")
Back to Journals » Veterinary Medicine: Research and Reports » Volume 10
Effects of astaxanthin supplementation in healthy and obese dogs
Authors Murai T , Kawasumi K, Tominaga K, Okada Y , Kobayashi M, Arai T
Received 2 September 2018
Accepted for publication 12 December 2018
Published 15 February 2019 Volume 2019:10 Pages 29—35
DOI https://doi.org/10.2147/VMRR.S186202
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Young Lyoo
Tae Murai,1 Koh Kawasumi,1 Kumi Tominaga,2 Yuki Okada,1 Motoo Kobayashi,1 Toshiro Arai1
1Laboratory of Veterinary Biochemistry, School of Veterinary Medicine, Nippon Veterinary and Life Science University, Musashino, Tokyo 180-8602, Japan; 2Research and Development Division, AstaReal Co. Ltd., Minato-ku, Tokyo 105-0011, Japan
Background: Since astaxanthin (ASX) has potent anti-oxidative effects with inhibitory action of lipid peroxidation and singlet oxygen quenching activity, it is widely used as a functional food for keeping good health in human. Obesity is a risk factor for various metabolic disorders. It is characterized by low-grade chronic inflammation based on oxidative stress by excessively produced ROS. From the point of preventive medicine, natural compounds have been proposed as potential therapeutic agents in the prevention of metabolic disorder in companion animals. The purpose of this study is to evaluate the effects of ASX supplementation in healthy and obese dogs.
Materials and methods: Ten healthy beagle dogs and 5 clinically obese dogs were used in this study. The healthy beagle dogs were randomly divided into 2 groups as follows: control and test groups. The test group dogs received ASX supplementation mixed with the food for 6 weeks. Five clinically obese dogs received ASX supplementation for 8 weeks. Metabolites, hormones and enzymes were measured before and after ASX supplementation.
Results: In the healthy dog groups, after 6 weeks, plasma triglyceride (TG) and malondialdehyde concentrations and lactate dehydrogenase (LDH) values significantly decreased in the test group. There was no significant difference in the control group. In clinically obese dogs, plasma TG concentration decreased after 8 weeks of ASX supplementation. Plasma alanine aminotransferase and LDH values clearly decreased in all 5 dogs and 4 dogs out of 5 dogs, respectively.
Conclusion: ASX supplementation (0.3 mg/kg body weight/day) for 6 weeks in healthy dogs and 8 weeks in obese dogs induced the elevation of antioxidant function and of liver function by ameliorating lipid metabolism.
Keywords: astaxanthin, obese dogs, lipid metabolism, anti-oxidative activity, liver function
Introduction
The incidence of obesity and its associated diseases has been increased in dogs and cats as well as in human.1,2 Since obesity causes physical inactivity and oxidative stress related diseases that are induced by obesity-based metabolic syndrome, body weight (BW) reduction is required for obese animals. However, the satisfactory result of weight reduction is limited in the animals with pathological obesity induced by accumulated visceral fat, causing slight systemic inflammation.3 Fat accumulation and oxidative stress impair the function of mitochondria via morphological alteration, increased membrane peroxidation, decreased ATP level, increased ROS production, defective mitochondrial β-oxidation and increased mitochondrial permeabilitization.4 Increase in circulating nonesterified fatty acids (NEFAs) due to excessive accumulated visceral fat is confirmed in obese animals. Fat accumulation triggers free radical production and insults additional inflammation.5 Excess amount of ROS is also produced via accelerated β-oxidation of fatty acids. Such overproduced ROS is attributed to one of the pathogens for obesity and its associated diseases.6 Consequently, some antioxidants appear to be effective to ameliorate obesity conditions in animals.7–9 Experimental studies of mice administered obesity-inducing diet combined with ASX showed anti-diabetic and anti-obesity effects by improved insulin (INS) sensitivities and liver function.10,11 The study showed the suppression of fat tissue weight gain by ASX in a dose-dependent manner.11
Haematococcus pluvialis, known as an important source of natural astaxanthin (ASX), is a freshwater microalga belonging to the family Chlamydomonadaceae. When the alga experiences environmental stress conditions, ASX is created and acts like a force field that protects the nuclear DNA and lipids against UV-induced oxidation.12 ASX (3,3′-dihydroxy-β,β′-carotene-4,4′-dione) is a nontoxic and organic fat-soluble xanthophyll carotenoid. In comparison to other phytochemicals, ASX has previously been reported to possess a significantly greater antioxidant function,13,14 with its antioxidant activities quantified as 10-fold greater than other carotenoids, such as β-carotene, and 100-fold greater than α-tocopherol (vitamin E).15 ASX accumulates in the liver, especially in the microsomal and mitochondrial fractions of the liver tissue.16,17 This substance has been shown to prevent oxidative damage to the liver, improve metabolic profiles, and reduce hepatic inflammation.18–20 From the above, excessive fat accumulation and oxidative stress and liver function are closely related.
In this study, we measured plasma metabolites and hormone concentrations and enzyme activities involved in energy metabolism in healthy and obese dogs with ASX supplementation for several weeks. The purpose of this study was to evaluate the effect of ASX supplementation in obese and healthy dogs.
