")
Back to Journals » Infection and Drug Resistance » Volume 12
Effects of sub-inhibitory concentrations of meropenem and tigecycline on the expression of genes regulating pili, efflux pumps and virulence factors involved in biofilm formation by Acinetobacter baumannii
Authors Navidifar T , Amin M, Rashno M
Received 21 January 2019
Accepted for publication 16 March 2019
Published 7 May 2019 Volume 2019:12 Pages 1099—1111
DOI https://doi.org/10.2147/IDR.S199993
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Tahereh Navidifar,1,2 Mansour Amin,2,3 Mohammad Rashno4
1Cellular and Molecular Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 3Health Research Institute, Infectious and Tropical Diseases Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 4Department of Immunology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Background: Sub-minimal inhibitory concentrations of antibiotics have been indicated to affect the biofilm formation in pathogens of nosocomial infections. This study aimed to investigate the effects of meropenem and tigecycline at their sub-minimum inhibitory concentrations (MICs) on the biofilm formation capacity of Acinetobacter baumannii (A. baumannii), as well as the expression levels of genes involved in biofilm formation, quorum sensing, pili assembly and efflux pumps.
Materials and methods: In this study, four non-clonal strains (AB10, AB13, AB32 and AB55), which were different from the aspects of antibiotic susceptibility and biofilm formation from each other were selected for the evaluation of antimicrobial susceptibility, biofilm inducibility at sub-MICs of meropenem and tigecycline and the gene expression levels (the abaI, abaR, bap, pgaA, csuE, bfmS, bfmR, ompA, adeB, adeJ and adeG genes).
Result: A significant increase in the MICs of all antibiotics was demonstrated in the biofilm cells in each four strains. The biofilm formation was significantly decreased in all the representative strains exposed to tigecycline. However, the biofilm inducibility at sub-MICs of meropenem was dependent on strain genotype. In concordance with these results, Pearson correlation analysis indicated a positive significant correlation between the biofilm formation capacity and the mRNA levels of genes encoding efflux pumps except adeJ, the genes involved in biofilm formation, pili assembly and quorum sensing following exposure to meropenem and tigecycline at their sub-MICs.
Conclusion: These results revealed valuable data into the correlation between the gene transcription levels and biofilm formation, as well as quorum sensing and their regulation at sub-MICs of meropenem and tigecycline.
Keywords: Acinetobacter baumannii, sub-MIC, meropenem, tigecycline, biofilm formation, gene expression
Introduction
Acinetobacter baumannii is one of the opportunistic bacterial pathogens that primarily associated with a wide variety of hospital-acquired infections, particularly those who have hospitalized for a long time.1 This bacterium has a high propensity to acquire a wide variety of antibiotic resistance determinants, as well as the capability of biofilm formation that these two characteristics play the important roles in treatment failure of this bacterium.2 Moreover, bacteria inside biofilms can tolerate the higher concentrations of antibiotics up to 1000 times more than their planktonic mode.3 The biofilm formation in A. baumannii is positively correlated with the transcription levels of several virulence factors, including two surface proteins of OmpA and Bap, the CsuABCDE operon that encodes type 1 pili, the pgaABCD locus that encodes proteins that synthesize cell-associated poly-ß-(1–6)-N-acetylglucosamine (PNAG) and the abaI gene that encodes acyl-homoserine lactones (AHL) as signal molecules.4
Moreover, the CsuABCDE operon is one of the key factors in the biofilm formation of A. baumannii that is controlled by a two-component regulatory system of BfmS/BfmR.5 Previous findings indicated that BfmR is essential for the stabilization of the csu operon, especially the csuE gene, as well as the biofilm formation.6
On the other hand, the efflux pumps involved in multidrug resistance especially the resistance-nodulation-cell division (RND) family display several different roles during the transition of planktonic cell to biofilm in A. baumannii. Moreover, these pumps have extruded actively the autoinducers associated with quorum sensing, as well as harmful molecules such as antibiotics and metabolic intermediates, resulting in the regulation of the biofilm formation and quorum sensing processes directly and indirectly.7
As described in previous studies, during the biofilm formation, a gradient of available substances such as oxygen, nutrient, pH, antibiotic is established; hence the cells within the inner layers of biofilm have a limited availability to the penetration of antibiotics, ie, these cells are exposed to sub-inhibitory concentrations of antibiotics.8 Moreover, several researchers showed that some antibiotics at sub-minimum inhibitory concentrations (MICs) can alter some bacterial functions such as the bacterial ultrastructure, the biofilm formation, the transcription of bacterial virulence factors and adhesions.9–12
Carbapenems as a sustainable group of antibiotics with the high activity and low toxicity are recommended for the treatment of infections associated with A. baumannii.13 However, in recent years, the emergence of the multidrug resistance A. baumannii (MDR-AB) isolates, which are resistant to carbapenems are increasing worldwide. So that the increasing resistance to carbapenems has limited their clinical use.14 Hence, the introduction of alternative antibiotic choices for the treatment of the MDR-AB infections is critical. Among antibiotic agents, polymyxins and tigecycline remain as the only active antibiotic choices against these infections.15 Moreover, a previous study by Sato et al indicated that colistin induced the biofilm formation in A. baumannii and increased the transcription levels of the genes associated with the biofilm.16 However, the effect of tigecycline at sub-MICs has already been not studied on the transcription levels of the genes associated with the A. baumannii biofilm. Hence, this current study was aimed to evaluate the effects of meropenem and tigecycline at their sub-MICs on the biofilm formation capacity of A. baumannii, as well as the expression levels of the genes involved in biofilm formation, efflux pumps and pili regulation.
Material and methods
Bacterial strains and antibiotic susceptibility
In this current study, four none-clonal strains (AB10, AB13, AB32 and AB55) based on ERIC–PCR patterns (data not shown) were selected for more analysis. Moreover, these four strains had differed from each other in aspects of the antibiotic susceptibility and the biofilm formation capability, as mentioned in Table 2. Identification of these isolates was performed using standard biochemical tests17 and confirmed by the amplification of blaOXA-51-like gene.18The study design was approved by the Research Ethics Committee of Ahvaz Jundishapur University of Medical Sciences (AJUMS.REC.1396.333), Iran.
 | Table 1 Primers used in this study |
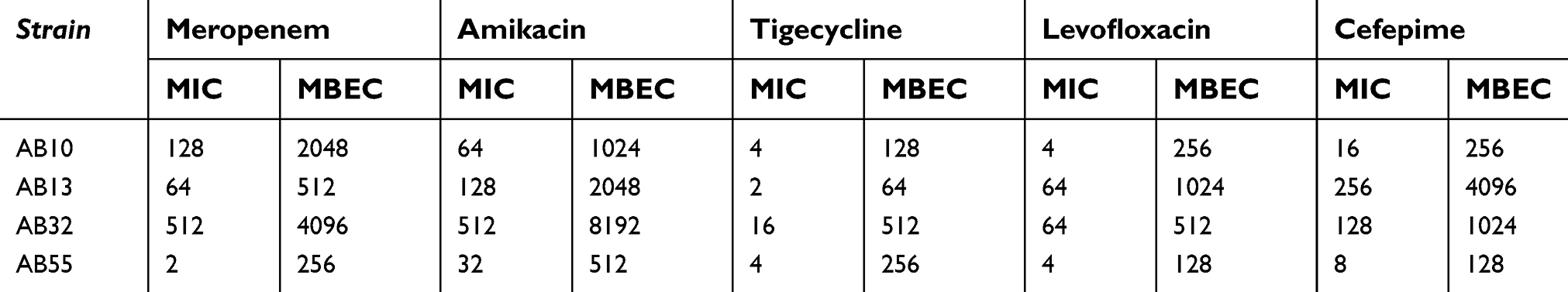 | Table 2 Antibiotic susceptibility of strains in planktonic and biofilm mode |
Biofilm formation determination and quantification
The biofilm formation capability of A. baumannii isolates was evaluated using the crystal violet staining method in the 96-well polystyrene microtiter plates, as previously described.19 Also, A. baumannii ATCC19606 and Muller Hinton Broth were used as positive and negative controls for the biofilm formation, respectively. The results were interpreted according to the criteria suggested by Zhang et al20.
