")
Back to Journals » Infection and Drug Resistance » Volume 14
Emergence of the Coexistence of mcr-1, blaNDM-5, and blaCTX-M-55 in Klebsiella pneumoniae ST485 Clinical Isolates in China
Authors Cao X, Zhong Q, Guo Y, Hang Y, Chen Y, Fang X, Xiao Y, Zhu H , Luo H, Yu F, Hu L
Received 21 March 2021
Accepted for publication 29 July 2021
Published 28 August 2021 Volume 2021:14 Pages 3449—3458
DOI https://doi.org/10.2147/IDR.S311808
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Xingwei Cao,1,* Qiaoshi Zhong,1,* Yinjuan Guo,2,3 Yaping Hang,1 Yanhui Chen,1 Xueyao Fang,1 Yanping Xiao,1 Hongying Zhu,1 Hong Luo,1 Fangyou Yu,2,3 Longhua Hu1
1Jiangxi Provincial Key Laboratory of Medicine, Clinical Laboratory of the Second Affiliated Hospital of Nanchang University, Nanchang, Jiangxi, People’s Republic of China; 2Department of Clinical Laboratory Medicine, Shanghai Pulmonary Hospital, Tongji University School of Medicine, Shanghai, 200082, People’s Republic of China; 3Shanghai Key Laboratory of Tuberculosis, Shanghai Pulmonary Hospital, Tongji University School of Medicine, Shanghai, 200082, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Longhua Hu; Fangyou Yu Email [email protected]; [email protected]
Introduction: Polymyxin resistance caused by the plasmid-mediated mcr-1 gene in gram-negative bacilli poses a huge threat to our health. In recent years, many regions have reported that mcr-1 and β-lactamase genes can coexist in a single strain.
Methods: In this study, 107 nonduplicate Klebsiella pneumoniae (K. pneumoniae) isolates were collected from a tertiary hospital in Jiangxi, China. Antimicrobial susceptibility testing of isolates was performed using gram-negative susceptibility cards on the VITEK system. The minimum inhibitory concentrations (MICs) of polymyxin B was detected using the microdilution broth method. The presence of resistance genes was assessed using polymerase chain reaction (PCR). We subjected isolates to genotyping using pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) and analyzed the transferability of plasmids with filter mating and electroporation. Subsequently, whole-genome sequencing was performed for plasmids.
Results: Of the 107 K. pneumoniae isolates, 15 (14.0%) were resistant to polymyxin B. All polymyxin B-resistant isolates harbored at least one of the extended-spectrum β-lactamase genes tested. Only one isolate simultaneously harbored mcr-1, blaNDM− 5, blaCTX-M-55, and blaSHV-27 genes. MLST results showed that 15 carbapenem-resistant K. pneumoniae isolates belonged to five sequence types (STs). PFGE results displayed nine different PFGE clusters. Conjugation and transformation experiments and sequencing analysis showed that the strain had three plasmids, and mcr-1, blaNDM− 5, and blaCTX-M-55 were located on different plasmids.
Conclusion: The present study demonstrated for the first time the coexistence of mcr-1, blaNDM− 5, and blaCTX-M-55 in a K. pneumoniae ST485 isolate. The three plasmids carrying the mcr-1, blaNDM− 5, and blaCTX-M-55 genes can be transmitted in Enterobacteriaceae strains, which may lead to more severe bacterial resistance.
Keywords: Klebsiella pneumoniae, polymyxin B, mcr-1, blaNDM-5, blaCTX-M-55, plasmid
Introduction
Klebsiella pneumoniae (K. pneumoniae) is an opportunistic pathogen and the leading cause of healthcare-associated infections.1 Multidrug-resistant (MDR) K. pneumoniae isolates are rapidly spreading, thus limiting the choice of antimicrobial agents for empiric treatment of infections caused by these microorganisms; hence, this is a public health challenge.2
Polymyxins are last-resort antibiotics used to treat infections caused by carbapenem-resistant K. pneumoniae (CRKP).3 The two polymyxins currently in clinical use are polymyxin B and colistin (polymyxin E). They have similar antibacterial activity, but their structures differ by only one amino acid.4
The antibacterial effect of polymyxins on gram-negative bacteria is mainly a two-step mechanism comprising initial binding to and permeabilization of the outer membrane, followed by the destruction of cytoplasmic membrane.5 Notably, with the increase in the clinical use of polymyxins, polymyxin resistance has emerged and is rising rapidly. Polymyxin-resistant K. pneumoniae often spread in different hospital wards, making clinical treatment more difficult.6 The previously reported mechanisms of polymyxin resistance are chromosomally mediated and involve the regulation of two-component regulatory systems (eg, pmrAB, phoPQ, and its negative regulator, mgrB, in the case of K. pneumoniae), leading to the modification of lipid A (phosphoethanolamine or 4-amino-4-arabinose) or in rare cases, the complete loss of the lipopolysaccharide.7
Researchers reported the first plasmid-mediated polymyxin resistance mechanism, mcr-1, in Enterobacteriaceae in China. This warranted immediate worldwide attention, and mcr-1 has since been detected in Enterobacteriaceae from animals, food, and healthy people outside of China, including in Europe and the USA.8 Recently, some countries have also reported that mcr-1 and blaNDM-5 genes coexist in Escherichia coli strains,9,10 which is a serious challenge to treatment efforts.
This study assessed the current status of polymyxin resistance in CRKP isolates and investigated the possible coexistence of mcr-1 and β-lactamase genes in K. pneumoniae in Nanchang, China.
Materials and Methods
Isolation and Identification of Bacterial Strains
From January 2018 to June 2019, a total of 107 nonduplicate CRKP isolates were isolated from hospitalized patients in different clinical departments in a tertiary teaching hospital in Nanchang, China. Different specimens were collected, and the K. pneumoniae isolates were identified using a VITEK-2 automated platform (bioMerieux, Marcy l’Etoile, France). E. coli ATCC 25922 was used as a control strain.