Materials and methods
Animals
Ten healthy beagle dogs and 5 clinically obese dogs were used in this study. Their body condition score (BCS) was evaluated by the 5-point scale system (1, very thin; 2, underweight; 3, ideal; 4, overweight and 5, obese). BCS of 10 healthy dogs was 3. The average age of them was 2 years (1–3 years), and the average BW was 10.4 kg (9.7–11.1 kg). They were randomly divided into 2 groups: control group (dog no 1–5) and test group (dog no 6–10). As preparation of the study, they were given commercial diet (Nippon Pet Food Co., Ltd., Tokyo, Japan) for 2 months. The nutrient composition of the food is crude protein 21.7%, crude fat 10.1%, crude ash 6.3%, crude fiber 2.7%, linoleic acid 1.72%, moisture 8.7% and nitrogen-free extract 50.5%. All the dogs were kept under controlled conditions and professionally supervised at Narita Animal Science Laboratory Co., Ltd. (Narita, Japan) prior to and during the study period. Ethical approval for this study was obtained from Narita Animal Science Laboratory co., Ltd. Research Animal Ethical Committee (17-C042).
Five clinically obese dogs were recruited from local primary veterinary practice as shown in Table 1. The selected 5 dogs met the following 3 conditions: 1) BW must be more than 20% of its ideal weight; 2) symptoms of chronic disease are properly managed and 3) owner is reliable, and good compliance can be received with the complete consent. Case numbers 1, 2 and 3 have been treated for their chronic diseases by certain therapeutic agents for years, and the clinical signs have been well controlled for long time. Case numbers 4 and 5 have taken no medications.
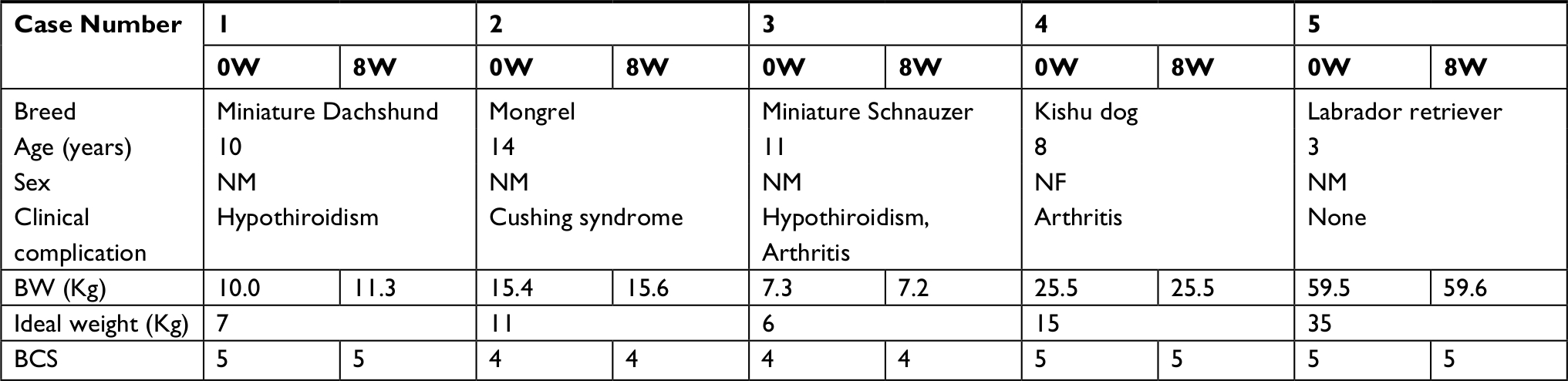 | Table 1 Profiles of clinically overweight and obese dogs Abbreviations: BCS, body condition score; BW, body weight; NF, neutered female; NM, neutered male; W, weeks. |
ASX supplementation
ASX that is H. pluvialis biomass (AstaReal® AW1011; AstaReal Inc., Moses Lake, WA, USA) was used in this study. In the healthy beagle dogs, 5 test group dogs were given 1 dose of 0.3 mg/kg ASX with a meal per day. BW and BCS were measured every week, and the ASX supplement dose was adjusted. For clinical cases, 0.3 mg /kg /day of ASX was given in a single dose or in divided doses with the food on a daily basis.
Blood sampling
Fasting blood samples were collected before initiation of the study, after 6 weeks in healthy dogs, and after 8 weeks in clinically obese dogs, respectively. Collected blood was dispensed in a heparinized tube and centrifuged at 400 × g for 10 minutes at 4°C to collect plasma. Plasma was stored at –80°C until use.
Metabolite, hormone and enzyme analyses
Glucose (GLU), total cholesterol (TC), triglyceride (TG), total protein (TP), blood urea nitrogen (BUN), creatinine (CRE) concentrations and alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP) and lactate dehydrogenase (LDH) activities were measured using an auto-analyzer (JCA-BM2250; JEOL, Tokyo, Japan) at FUJIFILM Monolith Co., Ltd (Tokyo, Japan). Plasma NEFA concentration was measured using the NEFA-C test kit (Wako Pure Chemical Industries, Ltd., Osaka, Japan). Plasma malondialdehyde (MDA) concentration was measured using the NWLSSTM Malondialdehyde assay kit (Northwest Life Science Specialties, LLC, Vancouver, Canada). Plasma INS, adiponectin (ADN) and TNFα were measured by the Rat Insulin ELISA kit (AKRIN-010T; Shibayagi Co., Gumma, Japan), mouse/rat adiponectin ELISA kit (Otsuka Pharmaceutical Co., Ltd., Tokyo, Japan) and TNFα Dog ELISA kit (LS-F1347-1; Life Span Bioscience, Inc, Seattle, WA, USA), respectively.
