")
Back to Journals » Journal of Pain Research » Volume 15
Effect of AMPK Subunit Alpha 2 Polymorphisms on Postherpetic Pain Susceptibility in Southwestern Han Chinese
Authors Mei Y , Mu Y, Wang W, Tan BT, Chen YH, Li YP, Zhu D, Li W , Cui J , Yu LH
Received 19 August 2022
Accepted for publication 5 October 2022
Published 21 October 2022 Volume 2022:15 Pages 3319—3326
DOI https://doi.org/10.2147/JPR.S385913
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qi Fang
Yang Mei,1,2 Yang Mu,3 Win Wang,2 Bo-Tao Tan,1 Yao-Hua Chen,4 Yu-Ping Li,4 Dan Zhu,4 Wei Li,2 Jian Cui,4 Le-Hua Yu1
1Department of Rehabilitation Medicine, The Second Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China; 2Department of Pain Medicine, Chongqing Hospital of Traditional Chinese Medicine, Chongqing, People’s Republic of China; 3Department of Rehabilitation Medicine, Chongqing University Fuling Hospital, Chongqing, People’s Republic of China; 4Department of Pain Medicine, Southwest Hospital, Army Medical University, Chongqing, People’s Republic of China
Correspondence: Jian Cui; Le-Hua Yu, Email [email protected]; [email protected]
Introduction: Adenosine 5’-monophosphate (AMP)-activated protein kinase (AMPK) can influence energy metabolism. Energy metabolism imbalance is closely associated with the occurrence of neuropathic pain (NeP). Rs10789038 and rs2796498 are genetic polymorphisms of PRKAA2, the gene encoding AMPK, which is closely related to energy metabolism imbalance. This study aimed to explore the relationship between PRKAA2 and postherpetic neuralgia (PHN) in the southwestern Chinese Han population.
Methods: This study enrolled 132 PHN patients and 118 healthy subjects. The rs10789038 and rs2796498 PRKAA2 genotypes were identified in all participants. The association between these single nucleotide polymorphisms and PHN susceptibility was evaluated in the dominant and recessive models. Haplotype analysis of patients with PHN and healthy controls was performed.
Results: The PHN patients were older than the healthy subjects (P < 0.05); however, the other clinical characteristics between two groups were not significantly different (all P > 0.05). Genotypes and allele frequencies differed significantly between PHN patients and healthy subjects in the rs10789038 polymorphism (P < 0.05), but not in rs2796498 (P > 0.05). In addition, the GG haplotype of rs10789038-rs2796498 correlated negatively with PHN occurrence in haplotype analysis (P < 0.05).
Conclusion: PHN occurrence may be related to the PRKAA2 rs10789038 A>G genetic polymorphism in the southwestern Chinese Han population.
Keywords: AMPK, PRKAA2, polymorphism, postherpetic neuralgia
Introduction
Herpes zoster is a viral skin disease that often causes severe pain.1 Postherpetic neuralgia (PHN) is defined as persistent or recurrent neuropathic pain (NeP) for more than three months after clinical healing of the rash.2 PHN incidence in patients with herpes zoster is approximately 5 to 30% and approximately 30–50% in patients with a course of more than one year.3 The daily life and mental state of patients are affected in severe cases.4 PHN is a typical form of NeP.5 NeP is clinically characterized by hyperalgesia, allodynia, and dysesthesia.6 Studies on NeP pathogenesis7–10 and treatment11,12 have made a lot of breakthrough progress. However, few studies have described the pathogenesis of PHN. Therefore, more studies on PHN pathogenesis and intervention measures are needed urgently.
Adenosine 5’-monophosphate (AMP)-activated protein kinase (AMPK) is an important protein that regulates cell energy homeostasis, as well as a hotspot in translational medicine research.13 In mammalian cells, AMPK may be involved in the regulation of cell energy balance.14 Our previous study found that abnormal energy metabolism in rat spinal cord astrocytes is closely related to NeP, and that AMPK plays an important role in abnormal cellular capacity metabolism.14 In our recent study, we also found that dexmedetomidine could alleviate NeP by activating AMPK in a mouse model of chronic constriction injury.15 AMPK consists of three subunits - α, β, and γ, the first of which is the functional unit.16 AMPKα has two subtypes, α1 and α2, encoded by the PRKAA1 and PRKAA2 genes, respectively.17–19 Studies have shown that drug transporter gene polymorphisms can affect the activity of their protein products and lead to abnormal drug metabolism.20,21 Therefore, AMPK gene polymorphisms may also affect AMPK activity and lead to disease. The current literature on PRKAA2 and diseases mainly includes diabetes-related studies.22,23 Other studies on AMPK polymorphisms have mainly focused on cancer and polycystic ovary syndrome,24–26 and no studies related to PHN have been reported. Thus, the purpose of this study was to investigate the relationship between PRKAA2 and PHN in the southwestern Chinese Han population.
