")
Back to Journals » Cancer Management and Research » Volume 11
Downregulation of CDH16 in Papillary Thyroid Cancer and Its Potential Molecular Mechanism Analysed by qRT-PCR, TCGA and in silico Analysis
Authors Li P, Wu Q, Sun Y, Pan X, Han Y, Ye B, Zhang Y, Dong J, Zheng Z
Received 2 September 2019
Accepted for publication 22 November 2019
Published 23 December 2019 Volume 2019:11 Pages 10719—10729
DOI https://doi.org/10.2147/CMAR.S229631
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Pihong Li,1,* Qiaolin Wu,2,* Yihan Sun,1,* Xiaoyu Pan,1 Yifan Han,1 Bing Ye,1 Yinlong Zhang,1 Jianda Dong,1 Zhouci Zheng1
1Departments of Neck Surgery, The Second Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, People’s Republic of China; 2Departments of Anesthesiology, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jianda Dong; Zhouci Zheng
Departments of Neck Surgery, The Second Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, People’s Republic of China
Email [email protected]; [email protected]
Objective: Thyroid cancer has the highest prevalence among the cancer types that affect the endocrine system; however, its molecular mechanisms are not yet determined. Cadherin-16 (CDH16) plays an important role in the tumorigenesis of human cancers, but its influence on papillary thyroid cancer (PTC) is poorly investigated. This study aimed to explore the role of CDH16 in PTC.
Methods: We performed quantitative real-time polymerase chain reaction to investigate CDH16 expression in PTC. The clinical significance of CDH16 expression in PTC was then evaluated using The Cancer Genome Atlas (TCGA) database. Bioinformatics analysis was also conducted to determine the potential molecular mechanisms of CDH16.
Results: CDH16 was remarkably downregulated in PTC tumors compared with that in corresponding normal thyroid tissues in the local and TCGA cohorts. This downregulation was associated with unfavorable clinicopathological features, including histological type, high tumor stage, aggressive lymph node metastasis (LNM), and advanced clinical stage. In addition, logistic analyses revealed that the reduced expression of CDH16 can aggravate the risk of LNM in PTC. Bioinformatics analysis indicated that the co-expressed CDH16 genes mainly participated in signaling the cancer-related pathways.
Conclusion: CDH16 is involved in PTC progression and acts as an LNM-related gene in PTC.
Keywords: CDH16, papillary thyroid cancer, lymph node metastasis, co-expression, mechanism
Introduction
Thyroid cancer has the highest prevalence among the cancers that affect the endocrine system worldwide. In 2017, approximately 56,870 cases of thyroid cancer were diagnosed in America, and approximately 2010 patients succumbed to the disease.1 In China, approximately 90,000 diagnosis with thyroid cancer and 6800 deaths were recorded in 2015.2 Papillary thyroid cancer (PTC) is the most common subtype of thyroid cancer and accounts for more than 80% of all the thyroid cancer cases.3,4
PTC patients have relatively better prognosis compared with other cancer patients. After proper surgical treatments and radiotherapy, more than 90% of the patients with PTC can have a 10-year survival.5 However, some patients might suffer from disease recurrence and death even after timely treatments.6 Lymph node metastasis (LNM) is a potential factor that can aggravate the risk of locoregional recurrence and mortality in patients with PTC.7–10 Recent studies revealed that several molecular biomarkers, such as BRAF11,12 TERT,13–15 PIK3CA16,17 can facilitate LNM to some extent. Despite the in-depth research and progress on PTC studies, comprehensive clinical evaluation and molecular mechanistic exploration remain lacking. Therefore, searching for novel functional biomarkers in PTC might provide preliminary data and a fundamental theoretical basis for the elucidation of PTC tumorigenesis.
Cadherins (CDHs) belong to a superfamily of glycoproteins that plays critical roles in calcium-dependent cell-to-cell adhesion.18,19 CDHs also participate in embryonic development, normal cell function, and tissue integrity preservation via homotypic and homophilic interactions between extracellular regions.20 Cadherin-16 (CDH16), a member of the CDH gene family and is localized to chromosome 16, might be involved in some certain diseases.21–25 However, studies on the clinical significance and regulatory mechanisms of CDH16 expression in PTC are required to verify this assumption. Further studies are necessary to identify the CDH16 expression in PTC, determine the regulatory mechanisms, and provide novel insights into PTC tumorigenesis.
In this study, we performed quantitative real-time polymerase chain reaction (qRT-PCR) to detect CDH16 expression in PTC tumors and corresponding normal thyroid tissues. The results were further validated using The Cancer Genome Atlas (TCGA) database. The relationship of CDH16 expression with clinicopathological features and LNM in PTC was also analyzed. Several bioinformatics analysis methods, including Gene Ontology (GO), Kyoto Encyclopedia of Gene and Genomes (KEGG) and protein–protein interaction (PPI), were performed to determine the potential molecular mechanisms of CDH16.
Materials and Methods
Clinical Samples
We collected 16 tissues of cases of clinical PTC tumors and corresponding noncancerous thyroid tissues during surgeries from February 2011 to June 2013 at the Department of Neck surgery of the Second Affiliated Hospital of Wenzhou Medical University. We used liquid nitrogen to snap-freeze these samples immediately after resection and then stored at −80°C for followed RNA isolation. Two senior pathologists were invited to review all the samples retrospectively and to confirm the histological diagnosis and to ensure abundant cancer content of the tumor using the double-blind method. All participants signed informed consent for the scientific use of biological material. This research protocol was approved by the Ethics Committee of the Second Affiliated Hospital of Wenzhou Medical University.
Clinicopathological Feature of Local Patients with PTC
This study involved 16 patients, including three men (18.8%) and 13 women (81.2%) (male-to-female ratio = 0.231). The patients were 27–75 years of age at diagnose with a mean age of 54.1 years. Among them, eight patients suffered from LNM. No significant difference was observed between the local and TCGA cohorts due to the small case series of the former. Detailed clinicopathological data are shown in Table 1.
