")
Back to Journals » Cancer Management and Research » Volume 12
Effect of RSK4 on Biological Characteristics of Gastric Cancer
Authors Hu C , Dai J , Lin X , Meng Y , Liang H
Received 11 November 2019
Accepted for publication 15 January 2020
Published 28 January 2020 Volume 2020:12 Pages 611—619
DOI https://doi.org/10.2147/CMAR.S238132
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Chien-Feng Li
Cong Hu,1 Jun Dai,1 Xu Lin,1 Ya Meng,2 Hui Liang1
1The First Department of General Surgery, Zhuhai People’s Hospital, Jinan University, Zhuhai City, Guangdong Province, People’s Republic of China; 2Zhuhai Precision Medicine Research Center, Zhuhai People’s Hospital, Jinan University, Zhuhai City, Guangdong Province, People’s Republic of China
Correspondence: Hui Liang
Zhuhai People’s Hospital, 79 Kangning Road, Zhuhai City, Guangdong Province, People’s Republic of China
Tel +86 756-2158341
Email [email protected]
Purpose: Gastric cancer is one of the most common cancers with high mortality. Emerging evidences show that ribosomal s6 kinase4 (RSK4) may be an anti-oncogene in several types of cancers, while its function in GC is still unclear. In the present study, we investigated the role of RSK4 in GC progression using MGC-803 and HGC-27 cell lines in vitro and in vivo.
Methods: The expression of RSK4 in gastric cancer cells was evaluated using RT-qPCR and Western blot analysis. We transfected cells with RSK4 siRNA to reduce the expression of RSK4 and then evaluated the effect of RSK4 on cellular function. MTT and cell cycle assays were used to study its effect on cell growth. Flow cytometry was used to evaluate cell apoptosis. Wound healing and Transwell assays were performed to investigate metastasis. Stable cell lines with or without RSK4 knockdown were constructed with lentivirus and tumor-bearing mice were used to investigate the effect of RSK4 on cancer progression.
Results: The results revealed that reduction of RSK4 expression inhibited cell apoptosis and promoted cell proliferation, migration, and invasion. Additionally, RSK4 knockdown promoted tumorigenesis in vivo.
Conclusion: Our study demonstrated that RSK4 serves as a tumor suppressor in GC.
Keywords: gastric cancer, RSK4, proliferation, apoptosis, migration, invasion
Introduction
Gastric cancer is the fifth most common cancer and the third leading cause of cancer-related death in the world, with nearly one million newly diagnosed cases every year.1 The prognosis of gastric cancer is poor and the average five-year survival rate is less than 20%.2 Cancer stem cells and metastasis usually cause refractory of gastric cancer.3 Furthermore, gastric cancer has caused an estimated 783,000 deaths in 2018.1 Thus, gastric cancer remains a clinical challenge and finding novel therapeutic targets is imperative.
RSK4, also known as RPS6KA6, is a member of the p90 ribosomal S6 kinase (RSKs) family, which are serine/threonine protein kinases that activate downstream of the Ras/MAPK pathway.4 RSKs participate in the regulation of cell proliferation, survival, invasion, and metastasis.5–9 Several reports have identified RSK4 as a tumor suppressor in the colon, breast, and renal carcinomas.10–13 The over-expression of RSK4 could restrict cell growth, demonstrating anti-invasive and anti-metastatic effects.14,15 In addition, RSK4 is recommended as a useful prognostic marker in patient evaluation.16 However, contradictory studies inconsistent with the findings are available. Thakur et al showed that the expression level of RSK4 was higher in some breast cancer tissues than in adjacent tissues.17 Sun et al suggested that whether RSK4 is oncogenic or tumor-suppressive depends on many factors.18 The purpose of our study is to explore the biological function of RSK4 in gastric cancer and provide a basis for its role in oncotherapy.
In this study, we observed that RSK4 played an important role in cell growth, apoptosis, and metastasis in vitro, and tumorigenesis in vivo. RSK4 serves as a tumor suppressor in gastric cancer, suggesting that RSK4 could be a potential target for patient therapy.
Methods and Materials
Cell Lines and Cell Culture
HEK293T, MGC-803, and HGC-27 cell lines were obtained from ATCC. Cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum (FBS),100 U/mL penicillin, 100 U/mL streptomycin, and 2 mM l-glutamine at 37°C in a 5% CO2 atmosphere with 21% oxygen.
Transfection
MGC-803 and HGC-27 cells were transfected with siRNA NC or RSK4 siRNA by ExFect2000 Transfection Reagent (Vazyme, Nanjing, Jiangsu, China), according to the manufacturer’s instruction. The sequences of siRNA are shown below.
siRNA NC: 5′-TTCTCCGAACGTGTCACGT-3′;
RSK4 siRNA-1: 5′-CATCGTTGTCTGCTAAATTAC-3′;
RSK4 siRNA-2: 5′-GTCCACAATATTCATACTATG-3′;
RSK4 siRNA-2: 5′-TTTACCTTGTTACGGATTTAA-3′.
