")
Back to Journals » Journal of Pain Research » Volume 12
Effects of an intrathecal TRPV1 antagonist, SB366791, on morphine-induced itch, body temperature, and antinociception in mice
Authors Sakakibara S, Imamachi N , Sakakihara M, Katsube Y, Hattori M, Saito Y
Received 28 May 2019
Accepted for publication 6 August 2019
Published 28 August 2019 Volume 2019:12 Pages 2629—2636
DOI https://doi.org/10.2147/JPR.S217439
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Satoshi Sakakibara, Noritaka Imamachi, Manabu Sakakihara, Yukiko Katsube, Mai Hattori, Yoji Saito
Department of Anesthesiology, Shimane University Faculty of Medicine, Shimane, Japan
Correspondence: Noritaka Imamachi
Department of Anesthesiology, Shimane University Faculty of Medicine, 89-1 Enyacho Izumo, Shimane
693-8501, Japan
Tel +81 85 320 2295
Fax +81 85 320 2292
Email [email protected]
Purpose: Transient receptor potential vanilloid 1 (TRPV1) not only is activated by multiple stimuli but also is involved with histamine-induced itch. The effects of TRPV1 on morphine-induced itch are unknown. We examined the effects of intrathecal administration of TRPV1 antagonist on morphine-induced itch, body temperature, and antinociception for mice.
Methods: Each C57/BL6j mouse was intrathecally administered with one of the following solutions: morphine, SB366791 (as the TRPV1 antagonist), morphine + SB366791, saline, or vehicle. For each mouse, each instance of observed scratching behavior was counted, the body temperature was measured, and the nociceptive threshold was determined using the tail-immersion test.
Results: SB366791 dose-dependently reduced the scratching behavior induced by the administration of morphine. SB366791 and the morphine + SB366791 groups did not manifest an increase in body temperature. Antinociceptive effects were observed to occur dose-dependently for morphine but not for SB366791. Compared with morphine alone, the administration of morphine + SB366791 did not reduce significant antinociceptive effects.
Conclusion: We propose that an intrathecal TRPV1 antagonist, SB366791, reduced morphine-induced itch without causing hyperthermia and did not suppress morphine-induced antinociception for mice.
Keywords: pruritus, opioid, TRPV1, nociception, hyperthermia
Introduction
Although morphine is indispensable and widely used for pain management, it presents some adverse effects, including nausea, vomiting, and respiratory depression, with itch being a major effect. The frequency and severity of morphine-induced itch vary with the dose and route of administration.1 The incidence of neuraxial morphine-induced itch is sometimes severe, ranging from 30% to 90% in patients receiving morphine.1 Previously, histamine antagonists have failed to suppress morphine-induced itch.2,3 Conversely, opioid receptor antagonists can block morphine-induced itch but are not clinically available because they inhibit opioids’ antinociceptive effects. Therefore, there is no standard treatment for morphine-induced itch.
Transient receptor potential vanilloid 1 (TRPV1) is the first cloned TRP family member channel.4 Recently, some TRP channels were considered as the molecular sensors of chemical, thermal, and mechanical noxious stimuli that evoke pain and itch.5 TRPV1 is activated by multiple stimuli, including capsaicin, heat, pH, endocannabinoids, and endogenous lipids.6 TRPV1 antagonists have shown antinociceptive effects in several pain models including inflammatory, cancer, and postoperative pain model.7–9 We previously demonstrated the important role of TRPV1 on histamine-induced itch.10 However, hyperthermia is a major side effect of the systemic administration of TRPV1 antagonists.11–13
The effects of an intrathecal TRPV1 antagonist on morphine-induced itch without causing hyperthermia remain unknown. Herein, we examined the effects of an intrathecal TRPV1 antagonist on morphine-induced itch, body temperature, and antinociception for mice.
Materials and methods
Animals
All experiments were approved by the Animal Care and Use Committee of Shimane university (No. IZ 27–117, 139) and conducted according to the regulations for animal experiment at Shimane university. The studies were conducted on male C57/BL6j mice (21–27 g), which were housed in light (lit from 8:00 to 20:00) and temperature-controlled (23–25°C) environment. Food and water were freely available.
Drugs
Morphine hydrochloride (Takeda Pharmaceutical, Tokyo, Japan) was dissolved in physiologic saline. Morphine hydrochloride was dissolved in vehicle consisting of ethanol and saline at a 1:9 dilution when we studied the effect of intrathecal SB366791 on morphine-induced itch. The dosage of morphine was determined based on the method described in a previous study.14 SB366791 (Wako Pure Chemical Industries, Osaka, Japan), the TRPV1 antagonist to be used in this study, was dissolved in a vehicle, and its dosage was determined as previously described.15 SB366791 cannot be dissolved in physiologic saline. Therefore, we used the vehicle.
