")
Back to Journals » International Journal of General Medicine » Volume 15
DOCK4 as a Potential Biomarker Associated with Immune Infiltration in Stomach Adenocarcinoma: A Database Analysis
Authors Lu Y, Yu J, Dong Q, Du Y, Liang Z
Received 14 February 2022
Accepted for publication 29 June 2022
Published 11 July 2022 Volume 2022:15 Pages 6127—6143
DOI https://doi.org/10.2147/IJGM.S357096
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Scott Fraser
Yi Lu,1 Jiaxi Yu,1 Qiuping Dong,2 Yan Du,3 Zheng Liang1
1Department of Otorhinolaryngology, Tianjin Medical University General Hospital, Tianjin, People’s Republic of China; 2Department of Cancer Cell Biology, Tianjin’s Key Laboratory of Cancer Prevention and Therapy, National Clinical Research Center for Cancer, Tianjin Medical University Cancer Institute and Hospital, Tianjin, People’s Republic of China; 3Department of Operating Theatre, Tianjin Medical University General Hospital, Tianjin, People’s Republic of China
Correspondence: Zheng Liang, Department of Otorhinolaryngology, Tianjin Medical University General Hospital, Tianjin, People’s Republic of China, Tel +8618802230827, Email [email protected]
Purpose: The involvement of dedicator for cytokinesis 4 (DOCK4), a guanine nucleotide exchange factor for Rac1, in immune infiltration in stomach adenocarcinoma (STAD) remains unclear.
Methods: The UALCAN database was used to analyze the expression of the DOCK family. The Kaplan–Meier method and Gene Expression Profiling Interactive Analysis (GEPIA) databases were used to assess the prognostic value of the DOCK family in STAD. Furthermore, the correlation between expression of DOCK4 as well as other immune-related marker genes and tumor immune infiltration in STAD was explored using the TIMER and GEPIA websites. Subsequently, the relationship between DOCK4 expression and clinical characteristics was verified using the UALCAN database. Finally, DOCK4 mutation was analyzed via the TIMER2.0 and cBioPortal databases and the DOCK4 protein-protein interaction networks were constructed using the GeneMANIA and STRING websites.
Results: DOCK4 was found to be a new prognostic biomarker in STAD. DOCK4 expression in tumors was thoroughly evaluated relative to paracancerous tissues; overexpression of DOCK4 had a negative impact on the prognosis of patients with STAD. DOCK4 was found to be significantly associated with tumor immune infiltration in STAD.
Conclusion: In summary, DOCK4 is a potential regulator of the recruitment and regulation of immune-infiltrating cells, thus serving as a valuable prognostic biomarker in STAD.
Keywords: DOCK4, stomach adenocarcinoma, immune infiltration, prognostic biomarker, TME
Introduction
Gastric cancer is a common malignancy worldwide with a complex etiology. Despite its declining incidence and mortality rates, the burden of gastric cancer remains high in many countries.1,2 The histological classification of gastric cancer mainly includes adenocarcinoma, adenosquamous carcinoma, squamous cell carcinoma, neuroendocrine carcinoma, and undifferentiated carcinoma.3 Adenocarcinomas account for approximately 90% of gastric cancers, and the World Health Organization (WHO) classification system divides gastric adenocarcinoma into five histologic subtypes: tubular adenocarcinoma, papillary adenocarcinoma, mucinous adenocarcinoma, poorly cohesive carcinoma, and mixed carcinoma.4 Studies have shown that the WHO histological classification and American Joint Committee on Cancer TNM staging systems can aid in the prediction of stomach adenocarcinoma (STAD) patient survival outcomes; however, these systems have limitations.5,6 Therefore, there is a need to discover new biomarkers to improve prognostic accuracy for STAD. It has been found that considering the data of tumor-infiltrating neutrophils in patients with STAD can improve the prognostic accuracy.7
The tumor microenvironment (TME) contains a complex variety of cells and molecules, most of which are reported to be related to tumor immune escape, growth, and metastasis.8–10 Moreover, the development of immune checkpoint inhibitors has led to improved clinical outcomes for patients with multiple types of solid tumors.11–13 For STAD, immune checkpoint inhibitors that block the cytotoxic T-lymphocyte antigen 4 (CTLA4) and programmed death-1 (PD-1) axes provide patients with new treatment strategies.14 However, all patients with gastric cancer are not sensitive to immune checkpoint inhibitors, such as PD-1 inhibitors,15 and the role of the TME in STAD is not completely understood. Therefore, there is a need to identify valuable prognostic biomarkers and immunotherapeutic targets for STAD.
DOCK family proteins are known to be activators of the Rho GTPases, Rac and Cdc42. The DOCK protein family has 11 members, from DOCK1 to DOCK11.16 According to their sequences and domains, the DOCK proteins are divided into four subgroups: DOCK-A, DOCK-B, DOCK-C, and DOCK-D.17 In addition, the DOCK family proteins are the most researched proteins in cell adhesion and migration, and interactions between the immune system and DOCK proteins have been reported. For example, DOCK2 and DOCK8 are widely expressed in immune cells, and as activators of Rac and Cdc42, they are necessary for mediating cytotoxic function and cell-cell adhesion of immune cells, respectively.16,18,19
Only a few studies have been conducted on the link between DOCK proteins and gastric cancer. One study has found that DOCK1 could be a target of mycophenolic acid in modulating gastric cancer cell migration.20 Moreover, DOCK5 could be a chemosensitivity-related gene in gastric cancer.21 It has also been reported that DOCK6 plays a pivotal role in regulating cancer stem cells in gastric cancer22 and promotes gastric cancer metastasis through the Rac1/Cdc42 axis.23 DOCK4, which belongs to the DOCK-B subgroup, is an atypical guanine nucleotide exchange factor for Rac1.24 Several studies have demonstrated that DOCK4 plays a carcinogenic role in multiple tumors. For example, DOCK4 overexpression promotes the proliferation, migration, and invasion of liver cancer cells.25 Furthermore, lncRNA AC073284.4 suppresses cell invasion, metastasis, and Epithelial-mesenchymal transition (EMT) in breast cancer via the miR‐18b‐5p/DOCK4 axis.26 DOCK4 also regulates the migration of breast cancer cells by binding to SH3YL1 to activate Rac1.27 Furthermore, DOCK4 promotes lung adenocarcinoma cell extravasation and metastasis through the TGF-β/Smad pathway.28 Moreover, DOCK4 was also determined to be a prognostic biomarker in ovarian cancer and has been shown to be associated with immune infiltration.29 However, the correlation between the expression of DOCK4 and the prognosis of patients with STAD remains unknown.
In the present study, we comprehensively analyzed the expression and clinical prognosis of members of the DOCK family in STAD. We then screened out DOCK4 and explored its correlation with the STAD TME. We aimed to reveal the prognostic value of DOCK4 in stomach adenocarcinoma and provide new evidence of the role of DOCK4 in tumor immunity.
Materials and Methods
UALCAN Database Analysis
UALCAN (http://ualcan.path.uab.edu.) aims to analyze the relative expression of genes as well as their correlation with diverse clinicopathologic features from TCGA, MET500, and CPTAC databases.30 Thus, this database was used to assess the expression level of DOCK proteins in STAD and paracancerous tumor tissues, as well as the relationship between DOCK4 expression and clinical characteristics.