Plasmid Construction and Lentiviral Transduction
The lentivirus-based vector pLKO.1-TRC-puro was used for the construction of the control short hairpin RNA (shctrl) lentivirus (siRNA NC) and shRSK4 lentivirus (RSK4 siRNA-1). HEK293T cells were used for the lentivirus package. Stable HGC-27 cells with or without RSK4 knockdown, used for tumor xenograft, were constructed by lentivirus transduction. HGC-27 cells were transfected with the lentivirus at 20 multiplicity of infection (MOI) and treated with puromycin (Yeasen, Shanghai, China) for 4 days.
qRT-PCR Assay
RNA isolated from cells was reverse transcribed using Superscript III transcriptase (Invitrogen). To determine the mRNA expression level of gene, primers were used in reverse transcription and qPCR; the assay was performed using the Bio-Rad CFX96 system with SYBR green (RSK4 and 18s RNA were transcribed using random reverse transcription primers):
RSK4, forward, 5′-CCTCCTTTCAAACCTGCTTCTGG-3′;
RSK4, reverse, 5′-GCTGATGAGCATTTGCACTGGC-3′;
18s, forward, 5′-CGACGACCCATTCGAACGTCT-3′;
18s, reverse, 5′-CTCTCCGGAATCGAA CCCTGA-3′.
Western Blot
Cells were lysed in ice-cold RIPA buffer and 20 µg protein was loaded for electrophoresis on 8–12% denaturing SDS-PAGE gels. The blots were probed with the primary antibodies overnight at 4ºC and followed by incubation with the appropriate secondary antibodies at room temperature for 1 h. The antibodies used in this study include Coronin3 (Cat: Ab76117, Abcam) and GAPDH (Cat: YM3029, ImmunoWay).
Cell Proliferation Assay
After transfection with siRNA ctrl or RSK4 siRNA, cells were seeded in 96-well plates at an initial density of (3 × 103 cells/well). The cells were stained with MTT (Cat: QF0025, Qiancheng Biotech, Shanghai) at each time point for 1 h at 37°C. The absorbance was measured at 490 nm. All experiments were performed in triplicates.
Cell Cycle Assay
Cells were seeded at a density of 2 × 105 cells in 6-well plates overnight. The next day, cells were transfected with siRNA ctrl or RSK4 siRNA. After 24–48 h the cells were harvested and fixed in 70% ethanol at 4°C overnight. The fixed cells were incubated with 0.5 mL phosphate-buffered saline (PBS) containing 10 μg/mL RNase and 0.2% Triton X-100 for 30 min at 37°C, then stained with 20 μg/mL propidium iodide (PI) for 30 min in a dark at room temperature, and finally analyzed using fluorescence-activated cell sorting (FACS).
Annexin V/Propidium Iodide Staining Assays
Annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) (Cat: A211-02, Vazyme) were used to detect cell apoptosis induced by cisplatin. MGC-803 and HGC-27 cells were seeded in 6-well plates, followed by transfection with siRNA ctrl or RSK4 siRN4 for 24–48 h. Then, the cells were collected, rinsed with PBS and stained with Annexin V-FITC (200 µg/mL) and 30 µg/mL PI in the dark at room temperature for 10 min. Immediately, the fluorescence was analyzed in 10,000 cells/sample using the flow cytometer NovoCyte (Cat:1300, ACEA). The results were expressed as the percentage of cells exhibiting apoptosis relative to the total number of cells analyzed.
Wound Healing Assay
MGC-803 and HGC-27 cells were seeded into 6-well plates at 100% confluence. After transfection with siRNA ctrl or RSK4 siRNA, the wound was stimulated with a 200 μL pipette tip with a straight scratch. Cells were continuously cultured in serum-free medium for 24 h and observed under the microscope.
Transwell Assay
Migration and invasion assays were performed using Transwell plates (Corning) with 8 μm-pore size membranes without matrigel (for migration assays) or with matrigel (for invasion assays). After transfection with siRNA ctrl and RSK4 siRNA for 24 h, 2–5×105 MGC-803 and HGC-27 cells were plated in the upper chambers of the Transwell. Twenty-four hours later, migrated cells were stained with 0.5% toluidine blue and counted from six random fields. For the invasion assay, the membrane of the upper chambers was pre-coated with 5 folds diluted matrigel (BD Biosciences, Sparks, MD) before use. The migrating or invading cells were counted and photographed.