Statistical analysis
All values were calculated using Microsoft Excel. The data were expressed as mean±standard error (SE). Statistical analysis was performed using the 2-tailed, paired t-test. Statistical significance was designated as P<0.05, and a high level of significance was designated as P<0.01.
Results
Comparisons of biomarker levels in healthy dogs in the control group and the test group are shown in Table 2. The values of plasma TG and MDA concentrations and LDH significantly decreased in the test group dogs (P<0.05, P<0.01), and there was no significant difference in the control group. After 6 weeks, TGs significantly decreased (P<0.05) in the test group, MDA and LDH also significantly decreased in the test group (P<0.01) (Figure 1). Moreover, upon comparing control and test groups in 6 weeks, the values of MDA and LDH significantly decreased (P<0.01) in the test group (P<0.01) (Figure 1). In healthy dogs, with or without ASX supplementation, plasma GLU, TC, NEFA, TP, BUN, CRE, INS, ADN, AST and ALP values showed no significant change before and after 6 weeks of trail (Table 2).
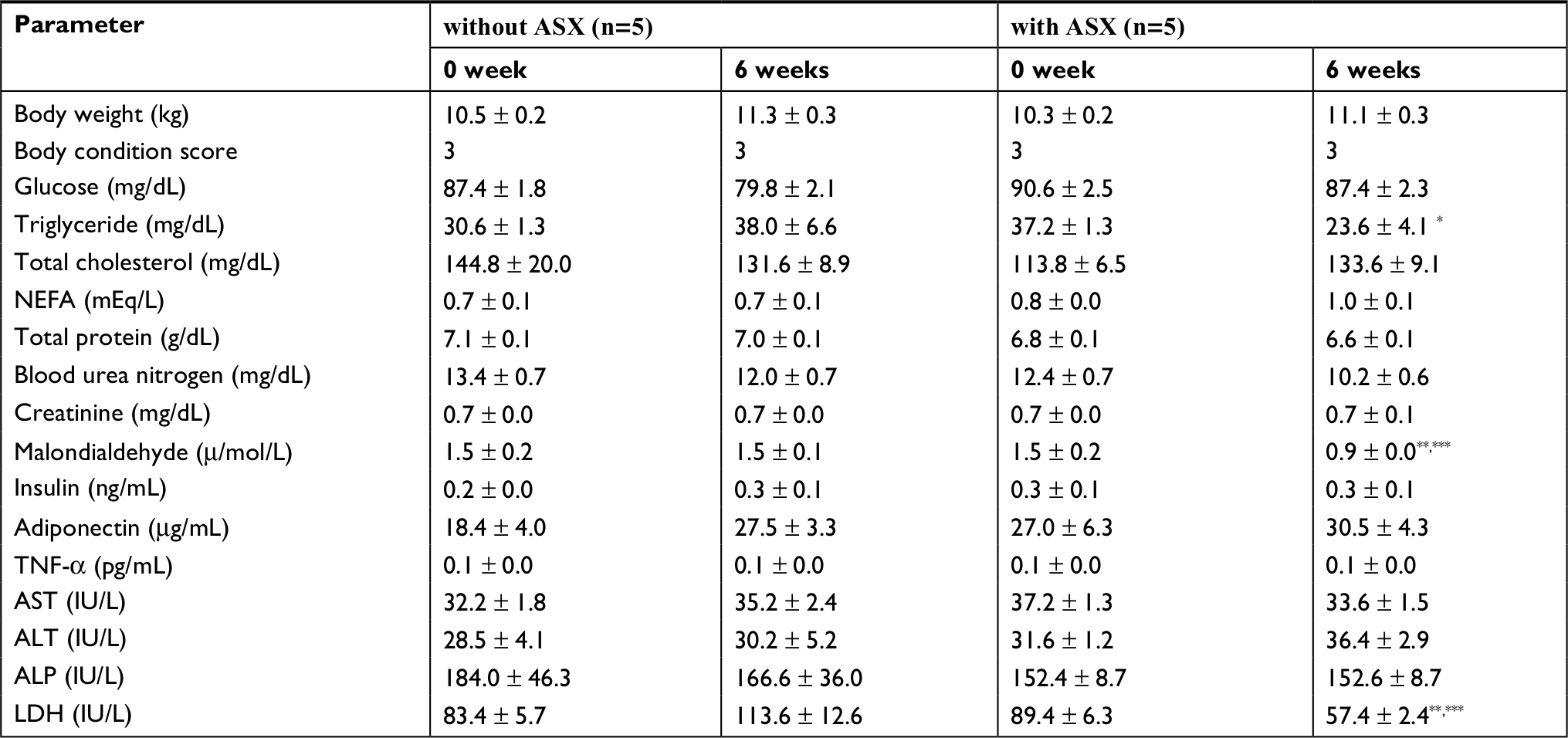 | Table 2 Comparison of biomarkers level of healthy dogs with and without astaxanthin supplementation Notes: Data are presented as the mean ± SE. Statistical significance is indicated by asterisks. *Significantly different (p<0.05) from the value at 0 week in the test group with ASX (paired