Antimicrobial Susceptibility Testing
The susceptibility of the K. pneumoniae clinical isolates to antimicrobials was determined using gram-negative susceptibility cards (AST-GN-16) on the VITEK system (bioMerieux, Marcy l’Etoile, France) following the manufacturer’s instructions; the results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) standards.11 The MICs of polymyxin B for CRKP were further determined using the microdilution broth method according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines.12 A total of 16 antimicrobial agents were tested, including carbapenems (imipenem), β-lactam/β-lactamase inhibitor complexes (amoxicillin/clavulanic acid and piperacillin–tazobactam), monocyclic β-lactam (aztreonam), cephalosporins (cefoxitin, cefepime, cefazolin, and ceftriaxone), aminoglycosides (gentamicin and amikacin), fluoroquinolones (levofloxacin and ciprofloxacin), folate metabolic pathway inhibitors (sulfamethoxazole), tetracyclines (tobramycin and tigecycline), and polymyxin B. E. coli ATCC 25922 was used as a control.
Detection of β-Lactamases and Resistance Genes
The carbapenemases produced by CRKP isolates were determined using a modified carbapenem inactivation test (mCIM) recommended by CLSI.11 In addition, a double-disc synergy test was performed to confirm the presence of metallo-β-lactamases (MBLs).11 The carbapenemase (blaKPC, blaGES, blaNDM, blaIMP, blaVIM, blaOXA-48, blaSIM, blaSPM, blaSME, and blaGIM), extended-spectrum β-lactamase (ESBLs; blaTEM, blaDHA, blaSHV, blaCMY-II, and blaCTX-M), and polymyxin B (mcr-1 to mcr-8) resistance genes were detected using polymerase chain reaction (PCR) and DNA sequencing as described previously.2,13
Multilocus Sequence Typing
Multilocus sequence typing (MLST) was performed on the polymyxin B-resistant K. pneumoniae isolates by amplifying and sequencing seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) according to a previously described protocol. Sequence types (STs) were assigned using the online database.
Pulsed-Field Gel Electrophoresis
Pulsed-field gel electrophoresis (PFGE) was performed to analyze the phylogenetic relatedness of the polymyxin B-resistant K. pneumoniae isolates. Genomic DNA was digested by XbaI for 4 h at 37 °C. Electrophoresis was performed for 19 h at 14 °C, at an angle of 120°, with switch times of 4 and 40 s at 6 V/cm using the CHEF III system (Bio-Rad Laboratories, Hercules, CA, USA). The Salmonella H9812 strain was used as the size marker. Analysis of the PFGE patterns using the Dice similarity coefficient was performed using the Bionumerics software (Applied Maths, Sint-Martens-Latem, Belgium). Clusters were defined as DNA patterns sharing more than 80% similarity.
Conjugation and Thermal Transfer Experiment
A donor isolate, N816, was cultured in lysogeny broth (LB), and an azide-resistant E. coli J53 strain was used as the recipient. Transconjugants were selected on LB agar plates with 2 mg/L of polymyxin B or imipenem and 150 mg/L of sodium azide. Multiple attempts to transfer blaNDM-5 plasmid failed. Plasmid DNA was extracted from N816, transferred to competent E. coli DH5α, and screened on LB agar plates with 2 mg/L imipenem. After the experiment, the transconjugant (JN816) and transformant (ZN816) were obtained and verified using PCR with previously described primers. Antimicrobial susceptibility testing was subsequently performed on JN816 and ZN816.
Genomic DNA Extraction and Analysis
Genomic DNA was extracted from JN816 and ZN816 with the Qiagen Midi kit (Qiagen, Hilden, Germany) and sequenced with an Illumina HiSeq 2000 sequencer following a paired-end 2×100-bp protocol.14 The raw data were mapped to a reference sequence found on the CLC genomics workbench version 8.0. Sequence comparison and alignment were performed using MEGA 5.01.15
Results
Antimicrobial Susceptibility
Among the 107 K. pneumoniae isolates, 15 (14.0%) were resistant to polymyxin B according to EUCAST 7.0 guidelines.12 The antimicrobial resistance rates of these isolates are shown in Table 1. These isolates were resistant to imipenem, aztreonam, cefazolin, cefepime, cefoxitin, ceftriaxone, ciprofloxacin, and sulfamethoxazole. The resistance rates of isolates for amikacin, gentamicin, tobramycin, and tigecycline were 46.7, 60.0, 53.3, and 13.3%, respectively.