Materials and Methods
Patients
This study included 132 PHN patients and 118 healthy subjects from the Chongqing Hospital of Traditional Chinese Medicine. The inclusion criteria were: PHN (persistent or recurrent pain for more than three months after clinical healing of the rash)2 and pain with a numerical rating scale ≥ 4. Patients with hepatic, renal, neuromuscular, or nerve system diseases were excluded from the study.
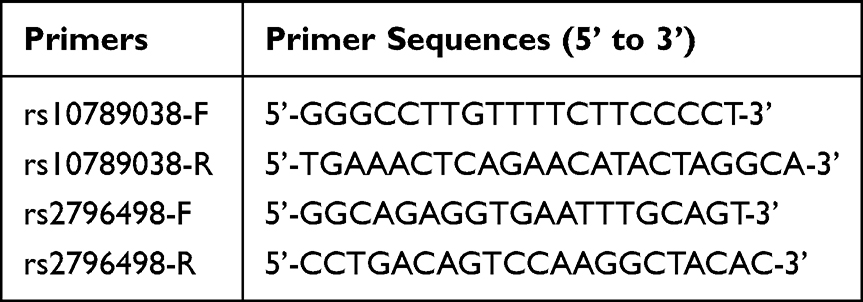 |
Table 1 PCR Primer Sequences of rs10789038 and rs2796498 Polymorphisms |
The clinical characteristics of PHN patients and healthy subjects are collected, including sex, age, body mass index (BMI), systolic blood pressure, diastolic blood pressure, triglycerides, total cholesterol, high-density lipoprotein cholesterol, low-density lipoprotein cholesterol, and fasting blood glucose. The research proposal was approved by the Ethics Committee of Chongqing Hospital of Traditional Chinese Medicine (2021-ky-67), and the study was conducted according to the Declaration of Helsinki principles. All subjects signed written consent forms. All subjects included in the experiment were of Han Chinese ethnicity.
DNA Extraction and Genotyping
Genomic DNA was isolated from peripheral venous blood using an SQ blood DNA kit (Cat. No. D5032-01, Omega, Norcross, Georgia, USA) according to the manufacturer’s protocol and stored at 4 °C (long-term storage at −80 °C). The rs10789038 and rs2796498 polymorphisms of PRKAA2 were used for genotyping. The primers used for the polymerase chain reaction (PCR) are listed in Table 1. The PCR reaction system consisted of 25 μL with less than 20 ng of DNA template, 0.4 μmol·L−1 of primer, and 12.5 μL PCR Mix [2X T5 Super PCR Mix (Basic), Cat. No. TSE008, Tsingke Biotechnology, Chongqing, China)]. The PCR reaction procedure was set according to the reference conditions of the kit, and the experiment was carried out after optimizing our preliminary experiment. The PCR products were sequenced by Tsingke Biotechnology Co. Ltd (Chongqing, China). The correlation between the rs10789038 and rs2796498 polymorphisms of PRKAA2 and PHN was analyzed according to the gene sequencing results.
Statistics
The Hardy–Weinberg equilibrium of rs10789038 and rs2796498 was determined using the Nielsen method.27 Genotypes and allele frequencies were compared between PHN patients and healthy subjects using the Chi-square test. Clinical data from PHN patients and healthy subjects were compared using the independent sample t-test. The correlation between each single nucleotide polymorphism (SNP) and PHN susceptibility was analyzed via binary logistic regression adjusted for age, sex, and BMI. The dominant and recessive models, as well as the homozygote and heterozygote comparisons were also assessed using binary logistic regression. Linkage disequilibrium and haplotypes were assessed as previously described.28 SPSS 26.0 (Chicago, Illinois, USA) was used to perform statistical analysis. Statistical significance was set at P < 0.05.
Results
Subject Characteristics
The clinical data of the PHN patients and healthy subjects are shown in Table 2. Overall, the PHN patients were older than the healthy subjects (P < 0.05); however, the other clinical data between the PHN patients and healthy subjects were not significantly different (all P >0.05).
