")
Back to Journals » Infection and Drug Resistance » Volume 15
Epidemiology of Klebsiella michiganensis Carrying Multidrug-Resistant IncHI5 Plasmids in the Southeast Coastal Area of China
Authors Luo X, Zhang J, Yuan M, Mou S, Xu M, Hu D, Ma Q, Sun L, Li P, Song Z, Yu L, Mu K
Received 26 January 2022
Accepted for publication 26 March 2022
Published 13 April 2022 Volume 2022:15 Pages 1831—1843
DOI https://doi.org/10.2147/IDR.S358839
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Xinhua Luo,1,* Jin Zhang,1,* Min Yuan,2 Sihua Mou,1 Mengqiao Xu,1 Dakang Hu,1 Qinfei Ma,1 Lingfen Sun,1 Piaopiao Li,1 Zhiwei Song,1 Lianhua Yu,1 Kai Mu3,4
1Department of Clinical Laboratory Medicine, Taizhou Municipal Hospital Affiliated with Taizhou University, Taizhou, 318000, People’s Republic of China; 2State Key Laboratory for Infectious Diseases Prevention and Control, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Disease, National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention, Beijing, 102206, People’s Republic of China; 3Beijing Institute of Radiation Medicine, Beijing, 100850, People’s Republic of China; 4Beijing Key Laboratory of New Molecular Diagnosis Technologies for Infectious Diseases, Beijing, 100850, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Kai Mu, Beijing Key Laboratory of New Molecular Diagnosis Technologies for Infectious Diseases, Beijing, 100850, People’s Republic of China, Tel +86-010-66874794, Email [email protected] Lianhua Yu, Department of Clinical Laboratory Medicine, Taizhou Municipal Hospital affiliated with Taizhou University, Taizhou, 318000, People’s Republic of China, Email [email protected]
Purpose: This study aimed to explore the genomic characterization of multidrug-resistant IncHI5-carrying Klebsiella michiganensis strains and detailed genomic dissection of the IncHI5 plasmids.
Materials and Methods: Through whole-genome sequencing, the IncHI5 plasmid pK92-qnrS was obtained from a single clinical K. michiganensis isolate K92. All complete genomes of K. michiganensis strains from the Genome database of NCBI were collected and used to construct a maximum likelihood (ML) phylogenetic tree. The epidemiology and geographic distribution of all the K. michiganensis strains were conducted. An extensive comparison of the seven IncHI5 plasmids of K. michiganensis (one from this study, six from GenBank) was applied.
Results: This study revealed that all K. michiganensis strains carrying IncHI5 plasmids from different clonal groups were located in the southeast coastal area of China. The backbone regions of IncHI5 plasmids were composed of replicon (repHI5B and repFIB), partition (parABC), and conjugal transfer (tra1/tra2). The main accessory resistant regions of IncHI5 could be divided into two categories, Tn 1696-related region and Tn 6535-related region. These seven IncHI5 plasmids carried multiple drug-resistance genes which were all mediated by the mobile genetic elements (MGEs).
Conclusion: Data presented here help to provide an overall in-depth understanding of epidemiology and geographic distribution of IncHI5-carrying K. michiganensis and the structure and evolutionary history of IncHI5 plasmids.
Keywords: Klebsiella michiganensis, mobile genetic elements, Tn 1696, Tn 6535, antimicrobial resistance
Introduction
Klebsiella michiganensis is a potential emerging pathogen causing serious and difficult-to-treat nosocomial infections. In 2012, K. michiganensis was first recovered from a toothbrush holder and relatedness with Klebsiella oxytoca was originally recognized by a list of housekeeping genes, although it was finally proved by DNA-DNA hybridization (DDH) that it should be a new species.1 Since then, clinical cases caused by K. michiganensis have been reported worldwide. Some K. michiganensis isolates showed antibiotic resistance, and even multidrug-resistance (MDR) through the production of carbapenemases.2–4 In existing reports, there have been multiple genetic recombination and integration events of drug resistance genes through mobile genetic elements (MGEs) such as transposons and plasmids carried by K. michiganensis.5–8 The existence and properties of plasmids make K. michiganensis isolates a serious threat, and it is of interest to conduct in-depth studies of the structure and function of the plasmids carried by K. michiganensis.
IncHI5 is one of the five subgroups of plasmids subdivided under IncHI based on its replication genes repHI5B and repFIB.9 IncHI5 plasmids, like most IncHI plasmids, have a size usually greater than 200 kb and a wide host range.10 IncHI5 plasmids are significant vectors carrying genes encoding resistance to heavy metals and antibiotics.11 It is necessary to continuously monitor the prevalence of IncHI5 plasmids in different species.12 However, research on the structure and evolution of IncHI5 plasmids is still limited, and there is a lack of an overall study on IncHI5 plasmids within specific species.13
IncHI5 plasmids carried by K. michiganensis were first reported in 2012 in a strain from Miaoli, Taiwan, China.14 There are limited reports on IncHI5 plasmids carried by K. michiganensis. The sequence of repHI5B (the key identified replicon of IncHI5 plasmids) was used to perform BLAST alignment against the GenBank15 of the National Center for Biotechnology Information (NCBI) to cover all known IncHI5 plasmids. From the alignment, a total of six IncHI5 plasmids whose host strain were K. michiganensis were obtained (last accessed on December 12, 2021): pKOX_R1 (accession number CP003684),14 p12084-HI5 (accession number MW810613), pCP024641 (accession number CP024641), pJNQH491-2 (accession number CP075883),16 p1 (accession number CP067094), and pKOX7525_1 (accession number CP065475).17
In this work, we characterized IncHI5 plasmid pK92-qnrS (accession number OL828743; from Taizhou, Zhejiang, China) and presented further comprehensive genomic comparison of all IncHI5 plasmids carried by K. michiganensis (six plasmids mentioned above and pK92-qnrS), and discussed epidemiology and geographic distribution of K. michiganensis, to gain a deeper understanding of the genomic characterization of multidrug-resistant IncHI5-carrying K. michiganensis strains and detailed genomic dissection of the IncHI5 plasmids.