Kaplan–Meier Plotter Database Analysis
The Kaplan–Meier plotter (http://kmplot.com/analysis/) was used to evaluate the prognostic impact of 54,675 genes in 10,461 cancer samples from 21 cancer types.31 We first used this database to explore the influence of DOCK expression on the overall survival (OS) of patients with STAD and analyze the impact of clinicopathological factors and immune cells on the OS of patients with STAD.
TIMER Database Analysis
The TIMER database (https://cistrome.shinyapps.io/timer/) comprehensively assesses the correlation between tumor-infiltrating immune cells and important tumoral genomic changes using six functional modules.32 In our study, the correlation between the expression of DOCK3 and DOCK4 with the degree of immune infiltration of B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells (DCs) was analyzed using the TIMER gene modules. Next, the tumor infiltration levels of cells with different somatic copy number alterations were compared using the SCNA module. The relationship between DOCK4 expression and immune marker genes was explored using the correlation module. Finally, DOCK4 mutations in 34 cancer types were analyzed using the gene mutation module in TIMER2.0.
Gene Correlation Analysis in GEPIA
The online database Gene Expression Profiling Interactive Analysis (GEPIA) (http://gepia.cancer-pku.cn/detail.php) provides RNA sequencing expression data to analyze the expression differences between tumor and control samples in multiple cancer types.33 This database was used to assess OS based on the expression of DOCK4, and the link between this gene and the expression of particular immune-related marker genes was verified.
cBioPortal Database Analysis
The cBioPortal website (http://cbioportal.org) has been used for analyzing multidimensional cancer genomics data from cancer tissues and cells to provide the genetic, epigenetic, gene expression, and proteomic information.34 This database was used to analyze the alterations in DOCK4 expression in stomach adenocarcinoma samples.
STRING Database Analysis
The STRING database (https://string-db.org/) aims to collect and integrate information on protein-protein interactions (PPI) to construct a comprehensive and objective global network for users.35 The database was used to construct the PPI network of DOCK4 and Cytoscape software was used to visualize the results.
GeneMANIA Database Analysis
GeneMANIA (http://genemania.org) is an open website that can analyze correlated genes and predict gene functions and their interactions.36 The database was used to analyze the genes correlated with DOCK4 and construct the PPI network.
Statistical Analysis
The hazard ratios (HRs) and p-values of the OS curves were computed using the Log rank test in the GEPIA and Kaplan–Meier Plotter databases. Spearman’s rank correlation coefficient was used to analyze the relationship between TIMER and GEPIA. The significance threshold was set at p < 0.05.
Results
Assessment of the Expression Level of DOCK Proteins in STAD
The mRNA expression of the DOCK family proteins in patients with STAD was analyzed using the UALCAN database (Figure 1). The expression of DOCK1 (Figure 1A, p<0.01), DOCK2 (Figure 1B, p<0.01), DOCK4 (Figure 1D, p<0.001), DOCK5 (Figure 1E, p<0.001), DOCK6 (Figure 1F, p<0.001), DOCK7 (Figure 1G, p<0.001), and DOCK10 (Figure 1J, p<0.001) was higher in tumor tissues than in paracancerous tissues, while DOCK3 (Figure 1C, p<0.01) and DOCK8 (Figure 1H, p<0.01) were lower in tumor tissues compared to paracancerous tissues. No statistically significant difference was observed in DOCK9 (Figure 1I) and DOCK11 (Figure 1K).
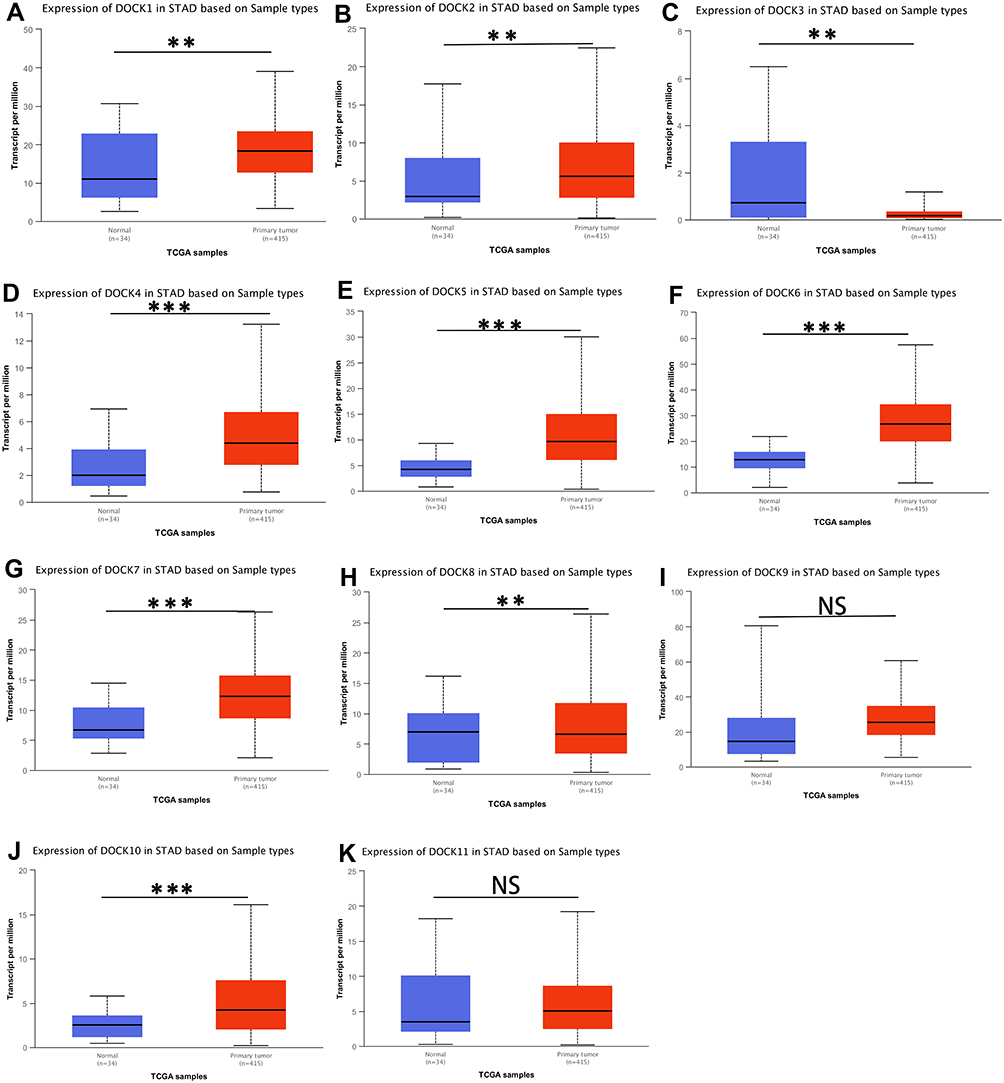 |
Figure 1 The expression of DOCK proteins in stomach adenocarcinoma (STAD) and paracancerous tissues via UALCAN database. (A) DOCK1, (B) DOCK2, (C) DOCK3, (D) DOCK4, (E) DOCK5, (F) DOCK6, (G) DOCK7, (H) DOCK8, (I) DOCK9, (J) DOCK10, (K) DOCK11. (**P <0.01, ***P < 0.001). Abbreviation: NS, not statistically significant. |
The Prognostic Value of DOCK Proteins in STAD
Next, DOCK9 and DOCK11, which showed no statistical difference in expression were excluded. The impact of the expression of the remaining DOCK proteins on OS in patients with STAD was determined using the Kaplan–Meier plotter database. As shown in Figure 2, overexpression of DOCK3 (Figure 2C, HR=1.95, log-rank P= 4.5e−05) and DOCK4 (Figure 2D, HR=1.57, log-rank P=0.016) was associated with poor prognosis in patients with STAD. DOCK1 (Figure 2A, HR=1.28, log-rank P=0.15), DOCK2 (Figure 2B, HR=1.28, log-rank P=0.17), DOCK5 (Figure 2E, HR=0.84, log-rank P=0.31), DOCK6 (Figure 2F, HR=0.81, log-rank P=0.26), DOCK7 (Figure 2G, HR=0.79, log-rank P=0.2), DOCK8 (Figure 2H, HR=0.86, log-rank P=0.38), and DOCK10 (Figure 2I, HR=1.29, log-rank P=0.13) showed no statistically significant difference in OS. Furthermore, to verify the results of the GEPIA website, we reached a consistent conclusion. The details are provided in Supplementary Figure 1. The results suggest that DOCK3 and DOCK4 are correlated with unfavorable prognosis in patients with STAD.