Antibiotic susceptibility testing
The minimum inhibitory concentrationsof levofloxacin, amikacin, meropenem, tigecycline and cefepime were determined using broth microdilution method and their results were interpreted according to the Clinical and Laboratory Standards Institute (CLSL) guidelines (CLSL, 2018).21 Briefly, for levofloxacin, amikacin, meropenem, and cefepime, the MICs of greater than or equal to 8, 64, 8 and 32 µg/mL are considered as the resistant breakpoints, respectively. In addition, for tigecycline a MIC of greater than or equal to 8 µg/mL is proposed as the resistant breakpoint according to the criteria suggested by Jones et al.22
Biofilm antibiotic susceptibility testing
The minimum biofilm eradication concentration (MBEC) values of levofloxacin, amikacin, meropenem, tigecycline and cefepime in A. baumannii isolates were measured using the broth microdilution method.19 First, the isolates were cultivated in the sterile 96-well polystyrene microtiter plates for an overnight at 37°C to allow for the biofilm formation. The biofilms were then exposed to the concentrations of 2–4,096 µg/mL of levofloxacin, 4–8,192 µg/mL of amikacin, 2–8,192 µg/mL of meropenem, 0.5–2048 µg/mL of tigecycline and 16–16,384 µg/mL of cefepime for an overnight at 37°C.Then, the wells were washed with sterile PBS three times, and incubated with Muller Hinton Broth (Merck, Darmstadt, Germany) for an overnight at 37°C. The MBEC was proposed as any viable cell was not recovered from the biofilm material or, ie, OD of 570nm (OD570) was <0.1. All tests were repeated in triplicate.
Biofilm formation in the presence of sub-MICs of tigecycline and meropenem
First, each strain was inoculated in the 96-well polystyrene microtiter plates at approximately 106 CFU/ml in cation-adjusted Mueller–Hinton broth with the different sub-inhibitory concentrations (1/8, 1/4 and 1/2×the MIC) of either tigecycline or meropenem. Then, the plates were incubated at 37°C for an overnight and the quantification of biofilms was performed as mentioned in the previous section. The antibiotic-free medium in well was used as negative control. Also, A. baumannii ATCC19606 was used as the positive control strain for the biofilm formation in the presence of sub-MICs of tigecycline and meropenem. The results were described as the OD570 ratio of the sub-MICs, ie, the 1/8×MIC, 1/4×MIC or 1/2×MIC of tigecycline or meropenem to the OD570 of control sample (0 MIC).23
Quantitative real-time PCR assay
First, these four representative A.baumannii strains were exposed to sub-inhibitory concentrations of either tigecycline or meropenem as described in before section. Then, RNA extraction was performed using an RNeasy plus Mini kit (Qiagen, Tokyo, Japan). The quality and integrity of the total RNA were evaluated with the NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and electrophoresed on 1% agarose gel. The final concentration of the RNA extracts of these four strains was adjusted to 400 ng/µL. The RNA was reverse transcribed to cDNA using PrimeScript™ 1st strand cDNA Synthesis Kit (Qiagen) according to the manufacturer’s procedure (Transgen Bio-Technology Company, Beijing, China). The cDNA was kept at -20°C. Real-time PCR amplification reaction was prepared in a final volume 20 µL, with 400 ng cDNA, 10 µL RealQ Plus Master Mix Green (Amliqon, Denmark) and 0.5 µL each of forward and reverse primers (10 nM each) and RNase- and DNase-free water up to in the final volume 20 µL. The primer sequences used for the genes involved in biofilm formation (bap, ompA, csuE and pgaA), quorum sensing (abaI and abaR), pili regulation (bfmS and bfmR) and efflux pumps (adeB, adeG and adeJ) are shown in Table1.4,5,23,24 The 16rRNA gene was used as an internal control for the normalization of the mRNA expression. Real-time PCR was performed using a Step One Real-Time PCR System (Applied Biosystems, CA, USA) as follows: on cycle of initial denaturation at 95°C for 15 mins, 40 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 30 s. The relative expression fold changes of mRNAs were calculated using the 2-ΔΔCt method. The relative expression of each gene after the exposure of the bacteria at sub-MICs of meropenem and tigecycline was normalized to the control sample (0 MIC), which was assigned a value of 1 arbitrary unit.
Statistical analysis
The mRNA expression analysis was performed using Student’s t test and one-way ANOVA, followed by the Tukey multiple comparison test. Pearson correlation analysis was used to analyze the gene expression levels and biofilm formation as well as quorum sensing. In all analyses, a two-sided significance level of <0.05 was considered statistically significant.
Results
Antibiotic susceptibility of strains in planktonic and biofilm mode
The values of MIC and MBEC of these representative four strains to antibiotic agents mentioned above is shown in Table 2. According to these results, the MIC values of meropenem, levofloxacin, cefepime, tigecycline and amikacin of these strains ranged from 2 to 512 µg/mL, 4 to 64 µg/mL, 8 to 256 µg/mL, 2 to 16 µg/mL and 32 to 512 µg/mL, respectively. As expected, the MBECs of these antibiotics were higher than their respective MICs, followed by 512–8192 µg/mL for amikacin, 128–4,096 µg/mL for cefepime, 128–1,024 µg/mL for levofloxacin, 256–4,096 µg/mL for meropenem and 64–512 µg/mL for tigecycline. With analysis of MBEC and MIC values of these antibiotics, we indicated an increase of 16-fold higher MBEC values rather than MIC values for amikacin, 8- to 128-fold for meropenem, 8- to 16-fold for cefepime, 8- to 64-fold for levofloxacin and 32- to 64-fold for tigecycline.
Effects of sub-MICs of tigecycline and meropenem on the biofilm formation
The greatest ability of the biofilm formation in the absence of antibiotics was belonged to strain AB55 (OD570: 0.984), followed by strain AB10 (OD570: 0.271), strain AB13 (OD570: 0.241) and strain AB32 (OD570: 0.152). Figure 1 demonstrates the biofilm formation capacity of the representative strains in the presence of levofloxacin and meropenem at 1/8, 1/4, and 1/2× the MICs rate to the biofilm formation in the absence of these antibiotics.