Tumor Xenograft
Six-week-old female nude mice were obtained from Charles River (Beijing, China). Stably RSK4-knockdown HGC-27 cells or control cells (4 x 106) were subcutaneously injected into the right side dorsal flank of each mouse. From the 14th day after injection, the tumor size was recorded every 4 days. The tumor volume was calculated using the formula of V = ab2/2 (cm3). Thirty-four days after injection, the tumors were obtained and photographed
Statistics
All statistics were analyzed using SPSS 22.0 (IBM SPSS, Armonk, NY, USA). Student’s t-test (unpaired) was performed for pairwise comparison of groups. ANOVA (Two-way analysis of variance) was performed to identify significant differences among multiple groups. A p < 0.05 was considered a statistically significant difference. GraphPad Prism 8.2.1 (GraphPad Software, Inc., La Jolla, CA, USA) was used to prepare the graphs.
Results
Low RSK4 Expression Level Was Linked to Poor Prognosis
To investigate the expression and developmental role of RSK4 in gastric cancer, we analyzed the mRNA expression in 25 patients from the TCGA database. As shown in Figure 1A, the RSK4 expression level was significantly lower in the gastric cancer tissues compared to the adjacent normal tissues. Clinical information from 126 gastric cancer patients was downloaded from the GEO database. The Kaplan-Meier survival analysis was used to determine the correlation of RSK4 expression with overall survival (OS) in patients with gastric cancer. The result indicated that low RSK4 expression in patients exhibited a shorter OS time (Figure 1B). These findings suggested that RSK4 could be a tumor suppressor.
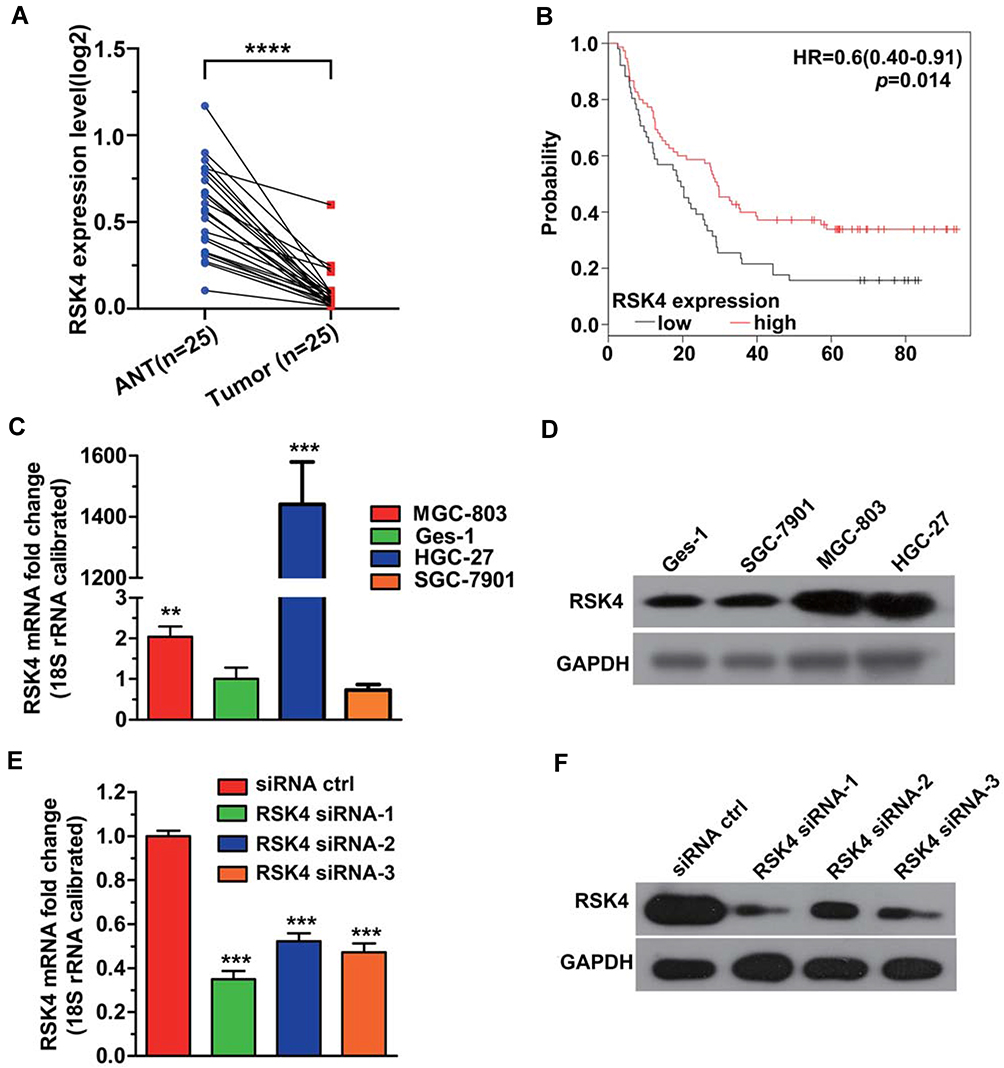 |
Figure 1 Expression levels of RSK4 in gastric cancer tissues and cell lines and the association of RSK4 expression with prognosis. (A) The mRNA level of RSK4 is lower in gastric cancer tissues than that in adjacent normal tissues. The data from 25 patients was obtained from the TCGA database. TCGA: The Cancer Genome Atlas. Mann–Whitney test: ****p < 0.0001. (B) The low mRNA level of RSK4 predicts a poor prognosis. Kaplan-Meier survival analysis was performed to analyze the correlation of RSK4 expression with OS in