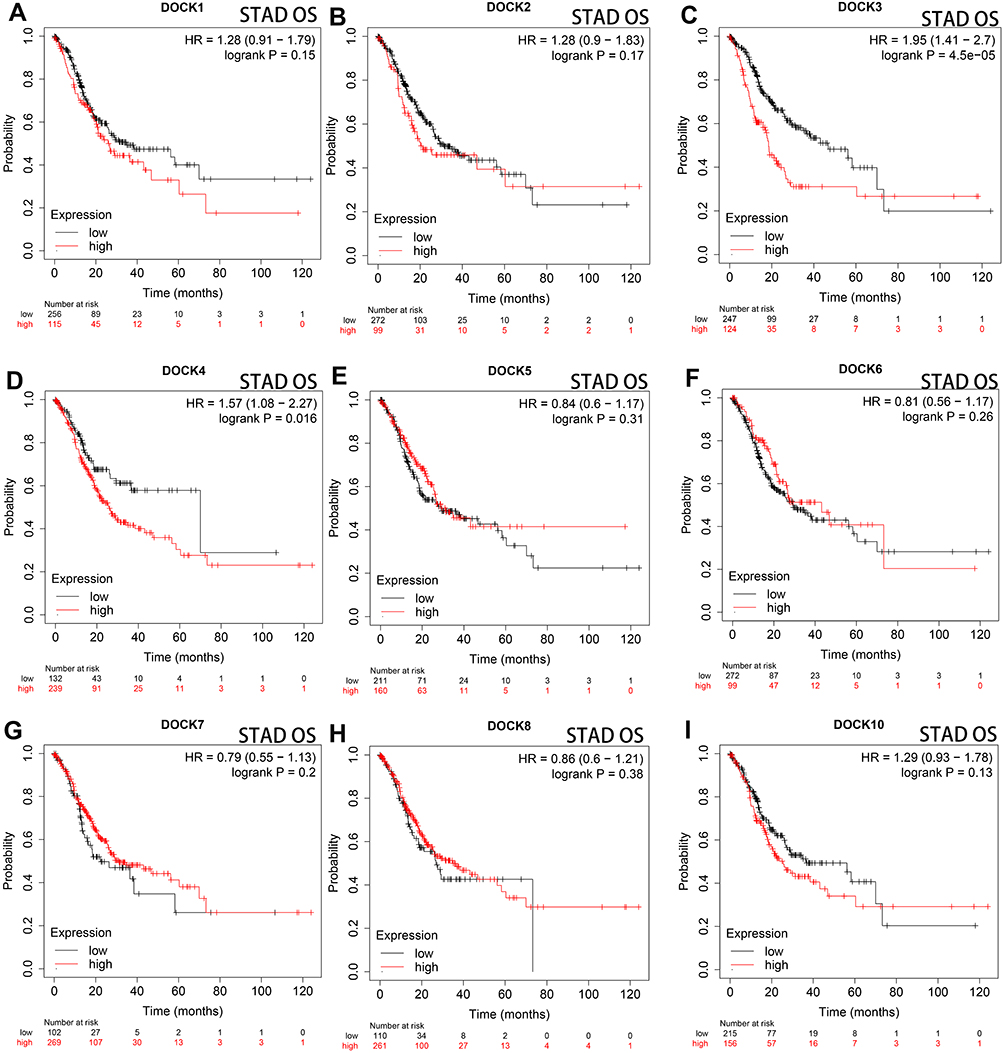 |
Figure 2 Comparison of Kaplan–Meier survival curves of DOCK proteins in patients with STAD. High DOCK3 (C) and DOCK4 expression (D) resulted in unfavorable OS in patients with STAD (n=371). The expression of DOCK1 (A), DOCK2 (B), DOCK5 (E), DOCK6 (F), DOCK7 (G), DOCK8 (H), DOCK10 (I) showed no significant correlation with OS. Abbreviations: HR, hazard ratio; OS, overall survival. |
Correlation Analysis of DOCK3 and DOCK4 Expression and Immune Infiltration in STAD
Next, we used the TIMER database to assess the correlation between DOCK3 and DOCK4 expression and the level of immune infiltrates, including B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and DCs, as well as tumor purity (Figure 3). DOCK3 expression was significantly related to B cells (r=0.253, p=8.61e−07), CD8+ T cells (r=−0.111, p=3.35e−02), CD4+ T cells (r=0.356, p=5.66e−13), and macrophages (r=0.308, p=1.45e−09) (Figure 3A). DOCK4 had a significant positive correlation with CD8+ T cells (r=0.25, p=1.14e−06), CD4+ T cells (r=0.39, p=9.40e−15), macrophages (r=0.518, p=8.17e−27), neutrophils (r=0.375, p=7.96e−14), and DCs (r=0.458, p=1.11e−20), while it was negatively correlated with tumor purity (r=−0.143, p=5.33e−03) (Figure 3B). Furthermore, we analyzed the association between copy number variations of DOCK3 and DOCK4 with the above six immune infiltrates (Figure 4). The results revealed that the levels of B cells (p<0.01), CD8+ T cells (p<0.001), CD4+ T cells (p<0.001), macrophages (p<0.001), neutrophils (p<0.001), and DCs (p<0.001) were relatively low, as indicated by the deletion of DOCK3 and DOCK4. These results indicate that DOCK4 plays a vital role in immune infiltration.