t-test). **Significantly different (p<0.01) from the value at 0 week in the test group with ASX (paired t-test). ***Significantly different (p<0.01) from the value at 6 weeks of the control group without ASX (paired t-test). Abbreviations: ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; ASX, astaxanthin; LDH, lactate dehydrogenase; NEFA, non-esterified fatty acid; SE, standard error.SE, standard error. |
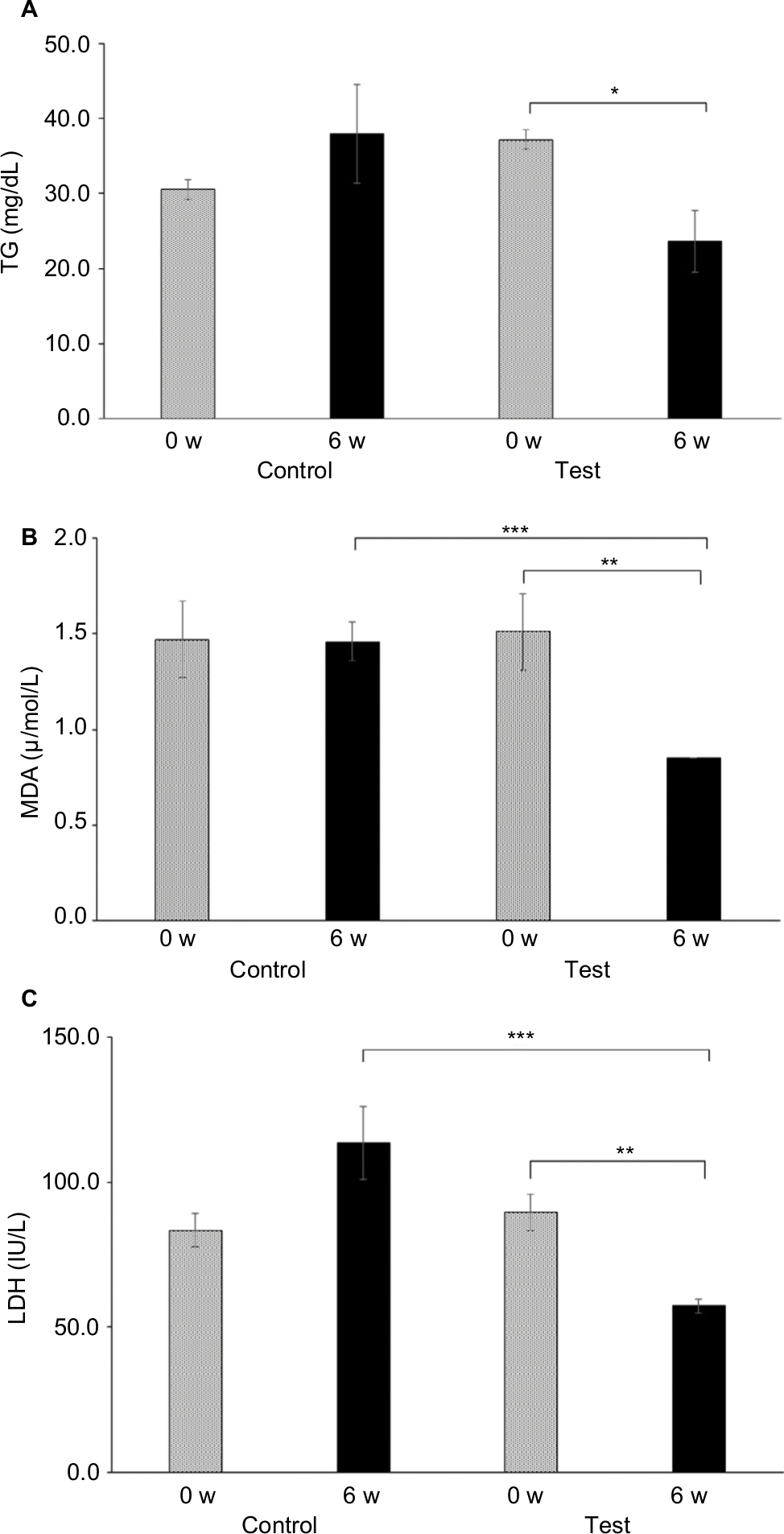 | Figure 1 Comparison of TG, MDA and LDH levels in healthy beagle dogs with or without ASX. (A) TG (mg/dL), (B) MDA (μ/mol/L) and (C) LDH (IU/L). Notes: Statistical significance is indicated by asterisks. *Significantly different (P<0.05) from the value at 0 week in the test group with ASX (paired t-test). **Significantly different (P<0.01) from the value at 0 week in the test group with ASX (paired t-test). ***Significantly different (P<0.01) from the value at 6 weeks of the control group without ASX (paired t-test). Abbreviations: ASX, astaxanthin; LDH, lactate dehydrogenase; MDA, malondialdehyde; TG, triglyceride; W, weeks. |
The results of obese dogs are shown in Table 3. TG values clearly decreased after 8 weeks of ASX supplementation, and ALT and LDH values also remarkably decreased in all 5 dogs and in 4 out of the 5 dogs, respectively. Plasma GLU, TC, NEFA, TP, BUN, CRE, INS, ADN, AST and ALP values showed no major change in clinically obese dogs on ASX supplementation after 8 weeks.