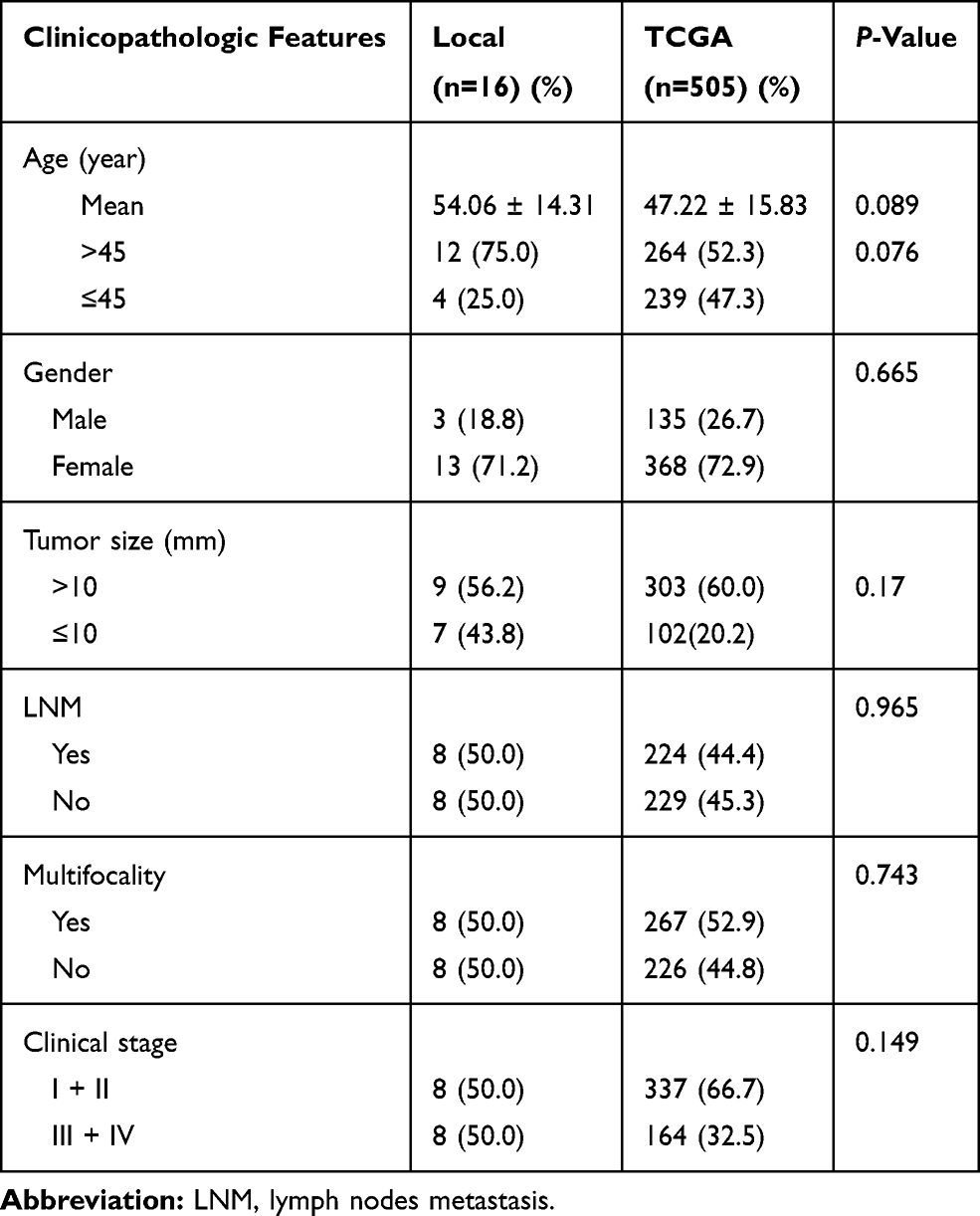 |
Table 1 The Clinicopathologic Features in the Local Cohort and in the TCGA cohort |
Detection of CDH16 Expression by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
We used TRIzol reagent (Life Technologies, Carlsbad, CA) to extract total RNA from the paired PTC tumors and noncancerous thyroid tissues according to the manufacturer’s indication, and then the RNA purity and concentration were measured by Plate reader Infinite 200 PRO (Tecan, Swiss). We used ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan) for cDNA preparation. qRT-PCR analysis of CDH16 expression was performed in triplicate using the Thunderbird SYBR qPCR Mix (Toyobo, Osaka, Japan) in the Applied Biosystems 7500 Real-time PCR system (Applied biosystems, Foster City, CA). We set the expression of GAPDH as an internal control. The primer sequences of CDH16 were as follows: forward 5ʹ- CCTCATCCTCATTTTCACC -3ʹ and reverse 5ʹ-GGGCTTCTACTCTGTCCTG -3ʹ.
Retrieval of the Cancer Genome Atlas (TCGA) Data
We downloaded the clinical information and corresponding RNA-seq data of CDH16, including 505 PTC tumors and 59 normal thyroid tissues, from TCGA database.
Retrieval of CDH16 Co-Expression Genes’ Data
To investigate the potential mechanisms of CDH16 expression in PTC tumors, we acquired CDH16-related co-expressed genes from the MEM and cBioPortal database. In MEM, the results of gene probe were extracted from MEM based on a P-value <0.0001. Pearson’s correlation analysis was used in cBioPortal data to evaluate the correlation between CDH16 and co-expressed genes. The present research included the genes with an absolute value of the Pearson’s correlation coefficient >0.4 in PTC. Then, the intersection results between MEM and cBioPortal were used for further study.
Bioinformatics Analyses
In this study, we used bioinformatics analyses to preliminarily discuss the intrinsic molecular mechanisms of CDH16 in PTC. The selected overlapping co-expressed genes in MEM and cBioPortal were used for GO and KEGG analyses in David v6.7 to elucidate the enrichment of genes in biological processes and signaling pathways. We also performed PPI analyses in STRING to explore proteins interaction association to hypothesize the potential pathways through which CDH16 participates in PTC tumorigenesis. Cytoscape 3.6.1 software was used to draw the functional networks.
Statistical Analyses
We used SPSS version 22.0 (IBM, New York, USA) to perform statistical analyses in the present study. The expression of CDH16 in PTC tumors and normal thyroid tissues detected by qRT-PCR were expressed as the means ± SD, and the Mann–Whitney U-test was used to determine significance. We used Student’s t-test to test the data on normal distribution. And chi-square test or Fisher’s exact test was used to elucidate the relationship between clinicopathological factors and CDH16 expression. We performed logistic regression analyses to evaluate the lymph node metastatic risk of CDH16. All P-values were two-sided, and a P-value of <0.05 was considered statistically significant. GraphPad Prism 5 (GraphPad Software, La Jolla, CA, USA) was used for graphs.
Results
CDH16 Expression in Patients with PTC
We detected the mRNA expression of CDH16 in 16 pairs of PTC tumors and corresponding noncancerous thyroid tissues by qRT-PCR to confirm the role of CDH16 in PTC. As shown in Figure 1A, CDH16 expression was significantly downregulated in the tumor samples compared with that in the corresponding noncancerous thyroid tissues (P < 0.05). This result was also validated by the TCGA database, which revealed that the CDH16 expression was lower in PTC samples than in normal thyroid tissues (Figure 1B). A receiver operator characteristic (ROC) curve was plotted to investigate the diagnostic value of CDH16. According to the TCGA database, 78.0% and 71.2% were the sensitivity and specificity rates, respectively, which can be used to distinguish the normal tissues from PTC tumors (Figure 1C).