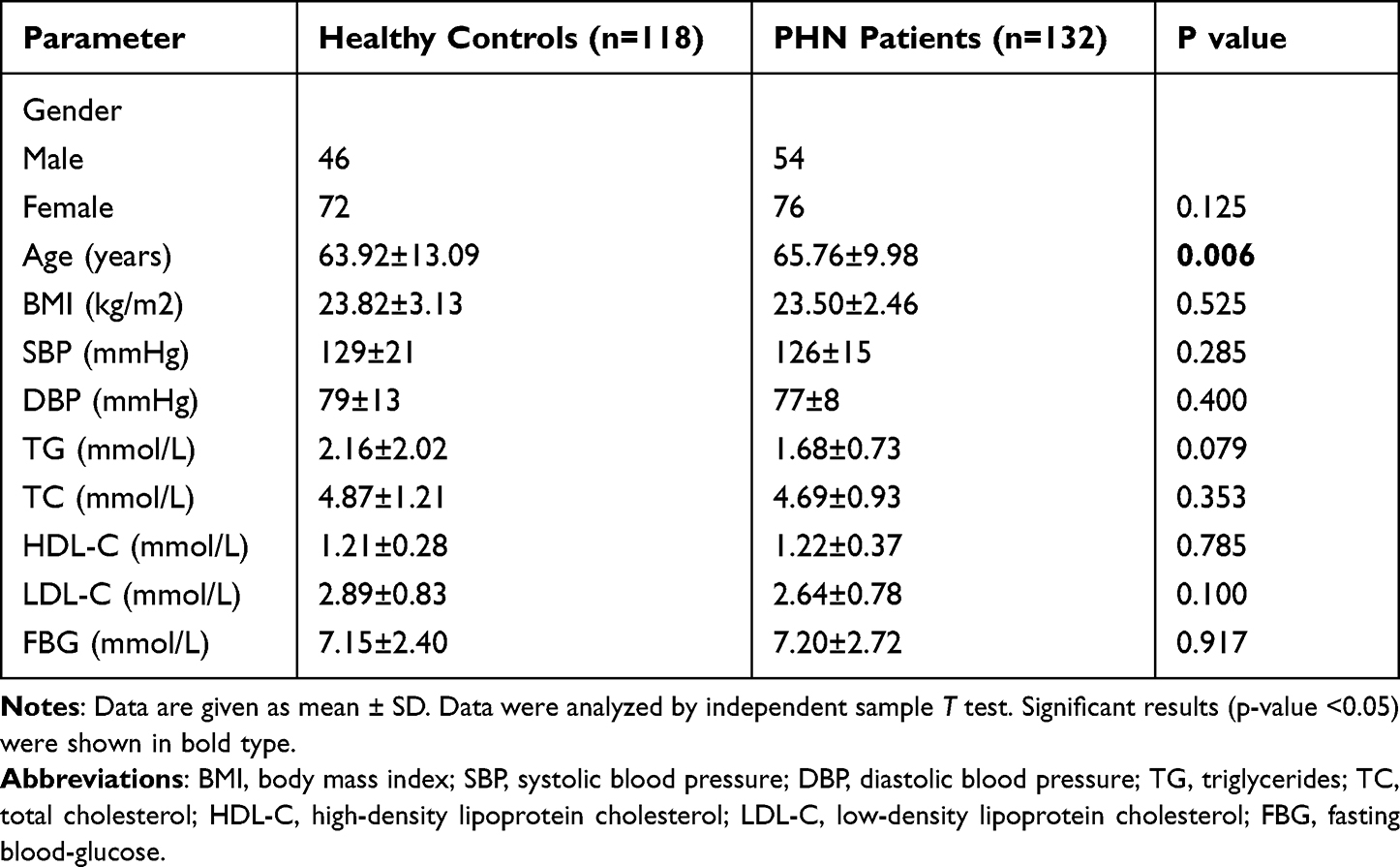 |
Table 2 The Demographic and Clinical Characteristics of PHN Patients and Healthy Subjects |
Association Study
The frequencies of the two SNPs are shown in Table 3. The rs10789038 and rs2796498 SNPs in PHN patients and healthy subjects conformed to the Hardy–Weinberg equilibrium (P > 0.05). This indicates that the population under investigation reached genetic equilibrium, showing that the data included in this study are credible. The genotype and allele frequencies of PHN patients and healthy subjects showed significant differences in rs10789038 (P < 0.05), but not in rs2796498 (P > 0.05).
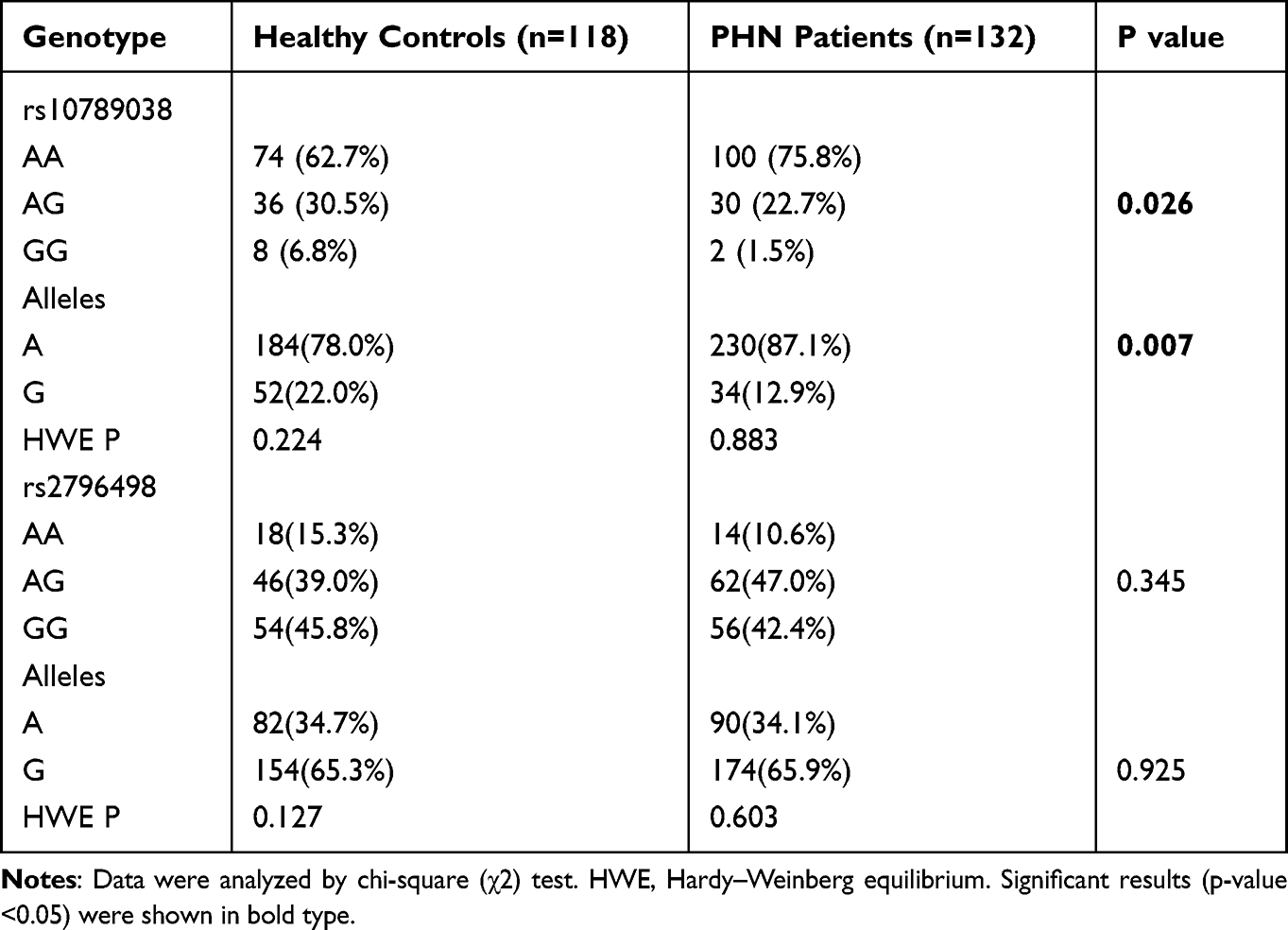 |
Table 3 Comparisons of Allelic Frequencies of PRKAA2 rs10789038 and rs2796498 Polymorphisms in PHN Patients and Healthy Subjects |
Table 4 shows the different analysis models of the effect of PRKAA2 gene polymorphisms on PHN. All statistical analyses were adjusted for sex, age, and BMI. Among all PRKAA2 rs10789038 polymorphism genotypes, GG correlated negatively with AA; thus, subjects with the AA genotype might be at a higher risk of experiencing PHN [GG vs AA: aOR (95% CI) = 0.153 (0.032–0.731); aP = 0.019]. In the dominant model, the GG genotype would significantly decrease the risk of developing PHN, compared with the GG+AG genotype [AA+AG vs GG: aOR (95% CI) = 0.190 (0.039–0.927), aP = 0.040]. Compared with the AA genotype, AG+GG significantly decreased the risk of developing PHN in the recessive models [AA vs AG+GG: aOR (95% CI) = 0.547 (0.314–0.951), aP = 0.033]. Compared with the AA genotype of rs10789038, GG significantly decreased the risk of developing PHN [AA vs GG: aOR (95% CI) = 0.172 (0.035–0.842), aP = 0.030]. However, compared with the homozygous AA genotype of rs10789038, AG showed no statistical difference in PHN incidence [AA vs AG: aOR (95% CI) = 0.633 (0.354–1.133), aP = 0.124]. However, no significant correlation was seen between each genotype and the occurrence of PHN in all model analyses of the PRKAA2 rs2796498 gene polymorphism (P > 0.05).