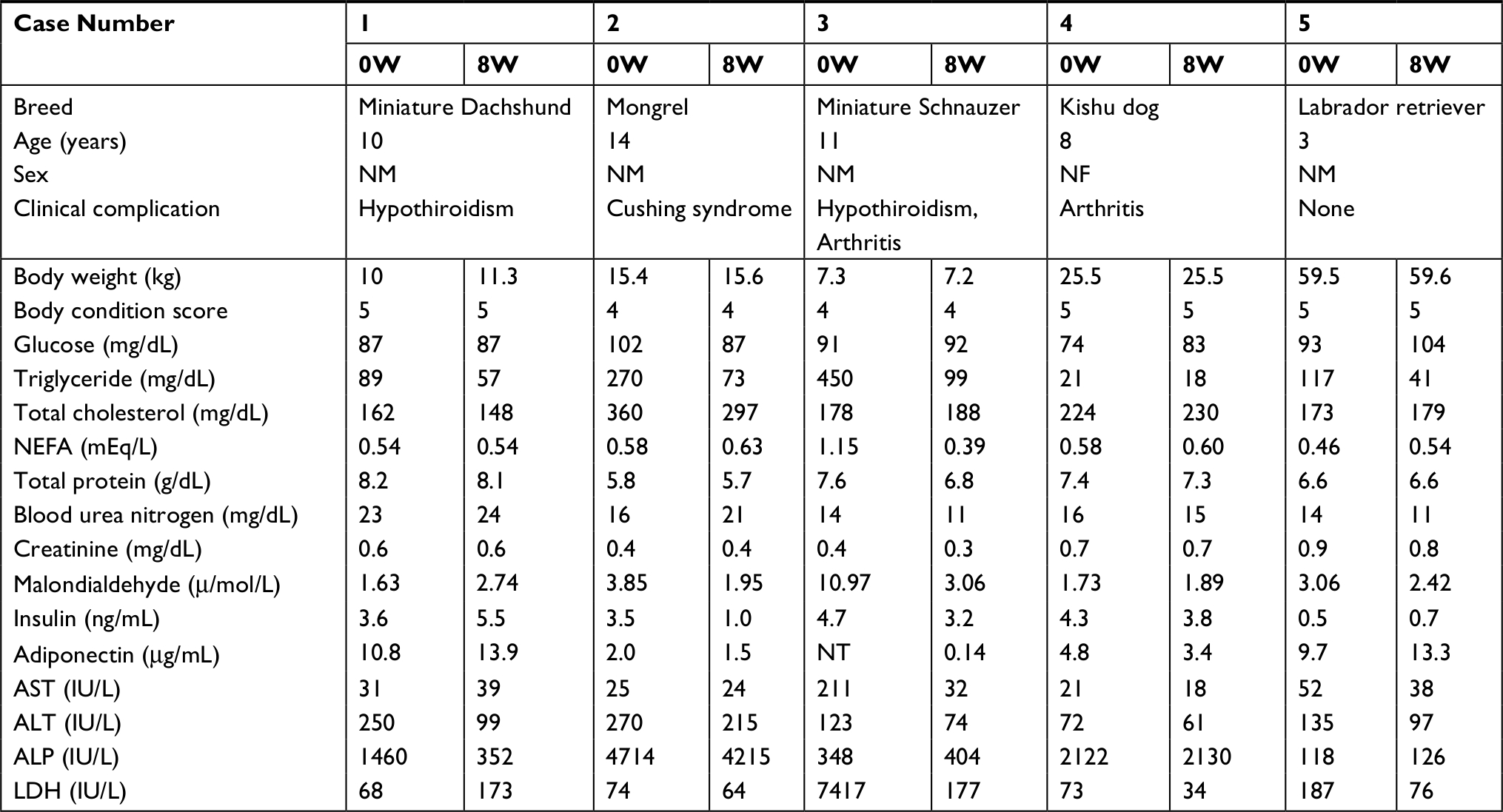 | Table 3 Changes in biomarkers level of clinically overweight and obese dogs with astaxanthin supplementation Abbreviations: ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; ASX, astaxanthin; LDH, lactate dehydrogenase; NEFA, non-esterified fatty acid; NF, neutered female; NM, neutered male; NT, not tested; W, weeks. |
Although statistical comparison is impossible, simple comparison of the TG, ALT, MDA and LDH values of the healthy dog groups with individual obese dogs is done (Figure 2). The change before and after ASX supplementation was more pronounced in obese dogs. In each obese dog, ALT values were always higher than those of healthy dogs regardless of ASX supplementation.
