")
Back to Journals » Infection and Drug Resistance » Volume 15
Exploring the Benefits of Metal Ions in Phage Cocktail for the Treatment of Methicillin-Resistant Staphylococcus aureus (MRSA) Infection
Authors Li X, Chen Y, Wang S, Duan X, Zhang F, Guo A, Tao P, Chen H, Li X, Qian P
Received 22 February 2022
Accepted for publication 15 April 2022
Published 27 May 2022 Volume 2022:15 Pages 2689—2702
DOI https://doi.org/10.2147/IDR.S362743
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Xinxin Li,1– 3 Yibao Chen,1– 3 Shuang Wang,1– 3 Xiaochao Duan,1– 3 Fenqiang Zhang,1– 3 Aizhen Guo,1– 3 Pan Tao,1– 3 Huanchun Chen,1– 3 Xiangmin Li,1– 3 Ping Qian1– 3
1State Key Laboratory of Agricultural Microbiology, Huazhong Agricultural University, Wuhan, Hubei, 430070, People’s Republic of China; 2College of Veterinary Medicine, Huazhong Agricultural University, Wuhan, Hubei, 430070, People’s Republic of China; 3Key Laboratory of Preventive Veterinary Medicine in Hubei Province, the Cooperative Innovation Center for Sustainable Pig Production, Wuhan, Hubei, 430070, People’s Republic of China
Correspondence: Ping Qian, Tel +86-27-87282608, Fax +86-27-87282608, Email [email protected]
Background: Methicillin-resistant Staphylococcus aureus (MRSA) is an important zoonotic pathogen worldwide. Infections due to MRSA are associated with higher mortality rates compared with methicillin-susceptible S. aureus. Meanwhile, bacteriophages have been shown to overcome the emergence of MRSA.
Methods: Phage PHB22a, PHB25a, PHB38a, and PHB40a were isolated. Here, we evaluated the ability of a phage cocktail containing phages PHB22a, PHB25a, PHB38a, and PHB40a against MRSA S-18 strain in vivo and in vitro. Phage whole-genome sequencing, host-range determination, lytic activity, and biofilm clearance experiments were performed in vitro. Galleria mellonella larvae and a mouse systemic infection model to evaluate the efficacy of phage therapy in vivo.
Results: The phage cocktail exhibited enhanced antibacterial and anti-biofilm effects compared to the single phage. Phage cocktail contained with Ca2+/Zn2+ significantly reduced the number of viable bacteria (24-h or 48-h biofilm) by more than 0.81-log compared to the phage cocktail alone. Furthermore, we demonstrated that the addition of Ca2+ and Zn2+ phage cocktail could increase the survival rate of G. mellonella larvae infected with S. aureus by 10% compared with phage cocktail alone. This was further confirmed in the mouse model, which showed a 2.64-log reduction of host bacteria S-18, when Ca2+ and Zn2+ were included in the cocktail compared with the phage cocktail alone.
Conclusion: Our results indicated that phage cocktail supplemented with Ca2+/Zn2+ could effectively remove bacteria in biofilms and mice tissues infected with S. aureus.
Keywords: methicillin-resistant Staphylococcus aureus, phage cocktail, metal ions, biofilm, phage therapy
Introduction
The Gram-positive Staphylococcus aureus, an important zoonotic pathogen caused bacteremia, endocarditis, and tissue infection in humans and animals,1–4 are listed as a “high priority” pathogen by the World Health Organization (WHO). It is estimated that in the absence of new antibiotic alternatives, by 2050, antibiotic-resistant pathogens will cause around 10 million deaths worldwide.5 The emergence of methicillin-resistant S. aureus (MRSA) and vancomycin-resistant S. aureus (VRSA) have brought challenges to clinical treatment.6,7
Bacteriophages (phages), first discovered in the early 20th century, are the most abundant organism in the world.8 Phage therapy has the advantages of high specificity to bacteria,9 reducing bacterial tolerance and removing biofilms.10–12 Recent studies have shown that phages could be a way to solve the crisis of extensively drug-resistant bacteria in animals and humans.6,13 For example, S. aureus phage Sb-1, successfully treated five diabetic foot ulcers patients for whom standard antibiotics were ineffective.14 The S. aureus phage cocktail is safe to give intravenously to patients with severe S. aureus infection.13 With the advantage of safety and tolerability in patients,13 combination of multiple phages as phage cocktail have been successfully used to treat wound infections in mice and chronic rhinosinusitis in humans caused by S. aureus.14,15 However, the NCBI database only had information on 69 staphylococcal virulent phage genomes in 2019.16 In contrast, temperate phages are abundant in nature and in bacteria.17 Meanwhile, the temperate phage JD419 of S. aureus exhibits a broad host range.18 There has been temperate phage ØCD27 for the treatment of Clostridium difficile infection.19 In addition, temperate phages MP22 and D3112 from clinical isolates of Pseudomonas aeruginosa for the treatment of peritonitis-sepsis caused by P. aeruginosa.20 Therefore, the temperate phage may be used as an alternative strategy to replace antibiotics and virulent phage in the future.21
The extracellular polysaccharides (EPS), extracellular DNA, and proteins form biofilm matrix protect bacteria from the external environment, which enhances bacteria’s resistance to antibiotics.22 Many antibiotics were less effective against S. aureus when it forms a biofilm.23 Phages and their derivatives have able to destroy the biofilm substrate.10,24 For instance, phages and their derivatives can remove the biofilm formed by S. aureus on different surfaces and mammary-gland tissue.10,24 In addition, the temperate phage vB_SauS_JS02 with a higher inhibiting and degrading activity against S. aureus biofilm.25 In previous studies, we found that the metal Ions mixed endolysin Lys08 can enhance its antibacterial activity and the ability to remove biofilms.26 Although studies showed the phage cocktail can remove the biofilm or treat Galleria mellonella larvae and mice with infection S. aureus, it is unknown whether addition of metal ions mixed phage cocktail to remove the biofilm or treat bacterial diseases of S. aureus.
In this study, we explored the effects of metal ions on phage therapy using a phage cocktail containing four phages PHB22a, PHB25a, PHB38a, and PHB40a against biofilms and MRSA S-18 infection in animal models. The addition of Ca2+ and Zn2+ significantly improved anti-biofilm activity of the phage cocktail. Meanwhile, treatment with phage cocktail supplemented with Ca2+ and Zn2+ increased the survival of G. mellonella larvae infected with MRSA S-18 and reduced the bacterial load of tissues in mice models. Our data highlight that phage cocktail with different metal ions is an effective strategy against MRSA infection.
Materials and Methods
Bacterial Strains
All the strains were present in our lab collection are listed in Table S1. ATCC25923 were obtained from American Type Culture Collection (ATCC). All strains were cultured on LB medium (Solarbio) or LB agar plates (1.5% agar) at 37°C for 12 h.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility testing was conducted using the disk diffusion method.27 Thirteen antibiotics disks, including Cefoxitin (30 μg), Penicillin G (10 U), Gentamicin (10 μg), Erythromycin (15 μg), Tetracycline (30 μg), Doxycycline (30 μg), Minocycline (30 μg), Furantoin (300 μg), Clindamycin (2 μg), Trimethoprim (5 μg), Sulfafurazole (300 μg), Chloramphenicol (30 μg), and Rifampicin (5 μg) were tested on Mueller-Hinton agar (Oxoid, UK) containing MRSA strain. After overnight incubation at 37°C, the diameter of the inhibition band (diameter length/mm) was measured. According to the Clinical & Laboratory Standards Institute (2018), the inhibition bands were interpreted as susceptible (S), intermediate (I), and resistant (R).28,29 S. aureus ATCC25923 was used as control strain.