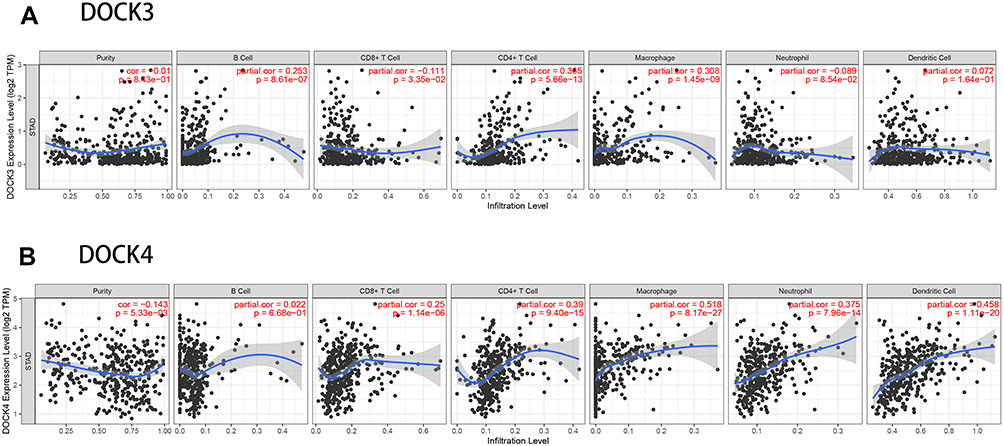 |
Figure 3 Correlation of DOCK3 and DOCK4 expression with immune infiltration levels in patients with STAD. The correlations between DOCK3 (A) and DOCK4 (B) expression with tumor purity and infiltrating levels of B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and DCs in STAD. |
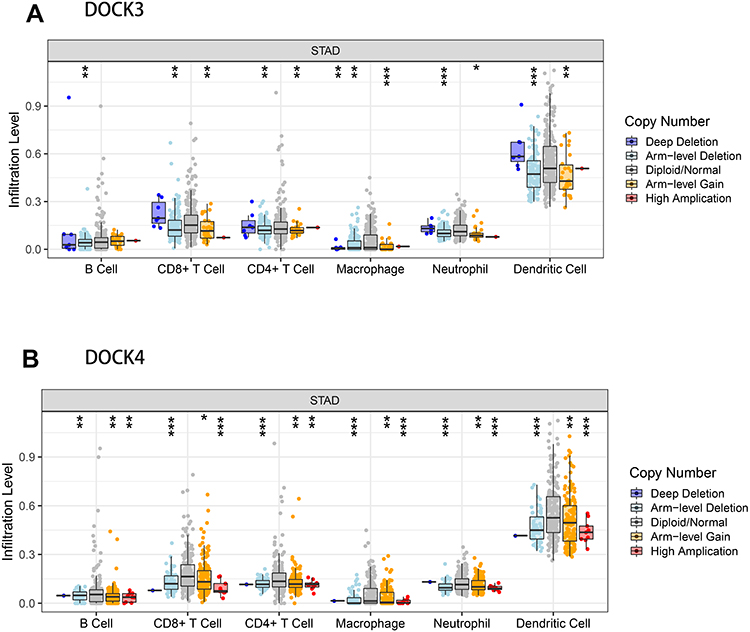 |
Figure 4 Correlation of somatic copy number variations of DOCK3 and DOCK4 with immune infiltration levels in patients with STAD. The correlation between the somatic copy number alterations of DOCK3 (A) and DOCK4 (B) with the immune infiltrating levels of B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and DCs in patients with STAD. (*P < 0.05, **P <0.01, ***P < 0.001). |
Correlation of DOCK4 and Immune Marker Expression
We analyzed the correlation between DOCK4 expression and immune marker genes in numerous immune cells, including CD8+ T cells, T cells (in general), B cells, monocytes, tumor-associated macrophages (TAMs), M1 and M2 macrophages, neutrophils, natural killer cells, and DCs in patients with STAD using the TIMER and GEPIA databases. We also analyzed T cells with different subsets, such as T helper 1 cells (Th1), T helper 2 cells (Th2), T helper 17 cells (Th17), follicular helper T cells (Tfh), regulatory T cells (Treg), and exhausted T cells. After adjusting for the correlation with purity, DOCK4 expression was significantly associated with most immune marker genes in patients with STAD (Table 1). We noticed that most markers of monocytes, TAMs, and M2 macrophages were strongly related to DOCK4 expression, including the monocyte markers, CD86 and CD115 (r >0.5, p<0.001), the TAM marker, IL10 (r>0.5, p<0.001), and the M2 macrophage markers, CD163, VSIG4, and MS4A4A (r>0.5, p<0.001). This suggests that DOCK4 regulates macrophage polarization in patients with STAD. We also found that DC immune marker genes, such as HLA-DPB1, HLA-DRA, HLA-DPA1, BDCA-1, BDCA-4, and CD11c, were significantly related to DOCK4 expression (p<0.001). Moreover, strengthening Treg function and reducing CD8+ T cell cytotoxicity is the main mechanism by which DCs promote the metastatic phenotype of tumors,37 which reveals the potential function of DOCK4. In Tregs, there was a significant relationship between FOXP3, TGFβ, CCR8, and STAT5B with DOCK4 (p<0.001). Finally, DOCK4 was positively correlated with T cell depletion genes, such as PD-1, CTLA4, LAG3, and TIM-3 (p<0.001).