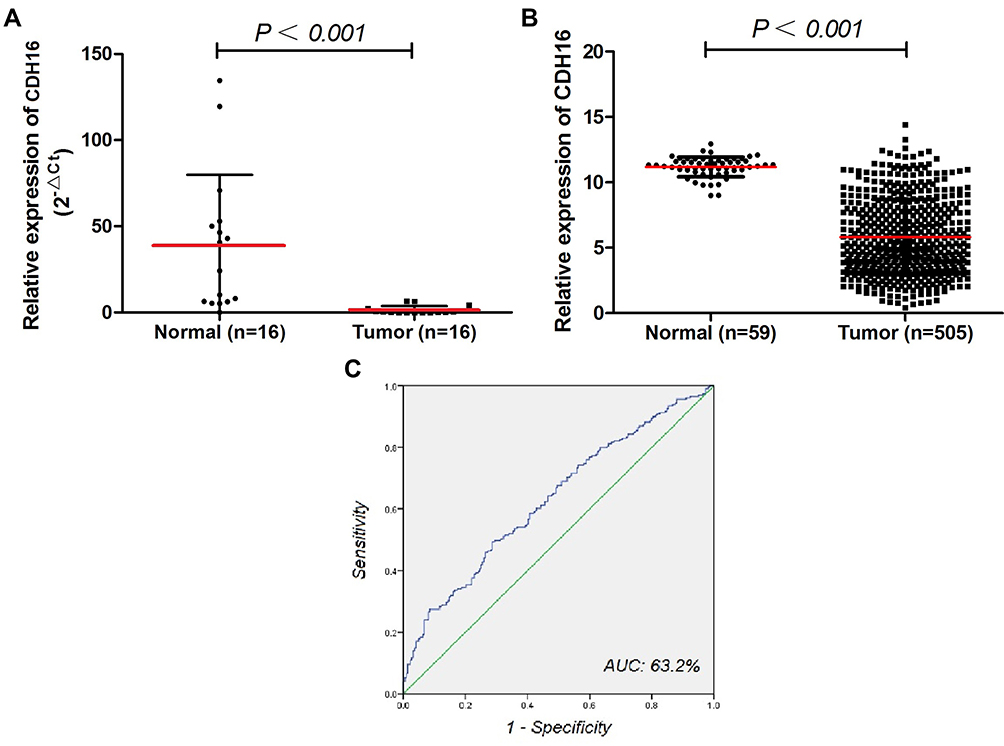 |
Figure 1 CDH16 expression was significantly downregulated in thyroid cancer samples compared with normal thyroid tissues in local and TCGA cohorts. Notes: (A) In the local cohort, CDH16 expression was examined by qRT-PCR in 16 paired PTC samples and adjacent normal tissues (Mann–Whitney U-test, P < 0.001). (B) The TCGA cohort contained 505 tumor samples and 59 normal tissue samples. (C) ROC curve for CDH16 expression to diagnose PTC in the TCGA cohort. |
Relationship Between CDH16 Expression and Clinicopathological Factors in PTC
We investigated the relationship between CDH16 expression and clinicopathological factors. The 505 patients with PTC from the TCGA database were divided into low (n=252) and high CDH16 expression (n=253) groups on the basis of the median value. As shown in Table 2, low CDH16 expression was significantly related to histological type (P < 0.001), signified large tumor size (P = 0.036), high tumor stage (P = 0.001), aggressive LNM (P = 0.001), and advanced clinical stage (P = 0.029) (Table 2). Compared with the high expression group, the low CDH16 expression group was inclined to the classical subtype (P < 0.001, Table 2). However, no significant difference was found among CDH16 expression and age, gender, and multifocality (P > 0.05).
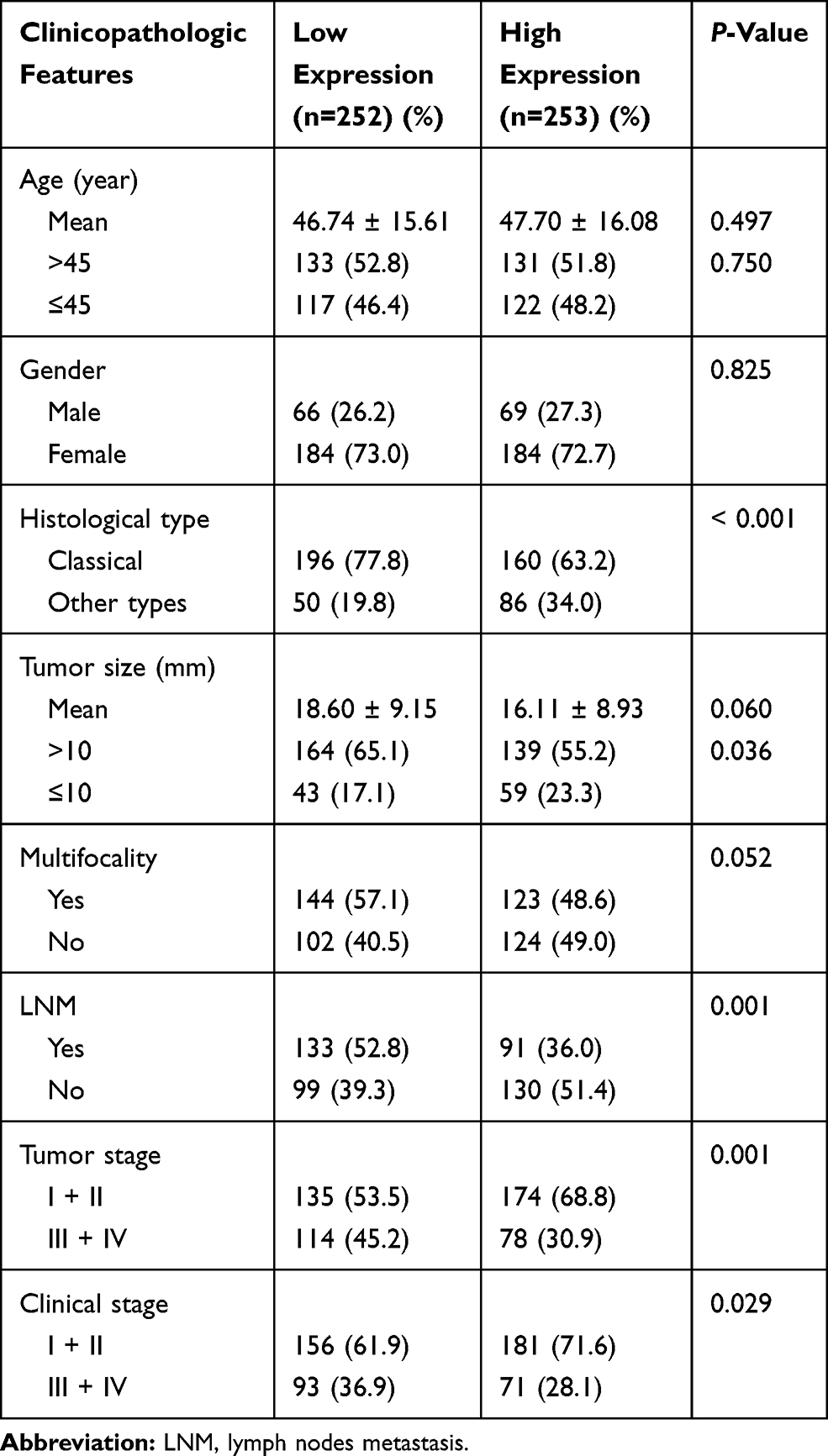 |
Table 2 The Relationship Between CDH16 Expression and Clinicopathologic Features in the TCGA Cohort |
Low CDH16 Expression Could Enhance the Risk of LNM in PTC
The association between CDH16 expression and LNM in PTC was further investigated through logistic regression analyses. Univariate logistic regression analysis revealed that age (OR = 0.614, 95% CI = 0.424–0.891, P = 0.01), gender (OR = 1.541, 95% CI = 1.016–2.337, P = 0.042), histological type (OR = 0.377, 95% CI =0.239–0.549, P < 0.001), clinical stage (OR = 3.429, 95% CI = 2.270–5.180, P < 0.001) and CDH16 expression (OR = 0.521, 95% CI = 0.359–0.757, P = 0.001) were significant variables for LNM in PTC (Table 3). Multivariate logistic analysis using these five parameters also indicated that age (OR = 0.042, 95% CI = 0. 015– 0.118, P < 0.001), histological type (OR = 0.331, 95% CI = 0.196–0.558, P < 0.001) and CDH16 expression (OR = 0.574, 95% CI = 0.369–0.893, P = 0.014) were protective factors for LNM in PTC, whereas clinical stage (OR = 47.93, 95% CI = 16.43–139.8, P < 0.001) was an independent high-risk factor for LNM (Table 4). In summary, the downregulation of CDH16 expression can aggravate the risk of LNM in patients with PTC.