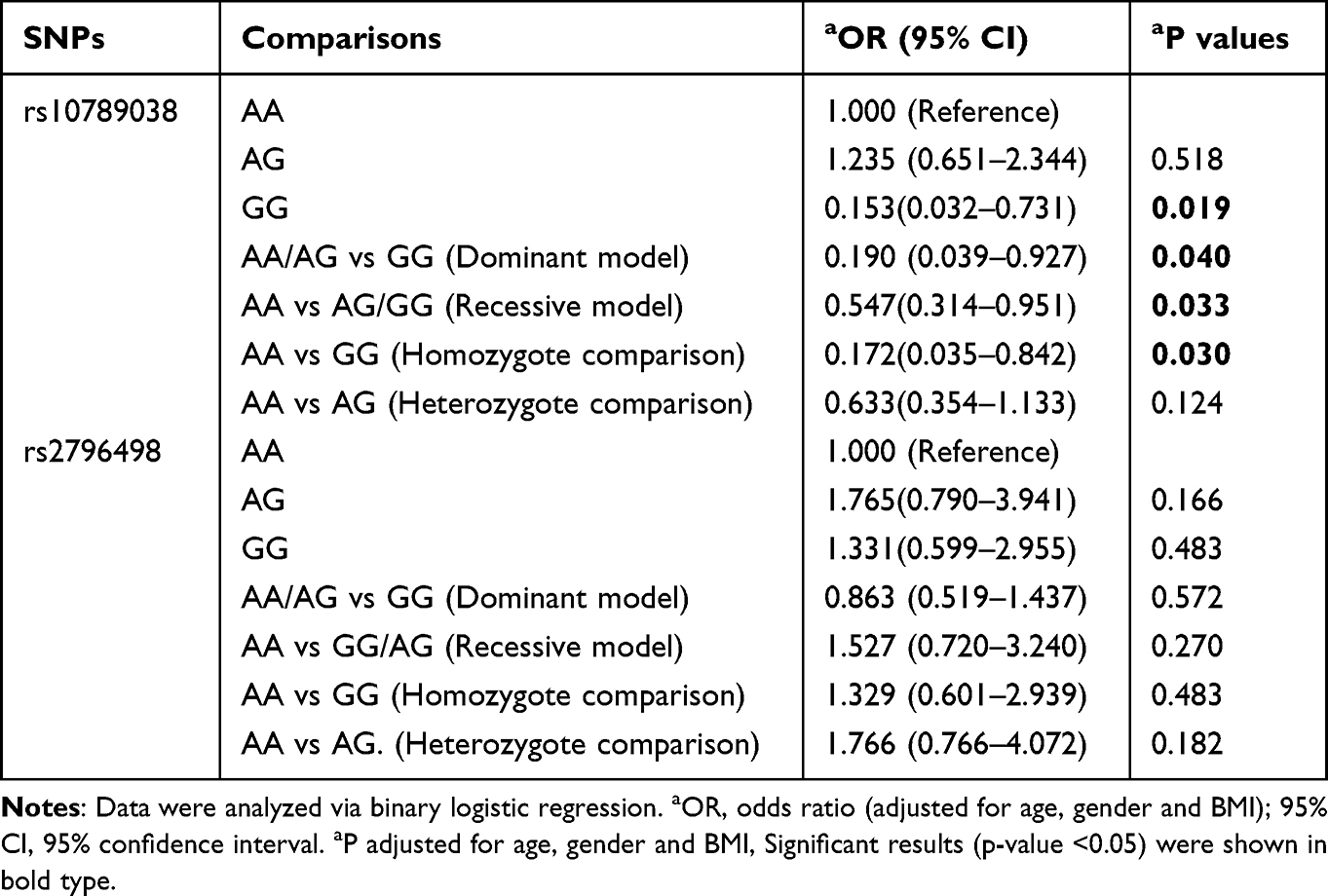 |
Table 4 The Association Between PRKAA2 Genetic Polymorphisms and the Risk of PHN (Adjusted for Age, Gender and BMI) |
Haplotype Analysis
The results showed no linkage disequilibrium between PRKAA2 rs10789038 and rs2796498. The D’ value was 0.783, and the r2 value was 0.243.
Haplotype analysis of PHN patients and healthy controls showed four haplotypes in the two SNPs, with significant differences between them (P < 0.05, Table 5). The GG haplotype was found only in healthy subjects, suggesting that it may reduce the risk of PHN (P < 0.001). There was no significant difference in the other three haplotypes between PHN patients and healthy subjects (P > 0.05).
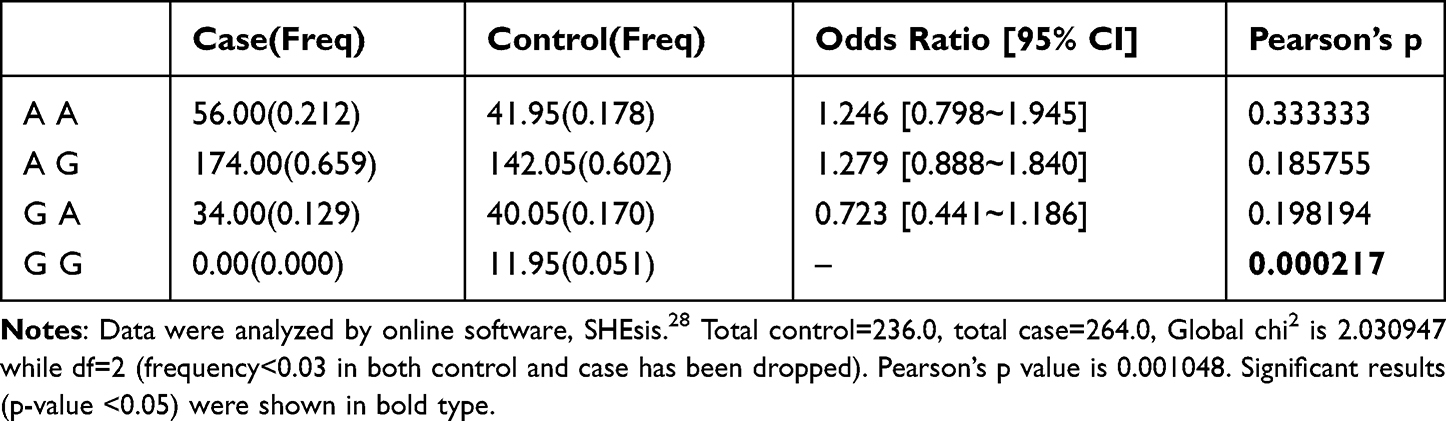 |
Table 5 Haplotype Analysis of PRKAA2 rs10789038 and rs2796498 with the Risk of PHN |
Discussion
This is the first study to investigate the relationship between AMPKα2 gene polymorphisms and PHN in the southwestern Chinese Han population.