Phage Isolation and Sequencing
Phages were isolated from sewage samples using a double-layer method.30 Firstly, the sewage samples were centrifuged at 12,000 × g for 10 min to remove the sediment. The supernatants were filtered using a 0.22 μm membrane (Millex–GP, USA) to remove bacteria. Then, 100 μL of the filtrates mixed with 200 μL of the host strain in the logarithmic phase were incubated at 37°C for 12 h. A single plaque was picked and resuspended in the host strain in the logarithmic phase. This step was repeated five times. The phage particles were purified by CsCl gradient ultracentrifugation as previously described.31 The transmission electron microscope (H-7650; Hitachi, Tokyo, Japan) was used for phage morphology with an acceleration voltage of 100 kV.27 Phage genomic DNA was extracted using phenol chloroform as previously described.32 Complete genome sequencing of phage PHB22a, PHB25a, PHB38a, and PHB40a was performed using the Illumina MiSeq (San Diego, CA, USA) system. The resulting data were assembled using Newbler v.3.0. Open reading frame (ORF) were predicted using RAST (http://rast.nmpdr.org/). Protein BLAST (BLASTP) (https://www.ncbi.nlm.nih.gov/BLAST/) was used to identify putative homology and proteins with similarity to predicted phage proteins. The short reads were assembled into the genome by means of SOAPdenovo (http://sourceforge.net/projects/soapdenovo2/files/SOAPdenovo2/).33 Phylogenetic analysis of phage large terminase subunit sequences was performed using the ClustalW program in MEGA6.34
Host Range of the Phages
The host range of phages was performed using the double-layer plate methods.32 Briefly, 200 μL of the bacterium in the exponential growth phase mixed with 5 mL of soft LB agar (0.75% agar) was poured on the double-layer plate. A 5 μL of the different titers of phages were spotted on a double-layer plate and then were incubated at 37°C for 12 h. The efficiency of plating (EOP) values was determined by calculating the ratio of PFUs of each phage-susceptible strain to its host strain.
Phages Biological Characterization
The phages stability of temperature and pH was performed as previously described.35 The thermal stability of phages was determined by incubation in phosphate buffer at 4, 20, 30, 40, 50, 60, 70, and 80°C (pH=7.0) for 1 h. To test the acid-base stability of phages, 100 μL phage was incubated with 900 μL different pH buffer (3, 5, 7, 9, and 11) for 1 h at 37°C. MRSA S-18 as the phages common host strain. Phage titers were determined by double-layer plate method. This experiment was repeated three times.
One-Step Growth Assay
One-step growth assay was performed as previously described.36 Briefly, the logarithmic-growth MRSA S-18 strain (20 mL) after addition of phage at the most appropriate multiplicity of infection (MOI) was incubated at 37°C for 5 min, followed by centrifugation at 12,000 × g for 1 min to remove unabsorbed phages. The pellet was resuspended in LB medium (20 mL) and incubated at 37°C at 200 × g. A 0.5 mL sample was collected after a total duration of 120 min. The PFU counts were obtained using the double-layer agar method. This experiment was repeated three times.
Phage Treatment of S. aureus Biofilm in 96-Well Cell Culture Plates
A 96-well plate was used to evaluate the phage’s ability to remove the biofilm as previously described.10 Firstly, overnight bacteria were diluted to a concentration of 1×107 CFU/mL. A 200 μL of diluent S-18 strain was added to each well. After incubation at 30°C for 24 h or 48 h, the biofilm was treated with the phages or phages contained with metal ions at 37°C for 4 h. The crystal violet staining was evaluated to the effect of phages on removal of biofilm. Specifically, each well was washed with sterile PBS, this step was repeated three times. After phage treatment, the biofilm was fixed with 1% crystal violet at room temperature for 30 min, and then was washed with sterile PBS, this step was repeated three times. Finally, each well was treated 200 μL of 33% glacial acetic acid for 30 min. The OD of each well was the measured at 590nm using a spectrophotometer. For detection of bacterial numbers in biofilm, the biofilm matrix each well was resuspended with sterile PBS buffer. The number of live bacteria (CFU/cm2) in the biofilm was calculated by plate counting of 10-fold serial dilution. This experiment was performed in triplicate. The morphology of MRSA S-18 biofilm was observed by scanning electron microscopy (SEM). Biofilm on coverslips were treated with different concentrations of ethanol (50%, 70%, 80%, 90% and 100% × 3 times) for 15 minutes. Then, the coverslips were sprayed with gold and observed under SEM (5000 ×).
Lytic Activity
The lytic activity of phages or phage cocktail was determined in a 96-well microtiter plate by analyzing the optical density (OD600) measurement method.37,38 Briefly, 20 mL host bacteria S-18 in logarithmic-growth was mixed with 200 μL phage of different MOIs (0.0001, 0.001, 0.01, 0.1, 1, and 10). The mixture was cultured at 37°C at 200 × g. Samples were collected from 0 h until 24 h and tested their OD600. This experiment was repeated three times.
Systemic Infection of G. mellonella Larvae
The weighing of 0.2–0.3 g G. mellonella larvae was cultured at 15°C in the dark environment. G. mellonella larvae was injected by using a 29-gauge (G) insulin syringe (U-40) from BD Micro-FineTM (Franklin Lakes, NJ, USA). The host strain S-18 was cultured at 37°C for 12 h, then was centrifuged and resuspended with PBS (pH 7.4) to final a concentration of 2×109 CFU/mL. The minimum lethal dose (MLD100) trial was determined as described previously.39 A 20 μL of S-18 strain at different concentrations (4 × 102 CFU, 4×103 CFU, 4×104 CFU, 4×105 CFU, and 4×106 CFU) were injected into separate groups of larvae (n = 15). A dose of 1 × MLD100 was used for the G. mellonella larvae challenge assays. For the infection assay, 20 μL of S-18 strain (4 × 106 CFU) was injected into the larvae belly (n = 20) on one side of the last proleg and the phages were injected into the larvae belly on the other side of the last proleg. After infection 1h or 0h, the equal volume of phage alone, phage cocktail (4 × 107 PFU), or phage cocktail (4 × 107 PFU) with 0.1mM Ca2+/Zn2+ was injected into the larvae belly. For pretreatment, the larvae (n = 20) were infected by the same dose of phages before infection 1 h. The equal volume PBS was injected as the negative control. The survival of larvae was calculated 48 h after infection. When the larvae did not respond to touch, they were recorded as dead.
Mouse Infection Assays
Five-week-old specific-pathogen-free (SPF) BALB/c female mice were purchased from the Experimental Animal Centre of Huazhong Agricultural University, China. The minimum lethal dose (MLD100) of strain S-18 on mice was determined as described above. A dose of 1 × the MLD100 (7.2 × 108 CFU) was used for the mouse challenge assays. Thirty BALB/c female mice were randomly divided into five groups (n = 6 per group), including phage cocktail treatment alone group, phage cocktail contained 0.1mM Ca2+ group, phage cocktail contained 0.1mM Zn2+ group, S-18 group, and PBS group. Administration before 1 h, the mice were inoculated intraperitoneally treated with 200 μL of phage (1 × 109 PFU) or equal volume PBS buffer. The survival of mice at 7 days was recorded.
For bacterial counts, a parallel test was performed according as described above. Every group included nine BALB/c female mice. The number of viable bacteria in the heart, liver, spleen, lungs, kidney, and blood were determined at 3h, 6h, and 12 h post-infection, respectively. Blood collected from mice at 12 h post-infection was centrifuged at 3000 × g for 15 min to separate the serum. The abundances of tumor necrosis factor alpha (TNF-α), gamma interferon (IFN-γ), interleukin 6 (IL-6), and interleukin 1β (IL-1β) in the serum samples were determined by enzyme-linked immunosorbent assay (ELISA) using commercially available cytokine kits (QuantiCyto® Mouse ELISA kit).
To mimic a clinical treatment situation, we implemented two treatment methods, including one with treatment administered simultaneously with infection and one in which treatment was delayed (1 h post-infection). In brief, a dose of 1×108 CFU was used for the mouse challenge assays. Every group included three BALB/c female mice. The number of viable bacteria in the heart, liver, spleen, lungs, kidney, and blood were determined at 12 h post-infection, respectively.
The ability of phage cocktail or phage cocktail with metal ions to induce acute toxicity in mice was described previously.40 Briefly, the mice were inoculated intraperitoneally with 200 μL of phage cocktail (109 PFU), 200 μL of phage cocktail (109 PFU) with 0.1 mM Ca2+, 200 μL of phage cocktail (109 PFU) with 0.1mM Zn2+ or 200 μL of PBS. Lungs, liver, and spleen from each group of mice (n=3) were taken for histopathological assessment at 7 days post inoculation. At the end of the experiment, the mice were sacrificed by CO2 asphyxiation method.