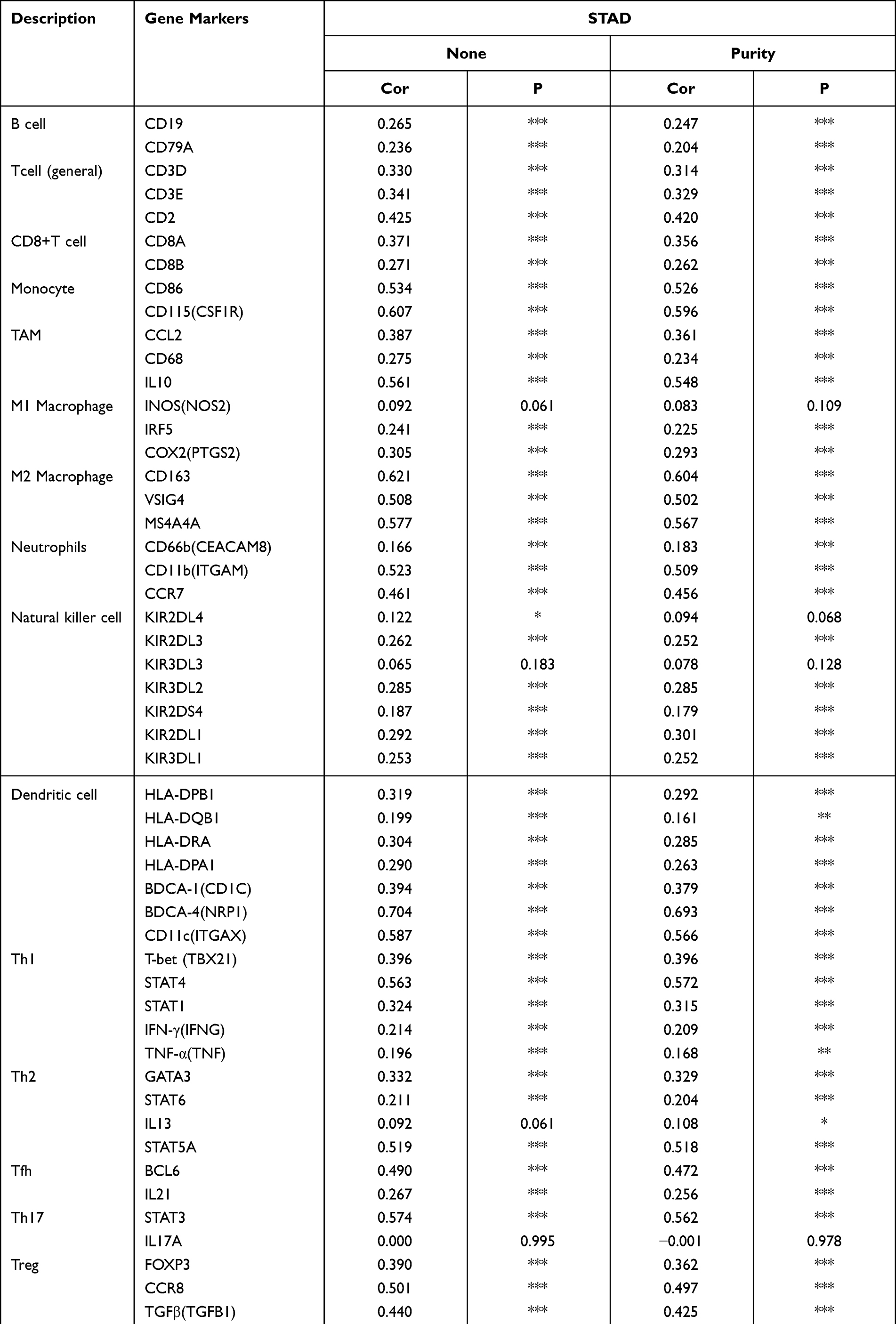 |
Table 1 Correlation Analysis Between DOCK4 and Relate Genes and Markers of Immune Cells in TIMER |
Analysis of the GEPIA database confirmed these findings. These immune marker genes, including CD86 and CD11 (r>0.5, p<0.001, Figure 5A), IL10 (r>0.5, p<0.001, Figure 5B), MS4A4A (r>0.5, p<0.001, Figure 5C), BDCA-4 and CD11c (r>0.5, p<0.001, Figure 5D), STAT5B (r>0.5, p<0.001, Figure 5E), TIM-3 (r>0.5, p<0.001, Figure 5F), CD11b (r>0.5, p<0.001, Figure 5G), STAT5A (r>0.5, p<0.001, Figure 5H) and STAT3 (r>0.5, p<0.001, Figure 5I) were strongly correlated with DOCK4 expression. These results indicate that DOCK4 plays a key role in immune escape of the TME in patients with STAD.
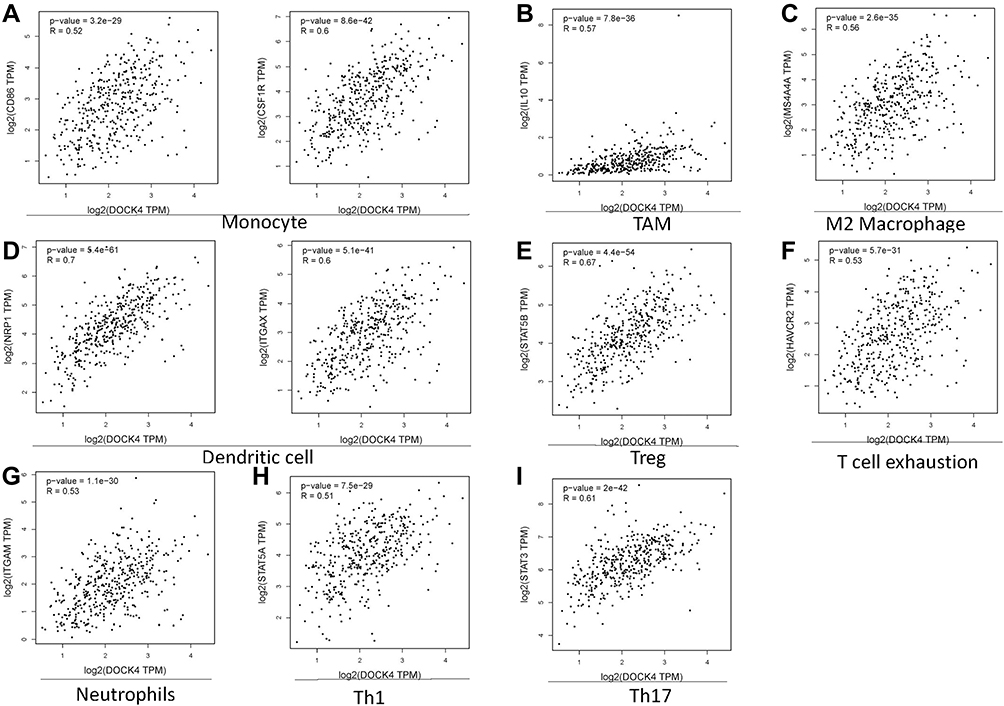 |
Figure 5 Correlation of DOCK4 expression with partial immune marker genes in patients with STAD. Markers include (A) CD86 and CD115 of Monocyte; (B) IL10 of TAM; (C) MS4A4A of M2 macrophages; (D) BDCA-4 (NRP1) and CD11c (ITGAX) of DCs; (E) STAT5B of Tregs; (F) TIM-3 (HAVCR2) of T cell exhaustion; (G) CD11b (ITGAM) of neutrophils; (H) STAT5A of Th1; and (I) STAT3 of Th17. |
Prognostic Value of DOCK4 Expression in Patients with STAD Based on Immune Cells
We further explored the ability to predict prognosis of patients with STAD based on immune cells, including basophils, B cells, CD4+ memory T-cells, CD8+ T cells, eosinophils, macrophages, mesenchymal stem cells, natural killer T-cells, Tregs, Th1 and Th2 cells using the Kaplan–Meier plotter (Table 2). The results revealed that high expression of DOCK4 in a decreased number of basophils (HR=1.68, P=0.021), CD8+ T cells (HR=1.96, P=0.022), macrophages (HR=1.95, P=0.013), mesenchymal stem cells (HR=1.9, P=0.013), natural killer T cells (HR=2.29, P=0.0018), and Th1 cells (HR=1.89, P=0.0038) displayed poor prognosis. Furthermore, the overexpression of DOCK4 in enriched immune cell subtypes, including B cells (HR=1.89, P=0.0042), CD4+ memory T cells (HR=1.84, P=0.0069), CD8+ T cells (HR=1.62, P=0.033), eosinophils (HR=1.78, P=0.021), macrophages (HR=1.73, P=0.021), Tregs (HR=1.81, P=0.015), and Th2 cells (HR=1.51, P=0.033) had an unfavorable impact on OS. The above results suggested that high DOCK4 expression in STAD may affect prognoses in part due to immune infiltration.