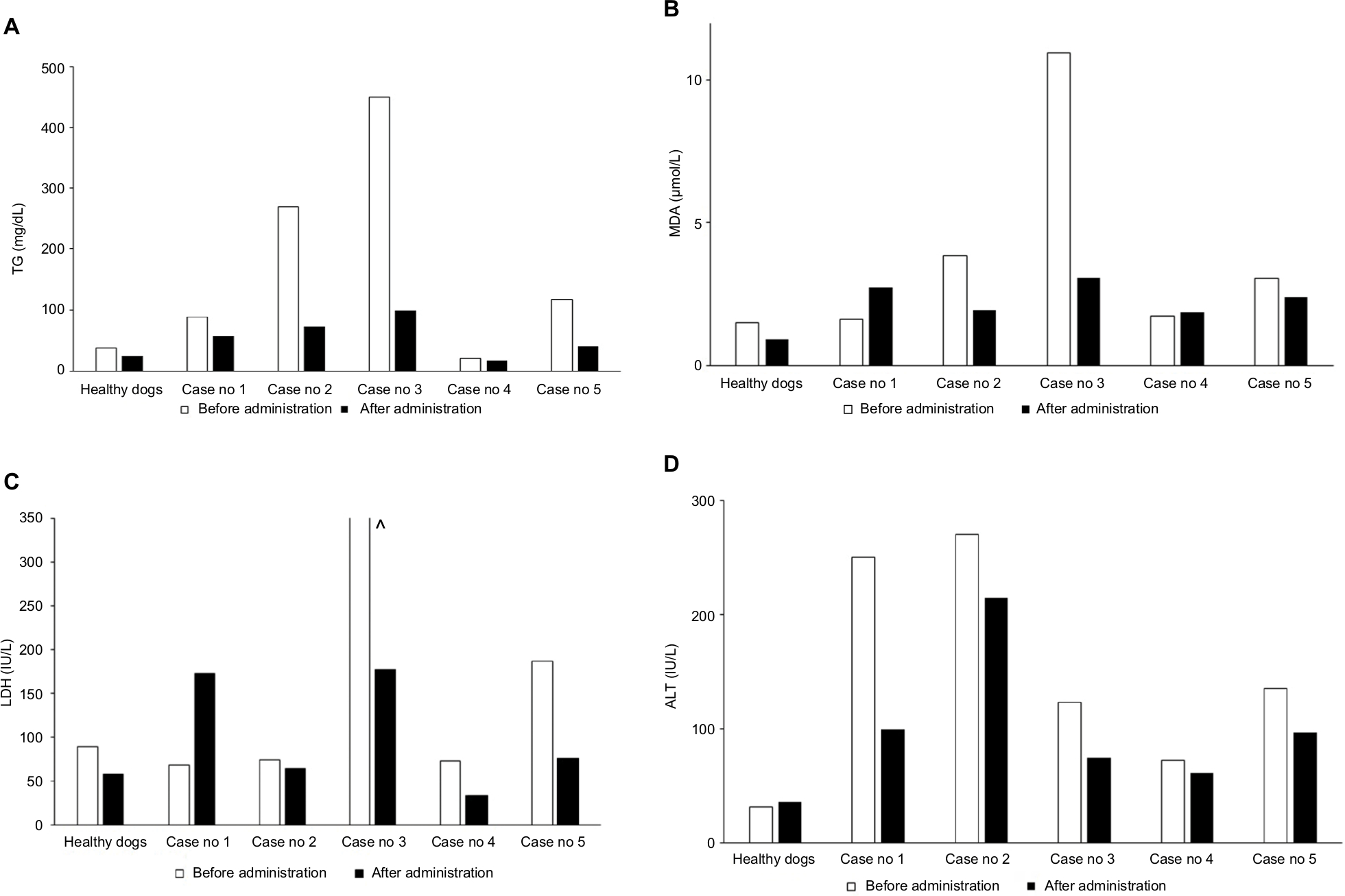 | Figure 2 Comparison of TG, MDA, ALT and LDH levels in healthy and obese dogs before and after ASX supplementation. Notes: (A) TG (mg/dL), (B) MDA (μ/mol/L), (C) LDH (IU/L) and (D) ALT (IU/L). ^The value exceeded the display range. Abbreviations: ALT, alanine aminotransferase; ASX, astaxanthin; LDH, lactate dehydrogenase; MDA, malondialdehyde; TG, triglyceride; W, weeks. |
BW and BCS showed no changes after ASX supplementation.
Discussion
Obesity is characterized by low-grade chronic inflammation.21 This continuous inflammation due to obesity induces severe metabolic disorders such as hypertension, vascular disorders, diabetes mellitus and others.22 Increased circulating NEFAs from accumulated visceral fat cause inflammation and INS resistance by directly activating plasma membrane receptors, such as toll-like receptor 4,23 followed by elevation of inflammatory reaction via NF-kβ.24,25 On the other hand, excessive amount of NEFA enhances overproduction of ROS in the process of β-oxidation of fatty acids in the mitochondria of various tissues. Overproduction of ROS induces oxidative stress. Consequently, systemic inflammatory components were confirmed in obesity.26,27 Adipose tissue, a population of adipocytes, not only acts as an energy reservoir but also has physiological activities such as angiogenesis and wound healing, and adipocytes produce and secrete adipokines involved in energy metabolism.28 Accumulated visceral fat in obese animals induces high concentrations of plasma NEFA, circulating C-reactive protein (CRP) and MDA.29,30 Excessive amount of visceral fat is suspected as a contributing factor to various metabolic disorders in obese animals. From the above mentioned findings, antioxidant substances are considered to decrease oxidative stress and to ameliorate metabolic disorders caused due to obesity.
Obese animals encountered in veterinary practice are individually unique in their background. In general, dietary supplements and functional foods are often recommended for animals that require the health care intervention from veterinary professionals, such as aging, chronic diseases and critical obesity as well. Considering these circumstances, obese dogs participated in this experiment were various age, sex, and species.
In this study, ASX supplementation showed clear antioxidative effects in both healthy and obese dogs. In the healthy dogs, after ASX supplementation for 6 weeks, plasma TG and oxidative stress biomarker MDA concentrations decreased significantly. ASX revealed antioxidant activity in healthy dogs. In the same way, plasma TG, MDA, ALT and LDH values decreased, especially, TG and ALT values remarkably decreased. Antioxidant effects of ASX supplementation in obese dogs are more apparent than those in the healthy dogs. Since those positive effects were observed in all obese dogs with different underlying diseases, it is considered that ASX supplementation could be effective on the antioxidant activity and also improve the hepatic function.