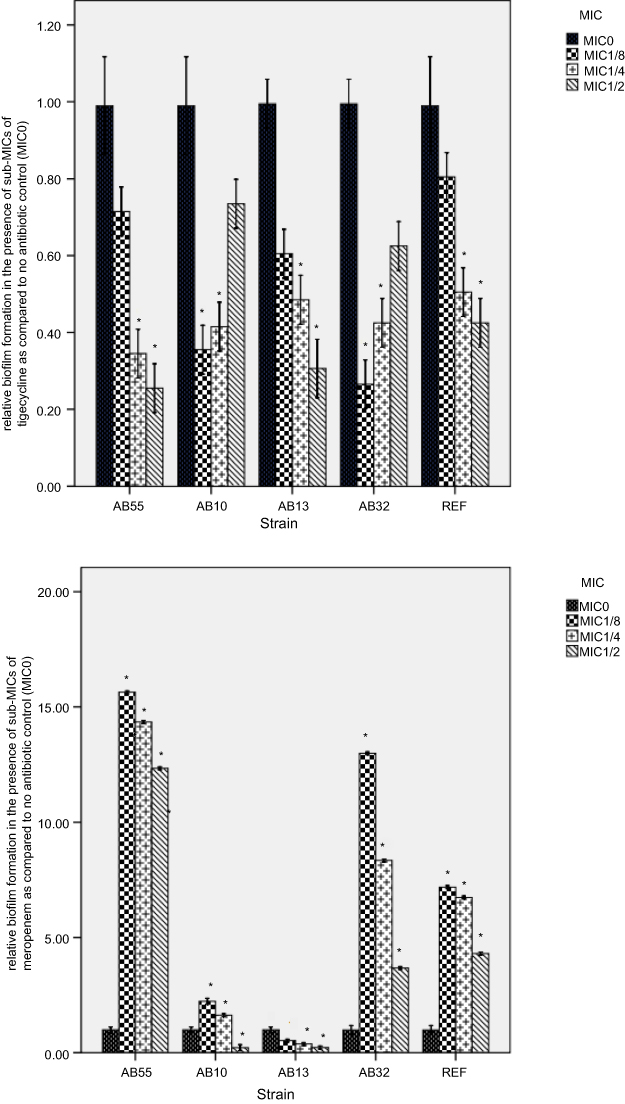 | Figure 1 Biofilm formation by A. baumannii strains in the presence of sub-MICs of tigecycline and meropenem. Error bars represent the standard deviations; *significant difference at a P-value of 0.05. REF: A. baumannii 19606. Abbreviations: A. baumannii, Acinetobacter baumannii; MIC, minimum inhibitory concentration; REF, reference srain. |
For strain AB55, following exposure to tigecycline, the biofilm formation was decreased significantly at concentrations of 1 and 2 µg/mL by 0.65- and 0.68-fold changes, whereas meropenem induced significantly the biofilm formation at concentrations of 0.25 µg/mL (15.64-fold), 0.5 (14.35-fold) and 1 µg/mL (12.33-fold).
For strain AB10, the significant decrease of the biofilm formation was observed in the presence of tigecycline at both the concentrations of 0.5 and 1 µg/mL, resulting in 0.65- and 0.58-fold changes, respectively. Also, following exposure to meropenem,the biofilm formation induced significantly at concentration of 16 µg/mL (a 2.23-fold change), whereas reduced significantly at concentration of 64 µg/mL (a 0.78-fold change).
For strain AB13, the biofilm formation was decreased significantly in the presence of tigecycline at concentrations of 0.5 and 1 µg/mL (by 0.52– and 0.69- fold changes) and meropenem at both concentrations of 16 and 32 µg/mL (by 0.62– to 0.78- fold changes).
For strain AB32, the significant decrease of the biofilm formation was observed in the presence of tigecycline at concentrations of 2 and 4 µg/mL, resulting in 0.73- and 0.57-fold changes, respectively. However, meropenem induced significantly the biofilm formation in a concentration-dependent manner, resulting in the changes of 15.64-, 14.35-, 12.34-fold at the concentrations of 64, 128 and 256 µg/mL respectively.
Expression levels of genes regulating pili, efflux pumps and virulence factors involved in the presence of sub-MICs of tigecycline
Figure 2 shows the effect of tigecycline at sub-MICs on the expression levels of the efflux pumps, pili regulation and biofilm involved genes in A. baumannii strains.
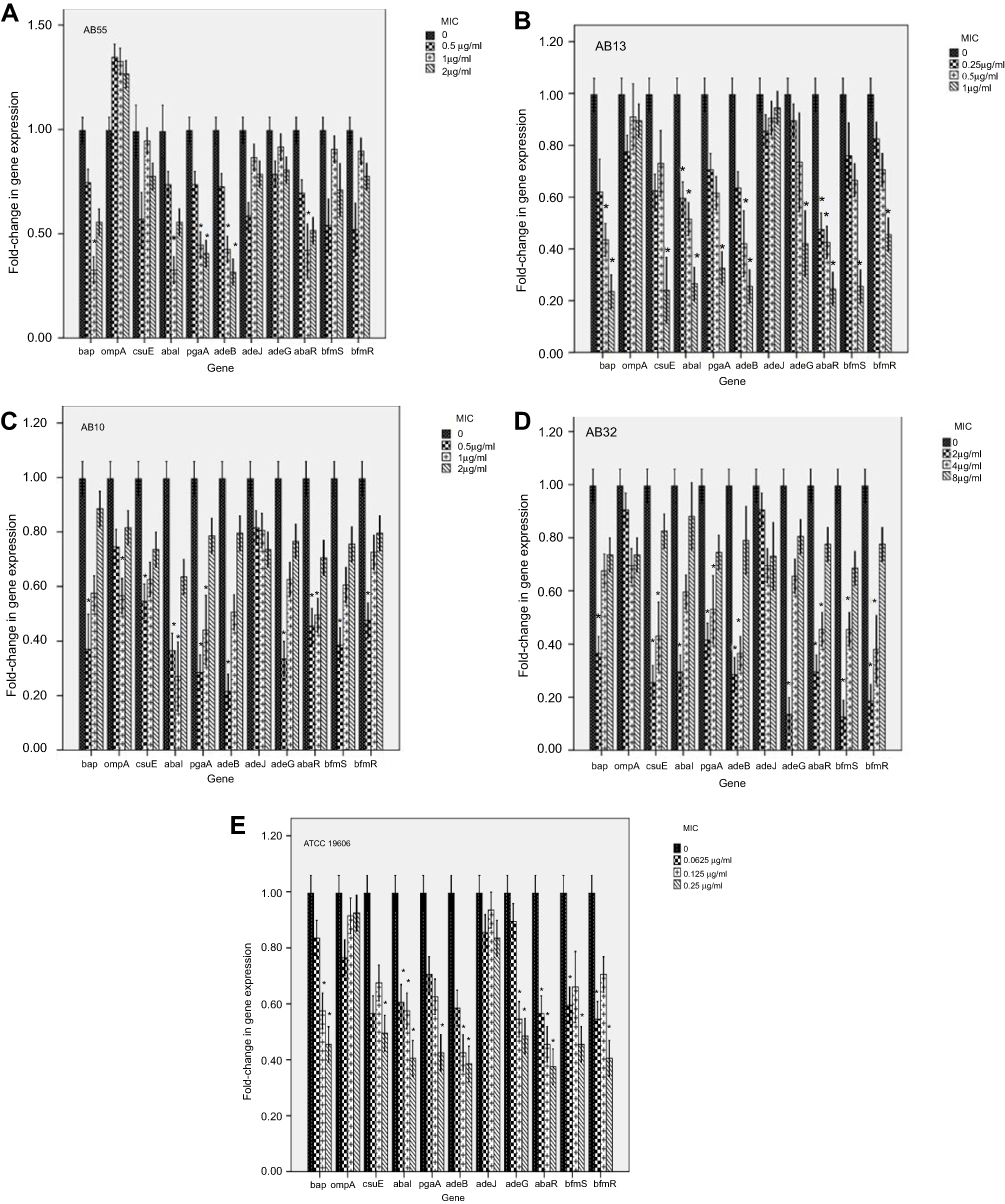 | Figure 2 Effect of tigecycline at sub-MICs on the expression levels of the efflux pumps, pili regulation and biofilm involved genes in A. baumannii strains. Error bars represent the standard deviations; *significant difference at a P-value of 0.05. Abbreviations: A. baumannii, Acinetobacter baumannii; MIC, minimum inhibitory concentration. |
For strain AB55, the gene expression levels of the bap (0.68- fold), the abaI (0.68- fold), the abaR (0.58- fold) were significantly decreased at the concentration of 0.25 µg/mL, as well as the pgaA (0.55- and 0.60- fold) and the adeB (0.57- and 0.69- fold) at concentrations of 0.25 and 0.5 µg/mL, respectively. However, the relative expression levels of the ompA, bfmS, bfmR, csuE and adeJ genes were not significantly changed at any concentration (P>0.05).