We hypothesized that PRKAA2 gene polymorphisms may also cause changes in AMPK function, leading to unidentified changes in the body. A literature search revealed many studies on PRKAA2 gene polymorphisms. A large part of the research has focused on the relationship between PRKAA2 and diabetes mellitus and its related complications. For example, Li et al22 found that the rs10789038 and rs2796498 gene polymorphisms were related to the incidence of type 2 diabetes, and rs2796498 was associated with the occurrence of diabetic nephropathy. Keshavarz et al29 reported that PRKAA2 rs1418442 was slightly associated with the incidence of type 2 diabetes. Shen et al30 reported that the PRKAA2 rs2746342 polymorphism was significantly related to the occurrence of type 2 diabetes, while the rs2143754 polymorphism participates in fasting plasma glucose modulation. Horikoshi et al23 found a PRKAA2 gene polymorphism associated with insulin resistance and type 2 diabetes. In addition, studies have reported the association between PRKAA2 gene polymorphisms and other diseases. Hoffman et al25 demonstrated PRKAA2 gene polymorphism involvement in non-Hodgkin lymphoma pathogenesis and progression. However, variants in PRKAA2 genes were not associated with polycystic ovary syndrome.24 Campa et al26 found no statistically significant correlation between breast cancer risk and SNPs in PRKAA2 genes in the European Prospective Investigation on Cancer.
In the present study, the correlation between the PRKAA2 SNPs (rs10789038 and rs2796498) and PHN was investigated in the southwestern Chinese Han population. PHN patients were older than the healthy subjects. Although the healthy group was statistically significantly younger, the difference in age was only 1.8 years. Due to the small SD and large sample size, the mean was significantly different in the statistical test, but not large enough to assume that it influenced the makeup of the groups. Therefore, we believe that the difference was casual. In healthy subjects, the variation frequency of rs10789038 A>G in our study was 37.29%, which was similar to the 36.92% reported by Li et al.22 However, Li et al22 reported that the rs2796498 A>G gene variant frequency was 64.02%, which is different from the 84.75% that healthy subjects showed in this study. Gene-environment or gene-region interactions may be the reason for this difference. Our subjects were from southwest China (Chongqing City and surrounding provinces, including Guizhou, Sichuan, and Yunnan), while the participants of Li et al22 were from north China (Zhengzhou, Henan Province). In addition, the subjects in our study were from a specific ethnicity (Chinese Han population), while Li et al had a more diverse population. Therefore, the different genetic makeup (phenotype) may also affect studies. Li et al22 found that rs2796498 was closely related to diabetic nephropathy. However, we found no significant correlation between rs2796498 and PHN occurrence, which may be related to different gene frequencies. On the other hand, rs10789038 correlated significantly with PHN occurrence (P < 0.001). These results also indicate that the PRKAA2 rs10789038 GG genotype may have reduced PHN incidence, whereas rs2796498 did not show this correlation (adjusted for sex, age, and BMI).
Haplotype analysis enables the combined detection of interacting susceptibility alleles on complex traits and the effects of linkage disequilibrium variants with specific haplotypes.31,32 This approach is more effective than analyzing SNPs alone. In the present study, haplotype analysis of PHN patients and healthy controls showed four haplotypes in the two SNPs, and significant differences were found between the four haplotypes (P < 0.05). The GG haploid type was found only in healthy subjects, suggesting that it may reduce the risk of PHN (P < 0.001).
There are several strengths of this study. This study was the first to investigate the relationship between PRKAA2 and PHN in the Chinese Han population. In addition, we show the first evidence that the rs10789038 polymorphism is significantly associated with susceptibility to PHN.
However, this study also has some limitations. The sample size was relatively small, and we selected only two SNPs to conduct genotype analysis, which cannot represent all PRKAA2 gene polymorphisms. Second, the participants in this study were of southwestern Chinese Han ethnicity, a genetically homogenous group; therefore, this study may provide basis for further studies that evaluate the hypothesis tested here in other populations in China or in other ethnicities. Further research with larger sample sizes, more ethnic populations, and more polymorphic loci is required.