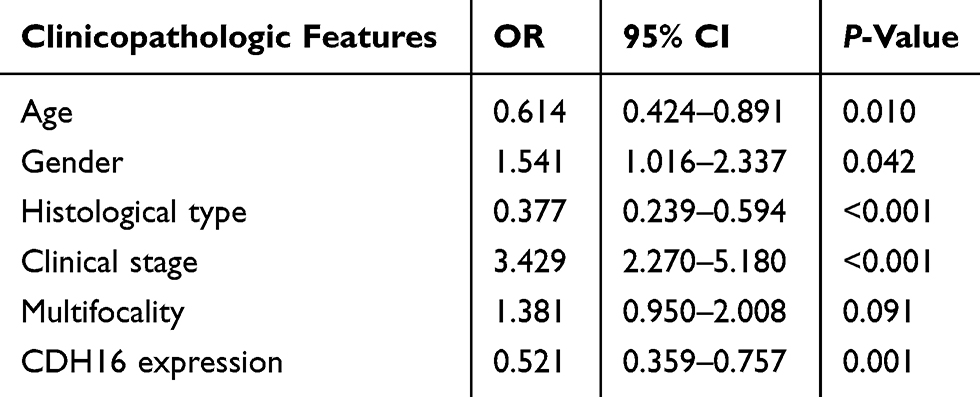 |
Table 3 Univariate Logistic Regression Analysis for the Lymph Node Metastatic Risk |
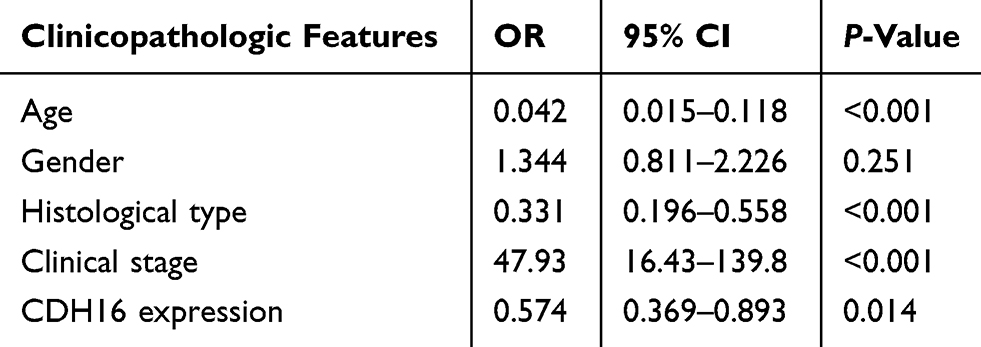 |
Table 4 Multivariate Logistic Regression Analysis for the Lymph Node Metastatic Risk |
CDH16 Co-Expressed Genes and Bioinformatics Analyses
Based on MEM, 5000 of the most significant CDH16 co-expressed mRNA genes were selected. The data from the cBioPortal for Cancer Genomics showed 20,033 CDH16 co-expressed mRNA genes in the PTC patients. A total of 1495 genes were obtained by excluding the genes with an absolute value of the Pearson’s correlation coefficient <0.4. The intersection between the results of the two databases was collected, and 340 reliable co-expressed genes were extracted for further study. GO and KEGG analyses were performed to determine the biological processes involved in PTC tumorigenesis. The results of the GO term analysis (Table 5) revealed that the genes are involved in the following: (1) biological processes (Figure 2A), including the multicellular organismal process, the system development, the anatomical structure development, the developmental process, the organ development, etc.; (2) cellular components (Figure 2B), such as plasma membrane part, integral to plasma membrane, cell fraction, and intrinsic to plasma membrane; and (3) molecular functions (Figure 2C), such as channel activity, passive transmembrane transporter activity, serine-type endopeptidase activity, and transporter activity . The results of the KEGG pathway analysis showed that CDH16 participated in the functional regulation of six signaling pathways (Table 6). The relatively important pathways include tryptophan metabolism, tyrosine metabolism, and pathways in cancer (Figure 3). PPI analyses were performed and the interaction nodes are shown in Figure 4. The top 10 nodes with the highest overall scores are listed in Table 7. We also focused on the genes enriched in “pathways in cancer”. Ten interaction nodes were obtained based on the results of PPI analyses (Figure 5). The findings suggested that WNT11, FZD9, FZD8, AXIN1, RXRG, all of which are common genes in cancer-related pathways, might play important connective roles in cancer initialization (Table 8) and therefore were further analyzed. According to the TCGA database, the results of Pearson’s correlation analyses demonstrated that AXIN1, RXRG exhibited a potential negative correlation with CDH16 expression, whereas WNT11, FZD9, FZD8 exhibited a possible positive correlation with CDH16 (Figure 6)
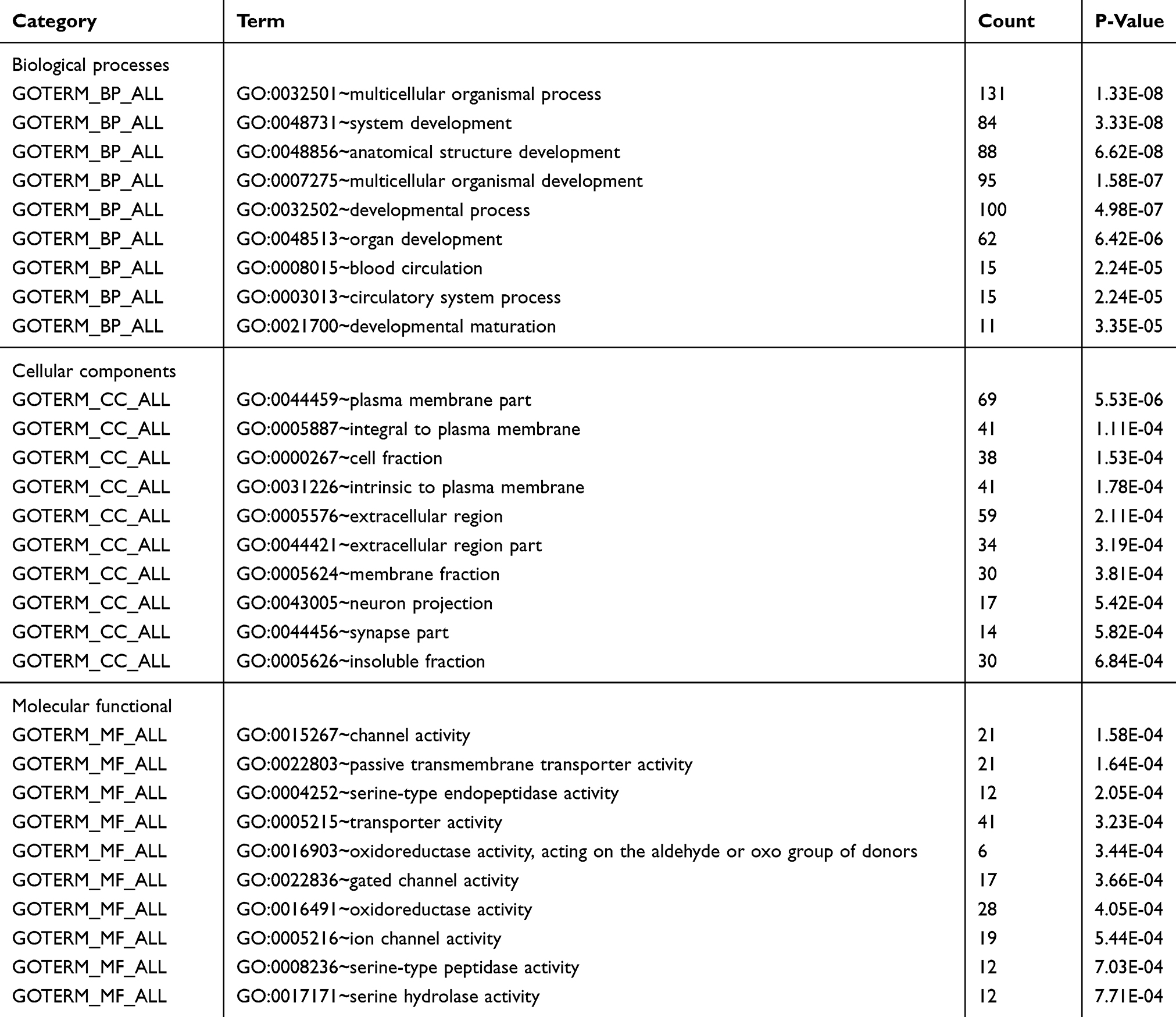 |
Table 5 The Top 10 Most Significant Items of Gene Ontology (GO) Terms of the Co-Expression Genes of CDH16 in PTC |
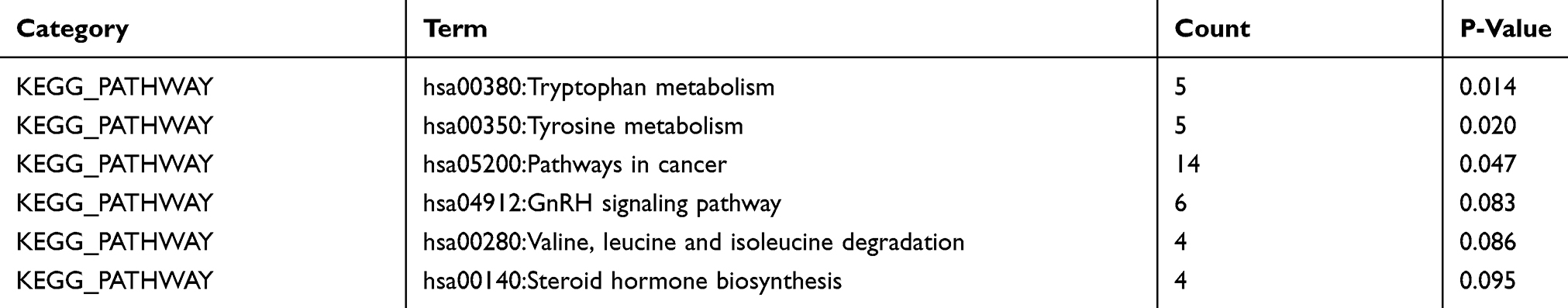 |
Table 6 KEGG Pathways Enriched by Co-Expression Genes of CDH16 in PTC |
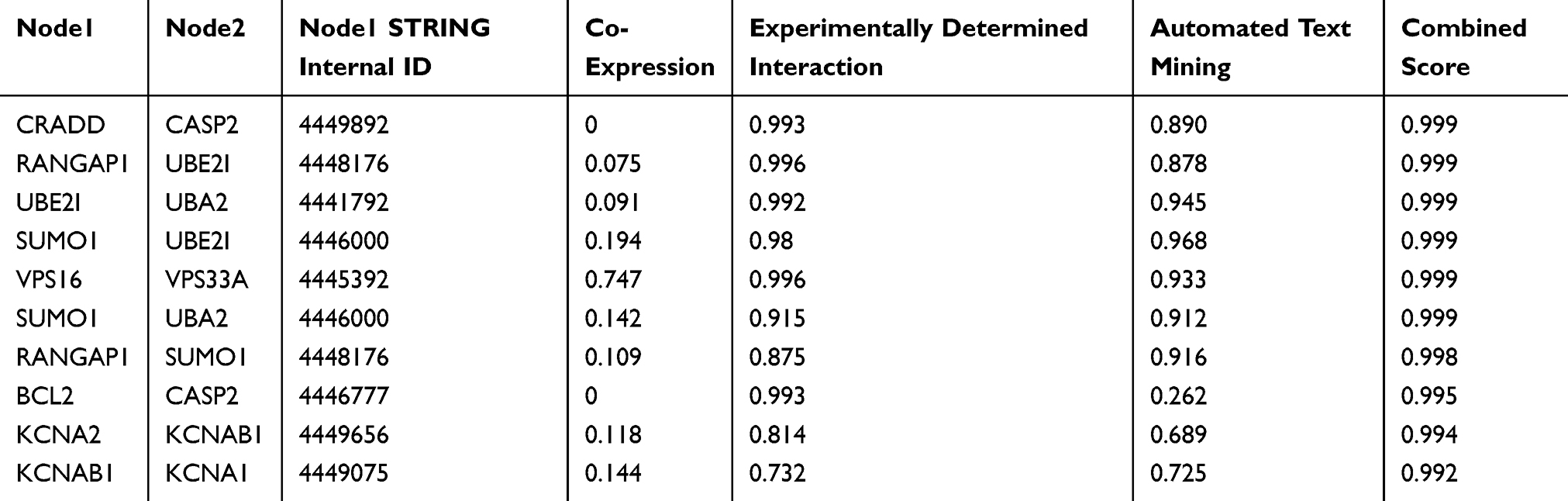 |
Table 7 The Top 10 with Combined Co-Expression Score That the Protein–protein Interaction (PPI) of CDH16 Co-Expression Genes in PTC Enriched in Significant Signaling Pathways |
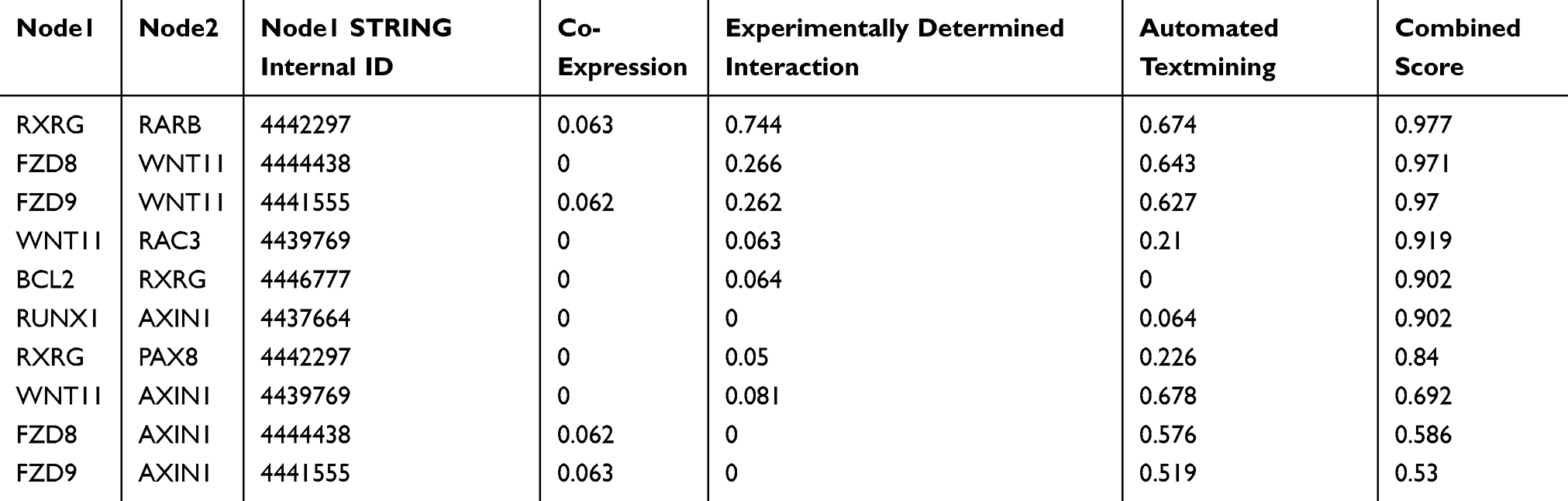 |
Table 8 The Top 10 with Combined Co-Expression Score That the Protein–protein Interaction (PPI) of CDH16 Co-Expression Genes in PTC Enriched in Pathways in Cancer |
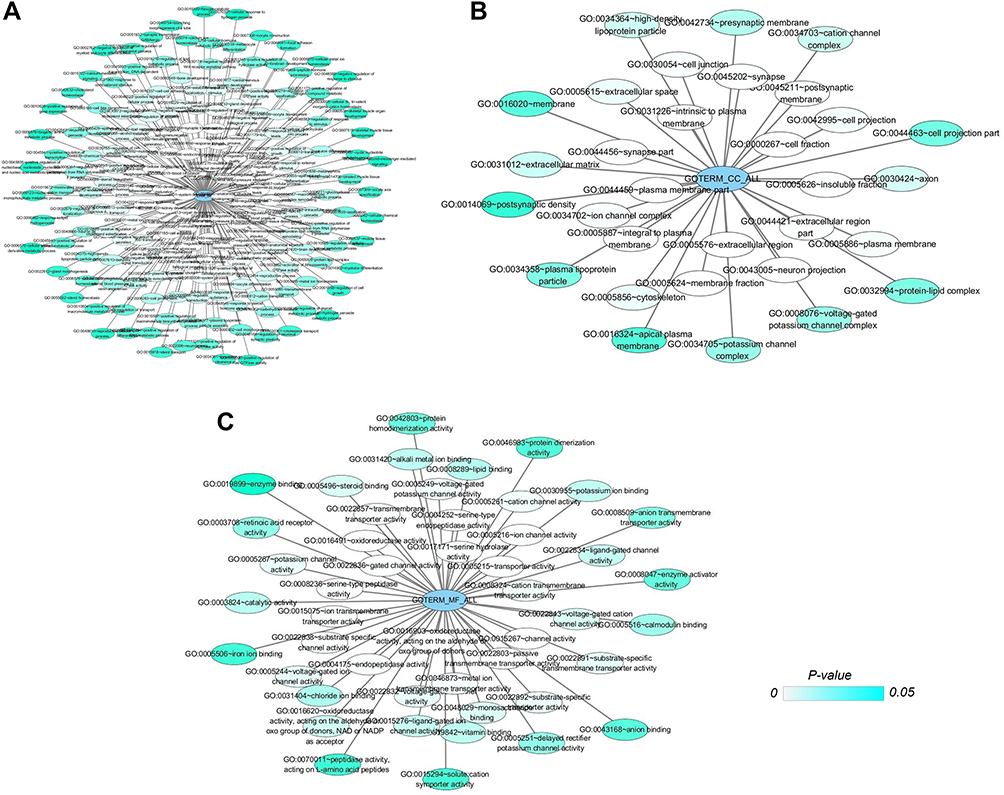 |
Figure 2 Networks of gene ontology (GO) terms of CDH16 co-expressed genes in PTC. Notes: (A) Network of biological processes (BPs) of CDH16 co-expressed genes. (B) Network of cellular components (CCs) of CDH16 co-expressed genes. (C) Network of molecular functions (MFs) of CDH16 co-expressed genes. In those networks, only terms with statistically significant (P < 0.05) are displayed. Each node represents a GO term, and the node color indicates the P-value of a GO term. |
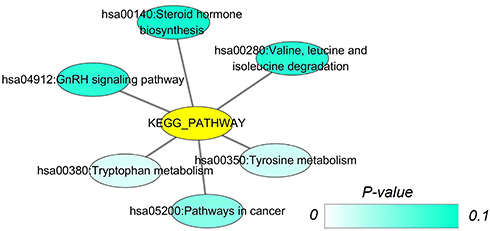 |
Figure 3 Network mappings of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways of CDH16 co-expressed genes in PTC. Notes: In this network, all terms related are displayed. Each node represents a KEGG term, and the node color indicates the P-value of a KEGG term. |