For strain AB10, the significant decreases in the relative expression levels were observed for the bap (0.63-fold), the csuE (0.46-fold), the adeB (0.78-fold), the adeG (0.66-fold), the bfmS (0.61-fold) and the bfmR (0.52-fold) at concentration of 0.5 µg/mL, the ompA (0.43-fold) at the concentration of 0.5 µg/mL, as well as the abaI (0.63- and 0.73-fold), the abaR (0.55- and 0.50-fold) and the pgaA (0.71- and 0.56-fold) at both concentrations of 0.5 and 1 µg/mL, respectively. However, the relative expression level of the adeJ gene was not significantly changed at any concentration (P>0.05).
For strain AB13, the significant decreases in the relative gene expression levels were observed for the csuE gene (0.76-fold), the pgaA (0.67-fold), the adeG (0.58-fold), the bfmS (0.75-fold) and the bfmR (0.55-fold) at the concentration of 1 µg/mL, the bap (0.57- and 0.77-fold) and adeB (0.58- and 0.75- fold) at both the concentrations of 0.5 and 1 µg/mL, respectively; as well as the abaI (0.41- to 0.74- fold) and the abaR(0.53- to 0.75- fold) in a concentration dependent manner (0.25–1 µg/mL). However, the relative expression levels of the ompA and adeJ genes were not significantly changed at any concentration (P>0.05).
For strain AB32, the significant decreases in the relative expression levels were indicated for the bap (0.63- fold), the abaI (0.70- fold) and the adeG (0.86- fold) at concentration of 2 µg/mL, as well as the csuE (0.75- and 0.57-), the pgaA (0.58- and 0.47- fold), the adeB (0.72-and 0.64- fold), the abaR (0.70- and 0.55- fold), the bfmS (0.87- and 0.55- fold) and the bfmR (0.81- and 0.63- fold) at both concentrations of 2 and 4 µg/mL, respectively. However, the relative expression levels of the ompA and adeJ genes were not significantly changed at any concentration (P>0.05).
Expression levels of genes regulating pili, efflux pumps and virulence factors involved in the presence of sub-MICs of meropenem
Figure 3 shows the effect of meropenem at sub-MICs on the expression levels of the efflux pumps, pili regulation and biofilm involved genes in A. baumannii strains.
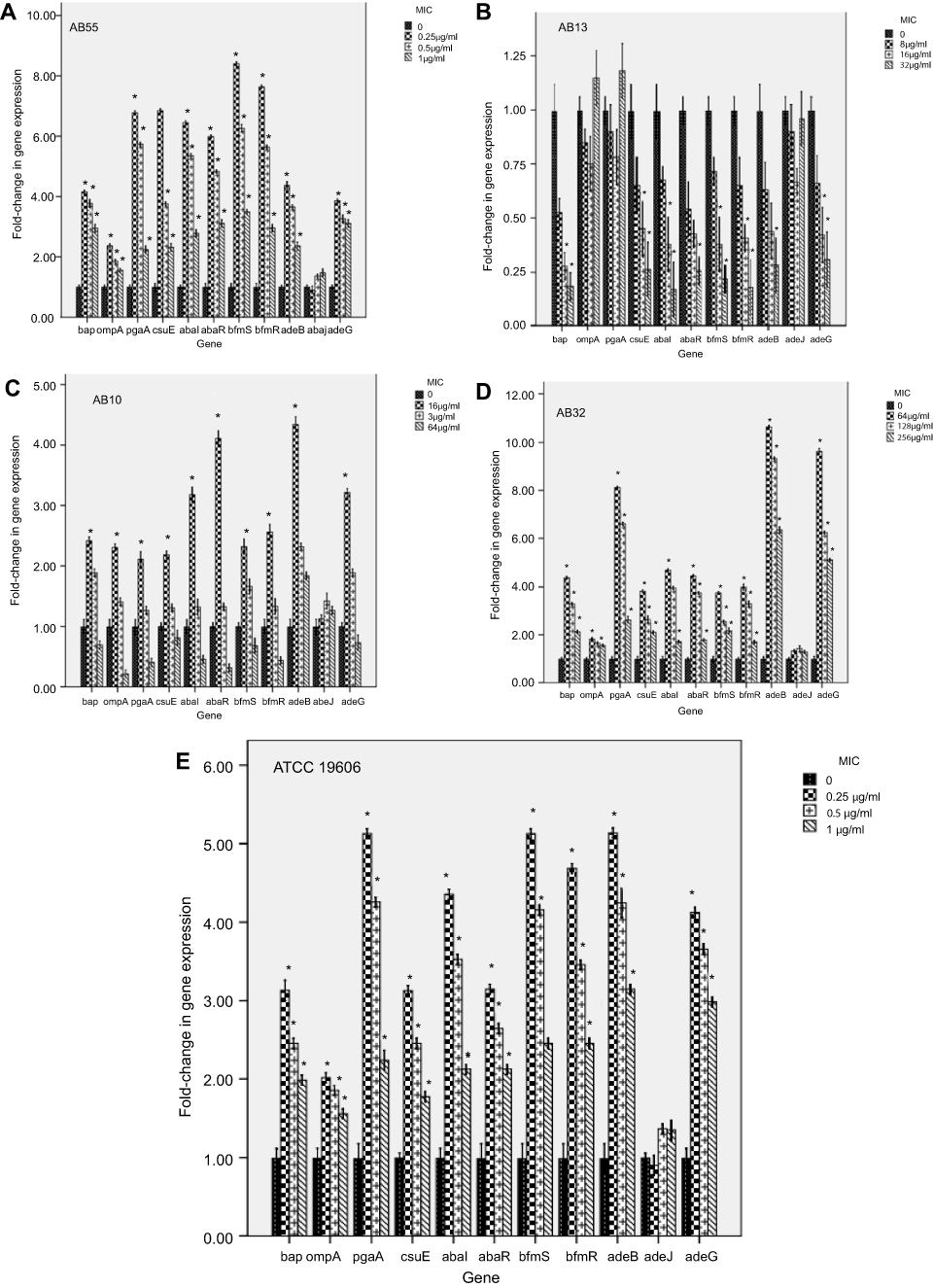 | Figure 3 Effect of meropenem at sub-MICs on the expression levels of the efflux pumps, pili regulation and biofilm involved genes in A. baumannii strains. Error bars represent the standard deviations; *significant difference at a P-value of 0.05. Abbreviations: A. baumannii, Acinetobacter baumannii; MIC, minimum inhibitory concentration. |
For strain AB55, the significant increases in the relative gene expression levels were observed for all of genes except the adeJ gene in a concentration-dependent manner (0.25–1 µg/mL).
For strain AB10, the significant increases in the relative gene expression levels were observed for the bap (2.41- fold), the csuE (2.19-), the pgaA (2.11-fold), the ompA(2.3- fold), the abaI (3.18- fold), the abaR (4.11- fold), the bfmS (2.23- fold), the bfmR (2.56- fold), the adeB (4.43- fold) and the adeG (3.21- fold) at the concentration of 16 µg/mL. However, the gene expression level of the adeJ was not significantly changed at any concentration (P>0.05).