Materials and Methods
Identification of Bacterial Strain and Conjugal Transfer
K. michiganensis strain K92 was isolated from a patient’s sputum in Taizhou Municipal Hospital affiliated with Taizhou University of China in 2018. Strain K92 was initially identified as K. oxytoca by Vitek 2. Moreover, bacterial species identification was performed using chromosomal genome sequence-based average nucleotide identity (ANI) analysis and finally proved that K92 belongs to K. michiganensis.18 THO-011 (accession number AP022547), the standard strain of K. michiganensis, was used as reference. The OrthoANI value between THO-011 and K92 was 98.83% (> 95% cut-off), which was calculated by OAT 0.93.1.19
Conjugal transfer experiments were carried out with sodium azide-resistant Escherichia coli J53 being used as a recipient, and strain K92 as a donor. The donor and recipient strains were grown in three milliliters (mL) brain heart infusion (BHI) broth overnight at 37°C. And then, 50 μL of donor strain culture was mixed with 500 μL of recipient strain culture (v:v = 1:10) and 4.5 mL of fresh BHI broth. In addition, 100 μL of the mixture was applied onto a cellulose filter membrane (pore size, 0.22 μm) already placed on a BHI agar plate. After incubation at 25 °C for 16–18 h, the filter membrane was taken out and vortexed in 1 mL of BHI broth. The vortex mixtures were plated on BHI agar plates containing 40 mg/L ciprofloxacin and 150 mg/L sodium azide for the selection of the E. coli J53 transconjugants. However, repeated conjugation experiments failed to transfer the qnrS marker from K92 to E. coli J53 (sodium azide resistance).
Sequencing and Sequence Assembly
Genomic DNA was extracted from strain K92 using a Gentra Puregene Yeast/Bact. Kit (Qiagen, Valencia, CA). Libraries were prepared separately using the TruePrepTM DNA Library Prep Kit V2 and the SQU-LSK109 Ligation Sequencing kit. After the preparation of the library was completed, it was separately sequenced on an Illumina HiSeq X Ten platform using the 2 × 150-base-pair paired-end mode (Illumina Inc., San Diego, CA, USA) and GridION X5 platform using the long reads (Oxford Nanopore, UK), the sequencing depth was 100x. To improve the reliability of data processing, raw data from HiSeq X Ten platform and GridION X5 platform were trimmed to obtain the high-quality clean reads (clean data) by Canu v1.8 (https://canu.readthedocs.io/en/latest/index.html). The paired-end short Illumina reads and the long Nanopore reads were assembled de novo utilizing Unicycler (v0.4.5) (https://github.com/rrwick/Unicycler).
Whole Genome Phylogeny and Genetic Background Analysis
A total of 32 publicly available complete genome sequences of K. michiganensis were downloaded from NCBI (last accessed on December 12, 2021), which were isolated from various sources in 2011–2021. Genomes were aligned against the reference genome (THO-011, the standard strain of K. michiganensis) using MUMmer v3.1,20 to identify the SNPs in the backbone regions. A total of 484632 SNPs (from 3011423-bp length core genomes) were identified in the seven K. michiganensis strains chromosome sequences, and an ML phylogenetic tree was constructed using this SNPs dataset. Phylogenetic trees were drawn using the Interactive Tree of Life (iTOL) programs.21 The evolution analysis of the seven IncHI5 plasmids’ backbone regions was consistent with the above-mentioned phylogenetic analysis method.
Sequence Annotation and Comparison
Sequence annotation and prediction of ORFs and pseudogenes were performed using RAST2.0,22 and further manual annotation was done with BLASTP/BLASTN23 against the UniProtKB/Swiss-Prot24 and RefSeq databases.25 Annotation of drug resistance genes, MGEs, and other features was performed using online databases such as CARD,26 ResFinder,27 ISfinder,28 INTEGRALL,29 and the Tn Number Registry.30 MUSCLE 3.8.3131 and BLASTN were used for multiple and pairwise sequence comparisons. The genome map was drawn using Inkscape 1.1 (https://inkscape.org/en). The heatmap of resistance genes carried by IncHI5 plasmids included in this study was drawn with iTOL.
Nucleotide Sequence Accession Number
The complete sequences of the chromosome of K92 and plasmid pK92-qnrS were submitted to GenBank database, under accession numbers CP089315 and OL828743, respectively.
Results
Genomic Characterization of K. michiganensis Genomes
A total of 32 complete genomes of K. michiganensis strains with geographic distribution and other background information were obtained from the Genome database of NCBI (last accessed on December 12, 2021) (Figure 1A and Supplementary Table S1). The chromosomal sequences of these 32 strains were aligned with the chromosomal sequence (accession number CP089315) of K. michiganensis strain K92 in this study, and strain HS11286 (accession number CP003200),32 the standard strain of Klebsiella pneumoniae, was used as the outer group, and a total of 484,632 SNPs were obtained. Among them, K. michiganensis strain RC10 (accession number CP011077) was excluded because of the low coverage value of the comparison with core genome of the 32 strains (14.8%). A maximum likelihood (ML) phylogenetic tree was constructed using these SNPs’ dataset (Figure 1B). The population structure showed that K. michiganensis strains carrying IncHI5 plasmids were scattered in different clades of the entire phylogenetic tree (orange dots and red star in Figure 1B), instead of being clustered and distributed in a certain branch of the tree, which may imply the acquisition of IncHI5 plasmids was an overall behavior of K. michiganensis and was not related to a specific branch of K. michiganensis. The geographic distribution showed that K. michiganensis strains were mainly distributed in North America, Europe, East Asia, and Oceania (Figure 1A). Although K. michiganensis had also been reported in South Africa before,6,33 the complete genome was not available in the NCBI Genome database, so K. michiganensis reported in South Africa was not included in this study. Sixteen of these strains originated in China. Coincidentally, all the K. michiganensis strains carrying IncHI5 plasmids were located in the southeast coastal area of China such as Zhejiang, Fujian, Taiwan, and Guangdong (orange dots and red star in Figure 1A–C). This geographic distribution indicated that K. michiganensis strains carrying IncHI5 plasmids were spread in a southeast coastal area of China, although the underlying mechanism is currently unclear.