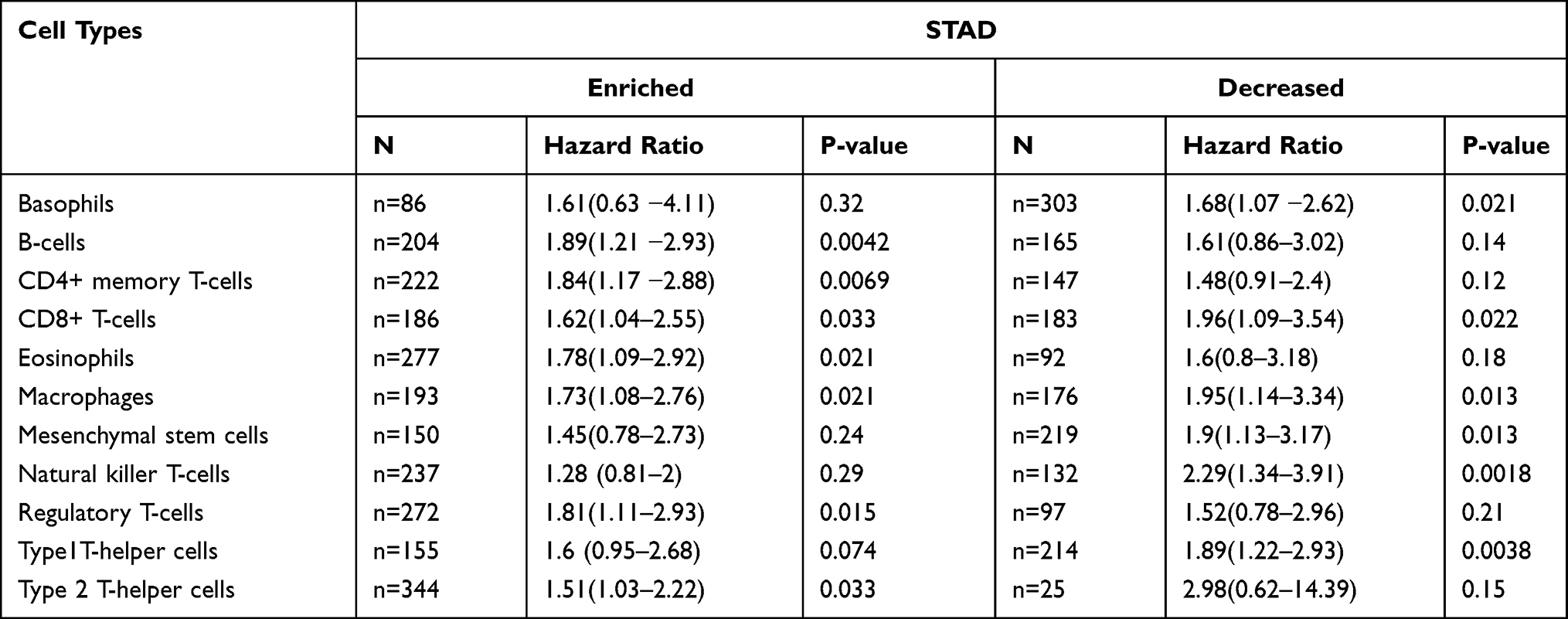 |
Table 2 Kaplan-Meier Plotter to Analyze the Overall Survival on the Expression of DOCK4 Based on Immune Cell Subgroups in STAD |
Correlation of DOCK4 Expression and Clinical Characteristics of Patients with STAD
We further analyzed the relationship between DOCK4 expression and clinical characteristics of patients with STAD in the UALCAN database. DOCK4 expression was higher in tumor tissues than in paracancerous tissues, as well as in stage 3 cancer than stage 2 (Figure 6A, p<0.01) and stage 1 cancer (Figure 6A, p<0.05). No statistically significant difference was observed according to race (Figure 6B) and sex (Figure 6C) of patients with STAD. In addition, DOCK4 expression was lower in patients 81–100 years of age than in those 61–80 years of age (Figure 6D, p<0.05). Based on the tumor grade, the expression of DOCK4 in grade 3 tumors was excessive compared to grade 2 and grade 1 tumors (Figure 6E, p<0.001). There was no statistically significant difference in the nodal metastasis status of patients with STAD (Figure 6F). Regarding TP53 mutation status, DOCK4 expression in patients with non-mutated TP53 was higher than that of patients with mutated TP53 (Figure 6G, p<0.05). No statistically significant difference was observed in the H. pylori infection status of patients with STAD (Figure 6H).
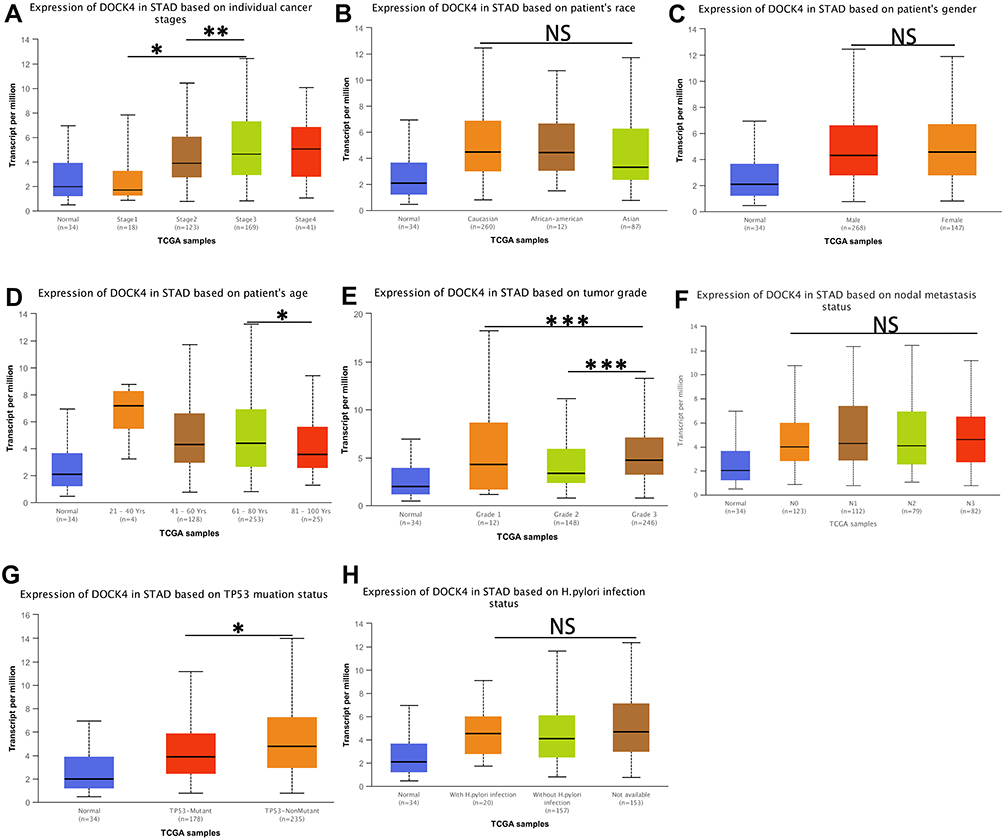 |
Figure 6 Correlation of DOCK4 expression with clinical characteristics of patients with STAD. (A) DOCK4 expression in according to STAD cancer stages; (B) DOCK4 expression according to race of patients with STAD; (C) DOCK4 expression according to sex of patients with STAD; (D) DOCK4 expression according to age of patients with STAD; (E) DOCK4 expression according to STAD tumor grade; (F) DOCK4 expression according to nodal metastasis status of STAD; (G) DOCK4 expression according to TP53 mutation status of patients with STAD; (H) DOCK4 expression according to H. pylori infection status of patients with STAD. (*P<0.05 **P<0.01 ***P<0.001). Abbreviation: NS, not statistically significant. |
In the stratified prognostic analysis of DOCK4 expression in patients with STAD, we found that upregulated DOCK4 had a negative impact on OS in clinical stage 3 (p=0.025) and clinical stage 4 patients (p=0.036) using the Kaplan–Meier database (Table 3). Furthermore, DOCK4 also played an unfavorable role in the following clinicopathological characteristics: female sex (p=0.00035), white race (p=0.01), and mutation burden (p=0.046) (Table 3). The other clinicopathological characteristics did not demonstrate statistically significant differences (p>0.05) (Table 3). These results suggest that DOCK4 expression may influence the prognosis of patients with clinically advanced STAD.