Intrathecal injection method
Lumbar punctures were performed as previously described.16 The experiments were performed only after each administrator had achieved a success rate of >90% in intrathecal injection training sessions, which involved the administration of 5 μL lidocaine (2%). The volume of each drug was 5 μL.
Scratching behavior
This experiment was conducted from 9:00 to 16:00. Scratching behavior was counted as previously described.17 Two days before starting this study, the mice were habituated each day under the same conditions of observation. After acclimation for 30 mins, each mouse was intrathecally administered with one of the following solutions: morphine (0.1, 0.3, or 1.0 nmol) dissolved in saline, 0.3-nmol morphine dissolved in vehicle, SB366791 0.1 nmol, 0.3-nmol morphine + SB366791 (0.01, 0.03, or 0.1 nmol), saline, or vehicle. After intrathecal solution administration, the scratching behavior of each mouse was videotaped for 60 mins under unmanned conditions. The temporal and total numbers of scratches performed by the individual mouse’s hind paws during the first 60 mins after intrathecal injection were counted. This test was performed in a blinded manner.
Observation of body temperature
This experiment was performed from 9:00 to 16:00. The mice were habituated every day for 2 days under the same conditions of observation. Each mouse received an intrathecal injection of one of the following agents: 0.3-nmol morphine dissolved in saline, 0.1-nmol SB, 0.3-nmol morphine +0.1-nmol SB, saline, or vehicle. The body temperature was measured using an infrared thermometer (Ubi-x, CISE 99TS, Tokyo, Japan) on the back of each mouse (the area shaved for intrathecal injection)18 at 10, 20, 30, 40, 50, and 60 mins after the performance of the intrathecal injection. This test was performed in a blinded manner.
Tail-immersion test
This experiment was performed from 9:00 to 17:00. The mice were habituated every day for 2 days under the same conditions of observation. The nociceptive threshold was determined as previously described.14 The tail of each mouse was submerged in water at 48.0±0.5°C, and the time to tail withdrawal was observed. On the testing day, each mouse was gently held in a soft cloth, and its tail was immersed in the heated water before and after 5, 15, 30, 60, 90, 120, and 150 mins of the performance of the intrathecal injection of one of the following solutions: morphine (0.1, 0.3, or 1.0 nmol) dissolved in saline, SB366791 (0.01, 0.03, or 0.1 nmol), 0.3-nmol morphine +0.1-nmol SB366791, saline, or vehicle. If a mouse did not remove its tail from the heated water, a 20 s cut-off was used to prevent tissue damage and an upper limit of latency of 20 s was recorded. This test was performed in a blinded manner.
Statistical analysis
The number of scratches, body temperature, and latencies of the tail-immersion test are presented as the mean ± standard error of the mean. The total number of scratches was analyzed using one-way analysis of variance (ANOVA), followed by Scheffe’s test. Changes in the number of scratches, body temperature, and tail-withdrawal latencies were analyzed using two-way repeated-measures ANOVA, followed by Scheffe’s test. Statistical analyses were performed by using Stat-View 5.0 (Abacus Concepts, Inc. Berkley, CA, USA). Differences were considered significant at P<0.05.
Results
The total number of mice used in all of the experiments was 156; the number of mice observed was 62 for scratching behavior, 40 for body temperature, and 54 for the tail-immersion test. We used eight mice for vehicle group of scratching behavior and all groups of body temperature. We used six mice for the other groups of scratching behavior and all groups of the tail-immersion test. No mouse showed any neurologic deficits resulting from intrathecal injection. No mouse was excluded from this study.
Scratching behavior
In the saline, 0.1-nmol morphine, and 1.0-nmol morphine groups, the total numbers of scratches were 12.7±1.7, 33.8±13.2, and 66.2±19.5, respectively. The number of scratches was significantly higher in the 0.3-nmol morphine group than in the saline group (127.5±23.2; P=0.001; F3,20=9.2) (Figure 1A). The peak of scratching behavior was seen at 10–20 mins after undergoing an intrathecal administration in the mice of the 0.3-nmol morphine group and at 0–10 mins after undergoing an intrathecal administration in the mice of the 1.0-nmol morphine group; the number of scratches decreased after these times in all of the groups (Figure 1B).
