013;50:48–60. doi:10.1053/j.seminhematol.2013.01.005
16. Kohli RM, Zhang Y. TET enzymes, TDG and the dynamics of DNA demethylation. Nature. 2013;502:472–479. doi:10.1038/nature12750
17. Kunimoto H, Nakajima H. Epigenetic dysregulation of hematopoietic stem cells and preleukemic state. Int J Hematol. 2017;106:34–44. doi:10.1007/s12185-017-2257-6
18. Branco MR, Ficz G, Reik W. Uncovering the role of hydroxymethylcytosine in the epigenome. Nat Rev Genet. 2011;13:7–13. doi:10.1038/nrg3080
19. Fu Y, Ito F, Zhang G, Fernandez B, Yang H, Chen XS. DNA cytosine and methylcytosine deamination by APOBEC3B: enhancing methylcytosine deamination by engineering APOBEC3B. Biochem J. 2015;471:25–35. doi:10.1042/BJ20150382
20. Mizuno S, Chijiwa T, Okamura T, et al. Expression of DNA methyltransferases DNMT1, 3A, and 3B in normal hematopoiesis and in acute and chronic myelogenous leukemia. Blood. 2001;97:1172–1179. doi:10.1182/blood.V97.5.1172
21. Coutinho DF, Monte-Mór BCR, Vianna DT, et al. TET2 expression level and 5-hydroxymethylcytosine are decreased in refractory cytopenia of childhood. Leuk Res. 2015;39:1103–1108. doi:10.1016/j.leukres.2015.07.005
22. Aoki E, Ohashi H, Uchida T, Murate T, Saito H, Kinoshita T. Expression levels of DNA methyltransferase genes do not correlate with p15INK4B gene methylation in myelodysplastic syndromes. Leukemia. 2003;17:1903–1904. doi:10.1038/sj.leu.2403046
23. Burns MB, Temiz NA, Harris RS. Evidence for APOBEC3B mutagenesis in multiple human cancers. Nat Genet. 2013;45:977–983. doi:10.1038/ng.2701
24. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 22DDCT. Method. 2001;25:402–408. doi:10.1006/meth.2001.1262
25. Langer F, Dingemann J, Kreipe H, Lehmann U. Up-regulation of DNA methyltransferses DNMT1, 3A, and 3 B in myelodysplastic syndrome. Leuk Res. 2005;29:325–329. doi:10.1182/blood.V97.5.1172
26. Hopfer O, Komor M, Koehler IS, et al. Aberrant promotor methylation in MDS hematopoietic cells during in vitro lineage specific differentiation is differently associated with DNMT isoforms. Leuk Res. 2009;33:434–442. doi:10.1016/j.leukres.2008.08.014
27. Trowbridge JJ, Snow JW, Kim J, Orkin SH. DNA methyltransferase 1 is essential for and uniquely regulates hematopoietic stem and progenitor cells. Cell Stem Cell. 2009;5:442–449. doi:10.1016/j.stem.2009.08.016
28. Zhang W, Xu J. DNA methyltransferases and their roles in tumorigenesis. Biomarker Res. 2017;5:1. doi:10.1186/s40364-017-0081-z
29. Challen GA, Sun D, Mayle A, et al. Dnmt3a and Dnmt3b have overlapping and distinct functions in hematopoietic stem cells. Cell Stem Cell. 2014;15:350–364. doi:10.1016/j.stem.2014.06.018
30. Lamba JK, Cao X, Raimondi SC, et al. Integrated epigenetic and genetic analysis identifies markers of prognostic significance in pediatric acute myeloid leukemia. Oncotarget. 2018;9:26711–26723. doi:10.18632/oncotarget.25475
31. Figueroa ME, Skrabanek L, Li Y, et al. MDS and secondary AML display unique patterns and abundance of aberrant DNA methylation. Blood. 2009;114:3448–3458. doi:10.1182/blood-2009-01-200519
32. Yang J, Wei X, Wu Q, et al. Clinical significance of the expression of DNA methyltransferase proteins in gastric cancer. Mol Med Rep. 2011;4:1139–1143. doi:10.3892/mmr.2011.578
33. Subramaniam D, Thombre R, Dhar A, Anant S. DNA methyltransferases: a novel target for prevention and therapy. Front Oncol. 2014;4:1–13. doi:10.3389/fonc.2014.00080
34. Scopim-Ribeiro R, Machado-Neto JA, Campos PDE M, et al. Ten-eleven-translocation 2 (TET2) is downregulated in myelodysplastic syndromes. Eur J Haematol. 2015;94:413–418. doi:10.1111/ejh.12445
35. Zhang W, Shao Z, Fu R, Wang H, Li L, Liu H. Down regulation of TET2 in CD3+ and CD34+ cells of myelodysplastic syndromes and enhances CD34+ cells proliferation. Int J Clin Exp Pathol. 2015;8:10840–10846.
36. Zou J, Wang C, Ma X, Wang E, Peng G. APOBEC3B, a molecular driver of mutagenesis in human cancers. Cell Biosci. 2017;7:29. doi:10.1186/s13578-017-0156-4
37. Sperling AS, Gibson CJ, Ebert BL. The genetics of myelodysplastic syndrome: from clonal haematopoiesis to secondary leukaemia. Nat Rev Cancer. 2016;17:5–19. doi:10.1038/nrc.2016.112
38. Hofmann I. Pediatric myelodysplastic syndromes. J Hematopathol. 2015;8:127–141. doi:10.1097/MPH.0000000000000046
39. Zhang W, Shao ZH, Fu R, et al. TET2 expression in bone marrow mononuclear cells of patients with myelodysplastic syndromes and its clinical significances. Cancer Biol Med. 2012;9:34–37. doi:10.3969/j.issn.2095-3941.2012.01.006
40. Waespe N, Van Den Akker M, Klaassen RJ, et al. Response to treatment with azacitidine in children with advanced myelodysplastic syndrome prior to hematopoietic stem cell transplantation. Haematologica. 2016;101(12):1508–1515. doi:10.3324/haematol.2016.145821
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.