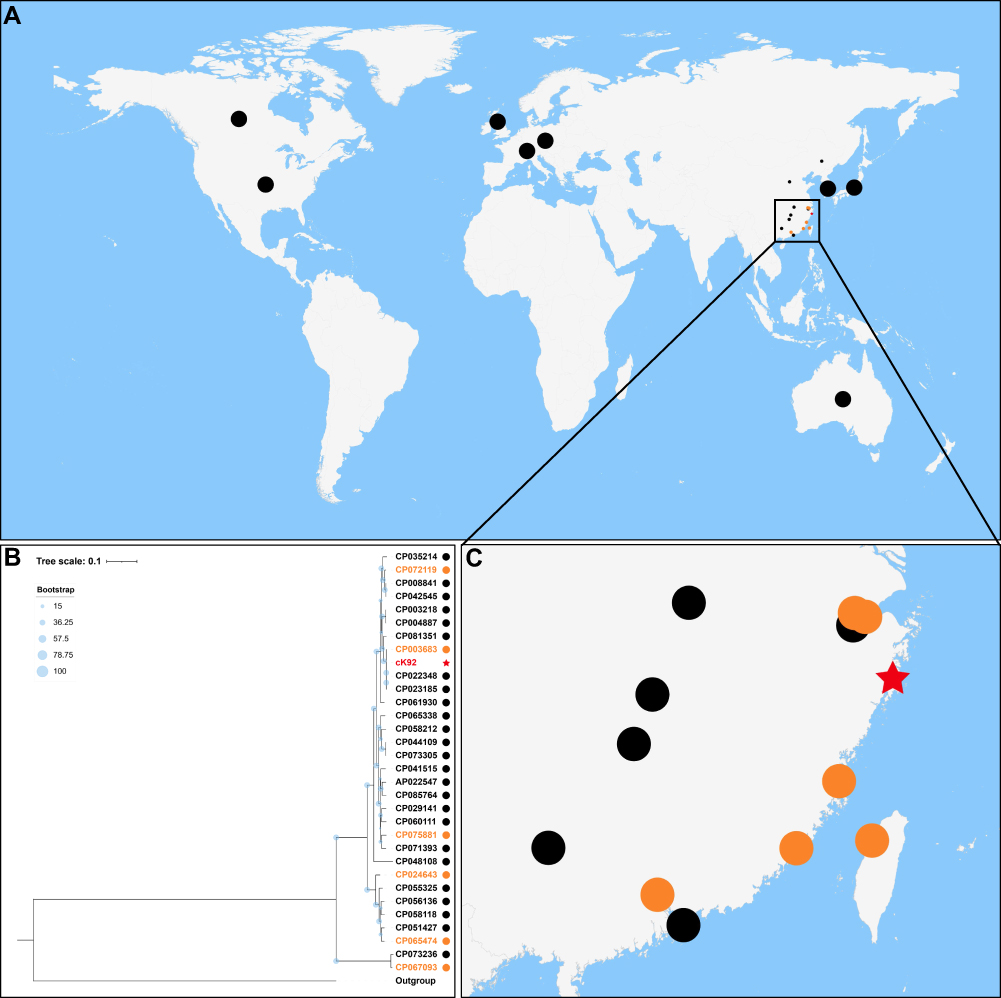 |
Figure 1 Population distribution and geographic distribution of all known Klebsiella michiganensis strains with complete genomes. (A) Geographic locations of 32 K. michiganensis strains in the world. The dots in the figure are used to mark the geographic distribution of strains. If the number of strains distributed in countries except China was greater than one, one dot is used to represent. Strains located in China are represented by smaller dots due to their dense distribution. The black dots represent the K. michiganensis strains that did not carry IncHI5 plasmids, the orange dots represent the K. michiganensis strains carrying IncHI5 plasmids, and the red star represents the K. michiganensis strain carrying pK92-qnrS from this study. (B) Maximum-likelihood tree. The degree of support (percentage) for each cluster of associated taxa, as determined by bootstrap analysis, is shown with blue dots next to each branch. The bar corresponds to the scale of sequence divergence. The meaning of the symbols corresponds to that in (A). (C) Geographic locations of K. michiganensis strains in China. The meanings of the symbols correspondto those in (A). |
Overview of All IncHI5 Plasmids Carried by K. michiganensis
The length of the seven plasmids mentioned above ranged from 274 kb to 397 kb, and the number of ORFs predicted on them ranged from 288 to 416 (Table 1 and Supplementary Figure S1). All the seven plasmids belonged to IncHI5 plasmids, because they all contained the replicon combination repHI5B and repFIB unique to IncHI5 plasmids, and had a similar conserved backbone regions structure.12 Table 1 showed the accession number, host bacterium, total length, total number of ORFs, mean G+C content, length of backbone, mean G+C content of backbone, origin, and other information of these plasmids. With the exception of plasmid pCP024641, the difference in the length of the backbone region of these plasmids was small. The extra part of the backbone region of pCP024641 revealed in the comparative genomics that it may be derived from the integration of part of the backbone regions of the IncC plasmids.34 All these seven IncHI5 plasmids had one or more relatively huge and complex MDR regions. Phylogenetic analysis, accurate annotation and detailed comparison were performed to gain an overall deeper evolution history and structural information of these plasmids.
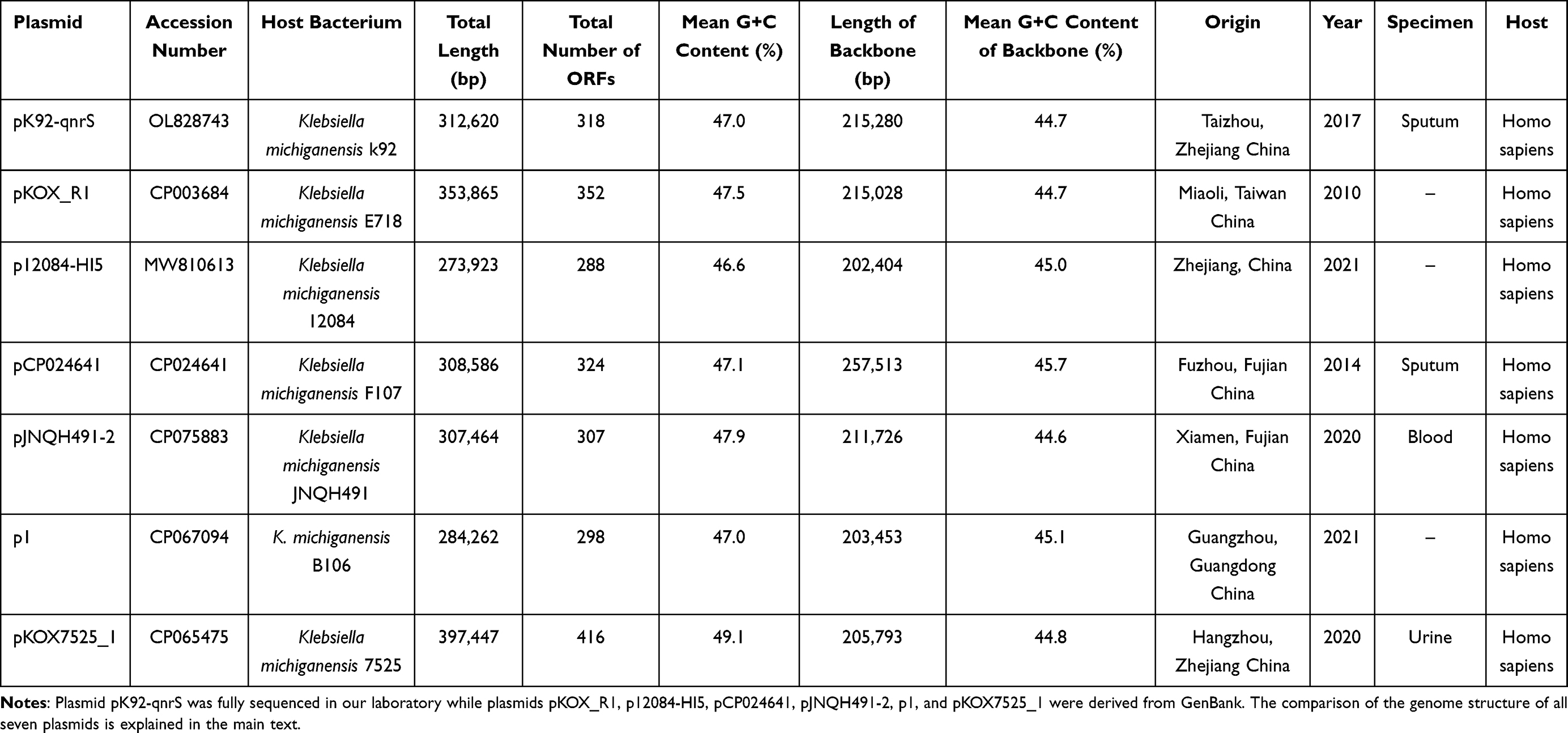 |
Table 1 Major Features of IncHI5 Plasmids Carried by Klebsiella michiganensis Included in This Study |
Comparison of the Seven IncHI5 Plasmids
The genetic structure of all IncHI5 plasmids can be divided into backbone regions and accessory regions. The length of the backbone regions of these seven plasmids varies from 202kb to 257kb (Table 1). Pairwise sequence comparison using BLASTN showed that they all had 99% nucleotide identity and coverage greater than 73% (Supplementary Table S2). A linear comparison of the backbone regions of these seven plasmids revealed the following characteristics (Supplementary Figure S2): (1) The major IncHI5 backbone genes or gene loci such as repHI5B, repFIB, parABC, and tra1/tra2 were conserved. (2) Among the seven plasmids, all except pCP024641 contained two conjugal