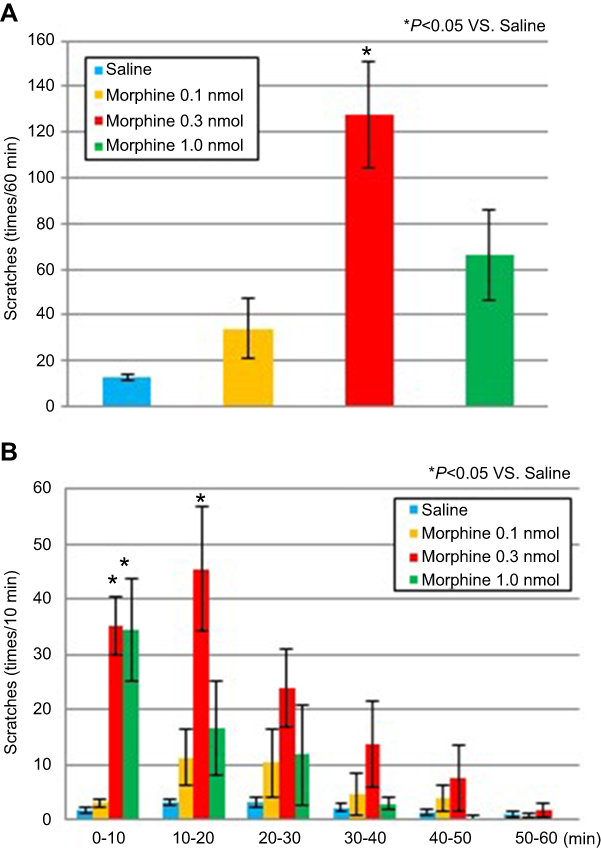 |
Figure 1 Scratching behavior induced by the intrathecal administration of morphine. Notes: (A) The total number of scratches was significantly higher for the 0.3-nmol morphine group than for the saline group (P=0.001). (B) The time course of scratching behavior after the administration of saline or morphine (0.1, 0.3, or 1.0 nmol). The peak of scratching behavior was at 10–20 mins after undergoing an intrathecal administration for the mice of the 0.3-nmol morphine group and at 0–10 mins after undergoing an intrathecal administration for the mice of the 1.0-nmol morphine group. Data are presented as the mean ± standard error of the mean (SEM) for the 6–8 mice in each group. *P<0.05 compared with the saline group. One-way analysis of variance (ANOVA), followed by Scheffe’s test in (A), two-way repeated-measures ANOVA, followed by Scheffe’s test in (B). |
The scratching behavior of the 0.1-nmol SB366791 group (10.2±1.0) was not significantly different from that of the vehicle group (9.8±2.2; P>0.99) (Figure 2). Scratching behavior was significantly increased for the 0.3-nmol morphine dissolved in vehicle (0.3-nmol morphine + vehicle) group (122.7±26.7) in comparison to that of the vehicle group (P=0.002). In contrast, compared with the vehicle group, the 0.3-nmol morphine +0.01-nmol, 0.03-nmol, and 0.1-nmol SB366791 groups did not exhibit significant increases in the number of scratches (68.3±12.5; P=0.12, 42.5±18.3; P=0.71, 29.2±10.7; P=0.95, respectively; F5,32=9.3). SB366791 dose-dependently reduced the scratching behavior that was induced by morphine at 0.3 nmol. In addition, the total numbers of scratches for the groups receiving 0.3-nmol morphine +0.03-nmol or 0.1-nmol SB366791 were significantly decreased compared with that of scratches of the 0.3-nmol morphine + vehicle group (P=0.02 and P=0.004, respectively) (Figure 2).