ASX prevents diseases in heart and kidneys31 and liver20 from oxidative stress, in addition ASX can decrease plasma MDA concentrations and improve the pathological signs of animal diabetic nephropathy.32 In this study, ASX supplementation indicated to be effective on elevation of antioxidant function and on amelioration of metabolic functions in liver. Long-term intake of ASX inhibits the elevations in BW and adipose tissue weight caused by a high-fat diet in mice tested for 60 days.11 Chronic ASX administration significantly improve increased body weight, hyperglycemia, hyperinsulinemia and increased plasma levels of TNFα and IL-6 observed in the study of obese model mice.33 However, ASX supplementation does not influence BW reduction in obese dogs in this study. In the mice experiments, ASX was given in relatively high dose such as 6, 12 and 30 mg/kg for 60 days. The dose of ASX used in this experiment was set up based on the standard recommended dose to human. Considering the lifespan of the mouse, the duration of 60 days for the mouse corresponds to 1 year or more in the dog. We speculate that longer administration period in obese dogs of more than 2 months could bring the farther positive effect. ASX improved oxidative stress biomarkers by suppressing lipid peroxidation and stimulating the liver function. However, weight reduction effect could not be achieved only by supplementation of ASX to obese dogs.
Moreover, safety of ASX supplementation is advocated by EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP) in previous report.34 Considering that ASX has wide safety range,35 continuous supplementation of ASX seems to be effective to prevent the prevalence of visceral fat-type obesity. Further studies with different doses of ASX supplementation are necessary to clarify the usefulness of ASX as an antioxidant supplement in a large number of dogs with different severities of obesity.
Conclusion
ASX supplementation (0.3 mg/kg BW/day) in food for 6 weeks in healthy dogs and for 8 weeks in obese dogs effectively activated antioxidant function and liver function followed by improved lipid metabolism.
Acknowledgments
The authors thank the staff of the Narita Animal Science Laboratory Co., Ltd for their reliable animal monitoring throughout the study period, and thank the owners of all the dogs for allowing them to participate in this experiment. The authors also thank Dr Atsuhiko Hasegawa and Dr Kohei Suruga for their advice on this manuscript.
Disclosure
Kumi Tominaga is a resercher working in AstaReal Co.Ltd. The authors report no other conflicts of interest in this work.
References
Edney AT, Smith PM. Study of obesity in dogs visiting veterinary practices in the United Kingdom. Vet Rec. 1986;118(14):391–396. | ||
Lund E, Armstrong P, Kirk C, Klausner J. Prevalence and risk factors for obesity in adult dogs from private US veterinary practices. Int J Appl Res Vet Med. 2006;4:177–186. | ||
Okada Y, Kobayashi M, Sawamura M, Arai T. Comparison of visceral fat accumulation and metabolome markers among cats of varying BCS and novel classification of feline obesity and metabolic syndrome. Front Vet Sci. 2017;4(22):17. | ||
Kim S, Kim H. Inhibitory effect of astaxanthin on oxidative stress-induced mitochondrial dysfunction-a mini-review. Nutrients. 2018;10(9):pii: E1137. | ||
Matteoni CA, Younossi ZM, Gramlich T, Boparai N, Liu YC, Mccullough AJ. Nonalcoholic fatty liver disease: a spectrum of clinical and pathological severity. Gastroenterology. 1999;116(6):1413–1419. | ||
Natoli R, Fernando N, Dahlenburg T, et al. Obesity-induced metabolic disturbance drives oxidative stress and complement activation in the retinal environment. Mol Vis. 2018;24:201–217. | ||
Ribeiro MR, Lima RPA, Lisboa JVC. Influence of the C677T polymorphism of the MTHFR gene on oxidative stress in women with overweight or obesity: response to a dietary folate intervention. J Am Coll Nutr. 2018;27:1–8. | ||
Kim K, Chung MH, Park S, et al. ER stress attenuation by Aloe-derived polysaccharides in the protection of pancreatic β-cells from free fatty acid-induced lipotoxicity. Biochem Biophys Res Commun. 2018;500(3):797–803. | ||
Otton R, Bolin AP, Ferreira LT, Marinovic MP, Rocha ALS, Mori MA. Polyphenol-rich green tea extract improves adipose tissue metabolism by down-regulating miR-335 expression and mitigating insulin resistance and inflammation. J Nutr Biochem. 2018;57:170–179. | ||
Bhuvaneswari S, Arunkumar E, Viswanathan P, Anuradha CV. Astaxanthin restricts weight gain, promotes insulin sensitivity and curtails fatty liver disease in mice fed a obesity-promoting diet. Process Biochem. 2010;45(8):1406–1414. | ||
Ikeuchi M, Koyama T, Takahashi J, Yazawa K. Effects of astaxanthin in obese mice fed a high-fat diet. Biosci Biotechnol Biochem. 2007;71(4):893–899. | ||