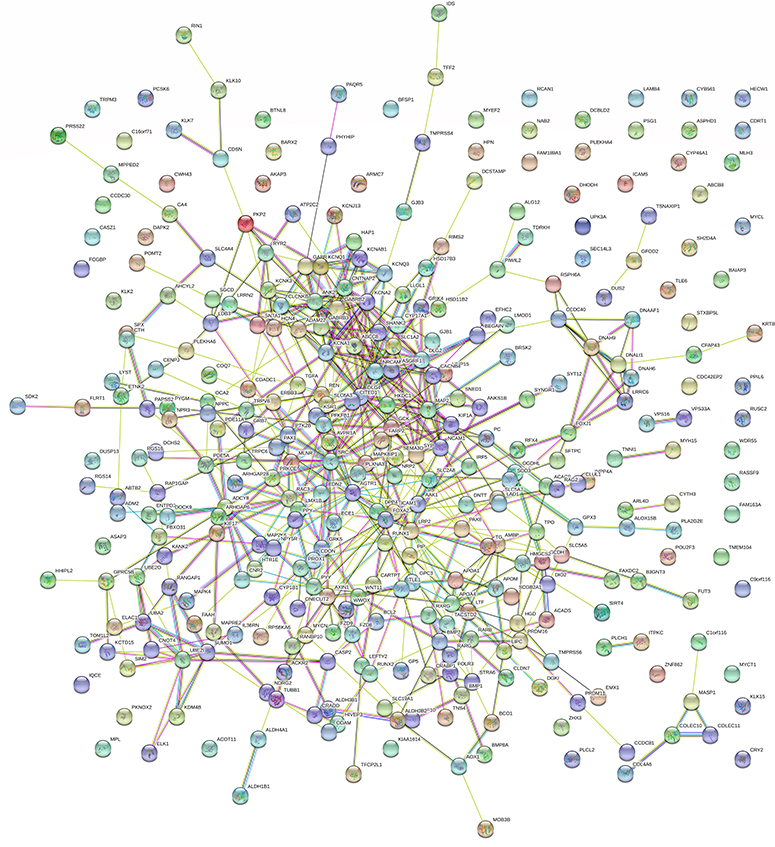 |
Figure 4 Network of protein–protein interaction (PPI) analyses of CDH16 co-expressed genes in PTC. |
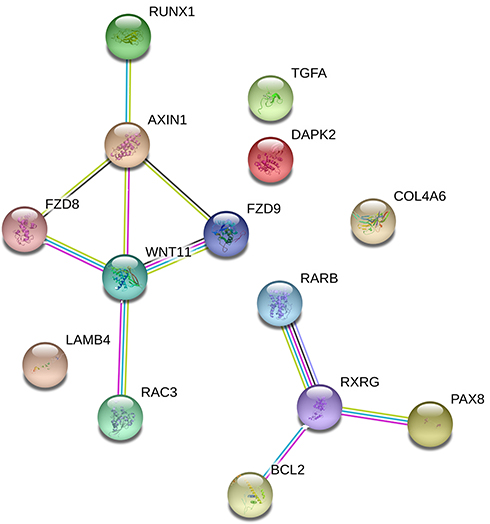 |
Figure 5 PPI network analyses of genes coexpressed in CDH16 in pathways in cancer. |
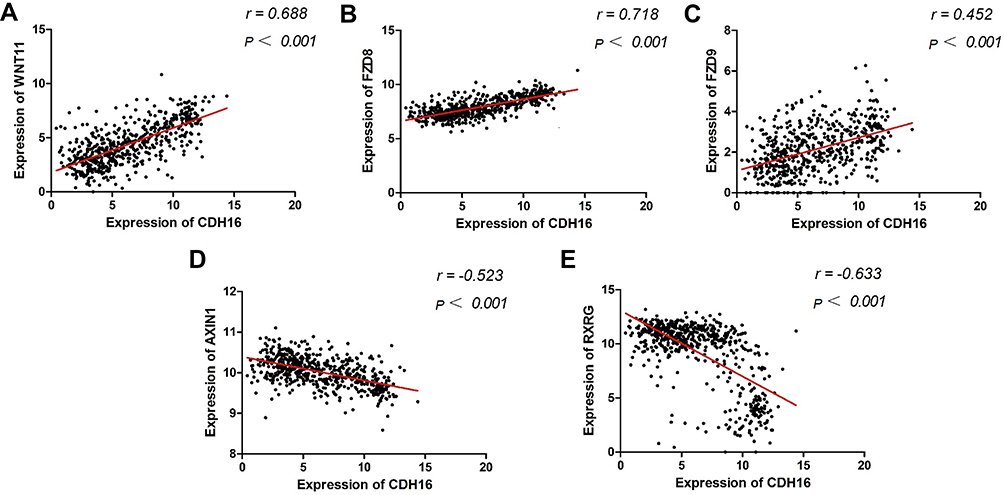 |
Figure 6 Correlations between the hub genes and the expression of CDH16 in PTC. Notes: (A) Correlation between expression of WNT11 and expression of CDH16 in PTC. r= 0.688, P < 0.001. (B) Correlation between expression of FZD8 and expression of CDH16 in PTC. r= 0.718, P < 0.001. (C) Correlation between expression of FZD9 and expression of CDH16 in PTC. r= 0.452, P < 0.001. (D) Correlation between expression of AXIN1 and expression of CDH16 in PTC. r= −0.523, P < 0.001. (E) Correlation between expression of RXRG and expression of CDH16 in PTC. r= −0.633, P < 0.001. |
Discussion
Thyroid cancer, which is one of the most common endocrine malignancies, has become the fastest growing type of cancer worldwide.1 This disease can be classified into several different subtypes according to molecular etiology.26,27 Patients with PTC have a statistically better prognosis and longer survival than those with other subtypes.4 However, the high incidence of LNM in PTC patients can significantly increase the risk of locoregional recurrence and mortality.7,8 Searching for potential biomarkers can provide new clinical insights into predicting the metastasis status of patients with PTC.
CDH16 plays an indispensable role in human development and growth.18,20 Thedieck et al found that CDH16 can facilitate tubulogenesis during human renal development and can be detected in the distal tubules of adults’ kidneys.23 Moreover, Morizane et al further proved that CDH16-positive cells derived from embryonic stem cells can reproduce tubular structures and differentiate into renal tubular cells.28 In addition, the CDH16 expression is associated with the fully differentiated state of the thyroid cells29 and partially influences the thyroid follicular polarity.30 