For strain AB13, the significant decreases in the relative gene expression levels were observed for the abaR (0.75-fold) and the adeB (0.72- fold) at the concentration of 32 µg/mL, the bap (0.48- and 0.73- fold), the abaI (0.62- and 0.83- fold), the bfmS (0.62- and 0.79- fold), the bfmR (0.60- and 0.80- fold) and the csuE(0.55- and 0.76- fold) and the adeG (0.58- and 0.69- fold) at both concentrations of 16 and 32 µg/mL, respectively. However, the relative expression levels of the ompA, pgaA and adeJ were not significantly changed at any concentration (P>0.05).
For strain AB32, the significant increases in the relative gene expression levels were observed for the bap, pgaA, csuE, abaI, abaR, bfmS, bfmR, adeB and adeG genes at each three concentrations in a concentration-dependent manner (64–256 µg/mL). Moreover, a significant increase in the relative expression level of the ompA (1.81- and 1.63- fold) was observed at both the concentrations of 64 and 128 µg/mL, respectively; whereas the gene expression of the adeJ was not significantly changed at any concentration (P>0.05).
Correlation between biofilm formation and gene expression
To understand the correlation between the biofilm formation and the relative gene expression levels, we calculated the Pearson correlation coefficients between the capability of biofilm formation and the relative expression levels of the target genes (bap, ompA, csuE, pgaA, abaI, abaR, bfmS, bfmR, adeB, adeG and adeJ) for four strains of AB10, AB13, AB32, and AB55 exposed to sub-MICs of tigecycline (Table 3) and meropenem (Table 4).
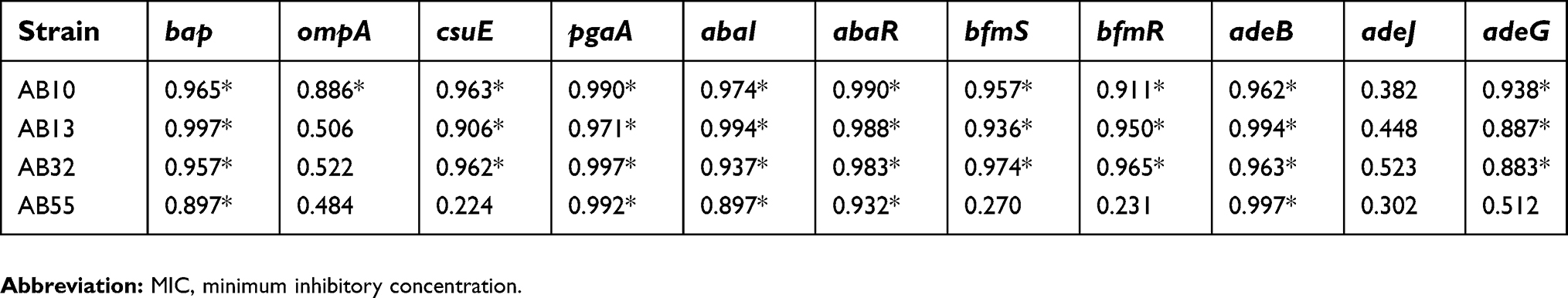 | Table 3 Association between biofilm formation and the gene expression profiles of A. baumannii strains at sub-MICs of tigecycline |
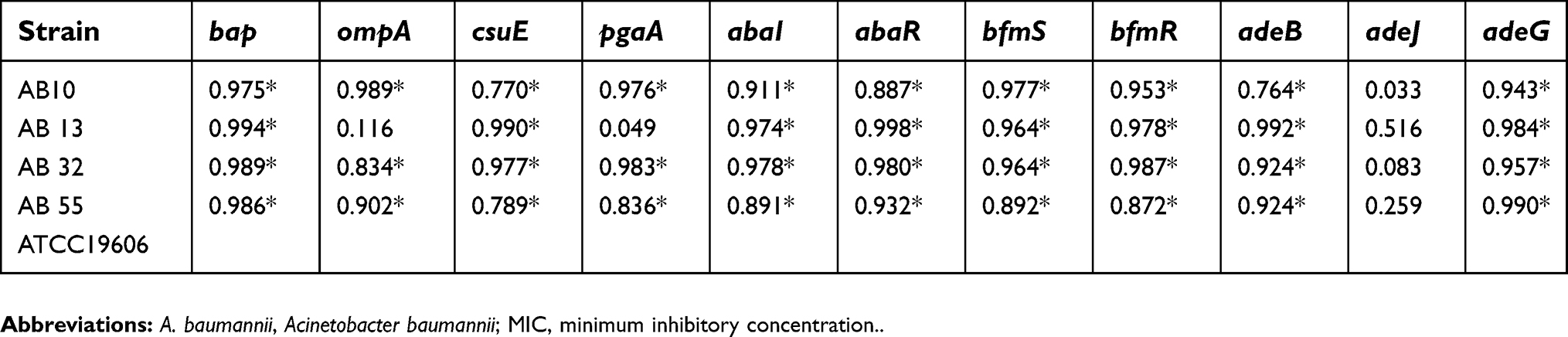 | Table 4 Association between biofilm formation and the gene expression profiles of A. baumannii strains at sub-MICs of meropenem |
In the presence of tigecycline, a significant positive correlation was indicated between the biofilm formation capacity and the gene expression levels of the bap, pgaA, csuE, pgaA, abaI, abaR, bfmS, bfmR, adeB and adeG in two strains of AB13 and AB32. Also, there was a significant correlation between the biofilm formation capacity and the expression levels of the bap, pgaA, abaR, adeG and adeB gene in strain of AB55. In addition, for strain of AB10, the biofilm formation capacity was highly correlated with the expression levels of all target genes except the adeJ gene.
In the presence of meropenem, a significant positive correlation was indicated between the biofilm formation capacity and the expression levels of all genes except the adeJ in three strains of AB55, AB32 and AB10. Also, the capability of biofilm formation was highly correlated with the expression levels of the bap, csuE, abaI, abaR, bfmS, bfmR, adeB and adeG genes in strain of AB13.
Correlation between quorum sensing and gene expression
To understand the correlation between the quorum sensing and the relative gene expression levels, we calculated the Pearson correlation coefficients between the capability of quorum sensing and the relative expression levels of the target genes (bap, ompA, pgaA, csuE, abaR,bfmS, bfmR, adeB, adeG and adeJ) for four strains of AB10, AB13, AB32, and AB55 exposed to sub-MICs of tigecycline (Table 5) and meropenem (Table 6).