Sarada R, Tripathi U, Ravishankar GA. Influence of stress on astaxanthin production in Haematococcus pluvialis grown under different culture conditions. Process Biochemistry. 2002;37(6):623–627. | ||
Kurashige M, Okimasu E, Inoue M, Utsumi K. Inhibition of oxidative injury of biological membranes by astaxanthin. Physiol Chem Phys Med NMR. 1990;22(1):27–38. | ||
Shimidzu N, Goto M, Miki W. Carotenoids as singlet oxygen quenchers in marine organisms. Fish Sci. 1996;62(1):134–137. | ||
Miki W. Biological functions and activities of animal carotenoids. Pure Appl Chem. 1991;63(1):141–146. | ||
Takahashi K, Watanabe M, Takimoto T, Akiba Y. Uptake and distribution of astaxanthin in several tissues and plasma lipoproteins in male broiler chickens fed a yeast (Phaffia rhodozyma) with a high concentration of astaxanthin. Br Poult Sci. 2004;45(1):133–138. | ||
Showalter LA, Weinman SA, Østerlie M, Lockwood SF. Plasma appearance and tissue accumulation of non-esterified, free astaxanthin in C57BL/6 mice after oral dosing of a disodium disuccinate diester of astaxanthin (Heptax). Comp Biochem Physiol C Toxicol Pharmacol. 2004;137(3):227–236. | ||
Curek GD, Cort A, Yucel G, et al. Effect of astaxanthin on hepatocellular injury following ischemia/reperfusion. Toxicology. 2010;267(1–3):147–153. | ||
Ferramosca, A, Di Giacomo M, Zara V. Antioxidant dietary approach in treatment of fatty liver: new insights and updates. World J Gastroenterol. 2017;23(23):4146–4157. | ||
Islam MA, Al Mamun MA, Faruk M. Astaxanthin ameliorates hepatic damage and oxidative stress in carbon tetrachloride-administered rats. Pharmacogn Res. 2017;9:S84–S91. | ||
Defronzo RA. Insulin resistance, lipotoxicity, type 2 diabetes and atherosclerosis: the missing links. The Claude Bernard Lecture 2009. Diabetologia. 2010;53(7):1270–1287. | ||
Prado NJ, Ferder L, Manucha W, Diez ER, Raul Diez E. Anti-inflammatory effects of melatonin in obesity and hypertension. Curr Hypertens Rep. 2018;20(5):45. | ||
Reyna SM, Ghosh S, Tantiwong P, et al. Elevated toll-like receptor 4 expression and signaling in muscle from insulin-resistant subjects. Diabetes. 2008;57(10):2595–2602. | ||
Ghanim H, Garg R, Aljada A, et al. Suppression of nuclear factor-kappaB and stimulation of inhibitor kappaB by troglitazone: evidence for an anti-inflammatory effect and a potential antiatherosclerotic effect in the obese. J Clin Endocrinol Metab. 2001;86(3):1306–1312. | ||
Evans JL, Goldfine ID, Maddux BA, Grodsky GM. Oxidative stress and stress-activated signaling pathways: a unifying hypothesis of type 2 diabetes. Endocr Rev. 2002;23(5):599–622. | ||
Dorresteijn JA, Visseren FL, Spiering W. Mechanisms linking obesity to hypertension. Obes Rev. 2012;13(1):17–26. | ||
Renna NF, Diez ER, Lembo C, Miatello RM. Role of Cox-2 in vascular inflammation: an experimental model of metabolic syndrome. Mediators Inflamm. 2013;2013:513251. | ||
Huby AC, Antonova G, Groenendyk J, et al. Adipocyte-derived hormone leptin is a direct regulator of aldosterone secretion, which promotes endothelial dysfunction and cardiac fibrosis. Circulation. 2015;132(22):2134–2145. | ||
Okamura A, Watanabe M, Fukudome I, et al. Relationship between visceral obesity and postoperative inflammatory response following minimally invasive esophagectomy. World J Surg. 2018;42(11):3651–3657. | ||
Ormazabal P, Scazzocchio B, Vari R, et al. Effect of protocatechuic acid on insulin responsiveness and inflammation in visceral adipose tissue from obese individuals: possible role for PTP1B. Int J Obes. 2018;42(12):2012–2021. | ||
Alam MN, Hossain MM, Rahman MM, et al. Astaxanthin prevented oxidative stress in heart and kidneys of isoproterenol-administered aged rats. J Diet Suppl. 2018;15(1):42–54. | ||
Zhu X, Chen Y, Chen Q, Yang H, Xie X. Astaxanthin promotes Nrf2/ARE signaling to alleviate renal fibronectin and collagen IV accumulation in diabetic rats. J Diabetes Res. 2018;article ID 6730315. | ||
Arunkumar E, Bhuvaneswari S, Anuradha CV. An intervention study in obese mice with astaxanthin, a marine carotenoid: effects on insulin signaling and pro-inflammatory cytokines. Food Funct. 2012;3(2):120–126. | ||
European Food Safety Authority. Scientific opinion on the safety and efficacy of synthetic astaxanthin as feed additive for salmon and trout, other fish, ornamental fish, crusrtaceans and ornamental birds. EFSAJ. 2014;12(6):3724. Available from: https://efsa.onlinelibrary.wiley.com/doi/pdf/10.2903/j.efsa.2014.3724. Accessed January 15, 2019. | ||
Brown DR, Gough LA, Deb SK, Sparks SA, Mcnaughton LR. Astaxanthin in exercise metabolism, performance and recovery: a review. Front Nutr. 2018;4:76. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.