However, the abnormal CDH16 expression can lead to tumorigenesis. Thedieck et al reported CDH16 downregulation in renal cell carcinoma and suggested the role of CDH16-related cell adhesion molecule in tumor suppression.23 As for the thyroid cancer, Cali et al confirmed that the downregulated CDH16 expression can be used as a marker to monitor thyroid cancer.25 In the present study, CDH16 expression was significantly downregulated in PTC. This finding is consistent with previous studies on human cancers indicating that CDH16 serves as a tumor suppressor in different kinds of human cancer.23,25 To the best of our knowledge, our study is the first to report the significant role of CDH16 in PTC.
We first reported that CDH16 plays an important role in LNM in PTC. qRT-PCR detection and TCGA RNA-Seq data were used to confirm that the CDH16 expression was significantly downregulated in PTC tumor tissues compared with that in noncancerous thyroid tissues. In addition, this downregulation of CDH16 was associated with unfavorable clinicopathological factors, such as aggressive LNM and high clinical stage. Subsequent logistic regression analyses showed that the downregulated expression of CDH16 can be an independent high-risk factor for LNM in PTC patients. The results confirmed that CDH16 acted as a tumor suppressor gene in PTC and was involved in the progression of LNM.
The molecular mechanisms of CDH16 in PTC were also explored in this study. We used GO and KEGG analyses to compare the enrichment of CDH16 co-expressed genes in PTC patients. The results suggested that the co-expressed genes play important roles in biological processes, cellular components, and molecular functions. According to the result of the KEGG pathways analysis, the genes were more concentrated on the pathways in cancer. The expression of CDH16 was positively related to WNT11, FZD9, FZD8 and negatively related to AXIN1 and RXRG. In conclusion, CDH16 performs regulatory functions in PTC through complex protein–protein interaction with the aforementioned genes.