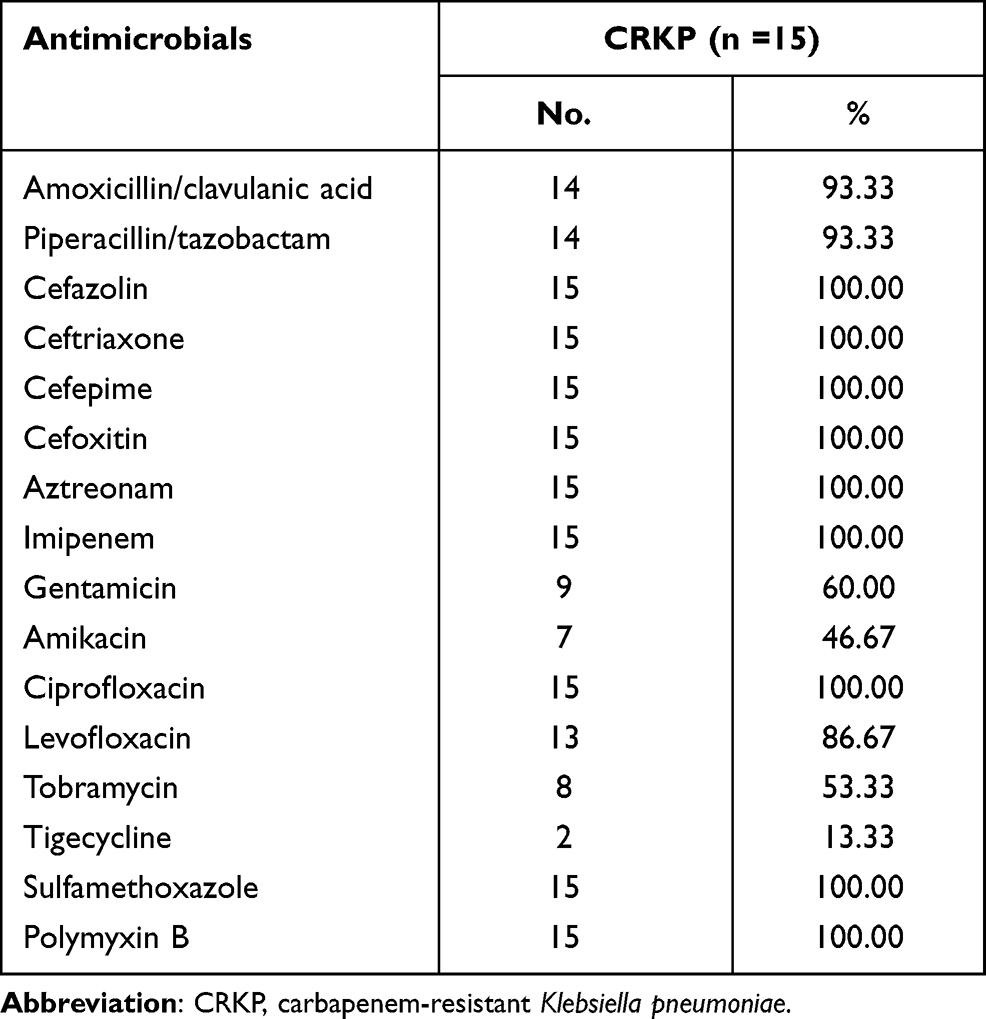 |
Table 1 Antimicrobial Resistance Profiling of 15 Carbapenem-Resistant Klebsiella pneumoniae Isolates |
Detection of Carbapenemase and Other Resistance Genes
Twelve of the 15 polymyxin B-resistant isolates were confirmed as carbapenemase producers as determined using the mCIM assay, among which two isolates had a positive result for the double-disc synergy test, indicating that they also produced an MBL. In addition, 10 CRKP isolates were positive for blaKPC-2, and two were positive for blaNDM. Other carbapenemase genes including blaGES, blaIMP, blaVIM, blaOXA-48, blaSIM, blaSPM, blaSME, and blaGIM were not detected in any of the tested isolates. In addition to blaKPC-2, all isolates were positive for blaSHV, and eight (53.3%) were positive for the ESBL gene, blaCTX-M-65. Only one CRKP isolate was positive for mcr-1, blaNDM-5, blaCTX-M-55, and blaSHV-27 (Figure 1).
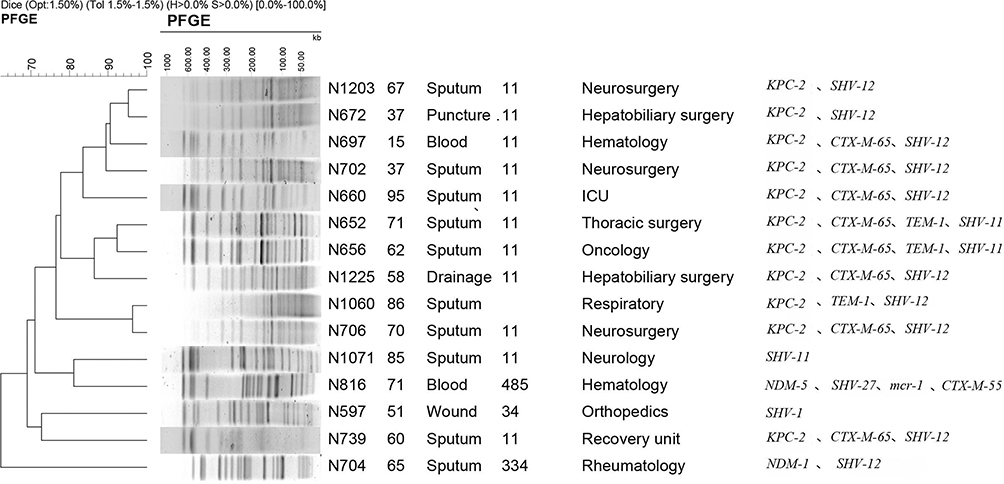 |
Figure 1 Pulsed-field gel electrophoresis results for 15 carbapenem-resistant Klebsiella pneumoniae isolates. |
Molecular Characteristics of CRKP Isolates
Among the 15 CRKP isolates, five STs were identified, including ST11 (11 isolates), and one isolate each in ST34, ST334, ST485, and a novel ST. The PFGE results showed that the 15 isolates were divided into nine different PFGE clusters; cluster A (4; 26.7%), cluster C (3; 20.0%), and cluster D (2; 13.3%). Each of the remaining six isolates were classified as singletons (Figure 1).
Location of mcr-1, blaCTX-M-55, and blaNDM-5 Genes
The two CRKP N816 plasmids harboring mcr-1 and blaCTX-M-55, designated as pMCR-1-N816 and pCTX-M-55-N816, respectively, were successfully transferred into the recipient strain (J53) via filter mating conjugation. We confirmed the presence of mcr-1 and blaCTX-M-55 genes in these plasmids. The antimicrobial resistance patterns of CRKP N816 and its transconjugant are shown in Table 2. The blaNDM-5-harboring plasmid of CRKP N816, designated as pNDM-5-N816, was electroporated into E. coli DH5α as described previously. Growth was observed only on plates with imipenem 2 mg/L, and the transformants were screened for the presence of blaNDM-5 using PCR, and blaNDM-5 was located on the plasmid. The antimicrobial resistance patterns of CRKP N816 and its transformants are shown in Table 2.