gastric cancer patients from the GEO database. OS, overall survival. GEO, Gene Expression Omnibus. (C, D) The mRNA expression and protein levels of RSK4 were analyzed using qRT-PCR and Western blot assay, respectively. Unpaired two-tailed t-test: **p < 0.01 and ***p < 0.001. (E, F) The RSK4 mRNA and protein expressions in HGC27 were measured after transfection with RSK4-si-NC, RSK4-si-1, RSK4-si-2 or RSK4-si-3. The data are represented as the mean ± s.d. Unpaired two-tailed t-test: ***p < 0.001. |
Expression Levels of RSK4 in Gastric Cancer Cell Lines and RSK4 siRNA Knockdown Efficiencies
The mRNA expression levels of RSK4 in gastric cancer cell lines (Ges-1, SGC-7901, MGC-803, and HGC-27) were detected using qRT-PCR. As shown in Figure 1C, the relative mRNA level of RSK4 in HGC-27 was the highest, followed by MGC-803. Similarly, the Western blot assay demonstrated that the expression levels of the RSK4 protein were the highest in HGC-27 and MGC-803 (Figure 1D). HGC27 cells were transfected with RSK4 siRNA and the knockdown efficiencies were evaluated by qRT-PCR and Western blot assay. The results indicated that the expression of RSK4 significantly decreased in both mRNA and protein levels (Figure 1E and F). Furthermore, the knockdown efficiency of RSK4-si-1 was superior. Based on the above results, we selected HGC-27, MGC-803, and RSK4 si-1 (named RSK4 siRNA) for the following experiments.
Knockdown of RSK4 Promotes Cell Growth
The MTT assay was used to detect the proliferation of BCG-803 and HGC-27 gastric cancer cell lines after transfection with siRNA ctrl and RSK4 siRNA, respectively. The results demonstrated that the cellular proliferation abilities were improved after transfection with RSK4 siRNA (Figure 2A), indicating that RSK4 suppresses cell growth. We then performed the cell cycle assay by flow cytometry (PI flow assay). As shown in Figure 2B and C, in the G0/G1 phase, the population of MGC-803 and HGC-27 cells was obviously decreased after transfection with RSK4 siRNA. RSK4 knockdown induced entry into the S phase to promote cell growth.
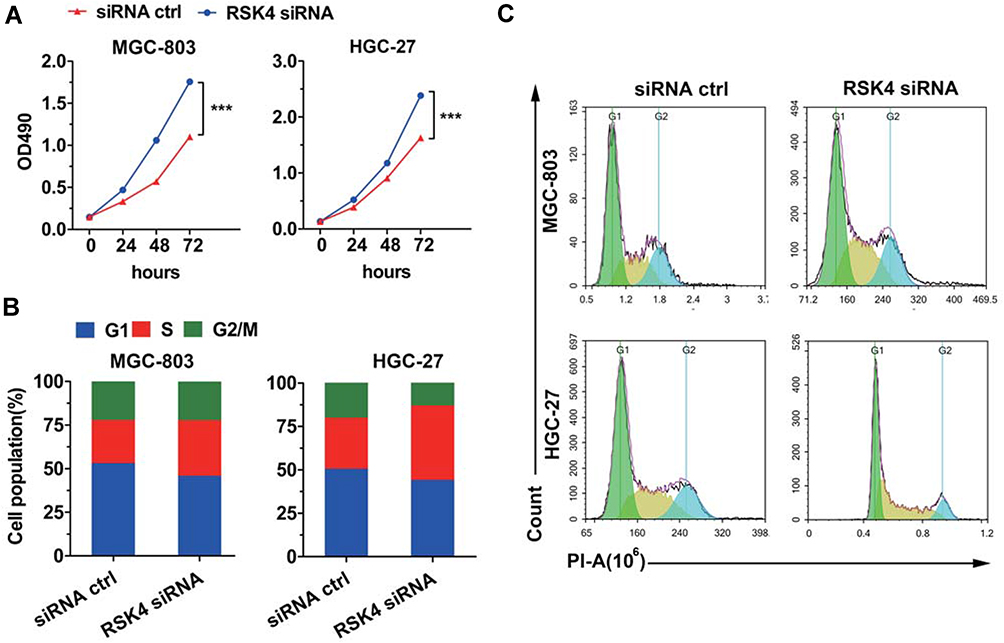 |