transfer regions (tra1/tra2). Although the structure of tra2 in different plasmids was not exactly the same, some lost a part of transfer genes, which may hinder their self-transferability. (3) pCP024641 had additional backbone regions compared with other plasmids, including a region of conjugative transfer. Comparing with IncC plasmid pR55,34 pCP024641 showed more than 95% identity, suggesting that this fragment may be derived from IncC plasmid. A total of 705 SNPs were identified from the backbone regions of these seven plasmids, and an ML phylogenetic tree was constructed using these SNPs dataset. Accordingly, these seven plasmids could be assigned into three clades: I (pKOX7525_1 and pJNQH491-2), II (pK92-qnrS and pKOX_R1), and III (p12084-HI5, p1, and pCP024641) (Figure 2).
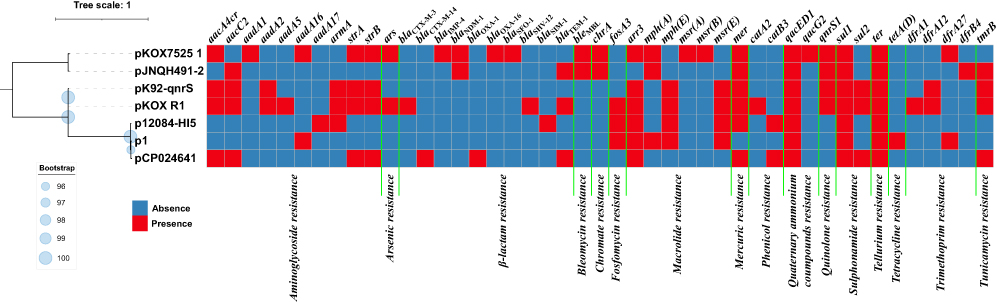 |
Figure 2 Maximum-likelihood tree and heatmap of prevalence of resistance genes in the seven plasmids. Bootstrap is shown with blue dots next to each branch. The bar corresponds to the scale of sequence divergence. |
Large accessory regions were inserted into the different loci of all the seven plasmid backbones, such as insertion sequences (ISs) and transposons. All the seven plasmids except p12084-HI5 had two main longer accessory resistance modules (Supplementary Figure S2). These accessory resistance modules could be divided into two main categories according to their structure and origin: Tn1696-related regions and Tn6535-related regions, and multiple drug-resistant genes were carried on these accessory resistance modules. (Figure 2 and Supplementary Table S3).
Comparison of Tn1696-Related Regions
There were nine Tn1696-related regions in these seven plasmids (Figure 3), including 81.1-kb MDR-1 region and 41.9-kb MDR-2 region from pKOX_R1, Tn7383 from pK92-qnrS, Tn7384 from p12084-HI5, Tn7385 from pCP024641, Tn7386 from pJNQH491-2, Tn7387 and 19.0-kb MDR region from p1, and 141.5-kb MDR-2 region from pKOX7525_1. Due to the similarity of structure, these regions can be regarded as derivatives of Tn1696. Tn1696 was a unit transposon of the Tn3 family,35,36 and its two ends were inverted repeat left (IRL) and inverted repeat right (IRR). The main backbone structure of Tn1696 was tnpA (transposase)–tnpR (resolvase)–res (resolution site)–mer (mercury resistance locus), and the concise class I integron In4 was inserted in the resolution site. Among the above nine Tn1696-related regions, five regions were identified as newly designated unit transposons, namely Tn7383, Tn7384, Tn7385, Tn7386, and Tn7387, because these five regions contained paired terminal 38-bp IRL/IRR on both sides, and all (except Tn7387) contained 5-bp direct repeats (DRs, target site duplication signals for transposition). In the structure of each remaining four Tn1696-related regions, there was no pair of IRL/IRR, so the remaining four regions could not be defined as transposons. Tn7383, 41.9-kb MDR-2 region from pKOX_R1, Tn7384, Tn7385, Tn7386, Tn7387, 19.0-kb MDR region from p1, and 141.5-kb MDR-2 region from pKOX7525_1 contained tnpA–tnpR fragments with high identity to Tn1696. 81.1-kb MDR-1 region from pKOX_R1, Tn7383, Tn7384, Tn7385, Tn7386, and Tn7387 contained the mer locus–IRR fragments that were partially or completely identical to Tn1696.