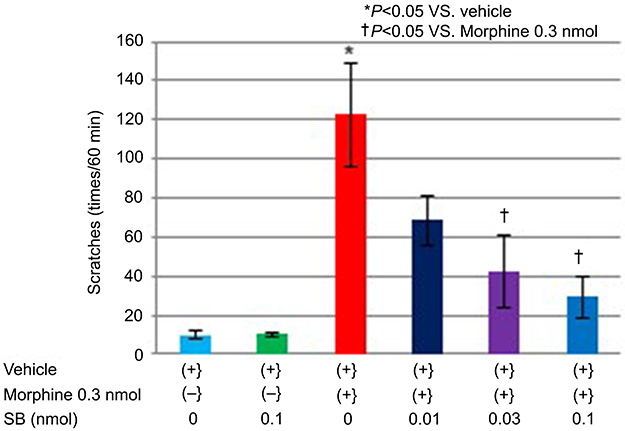 |
Figure 2 Scratching behavior induced by the intrathecal administration of morphine or/and SB366791. Notes: The total numbers of scratches for the groups receiving 0.3-nmol morphine +0.03-nmol or 0.1-nmol SB366791 were significantly decreased compared with the total number of scratches of the 0.3-nmol morphine group (P=0.02 and P=0.004, respectively). Data are presented as the mean ± SEM for the six mice in each group. *P<0.05 compared with the vehicle group. †P<0.05 compared with the 0.3-nmol morphine dissolved in vehicle group. One-way analysis of variance (ANOVA), followed by Scheffe’s test.Abbreviation: SB, SB366791. |
Body temperature
The body temperature of the mice ranged from 35.8°C to 36.2°C, among all of the groups for 60 mins after the intrathecal injection (P=0.087). Compared with the body-temperature measurements for the vehicle group, those of the SB366791 group and the morphine + SB366791 group did not manifest an increase in body temperature (Figure 3).
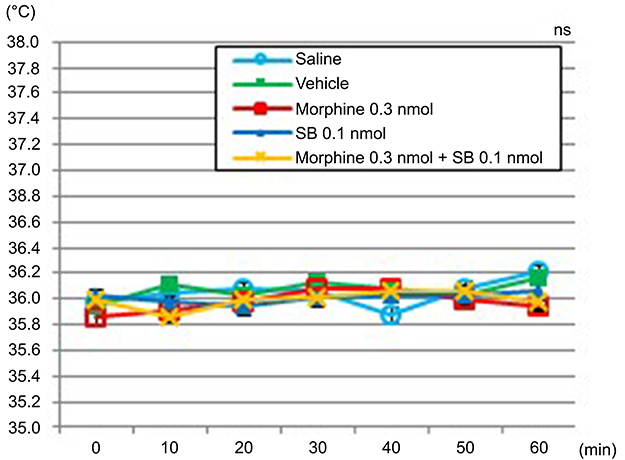 |
Figure 3 Time course of body temperature over the first 60 mins following the administration of saline, vehicle, 0.3-nmol morphine, 0.1-nmol SB366791, or 0.3-nmol morphine +0.1-nmol SB366791. Notes: The body temperature of the mice ranged from 35.8°C to 36.2°C in all groups for 60 mins after intrathecal injection (P=0.087). Data are presented as the mean ± SEM for the six mice in each group. Two-way repeated-measures analysis of variance. Abbreviations: SB, SB366791; ns, not significant. |
Tail-immersion test
Intrathecal morphine dose-dependently produced antinociceptive effects. The latency of withdrawal of the tail following tail immersion in heated water was significantly prolonged from 5 to 15 mins after administration for the 0.3-nmol morphine group (P=0.007 and P=0.0423, respectively) and from 5 to 90 mins and to 150 mins after administration for the 1.0-nmol morphine group, compared with the latency observed for the saline group (P=0.0001–0.044) (Figure 4A). Intrathecally administered SB366791 did not produce thermal antinociceptive effects, in comparison with the effects observed for the vehicle group (P=0.95) (Figure 4B). The latency was significantly prolonged from 5 to 120 mins after administration for the 0.3-nmol morphine group, compared with that for the saline group (P=0.001–0.015) (Figure 4C). Morphine at 0.3 nmol + SB366791 at 0.1 nmol produced antinociceptive effects corresponding to a latency increase from 5 to 120 mins, compared with the effects observed for the vehicle group (P<0.0001–0.025). Morphine at 0.3 nmol + SB366791 at 0.1 nmol did not produce significant thermal antinociceptive effects, compared with the effects observed for the 0.3-nmol morphine group (P=0.21–0.99) (Figure 4C).