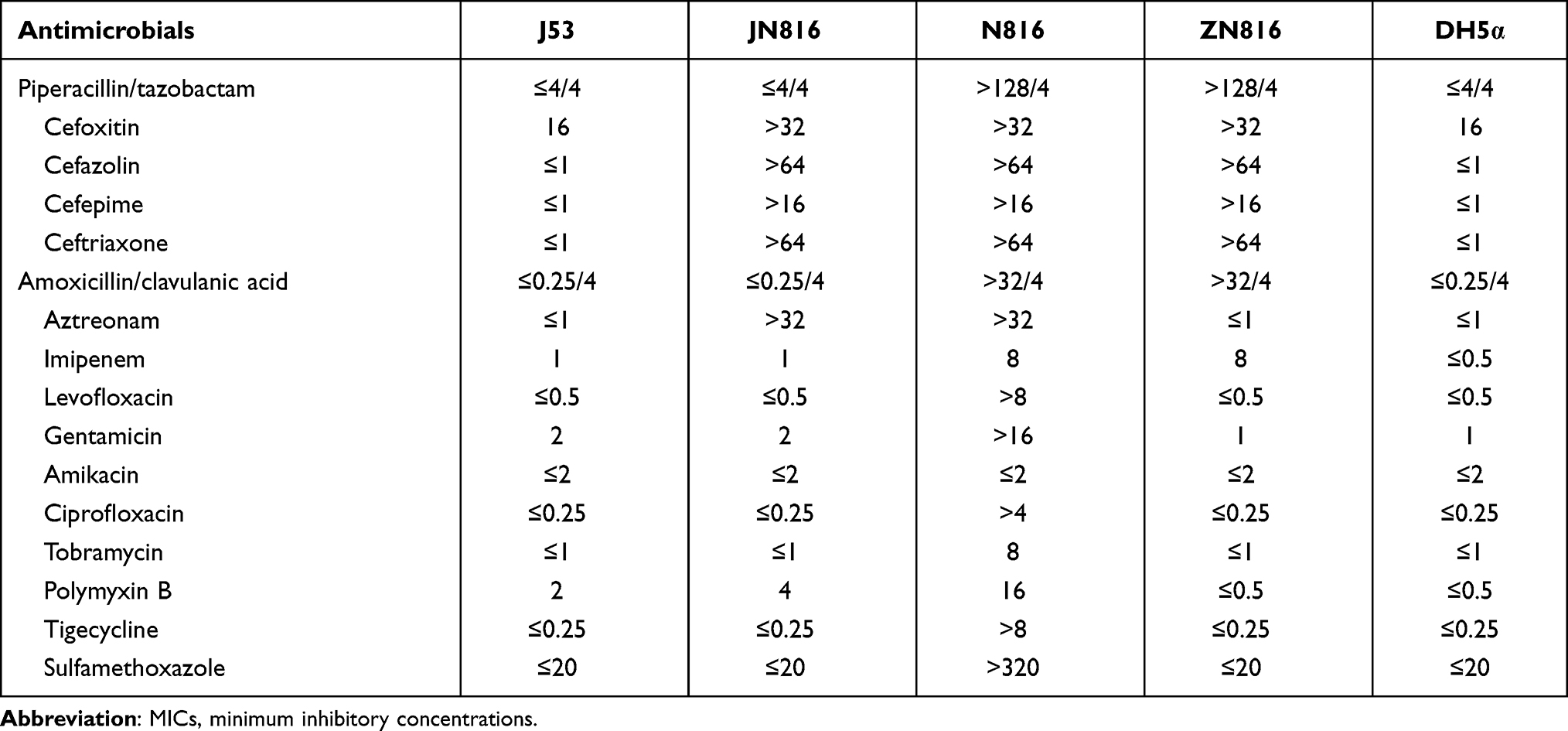 |
Table 2 Minimum Inhibitory Concentrations of Antimicrobials Against N816, JN816, ZN816, J53, and DH5α |
Complete Sequence of Plasmid
pMCR-1-N816 is 33309 base pairs (bp) long, with an average guanine-cytosine (GC) content of 41.84%. It has 41 predicted open reading frames (ORFs) and belongs to the IncX4 incompatibility group. A BLAST search of the plasmid sequences against the GenBank database showed that pMCR-1-N816 is similar (with 100% query coverage and >98.0% nucleotide identity) to pKP15450-MCR-1, an IncX4-type plasmid carrying mcr-1 among K. pneumoniae isolates in China (Figure 2). Plasmid pCTX-M-55-N816 is 76526-bp in length, with an average GC content of 51.93%. It has 105 predicted ORFs, and a BLAST search of the plasmid sequences against the GenBank database showed that pCTX-M-55-N816 is similar to pKP32558-4, with 89% query coverage and >98.0% nucleotide identity (Figure 3). Furthermore, pNDM-5-N816 is 46286-bp in length, with an average GC content of 46.63%, 59 predicted ORFs, and belongs to the IncX3 incompatibility group. A BLAST search showed that pNDM-5-N816 is similar to pNDM5-LDR, an IncX3-type plasmid carrying blaNDM-5 among K. pneumoniae isolates in China, with 100% query coverage and >99.9% nucleotide identity (Figure 4).