 |
Figure 3 Nine Tn1696-related regions from the seven plasmids. Genes are denoted by arrows. Genes, mobile genetic elements and other features are colored based on their functional classification. Shading denotes regions of homology (nucleotide identity > 95%). The accession numbers of Tn169635,36 and Tn154837 for reference are U12338 and AF550415, respectively. |
Besides the similarity of Tn1696 backbones, all the nine Tn1696-related regions contained different integrons, transposons, and putative resistance units which were related to resistant genes. Five Tn1696-related regions had different derivatives of Tn1548, which was a composite transposon first reported in 2005 to be identified as a vector of armA, a worldwide disseminated aminoglycoside resistance methylase gene.37 The 81.1-kb MDR region from pKOX_R1 contained a total of 18 resistance genes, which were located at nine different resistance loci: truncated aacC2–tmrB region, truncated IS26–catA2–IS26 unit, ars region, ΔTn1548, In797 [gene cassette array (GCA): aadA5–gcu37–dfrA1], truncated IS26–blaLAP-2–qnrS1–IS26 unit, Tn5393c, IS26–blaSHV-12–IS26 unit, and IS26–fosA3–IS26 unit. The resistance markers from Tn7383 were truncated aacC2–tmrB region and In263 (GCA: aacA4–arr3). The 41.9-kb MDR region from pKOX_R1 included In263, ΔTn6339 (containing blaCTX-M-3 and blaTEM-1B), and ΔTn1548. Tn7384 from p12084-HI5 contained three resistance loci: type A IS26–fosA3–IS26 unit, ∆Tn1548, and In1630 (GCA: aadA17–catB3–arr3–blaSIM-1). Tn7385 from pCP024641 was most similar in structure to Tn1696, except that In4 inserted in Tn1696 was replaced by In37 (GCA: arr3–catB3–blaOXA-1–aacA4). Tn7386 from pJNQH491-2 included truncated IS26–blaLAP-2–qnrS1–IS26 unit, blaNDM-1 and bleMBL containing ΔTn125, In211 (GCA: dfrB4). IS26–fosA3–IS26 unit, IS26–mph(A)–IS6100 unit, and In469 (GCA: arr3–dfrA27–aadA16) were distributed on the 19.0-kb MDR region from p1, and these two units shared the same IS26. 141.5-kb MDR-2 region from pKOX7525_1, a huge and complex Tn1696-related region, contained a total of 17 resistance genes associated with nine different resistance loci: ars region, Tn6399b, In1021, truncated IS26–blaLAP-2–qnrS1–IS26 unit, ΔTn5393c, blaSFO-1 region, ΔTn5053, ΔTn5563, and In1377 (GCA: blaIMP-4–qacG2–aacA4–∆catB3) (Figure 3 and Supplementary Table S3).
Comparison of Tn6535-Related Regions
Tn6535 was a unit transposon of the Tn3 family, and its backbone structure was composed of tnpA–res–tnpR and the ars locus.38 In the accessory regions of these seven plasmids, there were three Tn6535-related regions, which were 60.6-kb MDR region from pK92-qnrS, 34.8-kb MDR-1 region from pJNQH491-2, and 37.0-kb MDR-1 region from pKOX7525_1. Since there were no IRL/IRR pairs at both ends of these three regions, they could not be identified as intact transposons. The partial fragments of these three regions showed high identity to the tnpA–res–tnpR backbone region in Tn6535.
The 60.6-kb MDR region from pK92-qnrS had a lot of accessory resistant modules: ΔTn5393c, truncated IS26–blaLAP-2–qnrS1–IS26 unit, In263 (GCA: aacA4–arr3), ΔTn2 (containing blaTEM-1B), and ∆Tn1548. Moreover, the 60.6-kb MDR region from pK92-qnrS had six copies of IS26, which indicated IS26 played a crucial role in the formation of MDR region through homologous recombination.39 Both of 34.8-kb MDR-1 region from pJNQH491-2 and 37.0-kb MDR-1 region from pKOX7525_1 contained chrA–orf98 unit and IS26–mph(A)–IS6100 unit. Besides, there were also msrAB region (carrying msrAB, confer resistance to erythromycin and streptogramin B) and ΔTn5393c (carrying strA and strB) distributed on the 37.0-kb MDR-1 region from pKOX7525_1. In2132 (GCA: aacA4–arr3), aacC2–tmrB region, and ΔTn21 were existed in 34.8-kb MDR-1 region from pJNQH491-2 (Figure 4).
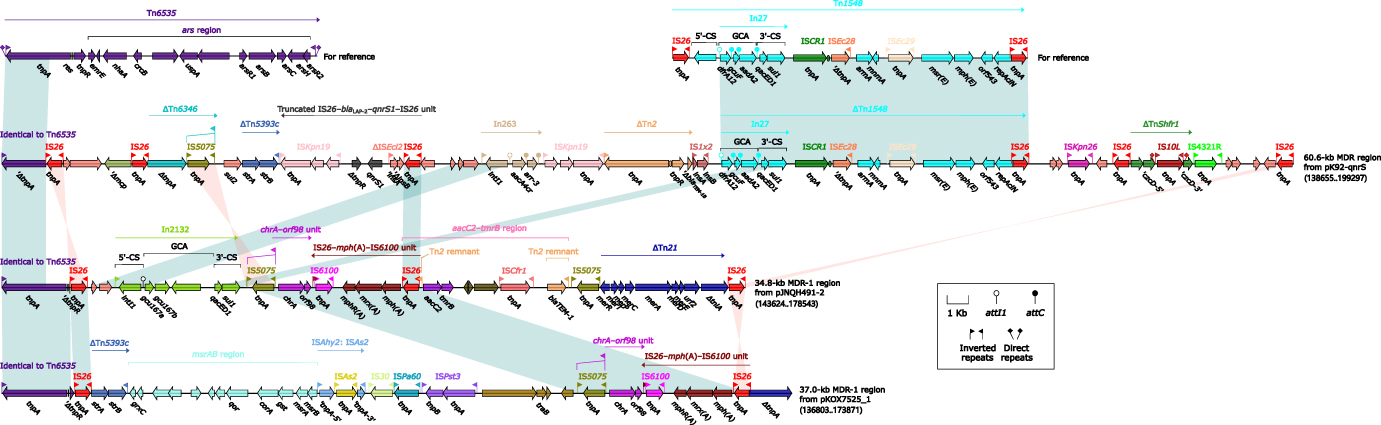 |
Figure 4 Tn6535-related regions from pK92-qnrS, pJNQH491-2, and pKOX7525_1. Genes are denoted by arrows. Genes, mobile genetic elements and other features are colored based on their functional classification. Shading denoted regions of homology (nucleotide identity > 95%). The accession numbers of Tn653538 and Tn154837 for reference are CP009706 and AF550415, respectively. |
18.4-Kb MDR Region from pCP024641
In addition, 18.4-kb MDR region from pCP024641 was different from other MDR regions and could not be divided into Tn1696-related regions or Tn6535-related regions according to the structure (Figure 5). It carried drug resistance genes such as strAB and blaTEM-1, locating in ΔTn5393c and aacC2–tmrB region.