Despite the substantial findings, this research still exhibits some limitations. First, the findings are mainly theoretical data that should be validated through in vitro and in vivo experiments. Second, the regulatory mechanism between CDH16 and its co-expressed genes should be further investigated.
In conclusion, CDH16 expression was downregulated in PTC tissues. The low expression of CDH16 can aggravate the risk of LNM in PTC. Moreover, CDH16 can collaborate with co-expressed genes, such as RXRG, WNT11 and so on, to regulate the pathways in cancer to change the LNM status. This research provides reliable molecular theories for further studies on the molecular mechanisms of CDH16 in PTC.
Ethical Approval
Ethical approval for this study was obtained from the Ethical Committee of the Second Affiliated Hospital of Wenzhou Medical University.
Acknowledgment
This work was supported by a grant from the Wenzhou Science and Technology Project (No. Y20180224).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.21387
2. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
3. Morris LG, Tuttle RM, Davies L. Changing trends in the incidence of thyroid cancer in the United States. JAMA Otolaryngol Head Neck Surg. 2016;142(7):709–711. doi:10.1001/jamaoto.2016.0230
4. Xing M. Molecular pathogenesis and mechanisms of thyroid cancer. Nat Rev Cancer. 2013;13(3):184–199. doi:10.1038/nrc3431
5. Leboulleux S, Rubino C, Baudin E, et al. Prognostic factors for persistent or recurrent disease of papillary thyroid carcinoma with neck lymph node metastases and/or tumor extension beyond the thyroid capsule at initial diagnosis. J Clin Endocrinol Metab. 2005;90(10):5723–5729. doi:10.1210/jc.2005-0285
6. Burns WR, Zeiger MA. Differentiated thyroid cancer. Semin Oncol. 2010;37(6):557–566. doi:10.1053/j.seminoncol.2010.10.008
7. Kim SK, Kwon AY, Back K, et al. Predictive factors of lymph node metastasis in follicular variant of papillary thyroid carcinoma. Ann Surg Oncol. 2017;24(9):2617–2623. doi:10.1245/s10434-017-5912-5
8. Blanchard C, Brient C, Volteau C, et al. Factors predictive of lymph node metastasis in the follicular variant of papillary thyroid carcinoma. Br J Surg. 2013;100(10):1312–1317. doi:10.1002/bjs.2013.100.issue-10
9. Lee YM, Sung TY, Kim WB, Chung KW, Yoon JH, Hong SJ. Risk factors for recurrence in patients with papillary thyroid carcinoma undergoing modified radical neck dissection. Br J Surg. 2016;103(8):1020–1025. doi:10.1002/bjs.10144
10. Liu FH, Kuo SF, Hsueh C, Chao TC, Lin JD. Postoperative recurrence of papillary thyroid carcinoma with lymph node metastasis. J Surg Oncol. 2015;112(2):149–154. doi:10.1002/jso.23967
11. Xing M. BRAF mutation in thyroid cancer. Endocr Relat Cancer. 2005;12(2):245–262. doi:10.1677/erc.1.0978
12. Ciarrocchi A, Cavuto S, Piana S. BRAF V600E mutation and papillary thyroid cancer. JAMA. 2013;310(5):534. doi:10.1001/jama.2013.8589
13. Blasco MA. Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet. 2005;6(8):611–622. doi:10.1038/nrg1656
14. Bu R, Siraj AK, Divya SP, et al. Telomerase reverse transcriptase mutations are independent predictor of disease-free survival in Middle Eastern papillary thyroid cancer. Int J Cancer. 2017;142(10).
15. Jin L, Chen E, Dong S, et al. BRAF and TERT promoter mutations in the aggressiveness of papillary thyroid carcinoma: a study of 653 patients. Oncotarget. 2016;7(14):18346–18355. doi:10.18632/oncotarget.7811
16. Abubaker J, Jehan Z, Bavi P, et al. Clinicopathological analysis of papillary thyroid cancer with PIK3CA alterations in a Middle Eastern population. J Clin Endocrinol Metab. 2008;93(2):611–618. doi:10.1210/jc.2007-1717
17. Garcia-Rostan G, Costa AM, Pereira-Castro I, et al. Mutation of the PIK3CA gene in anaplastic thyroid cancer. Cancer Res. 2005;65(22):10199–10207. doi:10.1158/0008-5472.CAN-04-4259
18. Hulpiau P, van Roy F. Molecular evolution of the cadherin superfamily. Int J Biochem Cell Biol. 2009;41(2):349–369. doi:10.1016/j.biocel.2008.09.027
19. Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell. 1996;84(3):345–357. doi:10.1016/S0092-8674(00)81279-9
20. Angst BD, Marcozzi C, Magee AI. The cadherin superfamily: diversity in form and function. J Cell Sci. 2001;114(Pt 4):629–641.
21. Berx G, van Roy F. Involvement of members of the cadherin superfamily in cancer. Cold Spring Harb Perspect Biol. 2009;1(6):a003129. doi:10.1101/cshperspect.a003129
22. Stemmler MP. Cadherins in development and cancer. Mol Biosyst. 2008;4(8):835–850. doi:10.1039/b719215k
23. Thedieck C, Kuczyk M, Klingel K, Steiert I, Muller CA, Klein G. Expression of Ksp-cadherin during kidney development and in renal cell carcinoma. Br J Cancer. 2005;92(11):2010–2017. doi:10.1038/sj.bjc.6602597
24. Kuehn A, Paner GP, Skinnider BF, et al. Expression analysis of kidney-specific cadherin in a wide spectrum of traditional and newly recognized renal epithelial neoplasms: diagnostic and histogenetic implications. Am J Surg Pathol. 2007;31(10):1528–1533. doi:10.1097/PAS.0b013e318058818c
25. Cali G, Gentile F, Mogavero S, et al. CDH16/Ksp-cadherin is expressed in the developing thyroid gland and is strongly down-regulated in thyroid carcinomas. Endocrinology. 2012;153(1):522–534. doi:10.1210/en.2011-1572
26. Wang O, Zheng Z, Wang Q, et al. ZCCHC12, a novel oncogene in papillary thyroid cancer. J Cancer Res Clin Oncol. 2017;143(9):1679–1686. doi:10.1007/s00432-017-2414-6
27. Lloyd RV, Buehler D, Khanafshar E. Papillary thyroid carcinoma variants. Head Neck Pathol. 2011;5(1):51–56. doi:10.1007/s12105-010-0236-9
28. Morizane R, Monkawa T, Fujii S, et al. Kidney specific protein-positive cells derived from embryonic stem cells reproduce tubular structures in vitro and differentiate into renal tubular cells. PLoS One. 2014;8(6):e64843. doi:10.1371/journal.pone.0064843
29. de Cristofaro T, Di Palma T, Fichera I, et al. An essential role for Pax8 in the transcriptional regulation of cadherin-16 in thyroid cells. Mole Endocrinol. 2012;26(1):67–78. doi:10.1210/me.2011-1090
30. Koumarianou P, Gomez-Lopez G, Santisteban P. Pax8 controls thyroid follicular polarity through cadherin-16. J Cell Sci. 2017;130(1):219–231. doi:10.1242/jcs.184291
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.