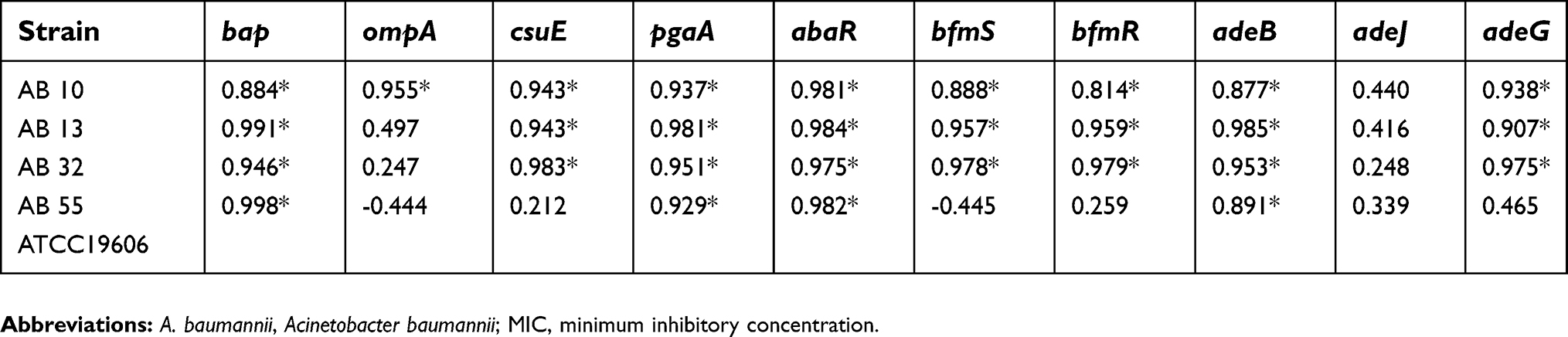 | Table 5 Association between Quorum sensing with the gene expression profiles of A. baumannii strains at sub-MICs of tigecycline |
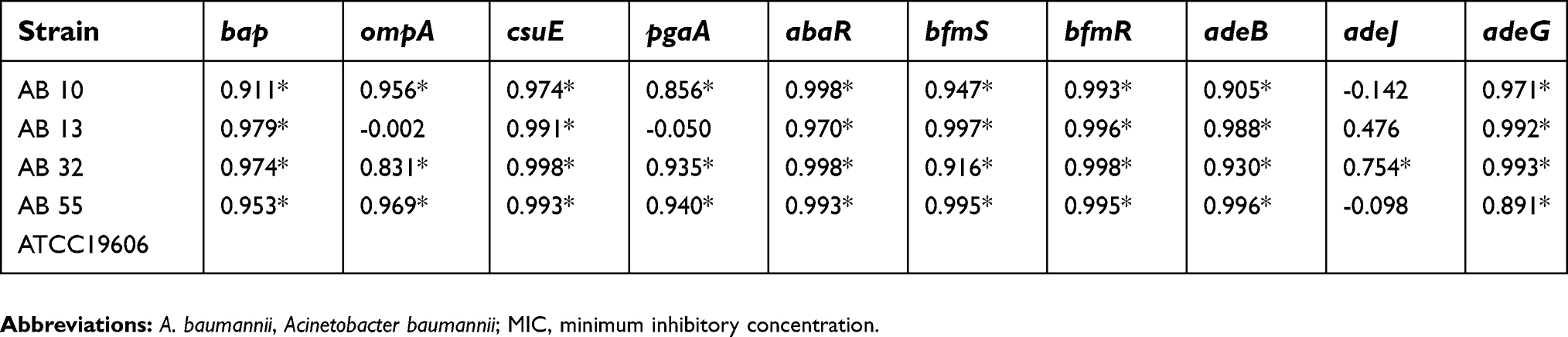 | Table 6 Association between Quorum sensing and the gene expression profiles of A. baumannii strains at sub-MICs of meropenem |
In the presence of tigecycline, a significant positive correlation was indicated between the quorum sensing (abaI) and the gene expression levels of the bap, pgaA, csuE, abaR, bfmS, bfmR, adeB and adeG in two strains of AB13 and AB32. Also, there was a significant correlation between the quorum sensing and the expression levels of the bap, pgaA, abaR and adeB gene in strain of AB55. In addition, for strain of AB10, the quorum sensing or the expression level of abaI gene was highly correlated with the expression levels of all target genes except the adeJ gene.
In the presence of meropenem, a significant positive correlation was indicated between the quorum sensing and the expression levels of all genes except the adeJ in two strains of AB55 and AB10. Also, for strain of AB13, the capability of biofilm formation was highly correlated with the expression levels of the bap, csuE, abaR, bfmS, bfmR, adeB and adeG genes. In addition, for strain of AB32, the quorum sensing was highly correlated with the expression levels of all target genes.
Discussion
Acinetobacter baumannii has emerged as one of the opportunistic pathogens causing nosocomial infections.1 The emergence of MDR strains as one of the main consequences of antibiotics excessive use in the treatment of human infections, compromises a major challenge to health systems worldwide.25 While most previous studies26–29 have investigated the different mechanisms of antibiotic resistance in A. baumannii, but there are few studies that evaluated the effects of antibiotics at sub-MICs on the biofilm formation and pathogenicity of A. baumannii.16,23 Hence, this study was aimed to investigate the effects of two antibiotics of meropenem and tigecycline on the biofilm formation capacity, as well as the expression levels of the genes involved in biofilm formation, efflux pumps and pili regulation in A. baumannii.
In this study, we indicated a significant increase of MBEC values compared to MIC values. This enhancement of MBEC values can be due to several factors such as the exopolysaccharide matrix of biofilm, overexpression of efflux pumps, persister biofilm cells and intrinsic characteristics of biofilm cells.30 Furthermore, the persister cells are metabolically dormant and are usually present in the stationary phase, as well as biofilm. These cells are extremely tolerant to antibiotics without undergoing any genetic change and may cause a relapse of infection.31
Carbapenems (meropenem and imipenem), as a class of ß-lactam antibiotics, are increasingly being used as first-line therapy of serious hospital-acquired infections.32 In the current study, we evaluated the effect of meropenem at sub-MICs on the biofilm formation capability in the four representative A. baumannii isolates. According to our results, meropenem induced significantly the capability of biofilm formation in two representative strains of AB55 and AB32, whereas decreased the biofilm formation in strain of AB13. Also, in AB10 strain, the biofilm formation was induced at the concentration of 1/8× the MIC while was decreased at the concentration of 1/2× the MIC. In agreement with our results, He et al23 demonstrated the different effects of meropenem at its sub-MICs on biofilm formation capability of non-clonal A. baumannii strains, indicating that meropenem has affected the biofilm formation dependent on strain type and highlight the importance of molecular typing methods prior to the choice of antibiotic therapy.
In this study, following exposure to sub-MICs of tigecycline, the ability of biofilm formation was decreased significantly in two strains of AB13 and AB55 at both concentrations of 1/4 and 1/2×the MIC, as well as two strains of AB32 and AB10 at both concentrations of 1/8 and 1/2×the MIC. Inconsistent with our results, Maestre et al33 and Chen et al34 reported that tigecycline at its sub-inhibitory concentrations interfered with forming biofilm by E. faecalis and A. baumannii strains, respectively. However, in contrast to our results, Szczuka et al35 and Weiser et al36 indicated that tigecycline induced forming biofilm by S. epidermidis through overexpression of extracellular matrix binding protein (Embp) and other biofilm-associated genes, suggesting that the effects of sub-MICs of tigecycline are almost dependent on bacterial species. In our study, tigecycline at sub-MICs decreased significantly the biofilm formation in these four representative strains whereas meropenem decreased significantly the biofilm formation only in two representative strains, suggesting that tigecycline rather than meropenem can interfere with the induction of biofilm formation in A. baumannii strains. Hence, exposure to the sub-MIC doses of tigecycline in patients is more effective than meropenem in killing A. baumannii strains without undergoing any effect on induction of biofilm formation.
In A. baumannii, AbaI/AbaR quorum sensing system is responsible for the synthesis and recognition of the AHLs. Following binding of the AHLs to AbaR, this conjugate binds to specific promoter DNA elements and regulate transcription of target genes such as genes involved in biofilm formation.37 Our results demonstrated a significant positive correlation between the expression levels of the abaI and abaR genes and biofilm formation at the sub-MICs of meropenem and tigecycline, suggesting a strong association between quorum sensing and forming biofilm by A. baumannii. Concordant to our results, previous studies also,16,23 confirmed a considerable correlation between the overexpression of the abaI gene and the biofilm formation when A. baumannii was exposed to levofloxacin, meropenem and colistin.
In addition, we studied the correlation between three RND efflux pumps of AdeABC, AdeFGH and AdeIJK with the biofilm formation, as well as quorum sensing. Our results indicated a significant positive correlation between the overexpression of the adeB and adeG genes and increased biofilm formation at the sub-MICs of meropenem and tigecycline in these four representative strains, that was in agreement with results obtained from the studies of Sato et al16 and He et al23 when A. baumannii was exposed to sub-MICs of antibiotics.