Statistical Analysis
All data were analyzed using GraphPad Prism 8.0 software (GraphPad Software Inc., La Jolla, CA, USA). Comparisons were performed through one- or two-way ANOVA followed by Tukey′s test. For the survival analysis of Galleria mellonella larvae, two-way ANOVA was performed. The data was represented from means ± standard deviation (SD) from three independent experiments. P < 0.05 was considered statistically significant.
Results
The Physical Properties of Four Phages PHB22a, PHB25a, PHB38a, and PHB40a Infecting Multiple MRSA Strains
The phages PHB22a, PHB25a, PHB38a, and PHB40a (MRSA S-18, S-11, S-16, and S-9 are their host strains, respectively) were isolated from environmental sewage using double-layer plate method. Then, the efficiency of plating (EOP) of each phage against 26 strains of MRSA was shown in Table S1. PHB25a could lyse 25 of 26 strains of MRSA tested in this study (Table S1). PHB25a showed a very low EOP for a part of MRSA, but PHB22a, PHB38a, and PHB40a could complement with PHB25a to improve the ability to kill host bacteria. Phages PHB22a, PHB25a, PHB38a, and PHB40a were combined into a phage cocktail, which infects all tested MRSA strains except MRSA S-17. Electron microscopy showed that three phages PHB22a, PHB25a, and PHB38a have a rectangular head and a long tail (Figure S1). The PHB40a has a regular icosahedral head and a long tail. According to the morphological characteristics and the International Committee on Taxonomy of Viruses (ICTV) classification, these phages were classified the members of the Siphoviridae family (Figure S1). Acid-base stability test indicated that they maintained excellent activity at pH 5.0 to 9.0 (Figure 1A-D). Thermolability tests showed that these phages were stable from 4 to 50°C (Figure 1E-H). One-step phage growth curve showed that the four phages had different latency and burst size, with latency ranging from 10 min to 20 min and burst size ranging from 25 to 180 phage particles per infected cell (PFU/cell) (Figure 1I-L). The complete genome of each phage has a linear double-stranded DNA, none of which show homology to any known virulence or antibiotic resistance genes. However, genomic analysis shows that the four phages are temperate phage because they carry the integrase and CI-like repressor. The NCBI GenBank accession numbers of these four phages was shown in Table S2. Phylogenetic analysis of phage large terminase subunit indicated that it belongs to the Siphoviridae family (Figure S2).
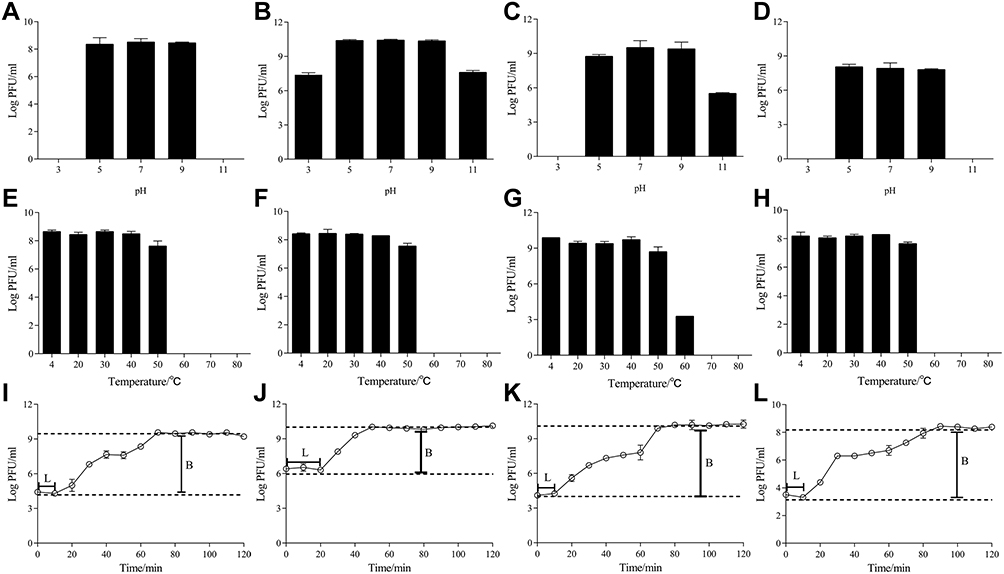 |
Figure 1 Biological properties of phages PHB22a, PHB25a, PHB38a and PHB40a. pH stability test of phages PHB22a (A), PHB25a (B), PHB38a (C), and PHB40a (D). Phages were incubated at 37°C for 1 h in buffer at pH 7.0 was considered 100% activity. Thermal stability test of phages PHB22a (E), PHB25a (F), PHB38a (G), and PHB40a (H). Phages were incubated at 37°C for 1 h was considered 100% activity. One step growth curves of phages PHB22a (I), PHB25a (J), PHB38a (K), and PHB40a (L) in host strain MRSA S-18. The average burst sizes were 37, 113, 180 and 25 for PHB22a, PHB25a, PHB38a, and PHB40a, respectively. And the latency periods of PHB22a, PHB25a, PHB38a, and PHB40a were 10 min, 20 min, 10 min, and 10 min respectively. L: Latency period; B: Burst size. Data are expressed as the mean ± SD. |
Supplementing with Metal Ions Enhances the Ability of Phage Cocktail to Remove Biofilms
In this study, the phage cocktail was made by composing of these four phages in equal proportions. The capacity of single phages and the phage cocktail were furtherly tested for removing the biofilms. The crystal violet staining assay showed that 69.1% (24-h-biofilm) and 66.7% (48-h-biofilm) of the biofilms could be removed with cocktail (108 PFU) treated, and most 59.5% (PHB25a) and 61.1% (PHB38a) of the biofilms could be removed with phage (108 PFU) treated (Figure S3). With phage (108 PFU) alone or phage cocktail (108 PFU) treated for 4 h, the results of the viable count showed the approximately 2.00 and 2.12 log CFU/cm2 (phage cocktail) or 1.69 and 1.97 log CFU/cm2 (phage alone) reduction, respectively (Figure S3). With advantages of phage cocktail, metal ions Ca2+ and Zn2+ mixed phage cocktail was performed subsequently. The results of crystal violet staining indicated that metal ions Ca2+ and Zn2+ mixed phage cocktail could not significantly change of absorbance values compared with the phage cocktail alone (Figure 2A and B). The viable-cell counts assay showed that the different concentrations of Ca2+ and Zn2+ mixed the phage cocktail significantly reduced the number of live bacteria (24-h or 48-h-old biofilms) compared with the phage cocktail alone (P < 0.01) (Figure 2C and D). Further, the 24-h-old or 48-h-old biofilms were treated with the phage cocktail (108 PFU) contained with 0.1 mM Ca2+ and Zn2+. Scanning electron microscopy assay indicated that a relatively small number of viable bacteria was observed in Ca2+ and Zn2+ treated groups, compared with phage cocktail alone group (Figure 3).