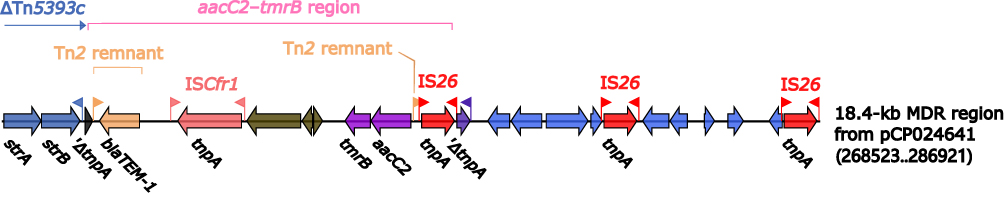 |
Figure 5 18.4-kb MDR region from pCP024641. Genes are denoted by arrows. Genes, mobile genetic elements and other features are colored based on their functional classification. |
Discussion
Since first being discovered in 2012, K. michiganensis has caused several serious and difficult-to-treat nosocomial infections.2–4,6–8 IncHI5 plasmids, a subgroup of IncHI, usually carry a variety of MGEs and drug resistance genes according to previous reports.9,12,40,41 The K. michiganensis carrying IncHI5 plasmids were identified frequently in the last two years.16,17 The similarities and differences of the genome structure of IncHI5 plasmids and the population and geographic distribution within the K. michiganensis species have urgently needed to be studied.
In this work, seven IncHI5 plasmids carried by K. michiganensis strains were accurately annotated and compared in detail, and their evolutionary history was analyzed (Table 1). Among the seven K. michiganensis strains carrying above-mentioned IncHI5 plasmids (Supplementary Table S1), strain E718 containing pKOX_R1 was the first reported case,14 and pKOX_R1 was reported as a multidrug resistant plasmid carrying three β-lactamase genes (blaCTX-M-3, blaTEM-1, and blaSHV-12).42 Strain E718 was obtained from urine and drainage from a pelvis cystic lesion of a patient in November 2010, and the original classification classified E718 as K. oxytoca, although it has now been reclassified as K. michiganensis. After that, IncHI5 plasmids and K. michiganensis have been reported worldwide, but there were no reports of their co-occurring in nearly eight years. Then, starting from 2020, K. michiganensis carrying IncHI5 plasmids have been reported several times.16,17 K. michiganensis carries IncHI5 plasmids at a low frequency, probably because some K. michiganensis was misclassified as K. oxytoca and thus not counted. Perhaps it is also because K. michiganensis have only recently attracted attention, and the rapid development of sequencing has also led to an increase in related sequencing samples in recent years.
There were five complete sequences of IncHI5 plasmids in K. michiganensis uploaded to NCBI since 2020 (p12084-HI5, pCP024641, pJNQH491-2,16 p1, and pKOX7525_117). Among them pCP024641 was the first reported case, which was collected from Fujian Provincial Hospital, Fuzhou Fujian China. From a geographic view, the distribution of pCP024641 and pKOX_R1 is relatively close; from the perspective of a phylogenetic tree, branches of pCP024641 and pKOX_R1 are also close, which may indicate a transmission event from Taiwan to Fujian. Later, IncHI5 plasmids coexisting with K. michiganensis were reported in Guangzhou, Hangzhou17 and other places,16 and all these locations are geographically located in the southeast coastal area of China. This distinctive geographic distribution may imply that the acquisition of IncHI5 plasmids in K. michiganensis is related to some unique geographic-related conditions, although the relevant internal mechanisms are still unclear. Phylogenetic analysis showed that K. michiganensis strains carrying IncHI5 plasmids are located in different branches of the phylogenetic tree (Figure 1), and their distribution on the tree was relatively scattered. This may indicate that the prevalence of IncHI5 plasmids carried by K. michiganensis are not limited to a specific branch, but have spread in different clonal groups. In summary, K. michiganensis strains carrying IncHI5 plasmids have appeared densely since 2020, and have a potential trend of outbreak; at the same time, multiple clonal groups of K. michiganensis carrying IncHI5 plasmids have spread in the southeast coastal area of China. Therefore, cases of IncHI5 plasmids occurring in K. michiganensis should be focused on and studied, and the attention should be paid to their transmission mechanism.
These seven IncHI5 plasmids shared a similar structure with high nucleotide identity and coverage (Supplementary Table S2), and two main large accessory resistance modules (Tn1696-related regions and Tn6535-related regions) were contained in all these plasmids except p12084-HI5. In the nine Tn1696-related regions (Figure 3), 81.1-kb MDR-1 region from pKOX_R1, Tn7383, Tn7384, Tn7385, Tn7386, and Tn7387 contained the mer locus–IRR fragments homologous to Tn1696. mer locus is a series of operons with mercury resistance.43 The upstream of these mer locus were all immediately adjacent to ISs such as IS26 and IS6100, indicating that the reason why these mer locus had different lengths was that mer locus were interrupted by the insertion of ISs. Tn7383, 41.9-kb MDR-2 region from pKOX_R1, Tn7384, Tn7385, Tn7386, Tn7387, 19.0-kb MDR region from p1, and 141.5-kb MDR-2 region from pKOX7525_1 contained the tnpA–tnpR fragments that were almost identical to Tn1696. In Tn1696, the tnpA–tnpR fragment is followed by concise class 1 integron In4, and after the tnpA–tnpR fragments of these Tn1696 derivatives, In4 was replaced by In263 (in Tn7383 and 41.9-kb MDR-2 region from pKOX_R1), In1630 (in Tn7384), In37 (in Tn7385), In211 (in Tn7386), In1706 (in Tn7387), In469 (in 19.0-kb MDR region from p1), and In1377 (in 141.5-kb MDR-2 from pKOX7525_1), respectively. This suggested integrations and replacements of integrons occurred downstream of tnpA–tnpR fragments in the Tn1696-related regions. The mer locus and tnpA–tnpR fragments were located at the two sides of the Tn1696-related regions (the junction with backbone regions). Although the insertion of ISs and the integration of integrons occurred upstream and downstream respectively, main structure of mer locus and tnpA–tnpR fragments was partially or completely retained, which indicated the homology of these Tn1696-related regions with Tn1696. In the three Tn6535-related regions (Figure 4), each of their internal upstream contained the part of the tnpA–res–tnpR backbone region of Tn6535, and they constituted the upstream boundary of each Tn6535-related regions respectively. All these three Tn6535-related regions did not have the complete tnpA–res–tnpR backbone region of Tn6535. The tnpA–res–tnpR regions were truncated due to the insertion of IS26 in the downstream, which suggested that these Tn6535-related regions were formed by Tn6535 through genetic recombination.