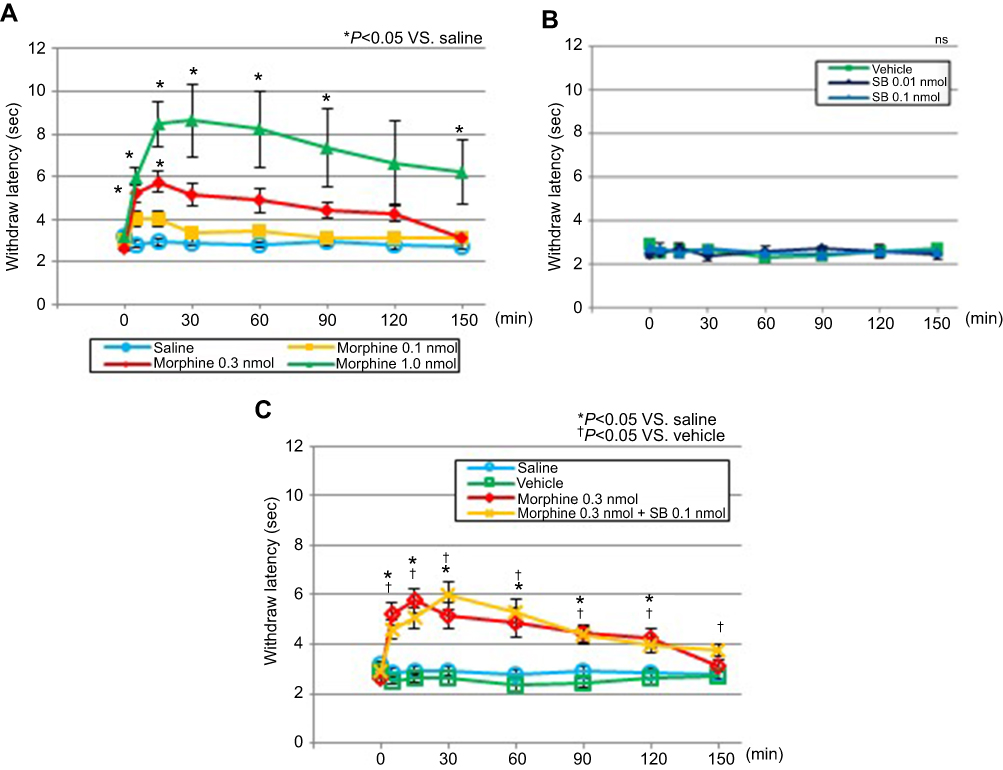 |
Figure 4 Antinociceptive effects measured by the tail-immersion test. Antinociceptive effects after the administration of (A) saline or morphine (0.1, 0.3, or 1.0 nmol); (B) vehicle or SB (0.01 nmol, or 0.1 nmol) and (C) saline, vehicle, 0.3-nmol morphine, or 0.3-nmol morphine +0.1-nmol SB. Notes: Antinociceptive effects were observed to occur dose-dependently for morphine but not for SB366791 (P=0.95). Compared with morphine alone, the administration of morphine + SB366791 did not reduce significant antinociceptive effects (P=0.21–0.99). Data are presented as the mean ± SEM for the eight mice in each group. *P<0.05 compared with the saline group. †P<0.05 compared with the vehicle group. Two-way repeated-measures analysis of variance followed by Scheffe’s test. Abbreviations: SB, SB366791; ns, not significant. |
Discussion
Three main findings were observed in this study. First, intrathecally administered SB366791, which is a TRPV1 antagonist, dose-dependently inhibited morphine-induced itch following an intrathecal administration in mice. Second, intrathecal SB366791 did not raise body temperature. Third, intrathecal SB366791 did not suppress morphine-induced antinociception of a thermal stimulus. Therefore, an intrathecal TRPV1 antagonist, SB366791, produced potent antipruritic effects for intrathecal morphine-induced itch, without serious adverse effects such as hyperthermia.
In clinical studies, several drugs have been used to treat morphine-induced itch.14,19–23 5-HT-receptor antagonists may relieve morphine-induced itch, although the results of clinical trials are debatable.19,20 Some reports have indicated that a subanesthetic dose of propofol resulted in antipruritic effects on morphine-induced itch; however, the efficacy of propofol for morphine-induced itch remains controversial.21,22 In both basic research and clinical studies, the administration of kappa-opioid receptor agonists was found to have antipruritic effects on morphine-induced itch,14,23 although the mice retained some scratching behavior;14 additionally, 50% of the patients continued to suffer from opioids-induced itch.23 There is no standard therapy for morphine-induced itch. It is well known that the systemic administration of morphine has a sedative effect.24 We showed that the intrathecal administration of high-dose morphine also causes sedation with a decrease in the scratching behavior in mice.14 Therefore, intrathecal morphine administration did not produce scratching behavior dose-dependently. Furthermore, 0.3-nmol morphine was selected as a combination dose with SB366791. To the best of our knowledge, no studies have reported the ability of intrathecal TRPV1 antagonists to attenuate morphine-induced itch. This study demonstrated that an intrathecal TRPV1 antagonist inhibited intrathecal morphine-induced itch.