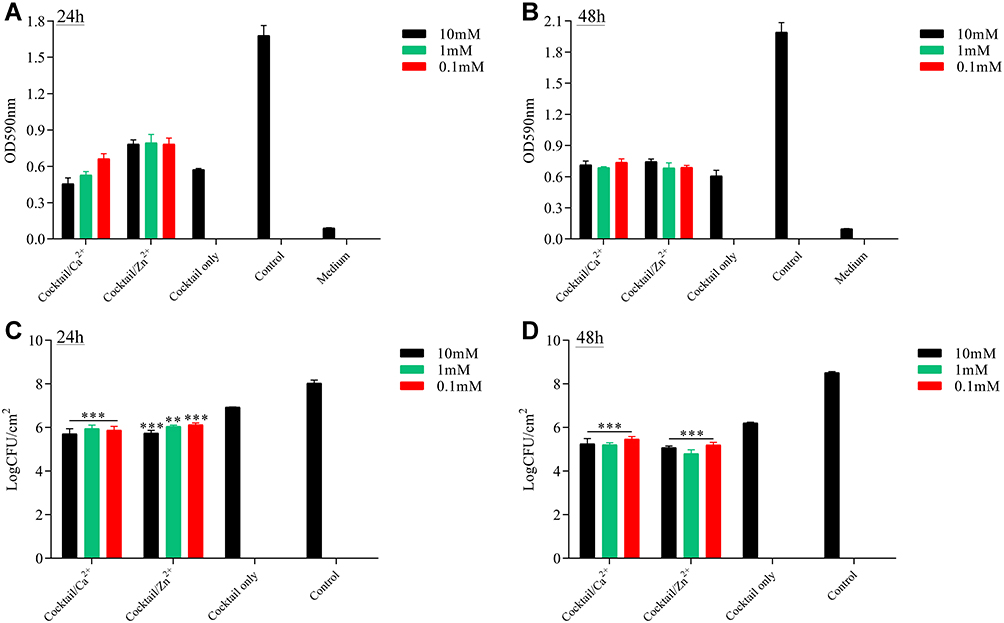 |
Figure 2 Antibiofilm activity of phage cocktail or cocktail with metal Ions Ca2+ and Zn2+. The value of OD590 after cocktail (108 PFU/mL) alone and cocktail (108 PFU/mL) with 10, 1.0 or 0.1mM Ca2+ or Zn2+ was applied to 24-h biofilm (A) and 48-h biofilm (B). The colony count (CFU/mL) after cocktail (108 PFU/mL) alone and cocktail (108 PFU/mL) with 10, 1.0 or 0.1mM Ca2+ or Zn2+ was applied to 24-h biofilm (C) and 48-h biofilm (D). Significant differences between the cocktail alone and cocktail with Ca2+ or Zn2+ treatment groups were determined by ANOVA (** p < 0.01, and *** p < 0.001). |
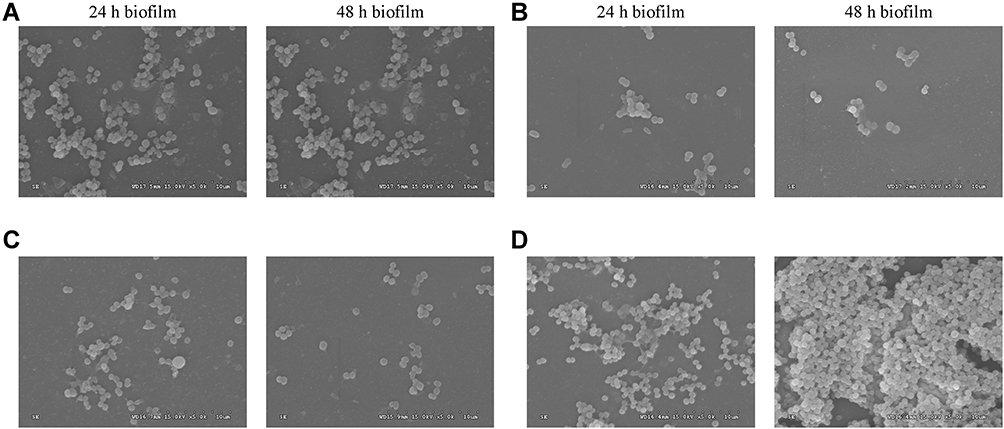 |
Figure 3 Scanning electron microscope observation. The magnification of the image is 5000. The 24-h and 48-h biofilm were treated with 1×108 PFU/mL cocktail alone (A), 1×108 PFU/mL cocktail contained with 0.1 mM Zn2+ (B), 1×108 PFU/mL cocktail contained with 0.1 mM Ca2+ (C) and PBS (D). |
The Antibacterial Effect of the Phage Cocktail in vitro and in vivo
The lysis kinetics of single phages or the phage cocktail was performed in LB medium (Figure 4A–E). The results of the lysis kinetics test of MRSA S-18 infected by the phage alone or phage cocktail showed that the OD600 values began to decrease at 0.5 h with MOI=10. Between MOI=0.001 and 10, phage cocktail could inhibit or kill the host strain S-18 in at least 10 h. Because, phage cocktail with Ca2+ and Zn2+ could enhance the ability to remove biofilms. To investigate the impact of single phages or phage cocktail on treatment of MRSA infection, we used the G. mellonella larvae modules for assessment of the effect of treatment in vivo. Administration of phage alone or phage cocktail 1 h after infection, resulted in the highest survival rate of 15% (3/20) in the single phage treatment group, while the phage cocktail treatment resulted in 35% (7/20) survival. Meanwhile, pretreatment with phage cocktail resulted in 85% (17/20) survival, and a single phage treatment resulted in the maximum 40% (8/20) survival. In addition, simultaneous treatment of S-18 and phage alone or phage cocktail, resulted in the highest survival rate of 30% (6/20) in the single phage treatment group, while the phage cocktail treatment resulted in 60% (12/20) survival (Figure 4F–J).
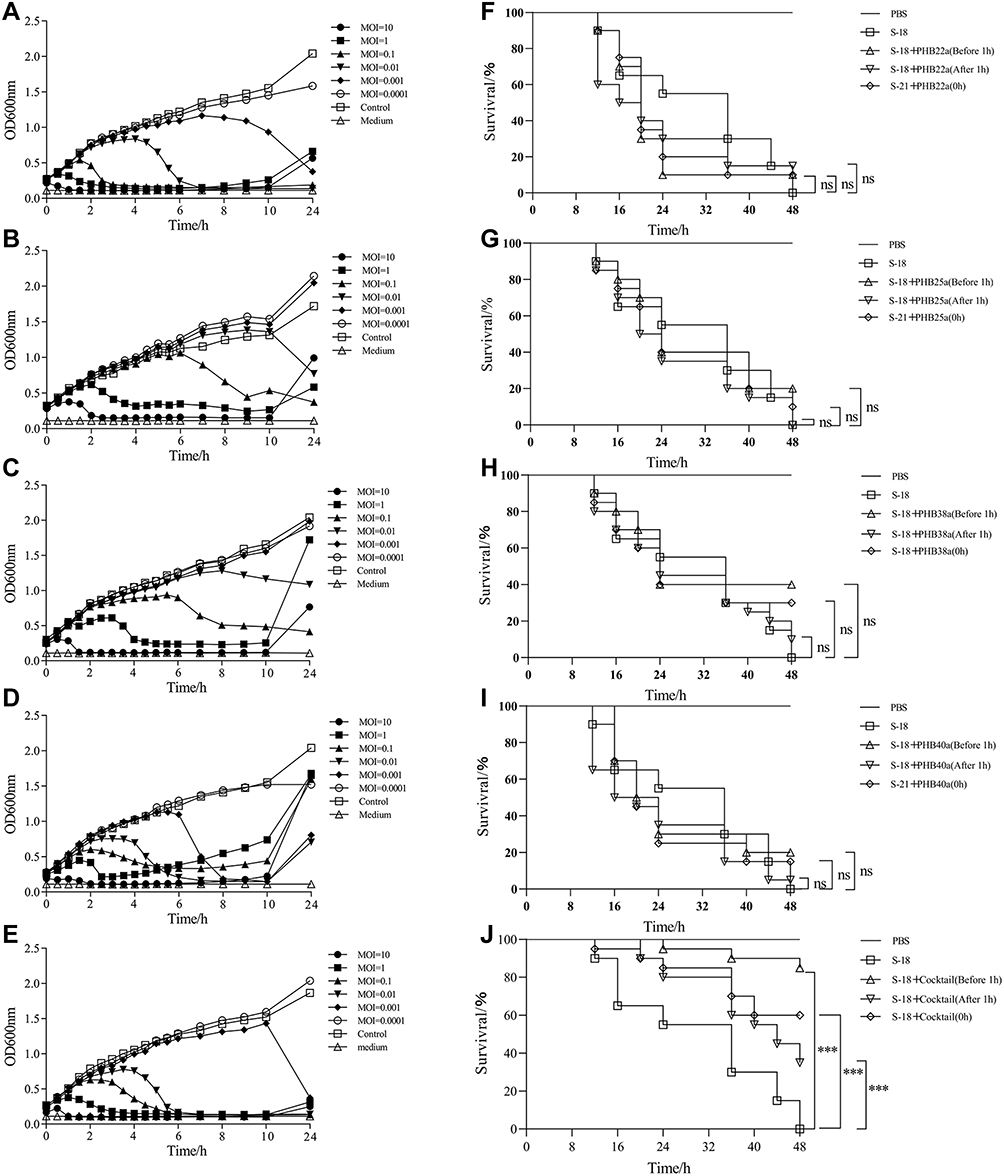 |
Figure 4 The effect of the phages or phage cocktail in vitro and in vivo. Effect of the phages PHB22a (A), PHB25a (B), PHB38a (C), PHB40a (D) and phage cocktail (E) on host bacteria growth of S-18. The therapeutic effect of phages PHB22a (F), PHB25a (G), PHB38a (H), PHB40a (I) and phage cocktail (J) on G. mellonella larvae with S-18 infection. Significant differences between the S-18 groups and single phages or cocktail treatment groups were determined by ANOVA (* *** p < 0.001). Data are expressed as means ± SD. |
Treatment with Phage Cocktail Containing Ca2+ and Zn2+ Increased the Survival of G. mellonella Larvae Against MRSA S-18 Infection
In pretreatment groups, compared with phage cocktail (85% survival), phage cocktail with 0.1 mM Ca2+ and Zn2+ resulted in 90% (cocktail with Ca2+) survival and 85% (cocktail with Zn2+) survival, respectively (Figure 5A). As showed in Figure 5B, administration of phage cocktail 1 h after infection, the larvae survival rate of the phage cocktail contained with 0.1 mM Ca2+ and Zn2+ resulted in 60% (12/20) survival, while phage cocktail alone resulted in 50% (10/20) survival. In simultaneously treated groups, phage cocktail with 0.1 mM Ca2+ and Zn2+ resulted in 65% (cocktail with Ca2+) survival and 70% (cocktail with Zn2+) survival, respectively (Figure 5C).