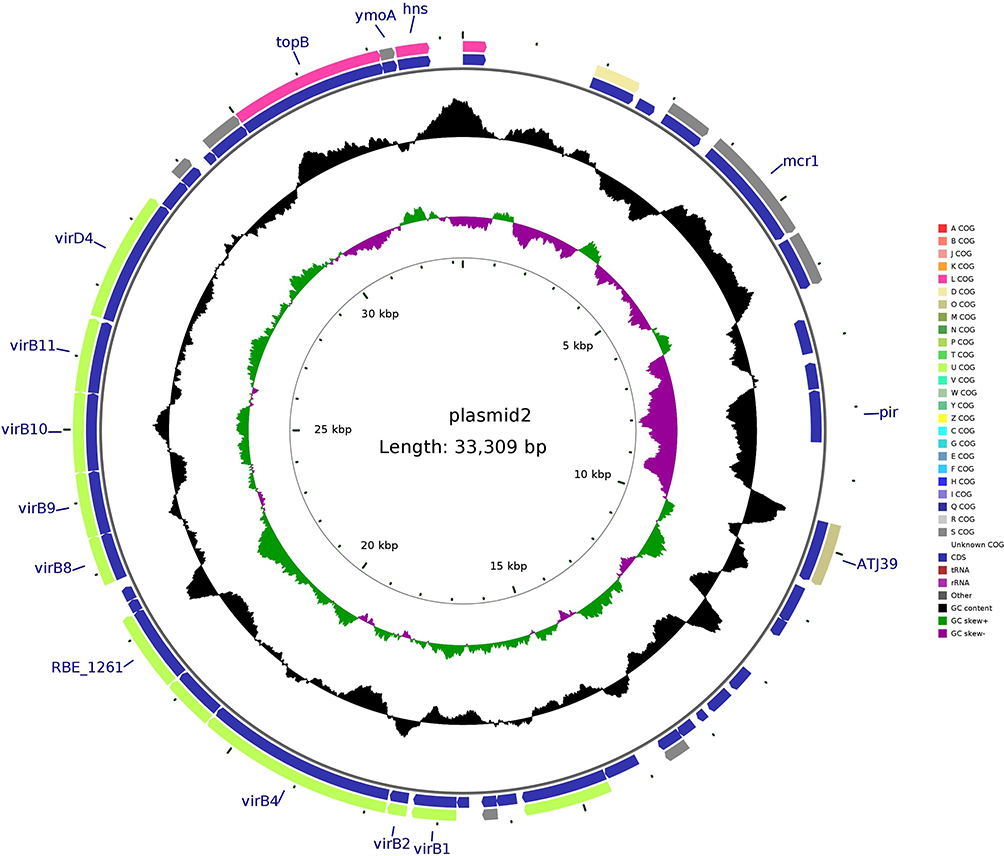 |
Figure 2 Structure of plasmid pMCR-1-N816 carrying mcr-1 from Klebsiella pneumoniae N816. |
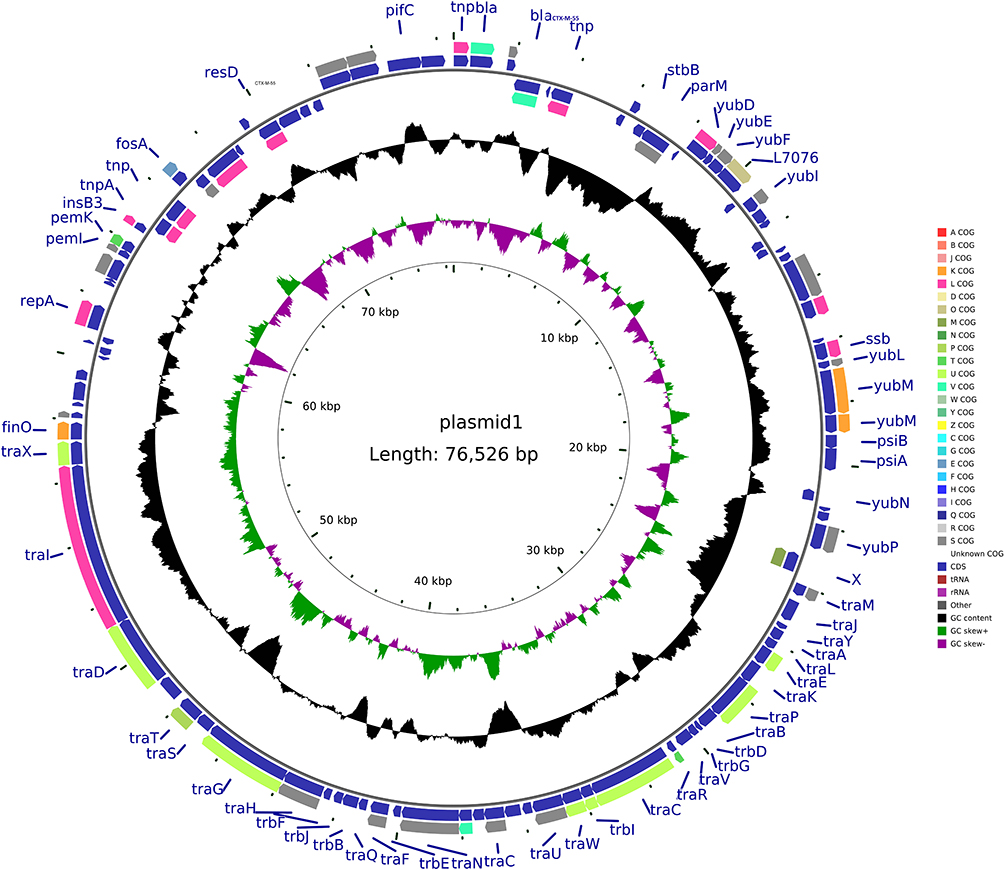 |
Figure 3 Structure of plasmid pCTX-M-55-N816 carrying blaCTX-M-55 from Klebsiella pneumoniae N816. |
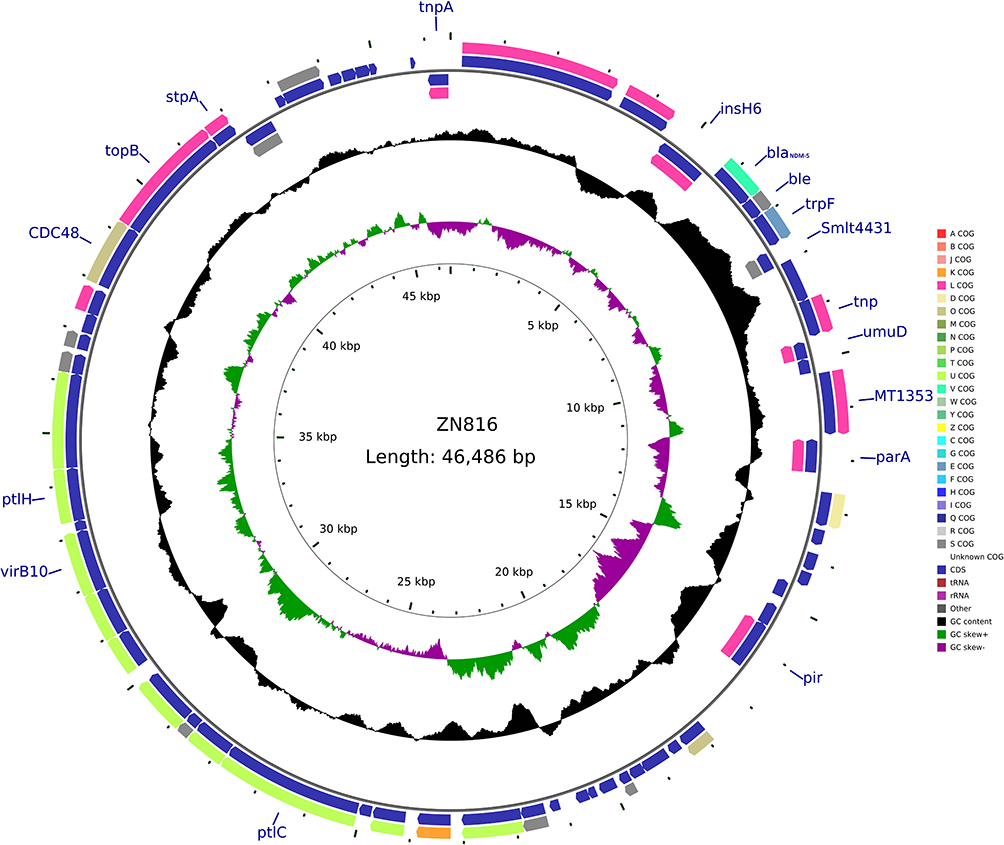 |
Figure 4 Structure of plasmid pNDM-5-N816 carrying blaNDM-5 from Klebsiella pneumoniae N816. |
Discussion
Carbapenems are the choice of treatment for infections caused by MDR K. pneumoniae. However, with the emergence of carbapenemase-producing bacteria, carbapenem resistance is increasing. The most common carbapenemase gene is blaKPC-2.16 Since the first discovery of the carbapenem resistance gene, blaNDM-1, in New Delhi, India,17 this gene and its multiple subtypes have been gradually discovered and reported worldwide. Moreover, the emergence of MBL-producing drug-resistant bacteria poses a great challenge for the treatment of drug-resistant bacterial infections. China reported a CRKP strain carrying the blaNDM-1 gene in 2013.18
Polymyxins have been used for many years in veterinary medicine, and nowadays, in human medicine, as a last resort for the treatment of MDR infections, especially CRKP. Thus, the increase in carbapenemase-producing Enterobacteriaceae has resulted in increased use of polymyxins with the inevitable risk of emerging polymyxin resistance.19,20 In this study, 107 CRKP isolates were tested for antimicrobial susceptibility; 15 (14.0%) of them were resistant to polymyxin B. The resistance rates of CRKP isolates to polymyxin B reported in Brazil and other European countries are 15.5% and 36%, respectively.20,21 The differences in antimicrobial resistance rates may be related to the different levels of antimicrobial usage in different countries.22
We found that 15 isolates were resistant to broad-spectrum antibiotics. Sequencing analysis showed that in addition to the blaKPC-2 gene, one or more other kinds of β-lactamase genes (such as blaCTX-M, blaSHV, and blaTEM) were identified in these KPC-producing K. pneumoniae strains, with 53% (8/15) of the strains carrying the ESBL gene, blaCTX-M; this is consistent with previous reports.23 Consistent with other studies, the isolates were also more resistant to quinolones and trimethoprim/sulfamethoxazole. Quite often, plasmids carrying ESBL genes also carry other drug-resistant genes including quinolone and trimethoprim/sulfamethoxazole resistance genes.24
The drug susceptibility results of this study showed CRKP has a low resistance to amikacin, possibly because amikacin has only been used for a short time in this region or owing to the presence of “restorative” mutations. It may also be because of the aminoglycoside-modifying enzymes produced when amikacin is used to treat CRKP; the 16S rRNA gene targeted by amikacin is prone to mutations, resulting in a decrease in the activity of the enzyme to hydrolyze it.25 Although the resistance rate of CRKP to tigecycline is