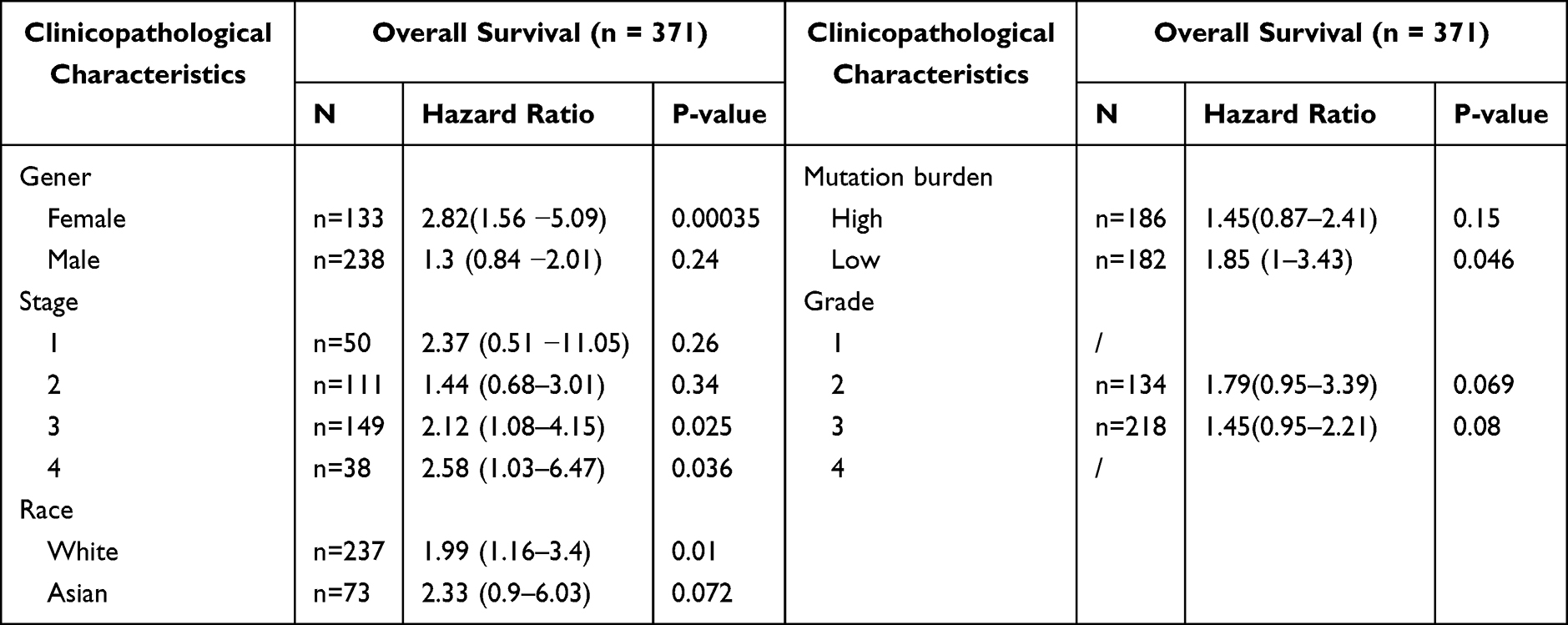 |
Table 3 Kaplan-Meier Plotter to Determine the Effect of Different Clinicopathological Factors on the Expression of DOCK4 Gene and Clinical Prognosis in STAD |
Mutation Analysis of DOCK4
The mutation profile of DOCK4 was analyzed using the TIMER2.0 and cBioPortal databases. The DOCK4 mutation rate was low in most of 34 TCGA cancer types, ranging from 0.02% (1/500 patients with Thyroid carcinoma (THCA)) to 11% (60/531 patients with Uterine Corpus Endometrial Carcinoma (UCEC)) (Figure 7A) in the TIMER2.0 database. The mutation rate of DOCK4 in patients with STAD was 7% (31/439), which was similar to the results from the cBioPortal database, which ranged from 5.44% (8/147) to 6.36% (28/440) (Figure 7B and Supplementary Table 1). In addition, the amplification rate of DOCK4 ranged from 1.36% (4/295) to 2.72% (4/147), and the rate of deep deletions ranged from 0.23% (1/440) to 0.68% (1/447) (Figure 7B) (Supplementary Table 1).
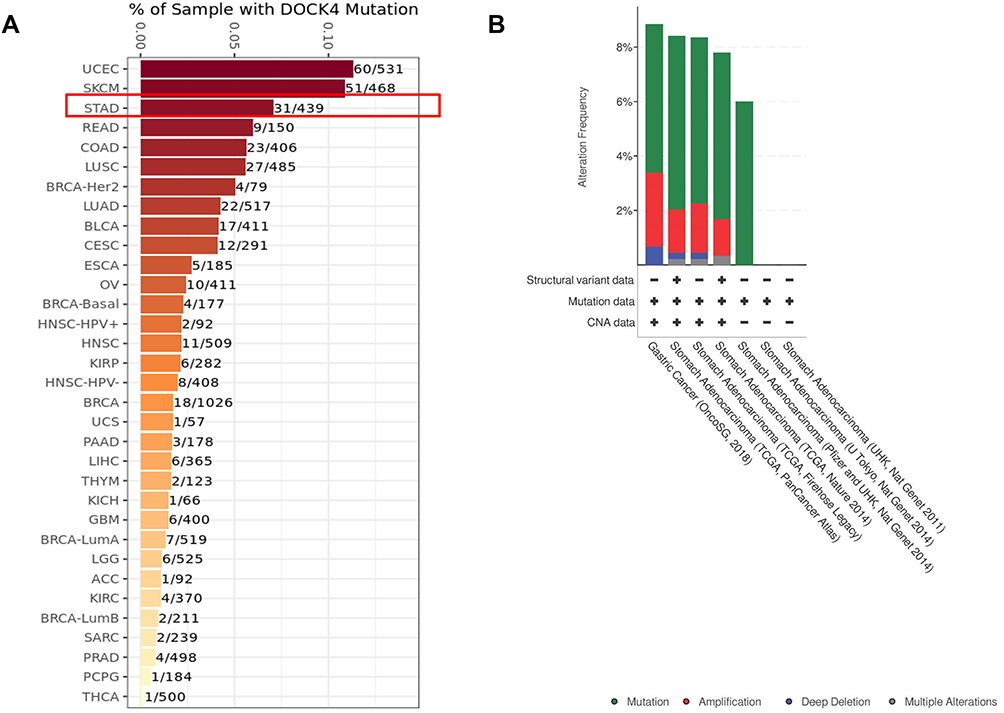 |
Figure 7 Mutational analysis of DOCK4. (A) Mutation of DOCK4 in 34 TCGA cancer types. (B) Alterations of DOCK4 in stomach adenocarcinoma samples. |
PPI Network of DOCK4 Analysis
The PPI networks of DOCK4 were constructed using the GeneMANIA website. The results revealed a list of genes correlated with DOCK4. The correlated genes were enriched in the following pathways: Rho GTPase binding, regulation of smooth muscle cell migration, smooth muscle cell migration, positive regulation of smooth muscle cell migration, regulation of vascular associated smooth muscle cell migration, vascular associated smooth muscle cell migration and muscle cell migration (Figure 8A). In addition, the PPI network data of DOCK4 from the STRING website was visualized using Cytoscape software (Figure 8B). DOCK4 interacted with ribosomal proteins, such as RPL6, RPL24, and RPL18, and ribosomal subunit proteins, such as RPS27A, RPS25, RPS29, and RPS15.