Conclusion
The present results indicate that PRKAA2 rs10789038 had a significant effect on PHN susceptibility, while rs2796498 did not affect southwestern Han Chinese patients. However, multi-locus, multi-ethnic, and large-sample studies are needed to evaluate the role of PRKAA2 polymorphisms in PHN.
Abbreviations
AMPK, adenosine 5’-monophosphate (AMP)-activated protein kinase; NeP, neuropathic pain; PHN, Postherpetic neuralgia; BMI, body mass index; SBP, systolic blood pressure; DBP, diastolic blood pressure; TG, triglycerides; TC, total cholesterol; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; FBG, fasting blood-glucose; PCR, polymerase chain reaction; OR, odds ratio; 95% CI, 95% confidence interval.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author, upon reasonable request.
Ethics Approval and Informed Consent
The research proposal was approved by the Ethics Committee of Chongqing Hospital of Traditional Chinese Medicine (2021-ky-67), and the study was conducted according to the Declaration of Helsinki principles. All subjects signed written consent forms.
Consent for Publication
Written consent was obtained from each of patients and healthy controls for all procedures and publication.
Acknowledgments
We thank all the patients and healthy subjects who participated in the trial. We also would like to thank Editage (www.editage.cn) for English language editing.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research has been granted by the National Natural Science Foundation of China (81870883), 2021 “Xinglin Scholar” Hospital Special Project of Chengdu University of Traditional Chinese Medicine (YYZX050), Chongqing Scientific Research Institution Performance Incentive and Guidance Special Project (cstc2021jxjl130022) and Natural Science Foundation of Chongqing (cstc2018jcyjAX0180).
Disclosure
The authors declared no conflict of interest.
References
1. Patil A, Goldust M, Wollina U. Herpes zoster: a review of clinical manifestations and management. Viruses. 2022;14(2):192. doi:10.3390/v14020192
2. Gan EY, Tian EA, Tey HL. Management of herpes zoster and post-herpetic neuralgia. Am J Clin Dermatol. 2013;14(2):77–85. doi:10.1007/s40257-013-0011-2
3. Kawai K, Gebremeskel BG, Acosta CJ. Systematic review of incidence and complications of herpes zoster: towards a global perspective. BMJ Open. 2014;4(6):e004833. doi:10.1136/bmjopen-2014-004833
4. Ebrahimiadib N, Yousefshahi F, Abdi P, Ghahari M, Modjtahedi BS. Ocular Neuropathic Pain: an Overview Focusing on Ocular Surface Pains. Clin Ophthalmol. 2020;14:2843–2854. doi:10.2147/OPTH.S262060
5. Finnerup NB, Haroutounian S, Kamerman P, et al. Neuropathic pain: an updated grading system for research and clinical practice. Pain. 2016;157(8):1599–1606. doi:10.1097/j.pain.0000000000000492
6. Meacham K, Shepherd A, Mohapatra DP, Haroutounian S. Neuropathic Pain: central vs. Peripheral Mechanisms. Curr Pain Headache Rep. 2017;21(6):28. doi:10.1007/s11916-017-0629-5
7. Fernandes V, Sharma D, Vaidya S, et al. Cellular and molecular mechanisms driving neuropathic pain: recent advancements and challenges. Expert Opin Ther Targets. 2018;22(2):131–142. doi:10.1080/14728222.2018.1420781
8. Tiwari V, Guan Y, Raja SN. Modulating the delicate glial-neuronal interactions in neuropathic pain: promises and potential caveats. Neurosci Biobehav Rev. 2014;45:19–27. doi:10.1016/j.neubiorev.2014.05.002
9. Shaw S, Uniyal A, Gadepalli A, et al. Adenosine receptor signalling: probing the potential pathways for the ministration of neuropathic pain. Eur J Pharmacol. 2020;889:173619. doi:10.1016/j.ejphar.2020.173619
10. Zhao X, Tang Z, Zhang H, et al. A long noncoding RNA contributes to neuropathic pain by silencing Kcna2 in primary afferent neurons. Nat Neurosci. 2013;16(8):1024–1031. doi:10.1038/nn.3438
11. Li Z, Tseng PY, Tiwari V, et al. Targeting human Mas-related G protein-coupled receptor X1 to inhibit persistent pain. Proc Natl Acad Sci U S A. 2017;114(10):E1996–E2005. doi:10.1073/pnas.1615255114
12. Uniyal A, Shantanu PA, Vaidya S, et al. Tozasertib Attenuates Neuropathic Pain by Interfering with Aurora Kinase and KIF11 Mediated Nociception. ACS Chem Neurosci. 2021;12(11):1948–1960. doi:10.1021/acschemneuro.1c00043