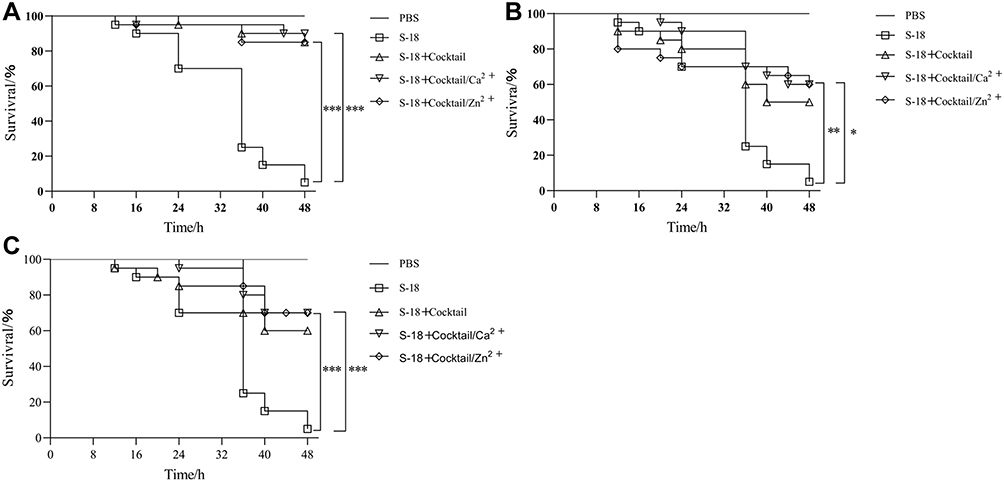 |
Figure 5 In vivo efficacy of phage cocktail with metal Ions Ca2+ and Zn2+ against MRSA S-18 in G. mellonella infection model. (A) Survival of G. mellonella treated with cocktail (4 × 107 PFU) alone and cocktail (4 × 107 PFU) with 0.1mM Ca2+ or Zn2+ before infection 1 h. (B) Survival of G. mellonella treated with cocktail (4 × 107 PFU) alone and cocktail (4 × 107 PFU) with 0.1mM Ca2+ or Zn2+ after infection 1 h. (C) Survival of G. mellonella simultaneously treated with cocktail (4 × 107 PFU) alone and cocktail (4 × 107 PFU) with 0.1mM Ca2+ or Zn2+. The larvae pre-treated with cocktail with 0.1mM Ca2+/Zn2+ showed significant increased survival rates compared to S-18 groups (p < 0.001). The larvae post-treated with cocktail with 0.1mM Ca2+ showed significant increased survival rates compared to S-18 groups (p < 0.01), and cocktail with 0.1mM Zn2+ showed significant increased survival rates compared to S-18 groups (p < 0.05). The larvae simultaneously treated with cocktail with 0.1mM Ca2+/Zn2+ showed significant increased survival rates compared to S-18 groups (p < 0.001). Significant differences between the S-18 groups and cocktail/Ca2+ or cocktail/Zn2+ groups were determined by ANOVA (* p < 0.05, ** p < 0.01, and *** p < 0.001). Data are expressed as means ± SD. |
Addition of Ca2+ and Zn2+ Enhance the Protection of Phage Cocktail Against MRSA S-18 Infection in Mouse Model
Phage cocktail with metal ions resulted in the better protection than cocktail alone in the G. mellonella modules. The results also showed that pretreatment resulted 100% survival in the cocktail (1 × 109 PFU) alone. And the cocktail (1 × 109 PFU) contained with metal ions resulted 100% survival. The results showed that pretreatment resulted in 66.7% (4/6) survival in the cocktail (5 × 108PFU) alone, 66.7% (4/6) survival in the cocktail (5 × 108PFU) with Ca2+, and 83.3% (5/6) survival in the cocktail (5 × 108PFU) with Zn2+ (Figure 6A).
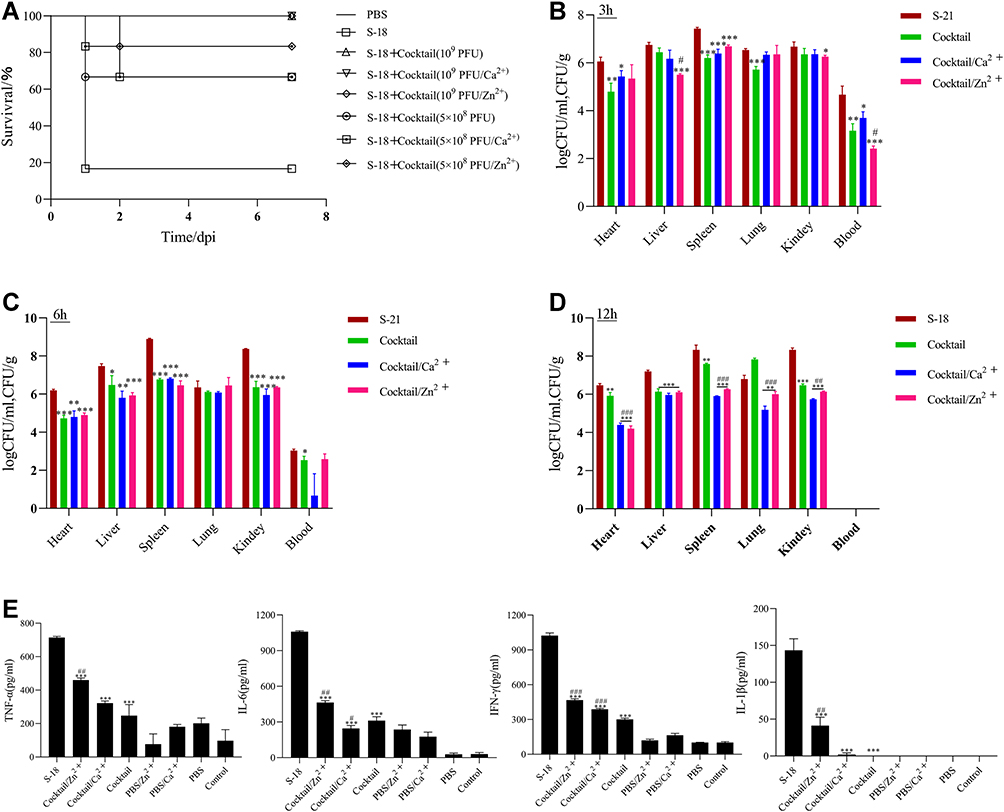 |
Figure 6 Protective effect of phage cocktail pretreatment in mice infected with MRSA S-18 strain. (A) Survival of mice were infected with MRSA S-18 treated with cocktail (1 × 109/5×108 PFU) alone, cocktail (1 × 109 /5×108 PFU) with 0.1mM Ca2+/Zn2+, and PBS. Bacterial abundances in different tissues from mice pre-treated with cocktail (1 × 109 PFU) alone or cocktail (1 × 109 PFU) with 0.1mM Ca2+/Zn2+ at 3h (B), 6h (C) 12h (D). (E) Levels of inflammatory cytokines in groups S-18, cocktail (1 × 109 PFU) alone, cocktail (1 × 109 PFU) with 0.1mM Ca2+ and cocktail (1 × 109 PFU) with 0.1mM Zn2+. Significant differences between the cocktail alone or cocktail with Ca2+/Zn2+ treatment groups and untreated groups were determined by ANOVA (* p < 0.05, ** p < 0.01, and *** p< 0.001). For the comparisons between the cocktail with Ca2+/Zn2+ treatment groups and the cocktail alone treatment groups, significance is indicated by ANOVA (#p < 0.05, ##p < 0.01, ### p < 0.001). Data are expressed as means ± SD from three independent experimental replicates (n = 3). |
Following cocktail pretreatment, the bacterial counts in the heart, spleen, lung, and kidney of the cocktail/Zn2+ group and the cocktail/Ca2+ group were significantly lower than the cocktail group at 12h post-infection (P < 0.01) (Figure 6D). Meanwhile, the bacterial counts in the blood in cocktail/Zn2+ group were significantly lower than the cocktail group at 3h post-infection (P < 0.05) (Figure 6B). Following cocktail simultaneous treatment, the bacterial counts in the liver, spleen, and lung of the cocktail/Zn2+ group were significantly lower than the cocktail group at 12h post-infection (P < 0.001). And the bacterial counts in the spleen, and lung of the cocktail/Ca2+ group were significantly lower than the cocktail group at 12h post-infection (P < 0.01). Following cocktail delay treatment, the bacterial counts in the spleen, and lung of the cocktail/Zn2+ group were significantly lower than the cocktail group at 12h post-infection (P < 0.05) (Figure S4).