The evolutionary history of these large accessory regions can be inferred from their internal structure and the distribution of MGEs on it. The formation of these large accessory regions is mostly due to the insertion, transposition, and integration of MGEs. Some are the overall activities of several adjacent MGEs, and some are the individual behaviors of a single MGE such as the insertion of a single IS. In 34.8-kb MDR-1 region from pJNQH491-2 and 37.0-kb MDR-1 region from pKOX7525_1 (Figure 4), chrA–orf98 unit and IS26–mph (A)–IS6100 unit seemed to form an overall putative resistance unit, which was bounded by IS5075 and IS26. This unit was likely to mobilize as a whole element and cannot be classified as a transposon. However, not all modules in the immediate vicinity can be regarded as a single component. aacC2–tmrB region and ISKpn19 were closely adjacent to each other in both 81.1-kb MDR-1 region from pKOX_R1 and Tn7383 (Figure 3), but they cannot be regarded as a single MGE. This is because from their downstream structure, their downstream difference was only an insertion of IS10L. Therefore, their formation was more likely to be caused by the insertion of this extra single IS10L. The evolutionary history caused by the insertions of ISs also existed in Tn7384: ISKpn21 and ISKml1 were inserted upstream of orf543 and armA in ΔTn1548, respectively, forming a new structure of accessory region. This view was also verified by comparison with the reference Tn1548.
There were a variety of resistance-related MGEs distributed on these plasmids, and various types of antibiotic resistance genes were discovered in these seven plasmids: a total of 14 drug-resistant genes phenotypes with 45 resistance markers, including β-lactam resistance, macrolide resistance, quinolone resistance, and so on (Figure 2 and Supplementary Table S3). Among them, the resistance genes for β-lactam resistance were blaCTX-M-3, blaCTX-M-14, blaIMP-4, blaNDM-1, blaOXA-1, blaOXA-16, blaSFO-1, blaSIM-1, and blaTEM-1. Except for pK92-qnrS and p1, the other five plasmids all contained one or more β-lactam resistance genes, suggesting that the IncHI5 plasmids in K. michiganensis were good vectors for antibiotic resistance genes.
Conclusions
In conclusion, IncHI5 plasmids have spread in different clonal groups of K. michiganensis strains and have recently appeared intensively, suggesting that they have a potential for outbreak. Up to now, K. michiganensis strains carrying IncHI5 plasmids have only appeared in the southeast coastal area of China, and their internal mechanism is urgently needed to be studied. The large accessory regions in the IncHI5 plasmids can be divided into two types: Tn6535-related regions and Tn1696-related regions. Their formation was related to the integration and insertion of MGEs, and the accumulation and expression of drug resistance genes carried on MGEs accelerated the spread of antimicrobial resistance in K. michiganensis strains. We expect that the data in this work can further decipher the structural diversification, population distribution, and evolutionary history of IncHI5 plasmids in K. michiganensis.
Data Sharing Statement
The data presented in this study are available on request from the corresponding author. The plasmid sequences analyzed in this study can be found in public NCBI GenBank database. The accession numbers were provided in this article when these plasmids were firstly indicated.
Ethics Approval
The specimens were obtained with the patient’s consent. The use of human specimens and all related experimental protocols was reviewed and approved by the Ethics Committee of Taizhou Municipal Hospital affiliated with Taizhou University, Zhejiang, China, in accordance with the medical research regulations of the Ministry of Health, China. Research and all related procedures involving biohazardous materials were approved by the Biosafety Committee of Taizhou Municipal Hospital affiliated with Taizhou University. This research was conducted in China.
Acknowledgments
This work was supported by the Foundation of Zhejiang Health Department (2022KY1395), and the Foundation of Taizhou Science and Technology Bureau (1902ky47).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Saha R, Farrance CE, Verghese B, Hong S, Donofrio RS. Klebsiella michiganensis sp. nov., a new bacterium isolated from a tooth brush holder. Curr Microbiol. 2013;66(1):72–78. doi:10.1007/s00284-012-0245-x
2. Seiffert SN, Wuthrich D, Gerth Y, Egli A, Kohler P, Nolte O. First clinical case of KPC-3-producing Klebsiella michiganensis in Europe. New Microbes New Infect. 2019;29:100516. doi:10.1016/j.nmni.2019.100516
3. Zheng B, Xu H, Yu X, et al. Identification and genomic characterization of a KPC-2-, NDM-1- and NDM-5-producing Klebsiella michiganensis isolate. J Antimicrob Chemother. 2018;73(2):536–538. doi:10.1093/jac/dkx415
4. Abed JY, Deraspe M, Berube E, et al. Complete genome sequences of Klebsiella michiganensis and citrobacter farmeri, KPC-2-producers serially isolated from a single patient. Antibiotics. 2021;10(11):11. doi:10.3390/antibiotics10111408
5. Ccorahua-Santo R, Cervantes M, Duran Y, Aguirre M, Marin C, Ramirez P. Draft genome sequence of Klebsiella michiganensis 3T412C, harboring an arsenic resistance genomic Island, isolated from mine Tailings in Peru. Genome Announc. 2017;5:28. doi:10.1128/genomeA.00611-17
6. King TL, Schmidt S, Thakur S, et al. Resistome of a carbapenemase-producing novel ST232 Klebsiella michiganensis isolate from urban hospital effluent in South Africa. J Glob Antimicrob Resist. 2021;24:321–324. doi:10.1016/j.jgar.2021.01.004
7. Kang M, Chmara J, Duceppe MO, Phipps-Todd B, Huang H. Complete genome sequence of a Canadian Klebsiella michiganensis strain, obtained using Oxford Nanopore Technologies Sequencing. Microbiol Resour Announc. 2020;9:46. doi:10.1128/MRA.00960-20
8. Campos-Madueno EI, Sigrist T, Fluckiger UM, Risch L, Bodmer T, Endimiani A. First report of a blaVIM-1 metallo-beta-lactamase-possessing Klebsiella michiganensis. J Glob Antimicrob Resist. 2021;25:310–314. doi:10.1016/j.jgar.2021.03.027
9. Liang Q, Jiang X, Hu L, et al. Sequencing and genomic diversity analysis of IncHI5 plasmids. Front Microbiol. 2018;9:3318. doi:10.3389/fmicb.2018.03318
10. Maher D, Taylor DE. Host range and transfer efficiency of incompatibility group HI plasmids. Can J Microbiol. 1993;39(6):581–587. doi:10.1139/m93-084
11. Cain AK, Hall RM. Evolution of IncHI2 plasmids via acquisition of transposons carrying antibiotic resistance determinants. J Antimicrob Chemother. 2012;67(5):1121–1127. doi:10.1093/jac/dks004
12. Liang Q, Yin Z, Zhao Y, et al. Sequencing and comparative genomics analysis of the IncHI2 plasmids pT5282-mphA and p112298-catA and the IncHI5 plasmid pYNKP001-dfrA. Int J Antimicrob Agents. 2017;49(6):709–718. doi:10.1016/j.ijantimicag.2017.01.021
13. Yang J, Long H, Hu Y, Feng Y, McNally A, Zong Z. Klebsiella oxytoca complex: update on taxonomy, antimicrobial resistance, and virulence. Clin Microbiol Rev. 2021;35:e0000621.