also low in this study, Its FDA approved uses include complicated skin/skin structure infections, complicated intra-abdominal infections, and community-acquired bacterial pneumonia, treatment of these infections limits its use.26 Studies have shown that polymyxin combined with amikacin has obvious synergistic and additive effects, and the MICs of this combination therapy are significantly lower than those of monotherapy.27 Polymyxin and amikacin may be sensitive to each other and as they target multiple proteins through different mechanisms to inhibit biofilm formation and increase membrane permeability, a synergistic effect to inhibit CRKP isolates is exhibited.28
Among the 15 polymyxin B-resistant CRKP isolates, most of the strains carried the blaKPC-2 gene, which is primarily responsible for carbapenem resistance, and this is consistent with our previous report.16 Among these strains, we only detected one isolate positive for mcr-1 gene. This strain was isolated from a blood culture specimen of a 71-year-old male patient and was resistant to multiple drugs, including polymyxin B, but not amikacin. This isolate also had blaCTX-M-55, blaNDM-5 and blaSHV-27 genes. Our experiments and sequencing results show that these mcr-1, blaCTX-M-55, and blaNDM-5 genes do not appear to be on the same plasmid, and the blaSHV-27 gene was found on the bacterial chromosome. Consistent with the above experimental results, the MICs of the corresponding antibiotic of the conjugants and transformants were altered accordingly (Table 2). The other 14 isolates did not harbor the mcr-1 gene. Other drug resistance mechanisms may be related to specific mutations within the genes encoding LPS-modifying enzymes, resulting in increased levels of the intrinsic regulator RamA and hyperproduction of CPS.29,30 However, this needs further testing. Unlike reports in other countries where the mcr-1 and blaNDM-5 genes were found to coexist in E. coli,9,10 this is the first time that K. pneumoniae has been reported to harbor both mcr-1, blaNDM−5, and blaCTX-M-55 genes. What is even more worrying is that the plasmids in which these three genes are located have the ability to transfer horizontally. Thus, the bacteria may develop more serious drug resistance and lead to a state where there will be no treatment options. However, PFGE and MLST results of this isolate indicated that it had a different homology from the other 14 polymyxin B-resistant K. pneumoniae isolates, indicating that this type of bacteria did not have an outbreak. This isolate is not part of the most common ST (ST11) in China,16 but is a rare ST485 isolate. There is no report of an outbreak caused by K. pneumoniae ST485 at home and abroad. The patient had no history of foreign travel in the inquiry; thus, we infer that the occurrence of this isolate is a rare phenomenon in this area, but continuous monitoring to prevent the spread of this type of bacteria, which may cause more serious drug resistance, is warranted.