Compared to S-18 group, a significant decrease in the levels of inflammatory factors, including TNF-α, IFN-γ, IL-6, and IL-1β, was observed in cocktail or cocktail with metal ions (Figure 6C). However, compared with cocktail alone group, a significant increase of inflammatory factors (TNF-α, IFN-γ, IL-6, and IL-1β) was observed in cocktail/Zn2+ group (P < 0.01). Meanwhile, a significantly increased of inflammatory factors (IFN-γ) was observed in cocktail/Ca2+ group compared to cocktail alone group (P < 0.01).
Pathological sections revealed no pathological changes were observed in the lung, liver, and spleen of the cocktail or cocktail Ca2+/Zn2+ treated mice compared with those of the PBS control group (Figure S5).
Discussion
S. aureus could colonize human skin and cause serious nosocomial infections.41 Although vancomycin has been widely used to treat MRSA infections in hospital patients, the widespread occurrence of vancomycin-intermediate S. aureus led to the failure of antibiotic treatment.7,42 Therefore, alternative strategies are urgently needed to overcome resistant S. aureus in hospitals. In this study, we composed a phage cocktail containing four phages isolated from sewage and explored the efficacy of the phage cocktail supplemented with Ca2+ and Zn2+ against MRSA strains in vitro and in vivo.
Phage cocktail is considered as a potential strategy for the prevention and treatment of drug-resistant bacterial infections.6,13–15 Furthermore, the therapeutic use of cocktail has been reported to improve the phage host range and reduce the emergence of phage-resistant mutants.43 A case of phages was successfully applied for the treatment of secondary drug-resistant bacterial infections in COVID-19 infection patients.44
The bacteria in the biofilms are protected by the biofilm matrix, which makes these fixed bacteria more likely to develop resistance to antibiotics than dissociated bacteria.45,46 Phages diffuse slowly in the viscous biofilm matrix, however, their derivatives (endolysins) have attracted attentions because they can degrade LPS, EPS, and CPS in biofilms.47 Our results showed that the addition of Ca2+ or Zn2+ to cocktail at a final concentration of 0.1 to 10 mM significantly enhanced the lysis of host cells in the 24-h-old and 48-h-old biofilms compared to the cocktail alone (P < 0.01). The previous reports indicated that zinc (Zn2+) are necessary for all living organisms, and it is essential for the activity of many metalloenzymes in vivo.48 Meanwhile, in our previous study, lysin contained metal ions Mg2+ and Mn2+ could significantly enhance the ability of removing alive bacteria in the biofilms.26 Moreover, Ca2+ and Mn2+ can enhance the antibacterial activity of S. aureus phage lysin LysCSA13.24 Because of the presence of calcium-binding site in the S. aureus phage lysin LysGH15, Ca2+ can promote the lytic activity of LysGH15.49 For chimeric phage lysin Csl2, the bactericidal capacity of lysin was enhanced by adding 10 mM Ca2+ and Mg2+ into medium.50 It has been reported that Ca2+ and Mg2+ could improve phage titer and enhance phage adsorption.51 Therefore, combined with our results, we predicted that Ca2+ and Zn2+ may promote the fluidity and stability of the cocktail in the biofilm and enhance the bactericidal activity of phages in biofilm. However, the different concentration of Ca2+ or Zn2+ had indistinctively affected on the ability of the phage to remove the biofilm. Therefore, we used a metal ion concentration of 0.1 mM for subsequent experiments. Meanwhile, we observed that no significant different of OD590 values in the cocktail contained metal ions compared to the cocktail alone group. One possible explanation is that crystal violet is mainly binds to negatively charged polysaccharides, proteins, and nucleic acids, which results in that crystal violet staining results cannot well reflect the changes of the number of sessile bacteria in the biofilms.52 Accordingly, further experiments are needed to reveal how Ca2+ and Zn2+ promote the phage cocktail to eliminate bacteria in the biofilm.
Previous in vivo studies of therapeutic potential of phages against S. aureus have used G. mellonella larvae infection model.53–55 For instance, the survival rate of larvae infected with MRSA is 20–35% after treatment with phages or phage cocktail.55 Pseudomonas aeruginosa phages resulted 50–60% survival of G. mellonella larvae.37,56 Meanwhile, it had been proven that intravenous phage cocktail is safe and tolerable for the treatment of infections caused by S. aureus.13 Then, some typical cases showed that phages could successfully cure the patients infected with MRSA,57,58 and rescue 91% of mice infected with MRSA from death.59 However, little research used mouse and G. mellonella larvae models to evaluate the efficiency of phage cocktail contained with metal ions against MRSA in vivo. In our study, phage cocktail treatment of larvae infected with MRSA could increase the survival rate of larvae by at least 20% compared with a single phage. The possible reasons are that multiple phages have synergistic action and reduce phage-resistant bacteria in vivo.37,60 The survival of mice in cocktail (5 × 108PFU) alone group was 66.7%, while the survival of mice in cocktail (5 × 108PFU) with Zn2+ group was 88.3%. It is worth noting that the survival of mice in the cocktail (5×108PFU) alone group was 66.7%, while the survival of mice in the cocktail (5×108PFU) with Ca2+ group was 88.3% at 24h post-infection. Meanwhile, we found that phage cocktail containing 0.1 mM Ca2+ and Zn2+ treatment could improve the survival rate of larvae by 10% at 48 h, and significantly reduce the bacterial load in mouse heart, spleen, lung, and kidney at 12 h (P < 0.01). Wang et al (2020) showed that Mn2+, which can activate innate immune by inducing type I–IFN response and cytokine production, is essential for the host to defense against viruses.60,61 Pathogen recognition receptors (PRRs), such as the Toll-like receptors (TLRs) can recognize invading bacteria. Specifically, TLR2 senses staphylococcal lipoproteins and TLR7, 9, and 13 recognizes staphylococcal nucleic acids.62 At the same time, when MRSA infection occurs, its lipoprotein promotes TLR2/1 to participate in the production of IL-8.63 Surprisingly, in our research, we found that phage cocktail supplemented with Ca2+ and Zn2+ could significantly increase the level of inflammatory factors in mice compared to the phage cocktail alone group, but did not lead to pathological changes of mouse organs (Figure S5). One possible explanation is that because the phage cocktail with Ca2+/Zn2+ could destroy more bacteria than the phage cocktail alone, the lipoproteins and nucleic acids released by the destroyed bacteria are sensed by TLRs and stimulate the production of more inflammatory factors. In y also play an indispensable role when the phage cocktail eliminates bacteria in mouse organs.