Figure 2 Knockdown of RSK4 promotes cell proliferation. (A) MGC-803 and HGC-27 cells were transfected with siRNA ctrl and RSK4 siRNA, respectively. MTT assay was used to detect cell viability at different time points. Two-way analysis of variance: ***p < 0.001. (B) The knockdown of RSK4 promotes cell entry in the S phase. MGC-803 and HGC-27 were transfected with siRNA ctrl and RSK4 siRNA, respectively. Cells were harvested and analyzed 24 h after transfection by flow cytometry. (C) Representative images of cell cycle assay. |
Knockdown of RSK4 Inhibits Cell Apoptosis
We used Annexin V/PI staining to analyze cell apoptosis. The results demonstrated that the proportion of apoptotic cells in the experimental groups markedly decreased (p < 0.01) (Figure 3A and B). After RSK4 knockdown, the percentage of apoptotic cells was reduced from 10.12±0.57% to 7.13±0.32% in MGC-803 and from 13.80±1.32% to 7.93%±0.75% in HGC-27, respectively (Figure 3B). These findings indicate that RSK4 promotes cell apoptosis.
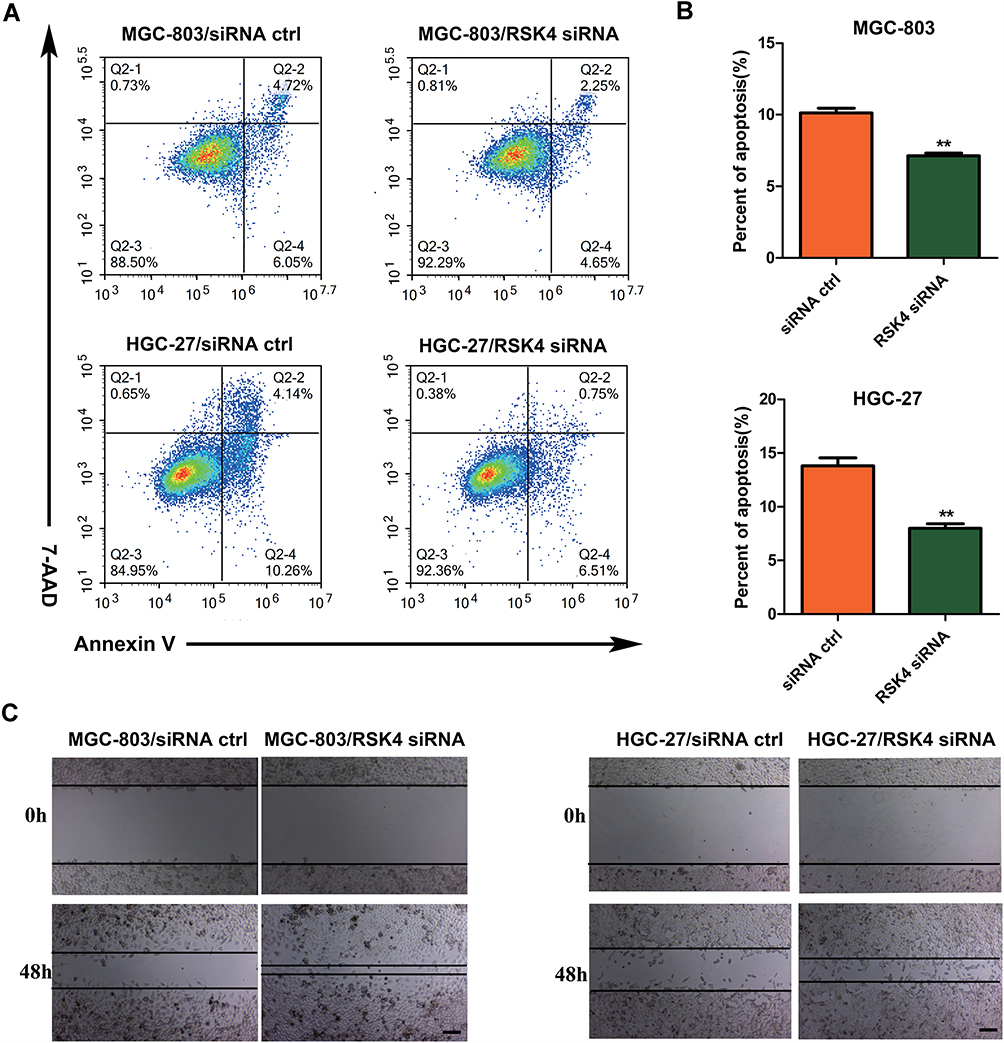 |
Figure 3 Apoptosis and wound healing assays. (A) Apoptosis was analyzed by flow cytometry for MGC-803 and HGC-27. MGC-803 and HGC-27 cells were transfected with siRNA ctrl and RSK4 siRNA, respectively. Cells were harvested and analyzed 24 h after transfection. (B) The knockdown of RSK4 reduces cell apoptosis. The data are represented as the mean ± s.d. Unpaired two-tailed t-test: **p < 0.01. (C) Movement of cells into the wound was shown for siRNA ctrl and RSK4 siRNA transfected cells at 0 h and 48 h post scratch. Magnification: 100×. |
Knockdown of RSK4 Promotes Migration and Invasion
To analyze the effect of RSK4 on cell migration, we performed wound healing and Transwell migration assays. As shown in Figure 3C, the wounds were almost the same size at 0 h; however, knockdown of RSK4 greatly accelerated wound closure at 48 h. Furthermore, the Transwell migration assay demonstrated that knockdown of RSK4 increased the number of migrating cells (Figure 4A and B). Next, we used the matrigel invasion assay to determine whether knockdown of RSK4 induces invasion. The results showed that knockdown of RSK4 improved the number of invasive cells (Figure 4C and D). All the above data suggested that RSK4 inhibited metastasis.
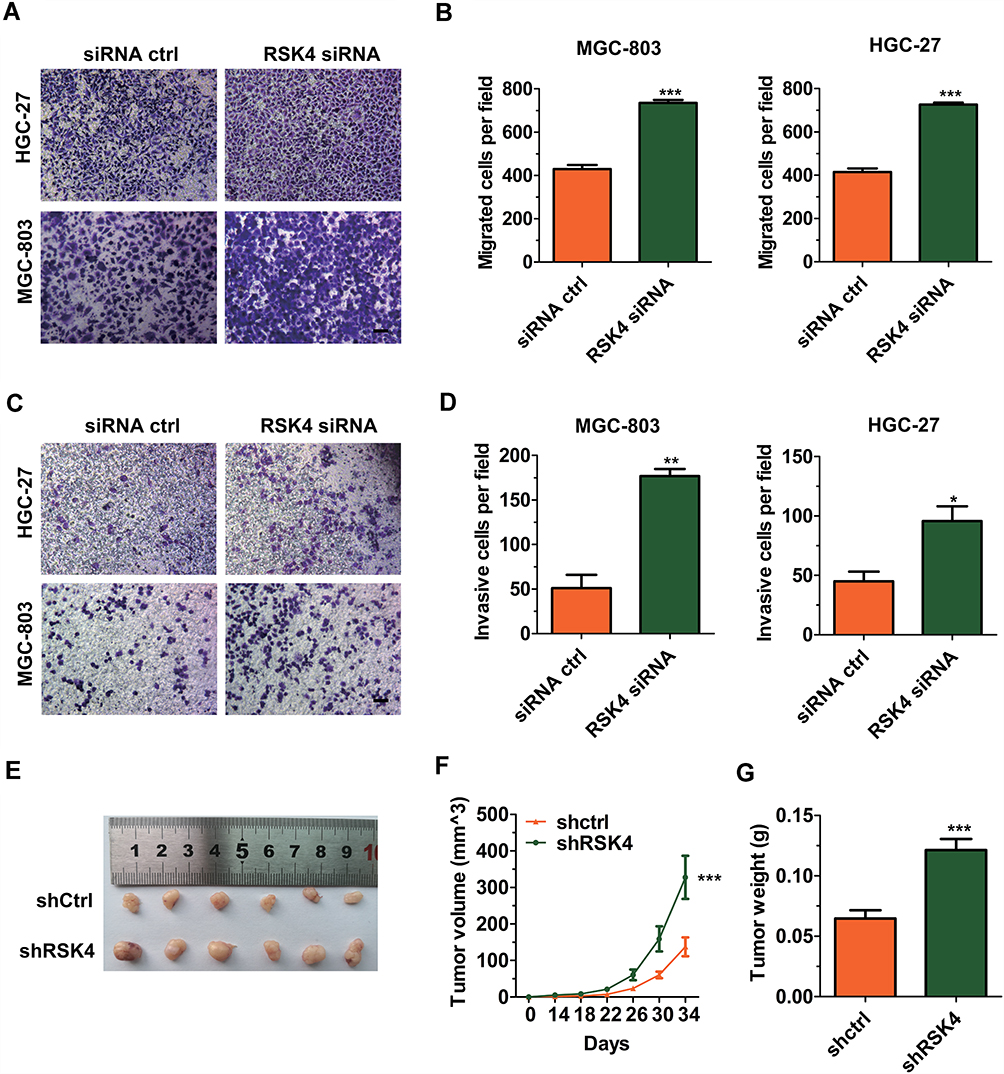 |