14. Huang TW, Wang JT, Lauderdale TL, et al. Complete sequences of two plasmids in a blaNDM-1-positive Klebsiella oxytoca isolate from Taiwan. Antimicrob Agents Chemother. 2013;57(8):4072–4076. doi:10.1128/AAC.02266-12
15. Sayers EW, Cavanaugh M, Clark K, et al. GenBank. Nucleic Acids Res. 2021;49(D1):D92–D96. doi:10.1093/nar/gkaa1023
16. Wang Y, Zhu B, Liu M, et al. Characterization of IncHI1B plasmids encoding efflux pump TmexCD2-ToprJ2 in carbapenem-resistant Klebsiella variicola, Klebsiella quasipneumoniae, and Klebsiella michiganensis strains. Front Microbiol. 2021;12:759208. doi:10.3389/fmicb.2021.759208
17. Li X, He J, Jiang Y, Peng M, Yu Y, Fu Y. Genetic characterization and passage instability of a hybrid plasmid co-harboring blaIMP-4 and blaNDM-1 reveal the contribution of insertion sequences during plasmid formation and evolution. Microbiol Spectr. 2021;9(3):e0157721. doi:10.1128/Spectrum.01577-21
18. Richter M, Rossello-Mora R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci U S A. 2009;106(45):19126–19131. doi:10.1073/pnas.0906412106
19. Lee I, Ouk Kim Y, Park SC, Chun J. OrthoANI:An improved algorithm and software for calculating average nucleotide identity. Int J Syst Evol Microbiol. 2016;66(2):1100–1103. doi:10.1099/ijsem.0.000760
20. Delcher AL, Salzberg SL, Phillippy AM. Using MUMmer to identify similar regions in large sequence sets. Curr Protoc Bioinformatics. 2003;1:10–13.
21. Letunic I, Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49(W1):W293–W296. doi:10.1093/nar/gkab301
22. Brettin T, Davis JJ, Disz T, et al. RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci Rep. 2015;5(1):8365. doi:10.1038/srep08365
23. Boratyn GM, Camacho C, Cooper PS, et al. BLAST: a more efficient report with usability improvements. Nucleic Acids Res. 2013;41(W1):W29–W33. doi:10.1093/nar/gkt282
24. Boutet E, Lieberherr D, Tognolli M, Schneider M, Bairoch A. UniProtKB/Swiss-Prot. Methods Mol Biol. 2007;406:89–112. doi:10.1007/978-1-59745-535-0_4
25. O’Leary NA, Wright MW, Brister JR, et al. Reference sequence (RefSeq) database at NCBI: current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016;44(D1):D733–D745. doi:10.1093/nar/gkv1189
26. Alcock BP, Raphenya AR, Lau TTY, et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020;48(D1):D517–D525. doi:10.1093/nar/gkz935
27. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
28. Siguier P, Perochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34:D32–D36. doi:10.1093/nar/gkj014
29. Moura A, Soares M, Pereira C, Leitao N, Henriques I, Correia A. INTEGRALL: a database and search engine for integrons, integrases and gene cassettes. Bioinformatics. 2009;25(8):1096–1098. doi:10.1093/bioinformatics/btp105
30. Roberts AP, Chandler M, Courvalin P, et al. Revised nomenclature for transposable genetic elements. Plasmid. 2008;60(3):167–173. doi:10.1016/j.plasmid.2008.08.001
31. Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792–1797. doi:10.1093/nar/gkh340
32. Liu P, Li P, Jiang X, et al. Complete genome sequence of Klebsiella pneumoniae subsp. pneumoniae HS11286, a multidrug-resistant strain isolated from human sputum. J Bacteriol. 2012;194(7):1841–1842. doi:10.1128/JB.00043-12
33. Founou RC, Founou LL, Allam M, Ismail A, Essack SY. Genomic characterisation of Klebsiella michiganensis co-producing OXA-181 and NDM-1 carbapenemases isolated from a cancer patient in uMgungundlovu District, KwaZulu-Natal Province, South Africa. S Afr Med J. 2018;109(1):7–8. doi:10.7196/SAMJ.2018.v109i1.13696
34. Doublet B, Boyd D, Douard G, Praud K, Cloeckaert A, Mulvey MR. Complete nucleotide sequence of the multidrug resistance IncA/C plasmid pR55 from Klebsiella pneumoniae isolated in 1969. J Antimicrob Chemother. 2012;67(10):2354–2360. doi:10.1093/jac/dks251
35. Partridge SR, Kwong SM, Firth N, Jensen SO. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol Rev. 2018;31(4):4. doi:10.1128/CMR.00088-17
36. Partridge SR, Brown HJ, Stokes HW, Hall RM. Transposons Tn1696 and Tn21 and their integrons In4 and In2 have independent origins. Antimicrob Agents Chemother. 2001;45(4):1263–1270. doi:10.1128/AAC.45.4.1263-1270.2001
37. Galimand M, Sabtcheva S, Courvalin P, Lambert T. Worldwide disseminated armA aminoglycoside resistance methylase gene is borne by composite transposon Tn1548. Antimicrob Agents Chemother. 2005;49(7):2949–2953. doi:10.1128/AAC.49.7.2949-2953.2005
38. Tan JY, Yin WF, Chan KG. Gene clusters of Hafnia alvei strain FB1 important in survival and pathogenesis: a draft genome perspective. Gut Pathog. 2014;6:29. doi:10.1186/1757-4749-6-29
39. Harmer CJ, Hall RM. IS26-mediated formation of transposons carrying antibiotic resistance genes. mSphere. 2016;1:2. doi:10.1128/mSphere.00038-16
40. Liu Z, Chen R, Xu P, Wang Z, Li R. Characterization of a blaNDM-1-Bearing IncHI5-like plasmid from Klebsiella pneumoniae of infant origin. Front Cell Infect Microbiol. 2021;11:738053. doi:10.3389/fcimb.2021.738053
41. Zhu Y, Liu W, Schwarz S, et al. Characterization of a blaNDM-1-carrying IncHI5 plasmid from Enterobacter cloacae complex of food-producing animal origin. J Antimicrob Chemother. 2020;75(5):1140–1145. doi:10.1093/jac/dkaa010
42. Lai CC, Lin TL, Tseng SP, et al. Pelvic abscess caused by New Delhi metallo-beta-lactamase-1-producing Klebsiella oxytoca in Taiwan in a patient who underwent renal transplantation in China. Diagn Microbiol Infect Dis. 2011;71(4):474–475. doi:10.1016/j.diagmicrobio.2011.09.004
43. Nascimento AM, Chartone-Souza E. Operon mer: bacterial resistance to mercury and potential for bioremediation of contaminated environments. Genet Mol Res. 2003;2(1):92–101.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.