In this study, phage cocktail can infect 25 out of 26 clinically isolated MRSA strains. Although these phages have a broad host range tested on MRSA strains, they were definitively classified as temperate phage. Although lytic phages are generally permitted to treat bacterial infections, lytic phage cocktails also have been shown numerous times to be detrimental given inhibition and other aspects of phage–phage interactions.64–66 At present, several studies have also begun to explore the potential of temperate phages. For example, temperate phages (ZoeJ and BPs) were genetically engineered into lytic phages, and then combined with a lytic phage (Muddy) to form a phage cocktail, which is the first use of bacteriophage in the treatment of human mycobacterial infection and the first use of engineered bacteriophage.67 Additionally, phage cocktail composed of temperate phages for the treatment of acute C. difficile infection.68 Nevertheless, there are still many concerns about temperate phages for clinical use and also not allowed by the regulatory bodies. The main reason is the ability of temperate phages to mediate horizontal gene transfer between bacterial genomes through generalized and specialized transduction.69 Fortunately, the four phages did not carry known virulence and resistance genes in this study. Thus, prophage genes such as integrase should be removed by genetic engineering prior to treatment with the four phages to avoid potential lysogeny and enhance bactericidal activity. Recent studies have shown that it has become possible to use synthetic biology to make temperate phages into high-quality virulent phages.70 Take together, it is also indicated that once we solve the temperate phage, the phage cocktail will be used for phage therapy in animals and humans in the future.
Conclusion
Taken together, we isolated four novel phages and composed a phage cocktail with these phages, which showed admirable bactericidal activity in vivo and in vitro. More importantly, we found that the addition of Ca2+ and Zn2+ could enhance the ability of the phage cocktail to eliminate biofilm immobilized bacteria and the prophylactic effect of the cocktail on S. aureus infection in larvae and mouse. However, all prophage-related genes should be removed by genetic engineering prior to treatment with this phage cocktail to avoid potential lysogeny and enhance bactericidal activity. In brief, our study indicated that the phage cocktail supplemented with metal ions could be an effective approach to solve the drug-resistant S. aureus infections.
Approval for Use of Animal Subjects
This study was approved by the Laboratory Animal Monitoring Committee of Huazhong Agricultural University, and carried out in accordance with the corresponding guidelines for laboratory animal operations in Huazhong Agricultural University. The corresponding ethical approval code is HZAUMO-2020-0063.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grant from the National Program on Key Research Project of China (2018YFD0500204) and the Fundamental Research Funds for the Central Universities (2662016PY004).
Disclosure
We have no conflicts of interest to declare.
References
1. David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev. 2010;23(3):616–687.
2. Michels R, Last K, Becker SL, Papan C. Update on Coagulase-Negative Staphylococci-What the Clinician Should Know. Microorganisms. 2021;9(4):625.
3. Foster AP. Staphylococcal skin disease in livestock. Vet Dermatol. 2012;23(4):342–351, e363.
4. Lowder BV, Guinane CM, Ben Zakour NL, et al. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc Natl Acad Sci U S A. 2009;106(46):19545–19550.
5. O’Neill J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Review on Antimicrobial Resistance. London, UK; 2016.
6. Lehman SM, Mearns G, Rankin D, et al. Design and Preclinical Development of a Phage Product for the Treatment of Antibiotic-Resistant Staphylococcus aureus Infections. Viruses. 2019;11(1):53.
7. Cong Y, Yang S, Rao X. Vancomycin resistant Staphylococcus aureus infections: a review of case updating and clinical features. J Adv Res. 2020;21:169–176.
8. Yang H, Xu J, Li W, et al. Staphylococcus aureus virulence attenuation and immune clearance mediated by a phage lysin-derived protein. EMBO j. 2018;37:17.
9. Shkoporov AN, Clooney AG, Sutton TDS, et al. The Human Gut Virome Is Highly Diverse, Stable, and Individual Specific. Cell Host Microbe. 2019;26(4):527–541.e525.
10. Song J, Ruan H, Chen L, et al. Potential of bacteriophages as disinfectants to control of Staphylococcus aureus biofilms. BMC Microbiol. 2021;21(1):57.
11. Gutiérrez D, Vandenheuvel D, Martínez B, Rodríguez A, Lavigne R, García P. Two Phages, phiIPLA-RODI and phiIPLA-C1C, Lyse Mono- and Dual-Species Staphylococcal Biofilms. Appl Environ Microbiol. 2015;81(10):3336–3348.
12. Kifelew LG, Warner MS, Morales S, et al. Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms. Viruses. 2020;12(5):536.
13. Petrovic Fabijan A, Lin RCY, Ho J, Maddocks S, Ben Zakour NL, Iredell JR. Safety of bacteriophage therapy in severe Staphylococcus aureus infection. Nat Microbiol. 2020;5(3):465–472.
14. Kifelew LG, Warner MS, Morales S, et al. Efficacy of phage cocktail AB-SA01 therapy in diabetic mouse wound infections caused by multidrug-resistant Staphylococcus aureus. BMC Microbiol. 2020;20(1):204.
15. Ooi ML, Drilling AJ, Morales S, et al. Safety and Tolerability of Bacteriophage Therapy for Chronic Rhinosinusitis Due to Staphylococcus aureus. JAMA Otolaryngol Head Neck Surg. 2019;145(8):723–729.
16. Ingmer H, Gerlach D, Wolz C. Temperate Phages of Staphylococcus aureus. Microbiol Spectr. 2019;7(5):234.
17. Touchon M, Bernheim A, Rocha EP. Genetic and life-history traits associated with the distribution of prophages in bacteria. Isme j. 2016;10(11):2744–2754.
18. Feng T, Leptihn S, Dong K, et al. JD419, a Staphylococcus aureus Phage With a Unique Morphology and Broad Host Range. Front Microbiol. 2021;12:602902.
19. Meader E, Mayer MJ, Steverding D, Carding SR, Narbad A. Evaluation of bacteriophage therapy to control Clostridium difficile and toxin production in an in vitro human colon model system. Anaerobe. 2013;22:25–30.
20. Chung IY, Sim N, Cho YH. Antibacterial efficacy of temperate phage-mediated inhibition of bacterial group motilities. Antimicrob Agents Chemother. 2012;56(11):5612–5617.
21. Monteiro R, Pires DP, Costa AR, Azeredo J. Phage Therapy: going Temperate? Trends Microbiol. 2019;27(4):368–378.
22. Ribeiro M, Monteiro FJ, Ferraz MP. Infection of orthopedic implants with emphasis on bacterial adhesion process and techniques used in studying bacterial-material interactions. Biomatter. 2012;2(4):176–194.
23. Molina-Manso D, Del Prado G, Ortiz-Pérez A, et al. In vitro susceptibility to antibiotics of staphylococci in biofilms isolated from orthopaedic infections. Int J Antimicrob Agents. 2013;41(6):521–523.
24. Cha Y, Son B, Ryu S. Effective removal of staphylococcal biofilms on various food contact surfaces by Staphylococcus aureus phage endolysin LysCSA13. Food Microbiol. 2019;84:103245.
25. Zhang L, Shahin K, Soleimani-Delfan A, et al. Phage JS02, a putative temperate phage, a novel biofilm-degrading agent for Staphylococcus aureus. Lett Appl Microbiol. 2022.
26. Yang D, Chen Y, Sun E, Hua L, Peng Z, Wu B. Characterization of a Lytic Bacteriophage vB_EfaS_PHB08 Harboring Endolysin Lys08 Against Enterococcus faecalis Biofilms. Microorganisms. 2020;8:9.
27. Kirby-Bauer Disk JH. Diffusion Susceptibility Test Protocol. Am Soc Microbiol. 2016;1:1–23.
28. Mangieri N, Picozzi C, Cocuzzi R, Foschino R. Evaluation of a Potential Bacteriophage Cocktail for the Control of Shiga-Toxin Producing Escherichia coli in Food. Front Microbiol. 2020;11:1801.
29. Chen Y, Yang L, Yang D, et al. Specific Integration of Temperate Phage Decreases the Pathogenicity of Host Bacteria. Front Cell Infect Microbiol. 2020;10:14.
30. Chang HC, Chen CR, Lin JW, et al. Isolation and characterization of novel giant Stenotrophomonas maltophilia phage phiSMA5. Appl Environ Microbiol. 2005;71(3):1387–1393.