Figure 4 Knockdown of RSK4 promotes cell migration, invasion and tumorigenesis. (A) Representative images of Transwell migration assay. Magnification: 100×. (B) Histogram of migrated cells per field. Unpaired two-tailed t-test: ***p < 0.001. (C) Representative images of matrigel invasion assay. The membrane of the upper chambers was pre-coated with 5 fold diluted matrigel. Magnification: 100×. (D) Histogram of invasive cells per field. Unpaired two-tailed t-test: *p < 0.05, **p < 0.01. (E) Images of the xenograft-derived tumors. The stable cell line of HGC-27 with or without RSK4 knockdown was subcutaneously injected into a nude mouse (4 × 106 per mouse). (F) Comparison of the tumor volume. The tumor size was recorded every 4 days from the 14th day after injection. The tumor volume was calculated by the formula of V = ab2/2(cm3). Two-way analysis of variance: ***p < 0.001. (G) Comparison of the tumor weight. The tumor was weighed immediately after harvesting. Unpaired two-tailed t-test: ***p < 0.001. |
Knockdown of RSK4 Promotes Gastric Cancer Cell Growth in Xenograft Mice
To study the effect of RSK4 on tumor growth in vivo, HGC-27 cells with or without RSK4 knockdown were injected subcutaneously into nude mice. The results indicated that knockdown of RSK4 promoted tumorigenesis (Figure 4E). The tumor size was recorded every 4 days from the 14th day after injection, and the tumor weight was measured immediately after dissection. As shown in Figure 4F and G, the tumor volume was significantly increased and the tumor weight was markedly elevated in the RSK4 knockdown group compared to the control group.
Discussion
The incidence of gastric cancer has been increasing every year in developed and developing countries. It is the fifth frequently diagnosed cancer and the third leading cause of cancer-related mortality worldwide.1 In recent years, a large number of investigations have been performed to improve the prognosis of patients with gastric cancer, with neoadjuvant chemotherapy, radiotherapy, and molecular-targeted therapies adopted as efficacious strategies.19 Despite this, the prognosis of gastric cancer is still dim. Thus, it is necessary to find a specific molecular target to attack the gastric tumor.