The molecular mechanisms of morphine-induced itch are not completely understood, but the mu-opioid receptor (MOR) isoform MOR1D and the gastrin-releasing peptide receptor (GRPR) are known to play a critical role in these mechanisms. MOR1D and GRPR are colocalized in the dorsal horn of the spinal cord.25 Morphine induces heterodimerization and co-internalization of MOR1D and GRPR. GRPR activates the phospholipase C (PLC)/inositol 1, 4, 5 triphosphate (IP3)/calcium signaling pathway. This PLC/IP3/calcium signaling pathway evokes morphine-induced itch.25
Although the mechanisms of antipruritic effects of TRPV1 antagonists on morphine-induced itch are unknown, possible mechanism may include as followed. TRPV1 is mainly expressed in the central and peripheral terminals of primary sensory neurons.26 The central terminal of the primary sensory neurons lies in the dorsal horn of the spinal cord and is concentrated in the superficial laminae. MOR and TRPV1 are colocalized in the superficial laminae of dorsal horn.26 Furthermore, TRPV1-expressing neurons release gastrin-releasing peptide (GRP) in the dorsal horn of the spinal cord, resulting in the evocation of the GRP–GRPR signaling pathway.27 These studies and our results indicate the possibility that TRPV1 interact between MOR1D and GRPR in the spinal cord.
Although SB366791 is not a pure TRPV1 antagonist, SB366791 is a potent and high selectivity TRPV1 antagonist which has little or no effect on the activity against a wide range of receptors, including opioids.28 In our data, SB366791 did not inhibit morphine-induced antinociception. Therefore, we suggest that SB366791 produces antipruritic effects through TRPV1 and not MOR.
It has been reported that a TRPV1 antagonist produces little or no antinociceptive effects in naïve models.29 Consistent with this report,29 an intrathecal TRPV1 antagonist did not produce antinociceptive effects in naïve mice in our study. However, TRPV1 antagonists have shown antinociceptive effects in several pain models.7–9 TRPV1 expression was increased in bone cancer pain mice model.30 In addition, TRPV1 is functionally upregulated in postoperative pain mice model.31 This difference between naïve models and pain models may depend on the activation of TRPV1, including the increase of TRPV1 expression or its up-regulation in the pain models.30,31 The antinociceptive effects of morphine vary depending on the animal models. Morphine is effective against postoperative and inflammatory pain compared with neuropathic pain.32,33 Bone cancer pain is resistant to morphine compared with inflammatory pain.34 Downregulation of MOR expression is thought to attenuate sensitivity of bone cancer pain to morphine.33 It has been reported that the combination of morphine and TRPV1 antagonists has potent analgesic effect on bone cancer model.35
Although TRPV1 antagonists have been widely accepted as next-generation pain therapies, many clinical studies of TRPV1 antagonists have been put on hold, mainly because of adverse events.36,37 In fact, the systemic use of TRPV1 antagonists in basic research studies has been shown to cause hyperthermia.11–13 The present study showed that an intrathecal TRPV1 antagonist did not affect the body temperature of mice.
There are some limitations to our study. First, the mechanisms of the antipruritic effects observed after the intrathecal administration of SB366791 combined with morphine were not clear. Further studies are needed to address these issues. Second, although no mice exhibited any side effects, such as motor dysfunction, after the intrathecal administration of SB366791, the neurotoxicity of SB366791 at the spinal level was not clarified.
Conclusion
This study demonstrated that intrathecal SB366791 reduced intrathecal morphine-induced itch without causing hyperthermia and did not suppress morphine-induced antinociception for mice.
Acknowledgment
This work was supported by Japan Society for the Promotion of Science KAKENHI Grant Number 17K09033.
Author contributions
SS and NI designed the project. SS, MS, YK, and MH performed the experiments. SS and NI wrote the manuscript. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
Dr. Yoji Saito reports personal fees from Kyowa Kirin, Phizer, Nipponzoki, Ayumi-Pharma, Tsumura, Maruishi, Japan Vaccine, Aspen Japan, and Bikken. He also reports personal fees including research funding and lecture fee from Daiichi Sankyo and AsahiKASEI, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Jannuzzi RG. Nalbuphine for treatment of opioid-induced pruritus: a systematic review of literature. Clin J Pain. 2016;32(1):87–93. doi:10.1097/AJP.0000000000000211
2. Ko MC, Song MS, Edwards T, Lee H, Naughton NN. The role of central mu opioid receptors in opioid-induced itch in primates. J Pharmacol Exp Ther. 2004;310(1):169–176. doi:10.1124/jpet.103.061101
3. Dunteman E, Karanikolas M, Filos KS. Transnasal butorphanol for the treatment of opioid-induced pruritus unresponsive to antihistamines. J Pain Symptom Manage. 1996;12(4):255–260.
4. Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389(6653):816–824. doi:10.1038/39807
5. Moore C, Gupta R, Jordt SE, Chen Y, Liedtke WB. Regulation of pain and itch by TRP channels. Neurosci Bull. 2018;34(1):120–142. doi:10.1007/s12264-017-0200-8
6. Bao Y, Gao Y, Yang L, et al. The mechanism of mu-opioid receptor (MOR)-TRPV1 crosstalk in TRPV1 activation involves morphine anti-nociception, tolerance and dependence. Channels (Austin, Tex). 2015;9(5):235–243. doi:10.1080/19336950.2015.1069450
7. Helyes Z, Sandor K, Borbely E, et al. Involvement of transient receptor potential vanilloid 1 receptors in protease-activated receptor-2-induced joint inflammation and nociception. Eur J Pain. 2010;14(4):351–358. doi:10.1016/j.ejpain.2009.07.005
8. Wong GY, Gavva NR. Therapeutic potential of vanilloid receptor TRPV1 agonists and antagonists as analgesics: recent advances and setbacks. Brain Res Rev. 2009;60(1):267–277. doi:10.1016/j.brainresrev.2008.12.006
9. Uchytilova E, Spicarova D, Palecek J. TRPV1 antagonist attenuates postoperative hypersensitivity by central and peripheral mechanisms. Mol Pain. 2014;10:67. doi:10.1186/1744-8069-10-67
10. Imamachi N, Park GH, Lee H, et al. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc Natl Acad Sci U S A. 2009;106(27):11330–11335. doi:10.1073/pnas.0905605106
11. Gavva NR, Bannon AW, Surapaneni S, et al. The vanilloid receptor TRPV1 is tonically activated in vivo and involved in body temperature regulation. J Neurosci. 2007;27(13):3366–3374. doi:10.1523/JNEUROSCI.4833-06.2007
12. Garami A, Shimansky YP, Pakai E, Oliveira DL, Gavva NR, Romanovsky AA. Contributions of different modes of TRPV1 activation to TRPV1 antagonist-induced hyperthermia. J Neurosci. 2010;30(4):1435–1440. doi:10.1523/JNEUROSCI.5150-09.2010
13. Alawi KM, Aubdool AA, Liang L, et al. The sympathetic nervous system is controlled by transient receptor potential vanilloid 1 in the regulation of body temperature. Faseb J. 2015;29(10):4285–4298. doi:10.1096/fj.15-272526
14. Sakakihara M, Imamachi N, Saito Y. Effects of intrathecal kappa-opioid receptor agonist on morphine-induced itch and antinociception in mice. Reg Anesth Pain Med. 2016;41(1):69–74. doi:10.1097/AAP.0000000000000326
15. Spicarova D, Adamek P, Kalynovska N, Mrozkova P, Palecek J. TRPV1 receptor inhibition decreases CCL2-induced hyperalgesia. Neuropharmacology. 2014;81:75–84. doi:10.1016/j.neuropharm.2014.01.041
16. Fairbanks CA. Spinal delivery of analgesics in experimental models of pain and analgesia. Adv Drug Deliv Rev. 2003;55(8):1007–1041.