It has long been believed that polymyxin is mediated by chromosomes.19 Until 2015, Chinese scholars reported that the plasmid-mediated polymyxin resistance gene mcr-1 was found in Enterobacteriaceae isolated from humans and animals.3 Since then, people have a new understanding of the mechanism of polymyxin resistance. The emergence of a new type of drug resistance mechanism immediately set off a research boom among microbiologists worldwide. After sequencing and analysis of the plasmid obtained in this study, it was found that the isolate contained three plasmids of different sizes (33, 46, and 76 kb), which carried the mcr-1, blaCTX-M-55, and blaNDM-5 genes, which also verified the results of our previous conjugation, transformation, and drug susceptibility experiments. Further analysis of the data obtained using sequencing revealed that the similarity between the plasmid carrying mcr-1 and the plasmid pKP15450-MCR-1 was 98.77%. The mcr-1 gene at the starting point of the plasmid is approximately 3826 to 5451 bp, which encodes 541 amino acids. Analysis of its upstream and downstream genes showed that there are no common insert elements, indicating that the mcr-1 gene on this type of plasmid is more prone to horizontal transfer. The plasmid carrying the blaCTX-M-55 gene has a similarity of 99.96% with the plasmid pKP32558-4. The start site of the blaCTX-M-55 gene is approximately 2052 to 2927 bp, which encodes 291 amino acids and has an inserted transposon IS431 around the gene. The above two plasmids can be successfully joined to the same strain at the same time, indicating that they are compatible with each other. The plasmid carrying the blaNDM-5 gene has a similarity of 98.4% with the plasmid pNDM5-LDR. The start site of the blaNDM-5 gene is approximately 5783 to 6595 bp, which encodes 270 amino acids and has an inserted transposon, IS1086 and IS5H, upstream of the gene. The IS1086 and IS5H sequences indicate that the plasmid can transfer horizontally and confirm the results of previous experimental research. The resistance gene expression and transferability of this isolate have been further verified, which may lead to serious drug resistance.
Conclusion
In conclusion, the present study demonstrated for the first time the coexistence of mcr-1, blaNDM−5, and blaCTX-M-55 in a K. pneumoniae ST485 isolate. Therefore, treatment strategies and monitoring should be implemented to limit the widespread of isolates containing mcr-1, blaNDM−5, and blaCTX-M-55.
Ethics Statement
As the Klebsiella pneumoniae clinical isolate in this study was part of the routine hospital laboratory procedure, we have confirmed that the isolate has no identifiable patient data, the Second Affiliated Hospital of Nanchang University Medical Research Ethics Committee exempted this research for review.
Acknowledgments
This study was supported by a grant from the Department of Science and Technology of Jiangxi Province (20181BBG70030), the Jiangxi Natural Science Foundation (No.20181BAB205066), Science and Technology Plan of Jiangxi Provincial Health Commission (NO.20195211).
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflict of interest.
References
1. Arato V, Raso MM, Gasperini G, et al. Prophylaxis and treatment against Klebsiella pneumoniae: current insights on this emerging anti-microbial resistant global threat. Int J Mol Sci. 2021;22(8):4042. doi:10.3390/ijms22084042
2. Zhan L, Wang S, Guo Y, et al. Outbreak by hypermucoviscous Klebsiella pneumoniae ST11 isolates with carbapenem resistance in a tertiary hospital in China. Front Cell Infect Microbiol. 2017;7:182. doi:10.3389/fcimb.2017.00182
3. Chavda B, Lv J, Hou M, et al. Coidentification of mcr-4.3 and blaNDM-1 in a clinical Enterobacter cloacae isolate from China. Antimicrob Agents Chemother. 2018;62(10):e00649–18. doi:10.1128/AAC.00649-18
4. Hussein M, Han ML, Zhu Y, et al. Metabolomics study of the synergistic killing of polymyxin B in combination with amikacin against polymyxin-susceptible and -resistant pseudomonas aeruginosa. Antimicrob Agents Chemother. 2019;64(1):e01587–19. doi:10.1128/AAC.01587-19
5. Moffatt JH, Harper M, Harrison P, et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob Agents Chemother. 2010;54(12):4971–4977. doi:10.1128/AAC.00834-10
6. Bonura CMC, Bernardo FD, Aleo A, et al. Ongoing spread of colistin-resistant Klebsiella pneumoniae in different wards of an acute general hospital, Italy, June to December 2011. Euro Surveill. 2012;17(33):20248. doi:10.1186/1475-2875-11-277
7. Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/s1473-3099(15)00424-7
8. Quan J, Li X, Chen Y, et al. Prevalence of mcr-1 in Escherichia coli and Klebsiella pneumoniae recovered from bloodstream infections in China: a multicentre longitudinal study. Lancet Infect Dis. 2017;17(4):400–410. doi:10.1016/s1473-3099(16)30528-x
9. Yang RS, Feng Y, Lv XY, et al. Emergence of NDM-5- and MCR-1-producing Escherichia coli clones ST648 and ST156 from a Single Muscovy Duck (Cairina moschata). Antimicrob Agents Chemother. 2016;60(11):6899–6902. doi:10.1128/AAC.01365-16
10. Zhang Y, Liao K, Gao H, et al. Decreased fitness and virulence in ST10 Escherichia coli harboring blaNDM-5 and mcr-1 against a ST4981 strain with blaNDM-5. Front Cell Infect Microbiol. 2017;7:242. doi:10.3389/fcimb.2017.00242