RSK4 has been reported as a tumor suppressor in multiple cancers,12,13,16 which is consistent with our study. In our study, we hypothesized that RSK4 was an anti-oncogene in gastric cancer. We transfected gastric cancer cells with RSK4 siRNA and then investigated the changes in cellular function after RSK4 knockdown. The results revealed that knockdown of RSK4 promoted proliferation, migration and invasion, confirming our hypothesis. Based on these results, the expression of RSK4 in carcinomas should be down-regulated as reported by Lopez-Vicente et al11 However, our qRT-PCR and Western blotting assays demonstrated that RSK4 expression in gastric cancer cell lines MGC-803 and HGC-27 was higher than that in the normal gastric cell line, Ges-1 (Figure 1). This could be attributed to the difference between tissues and cell lines. Hence, in our next investigation, we plan to compare the RSK4 expression between gastric carcinomas and adjacent tissues.
After transfection with RSK4 siRNA, the gastric cancer cells demonstrated a significantly reduced population in the G1/G0 phase and the percentage of cells in the S phase increased, suggesting that RSK4 induces G1/G0 phase arrest to inhibit cell proliferation.
RNA interference (RNAi) is a powerful tool to block the expression of the target gene, facilitating an understanding of gene function.20 RNAi is a post-transcriptional process whereby double-stranded short interfering RNA or microRNA target degradation or prevent the translation of an mRNA which contains complementarity to it.21 However, the gene function study could only be performed in vitro using RNAi. We analyzed the influence of RSK4 on the in vivo gastric cancer progress through the construction of stably over-expressive RSK4 cell lines using a lentivirus system.
Conclusion
Our study indicated that RSK4 was a significant anti-oncogene in gastric cancer and could be a potential target in cancer therapy.
Ethics Approval and Consent to Participate
All mouse studies were conducted in accordance with Laboratory Animal Management Regulations of Guangdong province and were approved by the Institutional Animal Care and Use Committee (IACUC) at Jinan University.