On the other hand, the up-regulation of the adeB and adeG genes was positively correlated with the transcription level of abaI gene, indicating a strong link between the RND efflux pumps (AdeABC and AdeFGH) and quorum sensing.
It seems that the overexpression of AdeABC and AdeFGH facilitate the transport of AHLs, resulting in the increase of the biofilm formation in A. baumannii. Hence, the inactivation of these two efflux pump by efflux pump inhibitors (EPIs) might be an alternative treatment approach to inhibit A. baumannii biofilm formation.23,38 In this study, the expression level of the adeJ gene was much low, so that any significant correlation was not found between the expression level of the adeJ gene and biofilm formation in these strains that is in agreement with the results of He et al23on A. baumannii biofilms. Moreover, several studies confirmed that the overexpression of the adeJ gene is lethal for the host; hence its expression is strictly regulated by A. baumannii biofilms.39–41
Our results showed that the transcription level of the csuE gene together with its regulatory genes, bfmS and bfmR, were positively correlated with the biofilm formation in all representative strains in the presence of either meropenem or tigecycline. Moreover, Tomaras et al42 and Pakharukova et al43 proved that the presence of type I pili on the surface of A. baumannii is critical in the early step of the biofilm formation on abiotic surfaces. So that the disruption of the csuC and csuE genes through direct mutagenesis resulted in non-piliated cells and abolishing the ability of the biofilm formation. Also, we indicated that the mRNA levels of the csuE gene together with the bfmS and bfmR were positively correlated with quorum sensing and implicitly the expression of the abaI gene. Also, the upregulation of the csuE gene was concordant to the expression levels of the BfmS and BfmR genes, as demonstrated by Luo et al5 Moreover, the researchers had proved that the increased expression of the BfmS and BfmR genes enhanced the expression level of the csu locus and subsequently forming pili for twitching motility in A. baumannii. Also, in support of our findings, they indicated the increased expression of all genes belonging to the csu locus together with chaperone-usher regulators (BfmS and BfmR) after addition of 100 µmol/L C6-HSL to culture medium of A. baumannii ATCC19606, suggesting a strong link between quorum sensing and forming type 1 pili.
OmpA and PNAG (encoded by the pga locus) in A. baumannii play the important roles in the colonization, immune evasion, antibiotic resistance and biofilm formation.44,45 Our results indicated a significant correlation between the expression level of the pgaA gene and biofilm formation when all of these representative strains were exposed to either tigecycline or meropenem (except strain AB05 at sub-MICs of tigecycline). Also, the transcription level of the ompA gene was positively correlated with biofilm formation in three strain of AB10, AB32, and AB55 in the presence of meropenem, as well as strain of AB10 at sub-MICs of tigecycline. In support of our findings, Sato et al16 indicated that the ompA and pgaA expression patterns were positively correlated with biofilm formation when A. baumannii strains were exposed to polymyxin B and colistin, respectively. Also, He et al23 proved that the expression regulation of the ompA gene was significantly correlated with forming biofilm at sub-MICs of either levofloxacin or meropenem.
Bap plays the important roles in the initial adherence to abiotic surfaces, the stabilization of mature biofilms, affecting both thickness and biovolume and subsequently the persistence in hospital infections.46 Our results showed that the biofilm formation at the sub-MICs of either levofloxacin or cefepime was positively correlated with the expression level of the bap gene in all of these representative strains. In agreement with our study, Sato et al16 also, demonstrated that polymyxin B altered the biofilm formation through the regulation of the bap gene.
Conclusion
In this study, we indicated that tigecycline rather than meropenem interfered with the induction of biofilm formation in A. baumannii strains. Also, the expression level of the adeB and adeG genes was positively correlated with the transcription level of abaI gene, indicating a strong link between the efflux pumps of AdeABC and AdeFGH and quorum sensing. In addition, we confirmed a positive correlation between the transcription level of the csuE gene together with its regulatory genes with the biofilm formation in all representative strains in the presence of either meropenem or tigecycline. Hence, blocking the efflux pump by EPIs or regulatory genes of type 1 pili might be an alternative treatment approach to inhibit A. baumannii biofilm formation.
Acknowledgments
This study was a part of the Ph.D. thesis of Tahereh Navidifar. This study was financially supported by the vice-chancellor of the Cellular and Molecular Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran (Grant No. CMRC-9614).
Author contributions
All authors contributed toward data analysis, drafting and revising the paper, gave final approval of the version to be published and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Mezzatesta ML, Trovato G, Gona F, et al. In vitro activity of tigecycline and comparators against carbapenem-susceptible and resistant acinetobacter baumannii clinical isolates in Italy. Ann Clin Microbiol Antimicrob. 2008;7:4. doi:10.1186/1476-0711-7-4
2. Longo F, Vuotto C, Donelli G. Biofilm formation in acinetobacter baumannii. New Microbiol. 2014;37(2):119–127.
3. Stewart PS. Antimicrobial Tolerance in Biofilms. Microbiol Spectr. 2015;3:3. doi:10.1128/microbiolspec.MB-0010-2014
4. Selasi GN, Nicholas A, Jeon H, et al. Differences in biofilm mass, expression of biofilm-associated genes, and resistance to desiccation between epidemic and sporadic clones of carbapenem-resistant acinetobacter baumannii sequence type 191. PLoS One. 2016;11(9):e0162576. doi:10.1371/journal.pone.0162576
5. Luo LM, Wu LJ, Xiao YL, et al. Enhancing pili assembly and biofilm formation in acinetobacter baumannii ATCC19606 using non-native acyl-homoserine lactones. BMC Microbiol. 2015;15:62. doi:10.1186/s12866-015-0397-5
6. Gaddy JA, Actis LA. Regulation of acinetobacter baumannii biofilm formation. Future Microbiol. 2009;4(3):273–278. doi:10.2217/fmb.09.5
7. Alav I, Sutton JM, Rahman KM. Role of bacterial efflux pumps in biofilm formation. J Antimicrob Chemother. 2018;73(8):2003–2020. doi:10.1093/jac/dky042
8. Lebeaux D, Ghigo JM, Beloin C. Biofilm-related infections: bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol Mol Biol Rev. 2014;78(3):510–543. doi:10.1128/MMBR.00013-14
9. Majtán J, Majtánová L, Xu M, Majtán V. In vitro effect of subinhibitory concentrations of antibiotics on biofilm formation by clinical strains of Salmonella enterica serovar typhimurium isolated in Slovakia. J Appl Microbiol. 2008;104(5):1294–1301. doi:10.1111/j.1365-2672.2007.03653.x
10. Denève C, Bouttier S, Dupuy B, Barbut F, Collignon A, Janoir C. Effects of subinhibitory concentrations of antibiotics on colonization factor expression by moxifloxacin-susceptible and moxifloxacin-resistant Clostridium difficile strains. Antimicrob Agents Chemother. 2009;53(12):5155–5162. doi:10.1128/AAC.00532-09
11. Zhou L, Li T, An J, Liao C, Li N, Wang X. Subminimal inhibitory concentration (sub-MIC) of antibiotic induces electroactive biofilm formation in bioelectrochemical systems. Water Res. 2017;125:280–287. doi:10.1016/j.watres.2017.08.059
12. Kaplan JB. Antibiotic-induced biofilm formation. Int J Artif Organs. 2011;34(9):737–751. doi:10.5301/ijao.5000027
13. Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. Carbapenems: past, present, and future. Antimicrob Agents Chemother. 2011;55(11):4943–4960. doi:10.1128/AAC.00296-11
14. Manchanda V, Sanchaita S, Singh N. Multidrug resistant acinetobacter. J Glob Infect Dis. 2010;2(3):291–304. doi:10.4103/0974-777X.68538
15. Chiotos K, Ross RK, Han JH, Miller M, Gerber JS. Use of carbapenems, polymyxins, and tigecycline in United States Children’s Hospitals, 2010–2014. Open Forum Infect Dis. 2017;4(2):ofx039. doi:10.1093/ofid/ofx039
16. Sato Y, Unno Y, Ubagai T, Ono Y. Sub-minimum inhibitory concentrations of colistin and polymyxin B promote acinetobacter baumannii biofilm formation. PLoS One. 2018;13(3):e0194556. doi:10.1371/journal.pone.0194556
17. Hall GS. Non-fermenting and miscellaneous gram-negative bacilli. In: Mahon CR, Lehman DC, Manuselis G, editors. Textbook of Diagnostic Microbiology. Maryland Heights, MO: Saunders/Elsevier; 2011:482–501.