11. CLSI. Performance Standards for Antimicrobial Susceptibility Testing.
12. The European Committee on Antimicrobial susceptibility testing breakpoint tables for interpretation of MICs and zone diameters. Version 6.0,2016. Available from: http://www.eucast.org. Accessed March 30,2016.
13. Yang F, Shen C, Zheng X, et al. Plasmid-mediated colistin resistance gene mcr-1 in Escherichia coli and Klebsiella pneumoniae isolated from market retail fruits in Guangzhou, China. Infect Drug Resist. 2019;12:385–389. doi:10.2147/idr.s194635
14. Chen L, Hu H, Chavda KD, et al. Complete sequence of a KPC-producing IncN multidrug-resistant plasmid from an epidemic Escherichia coli sequence type 131 strain inChina. Antimicrob Agents Chemother. 2014;58(4):2422–2425. doi:10.1128/aac.02587-13
15. Wang Z, Li M, Shen X, et al. Outbreak of blaNDM-5-harboring Klebsiella pneumoniae ST290 in a tertiary hospital in China. Microb Drug Resist. 2019;25(10):1443–1448. doi:10.1089/mdr.2019.0046
16. Hu L, Liu Y, Deng L, et al. Outbreak by ventilator-associated ST11 K. pneumoniae with co-production of CTX-M-24 and KPC-2 in a SICU of a tertiary teaching hospital in central China. Front Microbiol. 2016;7:1190. doi:10.3389/fmicb.2016.01190
17. Yong D, Toleman MA, Giske CG, et al. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53(12):5046–5054. doi:10.1128/AAC.00774-09
18. Hu L, Zhong Q, Tu J, et al. Emergence of blaNDM-1 among Klebsiella pneumoniae ST15 and novel ST1031 clinical isolates in China. Diagn Microbiol Infect Dis. 2013;75(4):373–376. doi:10.1016/j.diagmicrobio.2013.01.006
19. Mendes AC, Novais A, Campos J, et al. mcr-1 in carbapenemase-producing Klebsiella pneumoniae with hospitalized patients, Portugal, 2016–2017. Emerg Infect Dis. 2018;24(4):762–766. doi:10.3201/eid2404.171787
20. Wang R, van Dorp L, Shaw LP, et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat Commun. 2018;9(1):1–9. doi:10.1038/s41467-018-03205-z
21. Di Tella D, Tamburro M, Guerrizio G, et al. Molecular epidemiological insights into colistin-resistant and carbapenemases-producing clinical Klebsiella pneumoniae. Infect Drug Resist. 2019;Volume 12:3783–3795. doi:10.2147/idr.s226416
22. European Centre for Disease P., Control, A. European Food Safety. ECDC/EFSA/EMA second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals: Joint Interagency Antimicrobial Consumption and Resistance Analysis (JIACRA) Report. EFSA J. 2017;15(7):e04872. doi:10.2903/j.efsa.2017.4872
23. Liu H, Lin H, Sun Z, et al. Distribution of beta-lactamase genes and genetic context of bla KPC-2 in clinical carbapenemase-producing Klebsiella pneumoniae Isolates. Infect Drug Resist. 2021;14:237–247. doi:10.2147/IDR.S290434
24. Xiao Y, Hang Y, Chen Y, et al. A retrospective analysis of risk factors and patient outcomes of bloodstream infection with extended-spectrum beta-lactamase-producing Escherichia coli in a Chinese tertiary hospital. Infect Drug Resist. 2020;13:4289–4296. doi:10.2147/IDR.S269989
25. Zhang J, Yu L, Fu Y, et al. Tigecycline in combination with other antibiotics against clinical isolates of carbapenem-resistant Klebsiella pneumoniae in vitro. Ann Palliat Med. 2019;8(5):622–631. doi:10.21037/apm.2019.09.11
26. Gong J, Su D, Shang J, et al. Efficacy and safety of high-dose tigecycline for the treatment of infectious diseases: a meta-analysis. Medicine. 2019;98(38):e17091. doi:10.1097/MD.0000000000017091
27. Yu L, Zhang J, Fu Y, et al. Synergetic effects of combined treatment of colistin with meropenem or amikacin on carbapenem-resistant Klebsiella pneumoniae in vitro. Front Cell Infect Microbiol. 2019;9:422. doi:10.3389/fcimb.2019.00422
28. Ayoub Moubareck C. Polymyxins and bacterial membranes: a review of antibacterial activity and mechanisms of resistance. Membranes. 2020;10(8):181. doi:10.3390/membranes10080181
29. Poirel L, Jayol A, Nordmann P. Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin Microbiol Rev. 2017;30(2):557–596. doi:10.1128/CMR.00064-16
30. Petrosillo N, Taglietti F, Granata G. Treatment options for colistin resistant Klebsiella pneumoniae: present and future. J Clin Med. 2019;8(7):934. doi:10.3390/jcm8070934
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.