Disclosure
All authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.1002/jbm.b.34246. (1542-4863 (Electronic))
2. Correa P. Gastric cancer: overview. Gastroenterol Clin North Am. 2013;42(2):211–217. doi:10.1016/j.gtc.2013.01.002
3. Mariette C, JM B, Piessen G, et al. Pattern of recurrence following complete resection of esophageal carcinoma and factors predictive of recurrent disease. Cancer. 2003;97(7):1616–1623. (0008-543X (Print)).
4. Frodin M, Gammeltoft S. Role and regulation of 90 kDa ribosomal S6 kinase (RSK) in signal transduction. Mol Cell Endocrinol. 1999;151(1–2):65–77. (0303-7207 (Print)).
5. Lara R, Seckl OE, Pardo MF, Pardo OE. The p90 RSK family members: common functions and isoform specificity. Cancer Res. 2013;73(17):5301–5308. (1538-7445 (Electronic)).
6. Fogarty S, Hardie DG. C-terminal phosphorylation of LKB1 is not required for regulation of AMP-activated protein kinase, BRSK1, BRSK2, or cell cycle arrest. J Biol Chem. 2009;284(1):77–84. (0021-9258 (Print)).
7. Kang S, Elf S, Lythgoe K, et al. p90 ribosomal S6 kinase 2 promotes invasion and metastasis of human head and neck squamous cell carcinoma cells. J Clin Investig. 2010;120(4):1165–1177. (1558-8238 (Electronic)).
8. Lara R, Mauri FA, Taylor H, et al. An siRNA screen identifies RSK1 as a key modulator of lung cancer metastasis. Oncogene. 2011;30(32):3513. (1476-5594 (Electronic)).
9. Buck M, Poli V, Hunter T, et al. C/EBPbeta phosphorylation by RSK creates a functional XEXD caspase inhibitory box critical for cell survival. Mol Cell. 2001;8(4):807–816. (1097-2765 (Print)).
10. Li Q, Jiang Y, Wei W, et al. Frequent epigenetic inactivation of RSK4 by promoter methylation in cancerous and non-cancerous tissues of breast cancer. Med Oncol. 2014;31(1):793. (1559-131X (Electronic)).
11. Lopez-Vicente L, Armengol G, Pons B, et al. Regulation of replicative and stress-induced senescence by RSK4, which is down-regulated in human tumors. Clin Cancer Res. 2009;15(14):4546–4553. (1078-0432 (Print)).
12. Ye Q, Wang X, Jin M, et al. Effect of RSK4 on biological characteristics of colorectal cancer. World J Surg Oncol. 2018;16(1):240. doi:10.3389/fmicb.2018.01477. (1477-7819 (Electronic))
13. Zhu J, Li QY, Liu JL, et al. RSK4 knockdown promotes proliferation, migration and metastasis of human breast adenocarcinoma cells. Oncol Rep. 2015;34(6):3156–3162. (1791-2431 (Electronic)).
14. Dummler BA, Hauge C, Silber J, et al. Functional characterization of human RSK4, a new 90-kDa ribosomal S6 kinase, reveals constitutive activation in most cell types. J Biol Chem. 2005;280(14):13304–13314. (0021-9258 (Print)).
15. Thakur A, Sun Y, Bollig A, et al. Anti-invasive and antimetastatic activities of ribosomal protein S6 kinase 4 in breast cancer cells. Clin Cancer Res. 2008;14(14):4427–4436. (1078-0432 (Print)).
16. Cai J, Ma H, Huang F, et al. Low expression of RSK4 predicts poor prognosis in patients with colorectal cancer. Int J Clin Exp Pathol. 2014;7(8):4959–4970.
17. Thakur A, KW R, Wu J, et al. Aberrant expression of X-linked genes RbAp46, Rsk4, and Cldn2 in breast cancer. Mol Cancer Res. 2007;5(2):171–181. (1541-7786 (Print)).
18. Sun Y, Cao S, Yang M, et al. Basic anatomy and tumor biology of the RPS6KA6 gene that encodes the p90 ribosomal S6 kinase-4. Oncogene. 2013;32(14):1794–1810. doi:10.1038/onc.2012.200
19. Song Z, Wu Y, Yang J, et al. Progress in the treatment of advanced gastric cancer. Tumor Biol. 2017;39(7):1010428317714626. (1423-0380 (Electronic)).
20. Sen GL, Blau HM. A brief history of RNAi: the silence of the genes. FASEB j. 2006;20(9):1293–1299. (1530-6860 (Electronic)).
21. Elbashir SM, Lendeckel T, Fau-Tuschl W, Tuschl T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev. 2001;15(2):188–200. (0890–9369 (Print)).
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.