13. Trefts E, Shaw RJ. AMPK: restoring metabolic homeostasis over space and time. Mol Cell. 2021;81(18):3677–3690. doi:10.1016/j.molcel.2021.08.015
14. Cui J, Ou S, He WJ, Du L, Zhao YD, Ruan HZ. Prevention of extracellular ADP-induced ATP accumulation of the cultured rat spinal astrocytes via P2Y(1)-mediated inhibition of AMPK. Neurosci Lett. 2011;503(3):244–249. doi:10.1016/j.neulet.2011.08.045
15. Mu Y, Mei Y, Chen Y, et al. Perisciatic Nerve Dexmedetomidine Alleviates Spinal Oxidative Stress and Improves Peripheral Mitochondrial Dynamic Equilibrium in a Neuropathic Pain Mouse Model in an AMPK-Dependent Manner. Dis Markers. 2022;2022:6889676. doi:10.1155/2022/6889676
16. Carling D. The AMP-activated protein kinase cascade--a unifying system for energy control. Trends Biochem Sci. 2004;29(1):18–24. doi:10.1016/j.tibs.2003.11.005
17. Krishan S, Richardson DR, Sahni S. Gene of the month. AMP kinase (PRKAA1). J Clin Pathol. 2014;67(9):758–763. doi:10.1136/jclinpath-2014-202422
18. Lin L, Flisikowski K, Schwarzenbacher H, et al. Characterization of the porcine AMPK alpha 2 catalytic subunitgene (PRKAA2): genomic structure, polymorphism detection and association study. Anim Genet. 2010;41(2):203–207. doi:10.1111/j.1365-2052.2009.01971.x
19. Stapleton D, Mitchelhill KI, Gao G, et al. Mammalian AMP-activated protein kinase subfamily. J Biol Chem. 1996;271(2):611–614. doi:10.1074/jbc.271.2.611
20. Mei Y, Wang SY, Li Y, et al. Role of SLCO1B1, ABCB1, and CHRNA1 gene polymorphisms on the efficacy of rocuronium in Chinese patients. J Clin Pharmacol. 2015;55(3):261–268. doi:10.1002/jcph.405
21. Wang SY, Duan KM, Li Y, et al. Effect of quercetin on P-glycoprotein transport ability in Chinese healthy subjects. Eur J Clin Nutr. 2013;67(4):390–394. doi:10.1038/ejcn.2013.5
22. Li Q, Li C, Li H, et al. Effect of AMP-activated protein kinase subunit alpha 2 (PRKAA2) genetic polymorphisms on susceptibility to type 2 diabetes mellitus and diabetic nephropathy in a Chinese population. J Diabetes. 2018;10(1):43–49. doi:10.1111/1753-0407.12553
23. Horikoshi M, Hara K, Ohashi J, et al. A polymorphism in the AMPKalpha2 subunit gene is associated with insulin resistance and type 2 diabetes in the Japanese population. Diabetes. 2006;55(4):919–923. doi:10.2337/diabetes.55.04.06.db05-0727
24. Sproul K, Jones MR, Azziz R, Goodarzi MO. Association study of AMP-activated protein kinase subunit genes in polycystic ovary syndrome. Eur J Endocrinol. 2009;161(3):405–409. doi:10.1530/EJE-09-0245
25. Hoffman AE, Demanelis K, Fu A, Zheng T, Zhu Y. Association of AMP-activated protein kinase with risk and progression of non-Hodgkin lymphoma. Cancer Epidemiol Biomarkers Prev. 2013;22(4):736–744. doi:10.1158/1055-9965.EPI-12-1014
26. Campa D, Claus R, Dostal L, et al. Variation in genes coding for AMP-activated protein kinase (AMPK) and breast cancer risk in the European Prospective Investigation on Cancer (EPIC). Breast Cancer Res Treat. 2011;127(3):761–767. doi:10.1007/s10549-010-1269-1
27. Nielsen DM, Ehm MG, Weir BS. Detecting marker-disease association by testing for Hardy-Weinberg disequilibrium at a marker locus. Am J Hum Genet. 1998;63(5):1531–1540. doi:10.1086/302114
28. Li Z, Zhang Z, He Z, et al. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: update of the SHEsis. Cell Res. 2009;19(4):519–523. doi:10.1038/cr.2009.33
29. Keshavarz P, Inoue H, Nakamura N, Yoshikawa T, Tanahashi T, Itakura M. Single nucleotide polymorphisms in genes encoding LKB1 (STK11), TORC2 (CRTC2) and AMPK alpha2-subunit (PRKAA2) and risk of type 2 diabetes. Mol Genet Metab. 2008;93(2):200–209. doi:10.1016/j.ymgme.2007.08.125
30. Shen JZ, Ge WH, Fang Y, Liu H. A novel polymorphism in protein kinase AMP-activated catalytic subunit alpha 2 (PRKAA2) is associated with type 2 diabetes in the Han Chinese population. J Diabetes. 2017;9(6):606–612. doi:10.1111/1753-0407.12449
31. Zhao H, Pfeiffer R, Gail MH. Haplotype analysis in population genetics and association studies. Pharmacogenomics. 2003;4(2):171–178. doi:10.1517/phgs.4.2.171.22636
32. Morris RW, Kaplan NL. On the advantage of haplotype analysis in the presence of multiple disease susceptibility alleles. Genet Epidemiol. 2002;23(3):221–233. doi:10.1002/gepi.10200
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.