17. Kuraishi Y, Nagasawa T, Hayashi K, Satoh M. Scratching behavior induced by pruritogenic but not algesiogenic agents in mice. Eur J Pharmacol. 1995;275(3):229–233. doi:10.1016/0014-2999(94)00780-b
18. Saegusa Y, Tabata H. Usefulness of infrared thermometry in determining body temperature in mice. J Vet Med Sci. 2003;65(12):1365–1367. doi:10.1292/jvms.65.1365
19. Bonnet M-P, Marret E, Josserand J, Mercier FJ. Effect of prophylactic 5-HT3 receptor antagonists on pruritus induced by neuraxial opioids: a quantitative systematic review. Br J Anaesth. 2008;101(3):311–319. doi:10.1093/bja/aen202
20. George RB, Allen TK, Habib AS. Serotonin receptor antagonists for the prevention and treatment of pruritus, nausea, and vomiting in women undergoing cesarean delivery with intrathecal morphine: a systematic review and meta-analysis. Anesth Analg. 2009;109(1):174–182. doi:10.1213/ane.0b013e3181a45a6b
21. Liu X, Zhang J, Zhao H, Mei H, Lian Q, Shangguan W. The effect of propofol on intrathecal morphine-induced pruritus and its mechanism. Anesth Analg. 2014;118(2):303–309. doi:10.1213/ANE.0000000000000086
22. Warwick JP, Kearns CF, Scott WE. The effect of subhypnotic doses of propofol on the incidence of pruritus after intrathecal morphine for caesarean section. Anaesthesia. 1997;52(3):270–275. doi:10.1111/j.1365-2044.1997.069-az0060.x
23. Hirabayashi M, Doi K, Imamachi N, Kishimoto T, Saito Y. Prophylactic pentazocine reduces the incidence of pruritus after cesarean delivery under spinal anesthesia with opioids: a prospective randomized clinical trial. Anesth Analg. 2017;124(6):1930–1934. doi:10.1213/ANE.0000000000002060
24. Young-McCaughan S, Miaskowski C. Definition of and mechanism for opioid-induced sedation. Pain Manag Nurs. 2001;2(3):84–97. doi:10.1053/jpmn.2001.25012
25. Liu XY, Liu ZC, Sun YG, et al. Unidirectional cross-activation of GRPR by MOR1D uncouples itch and analgesia induced by opioids. Cell. 2011;147(2):447–458. doi:10.1016/j.cell.2011.08.043
26. Scherrer G, Imamachi N, Cao YQ, et al. Dissociation of the opioid receptor mechanisms that control mechanical and heat pain. Cell. 2009;137(6):1148–1159. doi:10.1016/j.cell.2009.04.019
27. Tsuda M. Astrocytes in the spinal dorsal horn and chronic itch. Neurosci Res. 2018;126:9–14. doi:10.1016/j.neures.2017.08.010
28. Gunthorpe MJ, Rami HK, Jerman JC, et al. Identification and characterisation of SB-366791, a potent and selective vanilloid receptor (VR1/TRPV1) antagonist. Neuropharmacology. 2004;46(1):133–149. doi:10.1016/s0028-3908(03)00305-8
29. Kitagawa Y, Tamai I, Hamada Y, et al. Orally administered selective TRPV1 antagonist, JTS-653, attenuates chronic pain refractory to non-steroidal anti-inflammatory drugs in rats and mice including post-herpetic pain. J Pharmacol Sci. 2013;122(2):128–137.
30. Niiyama Y, Kawamata T, Yamamoto J, Omote K, Namiki A. Bone cancer increases transient receptor potential vanilloid subfamily 1 expression within distinct subpopulations of dorsal root ganglion neurons. Neuroscience. 2007;148(2):560–572. doi:10.1016/j.neuroscience.2007.05.049
31. Barabas ME, Stucky CL. TRPV1, but not TRPA1, in primary sensory neurons contributes to cutaneous incision-mediated hypersensitivity. Mol Pain. 2013;9:9. doi:10.1186/1744-8069-9-9
32. Feehan AK, Morgenweck J, Zhang X, Amgott-Kwan AT, Zadina JE. Novel endomorphin analogs are more potent and longer-lasting analgesics in neuropathic, inflammatory, postoperative, and visceral pain relative to morphine. J Pain. 2017;18(12):1526–1541. doi:10.1016/j.jpain.2017.08.007
33. Yamamoto J, Kawamata T, Niiyama Y, Omote K, Namiki A. Down-regulation of mu opioid receptor expression within distinct subpopulations of dorsal root ganglion neurons in a murine model of bone cancer pain. Neuroscience. 2008;151(3):843–853. doi:10.1016/j.neuroscience.2007.11.025
34. Luger NM, Sabino MA, Schwei MJ, et al. Efficacy of systemic morphine suggests a fundamental difference in the mechanisms that generate bone cancer vs inflammatory pain. Pain. 2002;99(3):397–406.
35. Niiyama Y, Kawamata T, Yamamoto J, Furuse S, Namiki A. SB366791, a TRPV1 antagonist, potentiates analgesic effects of systemic morphine in a murine model of bone cancer pain. Br J Anaesth. 2009;102(2):251–258. doi:10.1093/bja/aen347
36. Gavva NR, Treanor JJ, Garami A, et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain. 2008;136(1–2):202–210. doi:10.1016/j.pain.2008.01.024
37. Rowbotham MC, Nothaft W, Duan WR, et al. Oral and cutaneous thermosensory profile of selective TRPV1 inhibition by ABT-102 in a randomized healthy volunteer trial. Pain. 2011;152(5):1192–1200. doi:10.1016/j.pain.2011.01.051
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.