31. Thomas JA, Hardies SC, Rolando M, et al. Complete genomic sequence and mass spectrometric analysis of highly diverse, atypical Bacillus thuringiensis phage 0305phi8-36. Virology. 2007;368(2):405–421.
32. Chen Y, Li X, Song J, et al. Isolation and characterization of a novel temperate bacteriophage from gut-associated Escherichia within black soldier fly larvae (Hermetia illucens L. [Diptera: stratiomyidae]). Arch Virol. 2019;164(9):2277–2284.
33. Li R, Zhu H, Ruan J, et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010;20(2):265–272.
34. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729.
35. Oliveira H, Mendes A, Fraga AG, et al. K2 Capsule Depolymerase Is Highly Stable, Is Refractory to Resistance, and Protects Larvae and Mice from Acinetobacter baumannii Sepsis. Appl Environ Microbiol. 2019;85:17.
36. Chen Y, Yang L, Sun E, Song J, Wu B. Characterisation of a newly detected bacteriophage infecting Bordetella bronchiseptica in swine. Arch Virol. 2019;164(1):33–40.
37. Forti F, Roach DR, Cafora M, et al. Design of a Broad-Range Bacteriophage Cocktail That Reduces Pseudomonas aeruginosa Biofilms and Treats Acute Infections in Two Animal Models. Antimicrob Agents Chemother. 2018;62:6.
38. Yu L, Wang S, Guo Z, et al. A guard-killer phage cocktail effectively lyses the host and inhibits the development of phage-resistant strains of Escherichia coli. Appl Microbiol Biotechnol. 2018;102(2):971–983.
39. Velikova N, Kavanagh K, Wells JM. Evaluation of Galleria mellonella larvae for studying the virulence of Streptococcus suis. BMC Microbiol. 2016;16(1):291.
40. Chen Y, Li X, Wang S, et al. A Novel Tail-Associated O91-Specific Polysaccharide Depolymerase from a Podophage Reveals Lytic Efficacy of Shiga Toxin-Producing Escherichia coli. Appl Environ Microbiol. 2020;86(9):56.
41. Pandey R, Mishra SK, Shrestha A. Characterisation of ESKAPE Pathogens with Special Reference to Multidrug Resistance and Biofilm Production in a Nepalese Hospital. Infect Drug Resist. 2021;14:2201–2212.
42. Foster TJ. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol Rev. 2017;41(3):430–449.
43. Chan BK, Abedon ST. Phage therapy pharmacology phage cocktails. Adv Appl Microbiol. 2012;78:1–23.
44. Wu N, Dai J, Guo M, et al. Pre-optimized phage therapy on secondary Acinetobacter baumannii infection in four critical COVID-19 patients. Emerg Microbes Infect. 2021;10(1):612–618.
45. Anderl JN, Franklin MJ, Stewart PS. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob Agents Chemother. 2000;44(7):1818–1824.
46. Van Laar TA, Chen T, You T, Leung KP. Sublethal concentrations of carbapenems alter cell morphology and genomic expression of Klebsiella pneumoniae biofilms. Antimicrob Agents Chemother. 2015;59(3):1707–1717.
47. Hughes KA, Sutherland IW, Jones MV. Biofilm susceptibility to bacteriophage attack: the role of phage-borne polysaccharide depolymerase. Microbiology. 1998;144(Pt 11):3039–3047.
48. Hood MI, Skaar EP. Nutritional immunity: transition metals at the pathogen-host interface. Nat Rev Microbiol. 2012;10(8):525–537.
49. Gu J, Feng Y, Feng X, et al. Structural and biochemical characterization reveals LysGH15 as an unprecedented “EF-hand-like” calcium-binding phage lysin. PLoS Pathog. 2014;10(5):e1004109.
50. Vázquez R, Domenech M, Iglesias-Bexiga M, Menéndez M, García P. Csl2, a novel chimeric bacteriophage lysin to fight infections caused by Streptococcus suis, an emerging zoonotic pathogen. Sci Rep. 2017;7(1):16506.
51. Zhang X, Lan Y, Jiao W, et al. Isolation and Characterization of a Novel Virulent Phage of Lactobacillus casei ATCC 393. Food Environ Virol. 2015;7(4):333–341.
52. Latka A, Drulis-Kawa Z. Advantages and limitations of microtiter biofilm assays in the model of antibiofilm activity of Klebsiella phage KP34 and its depolymerase. Sci Rep. 2020;10(1):20338.
53. Mannala GK, Rupp M, Alagboso F, et al. Galleria mellonella as an alternative in vivo model to study bacterial biofilms on stainless steel and titanium implants. Altex. 2021;38(2):245–252.
54. Xie N, Jiang L, Chen M, et al. In vitro and in vivo Antibacterial Activity of Linezolid Plus Fosfomycin Against Staphylococcus aureus with Resistance to One Drug. Infect Drug Resist. 2021;14:639–649.
55. Ngassam-Tchamba C, Duprez JN, Fergestad M, et al. In vitro and in vivo assessment of phage therapy against Staphylococcus aureus causing bovine mastitis. J Glob Antimicrob Resist. 2020;22:762–770.
56. Jeon J, Yong D. Two Novel Bacteriophages Improve Survival in Galleria mellonella Infection and Mouse Acute Pneumonia Models Infected with Extensively Drug-Resistant Pseudomonas aeruginosa. Appl Environ Microbiol. 2019;85:9.
57. Fish R, Kutter E, Bryan D, Wheat G, Kuhl S. Resolving Digital Staphylococcal Osteomyelitis Using Bacteriophage-A Case Report. Antibiotics. 2018;7(4):8978.
58. Morozova VV, Vlassov VV, Tikunova NV. Applications of Bacteriophages in the Treatment of Localized Infections in Humans. Front Microbiol. 2018;9:1696.
59. Prazak J, Valente L, Iten M, et al. Benefits of aerosolized phages for the treatment of pneumonia due to methicillin-resistant Staphylococcus aureus (MRSA): an experimental study in rats. J Infect Dis. 2021.
60. Roach DR, Leung CY, Henry M, et al. Synergy between the Host Immune System and Bacteriophage Is Essential for Successful Phage Therapy against an Acute Respiratory Pathogen. Cell Host Microbe. 2017;22(1):38–47.e34.
61. Lv M, Chen M, Zhang R, et al. Manganese is critical for antitumor immune responses via cGAS-STING and improves the efficacy of clinical immunotherapy. Cell Res. 2020;30(11):966–979.
62. Lentini G, Famà A, De Gaetano GV, et al. Role of Endosomal TLRs in Staphylococcus aureus Infection. J Immunol. 2021;207(5):1448–1455.
63. Hook JS, Patel PA, O’Malley A, et al. Lipoproteins from Staphylococcus aureus Drive Neutrophil Extracellular Trap Formation in a TLR2/1- and PAD-Dependent Manner. J Immunol. 2021;207(3):966–973.
64. Molina F, Menor-Flores M, Fernández L, Vega-Rodríguez MA, García P. Systematic analysis of putative phage-phage interactions on minimum-sized phage cocktails. Sci Rep. 2022;12(1):2458.
65. Ram G, Chen J, Kumar K, et al. Staphylococcal pathogenicity island interference with helper phage reproduction is a paradigm of molecular parasitism. Proc Natl Acad Sci U S A. 2012;109(40):16300–16305.
66. Abedon ST, Thomas-Abedon C. Phage therapy pharmacology. Curr Pharm Biotechnol. 2010;11(1):28–47.
67. Dedrick RM, Guerrero-Bustamante CA, Garlena RA, et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat Med. 2019;25(5):730–733.
68. Nale JY, Spencer J, Hargreaves KR, et al. Bacteriophage Combinations Significantly Reduce Clostridium difficile Growth In Vitro and Proliferation In Vivo. Antimicrob Agents Chemother. 2016;60(2):968–981.
69. Harrison E, Brockhurst MA. Ecological and Evolutionary Benefits of Temperate Phage: what Does or Doesn’t Kill You Makes You Stronger. Bioessays. 2017;39:12.
70. Kilcher S, Studer P, Muessner C, Klumpp J, Loessner MJ. Cross-genus rebooting of custom-made, synthetic bacteriophage genomes in L-form bacteria. Proc Natl Acad Sci U S A. 2018;115(3):567–572.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.