18. Turton JF, Woodford N, Glover J, Yarde S, Kaufmann ME, Pitt TL. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J Clin Microbiol. 2006;44:2974–2976. doi:10.1128/JCM.01021-06
19. Qi L, Li H, Zhang C, et al. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in acinetobacter baumannii. Front Microbiol. 2016;7:483. doi:10.3389/fmicb.2016.00483
20. Zhang D, Xia J, Xu Y, et al. Biological features of biofilm-forming ability of acinetobacter baumannii strains derived from 121 elderly patients with hospital-acquired pneumonia. Clin Exp Med. 2016;16(1):73–80. doi:10.1007/s10238-014-0333-2
21.
22. Jones RN, Ferraro MJ, Reller LB, Schreckenberger PC, Swenson JM, Sader HS. Multicenter studies of tigecycline disk diffusion susceptibility results for Acinetobacter spp. J Clin Microbiol. 2007;45:227–230. doi:10.1128/JCM.01588-06
23. He X, Lu F, Yuan F, et al. Biofilm formation caused by clinical acinetobacter baumannii isolates is associated with overexpression of the AdeFGH efflux pump. Antimicrob Agents Chemother. 2015;59(8):4817–4825. doi:10.1128/AAC.00877-15
24. López M, Mayer C, Fernández-García L, et al. Quorum sensing network in clinical strains of A. baumannii: aidA is a new quorum quenching enzyme. PLoS One. 2017;12(3):e0174454. doi:10.1371/journal.pone.0174454
25. Smaldone G, Marrone R, Cappiello S, et al. Occurrence of antibiotic resistance in bacteria isolated from seawater organisms caught in Campania Region: preliminary study. BMC Vet Res. 2014;10:161. doi:10.1186/1746-6148-10-1
26. Perez F, Hujer AM, Hujer KM, Decker BK, Rather PN, Bonomo RA. Global challenge of multidrug-resistant acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(10):3471–3484. doi:10.1128/AAC.01464-06
27. Lin MF, Lan CY. Antimicrobial resistance in acinetobacter baumannii: from bench to bedside. World J Clin Cases. 2014;2(12):787–814. doi:10.12998/wjcc.v2.i12.787
28. Jamal S, Al Atrouni A, Rafei R, Dabboussi F, Hamze M, Osman M. Molecular mechanisms of antimicrobial resistance in acinetobacter baumannii, with a special focus on its epidemiology in Lebanon. J Glob Antimicrob Resist. 2018;15(12):154–163. doi:10.1016/j.jgar.2018.05.022
29. Hujer KM, Hujer AM, Hulten EA, et al. Analysis of antibiotic resistance genes in multidrug-resistant acinetobacter sp. isolates from military and civilian patients treated at the Walter Reed Army Medical Center. Antimicrob Agents Chemother. 2006;50:4114–4123. doi:10.1128/AAC.00778-06
30. Singh S, Singh SK, Chowdhury I, Singh R. Understanding the Mechanism of bacterial biofilms resistance to antimicrobial agents. Open Microbiol J. 2017;11:53–62. doi:10.2174/1874285801711010053
31. Wood TK, Knabel SJ, Kwan BW. Bacterial persister cell formation and dormancy. Appl Environ Microbiol. 2013;79(23):7116–7121. doi:10.1128/AEM.02636-13
32. Hawkey PM, Livermore DM. Carbapenem antibiotics for serious infections. BMJ. 2012;344:e3236. doi:10.1136/bmj.e3236
33. Maestre JR, Aguilar L, Mateo M, et al. In vitro interference of tigecycline at subinhibitory concentrations on biofilm development by Enterococcus faecalis. J Antimicrob Chemother. 2012;67(5):1155–1158. doi:10.1093/jac/dks014
34. Chen H, Cao J, Zhou C, Liu H, Zhang X, Zhou T. Biofilm formation restrained by subinhibitory concentrations of tigecyclin in acinetobacter baumannii is associated with downregulation of efflux pumps. Chemotherapy. 2017;62(2):128–133. doi:10.1159/000450537
35. Szczuka E, Jablonska L, Kaznowski A. Effect of subinhibitory concentrations of tigecycline and ciprofloxacin on the expression of biofilm-associated genes and biofilm structure of staphylococcus epidermidis. Microbiology. 2017;163(5):712–718. doi:10.1099/mic.0.000453
36. Weiser J, Henke HA, Hector N, et al. Sub-inhibitory tigecycline concentrations induce extracellular matrix binding protein Embp dependent staphylococcus epidermidis biofilm formation and immune evasion. Int J Med Microbiol. 2016;306(6):471–478. doi:10.1016/j.ijmm.2016.05.015
37. Bhargava N, Sharma P, Capalash N. Quorum sensing in acinetobacter: an emerging pathogen. Crit Rev Microbiol. 2010;36(4):349–360. doi:10.3109/1040841X.2010.512269
38. Kvist M, Hancock V, Klemm P. Inactivation of efflux pumps abolishes bacterial biofilm formation. Appl Environ Microbiol. 2008;74(23):7376–7382. doi:10.1128/AEM.01310-08
39. Yoon EJ, Courvalin P, Grillot-Courvalin C. RND-type efflux pumps in multidrug-resistant clinical isolates of acinetobacter baumannii: major role for AdeABC overexpression and AdeRS mutations. Antimicrob Agents Chemother. 2013;57(7):2989–2995. doi:10.1128/AAC.02556-12
40. Yang YS, Chen HY, Hsu WJ, et al. Overexpression of AdeABC efflux pump associated with tigecycline resistance in clinical acinetobacter nosocomialis isolates. Clin Microbiol Infect. 2018;
41. Damier-Piolle L, Magnet S, Brémont S, Lambert T, Courvalin P. AdeIJK, a resistance-nodulation-cell division pump effluxing multiple antibiotics in acinetobacter baumannii. Antimicrob Agents Chemother. 2008;52(2):557–562. doi:10.1128/AAC.00732-07
42. Tomaras AP, Dorsey CW, Edelmann RE, Actis LA. Attachment to and biofilm formation on abiotic surfaces by acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology. 2003;149(Pt 12):3473–3484. doi:10.1099/mic.0.26541-0
43. Pakharukova N, Tuittila M, Paavilainen S, et al. Structural basis for acinetobacter baumannii biofilm formation. Proc Natl Acad Sci U S A. 2018;115(21):5558–5563. doi:10.1073/pnas.1800961115
44. Choi AH, Slamti L, Avci FY, Pier GB, Maira-Litrán T. The pgaABCD locus of acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J Bacteriol. 2009;191(19):5953–5963. doi:10.1128/JB.00647-09
45. Gaddy JA, Tomaras AP, Actis LA. The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect Immun. 2009;77(8):3150–3160. doi:10.1128/IAI.00096-09
46. Brossard KA, Campagnari AA. The acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect Immun. 2012;80(1):228–233. doi:10.1128/IAI.05913-11
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.