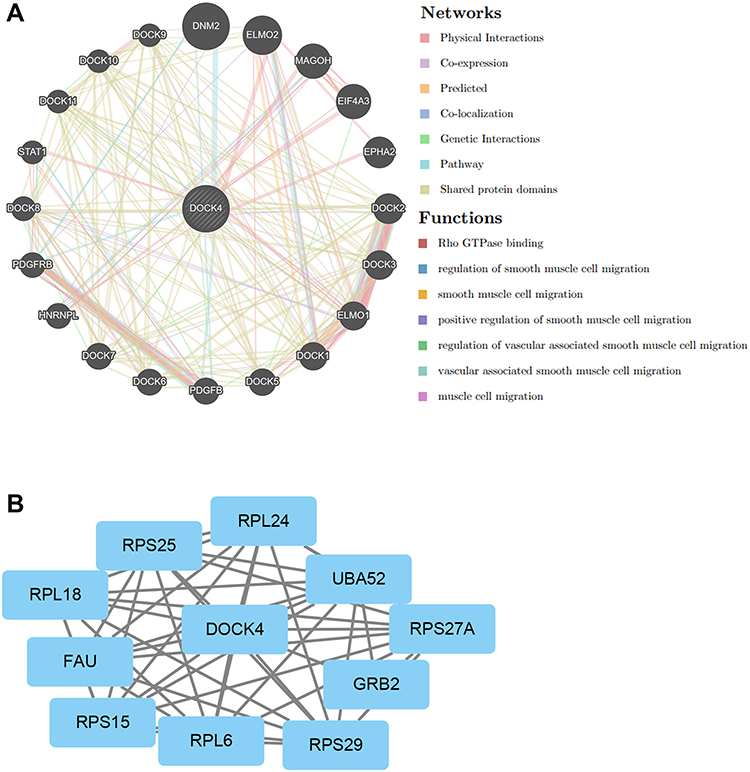 |
Figure 8 Gene network analysis of DOCK4. (A) DOCK4 with its correlated genes involved in physical interactions, co-expression, prediction, co-localization, pathways, genetic interaction, and shared protein domains. (B) Protein-protein interaction network of DOCK4-correlated genes based on Cytoscape. |
Discussion
Stomach adenocarcinoma is the fifth most common malignancy and ranks third in cancer-related mortality worldwide.1 With recent improvements in treatment methods for STAD, including surgical treatment, neoadjuvant radiotherapy, chemotherapy, and targeted therapy, the 5-year survival rate of patients with early diagnosis and treatment has increased; however, the survival outcomes of advanced/metastatic cancers remain unsatisfactory.38 A possible mechanism of development of stomach adenocarcinoma involves the malignancy of cancer cells and various immune cells in the TME.39 Thus, the development of immune checkpoint inhibitors is beneficial for patients with advanced or metastatic STAD. For example, pembrolizumab prolongs OS and reduces adverse effects of treatment in patients with advanced STAD.40 Moreover, the combination of nivolumab and ipilimumab is effective for chemotherapy-refractory gastroesophageal adenocarcinoma.14 However, the incomplete validity of immune checkpoint inhibitors in gastric cancer patients has pose a new challenge.15,41 Thus, to optimize individualized treatment strategies, we aimed to explore new immunotherapeutic-related molecules for STAD. There are 20 Rho GTPases in the human genome, including Rac1 and Cdc42, which have a deep impact on cell proliferation, differentiation, motility, adhesion, survival and secretion.42 As Rho GTPase activators, DOCK proteins were first identified as exchange factors for Rac1/2/3 and Cdc42 more than 10 years ago.16 Thus, the DOCK family plays an important role in carcinogenesis and central nervous and immune system diseases through the activation of Rho GTPases.43,44 Specifically, DOCK4 is an atypical Rho GTPase guanine nucleotide exchange factor and induces physiological processes and diseases that activate Rac1. Moreover, DOCK4 plays an important role in neurite differentiation, erythropoiesis, and vascular smooth muscle cell migration.45–48 Furthermore, in tumors, DOCK4 mainly promotes the migration, invasion, and EMT of cancer cells.25–27 This is the first study to report on the interactions between DOCK4 and the STAD TME. We found that high DOCK4 expression is a poor prognostic factor for STAD. Moreover, DOCK4 expression was significantly related to the expression of different immune-related markers, highlighting its potential role in STAD immunity. Thus, DOCK4 is a valuable biomarker for this cancer.
Using the UALCAN database, we found that the expression of most DOCK proteins was higher than that of the control samples (Figure 1). Based on their prognostic value in the Kaplan–Meier plotter and GEPIA databases, we selected DOCK3 and DOCK4 for further study; both showed an unfavorable impact on the OS of patients with STAD (Figure 2). DOCK3 is specifically expressed in neurons and participates in the development of Alzheimer’s disease.49 Furthermore, the Twist1 protein promotes EMT through the NEDD9/DOCK3/Rac axis in head and neck squamous cell carcinoma.50 DOCK3 also drives tumor cell adhesion, migration, and invasion by regulating Rac1 activity in lung cancer.51 In addition to metastasis, the relationship between immune infiltration and DOCK3 and DOCK4 in patients with STAD was explored using the TIMER and GEPIA websites. Only a weak relationship was observed between DOCK3 and B cells, CD8+ T cells, CD4+ T cells, and macrophages (Figure 3A). However, DOCK4 had a strong positive correlation with CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and DCs, and a negative correlation with tumor purity (Figure 3B). In addition to copy number variations, deletion of DOCK4 was significantly associated with lower levels of the six types of immune cells (Figure 4). Thus, DOCK4 is closely related to the TME of STAD. Although the structure of DOCK4 has high homology with DOCK3, it can also be detected as a secreted protein in peripheral blood.29
The function of chemokines in lymphocyte migration depends on the activation of DOCK2, and DOCK8 mutations cause functional defects in the immune system, suggesting that DOCK2 and DOCK8 are necessary regulators of the immune system.52,53 Thus, it can be predicted that DOCK4 has a correlation with immune infiltration similar to DOCK2 and DOCK8. Thus, we further verified the link between the expression of immune-related marker genes and DOCK4. There was a significant correlation between DOCK4 and diverse marker genes of immune–infiltrating cells, including monocytes, TAMs, M2 macrophages, DCs, Tregs, and exhausted T cells in the TIMER database. DOCK4 was weakly associated with M1 (antitumor) macrophage markers (IRF5 and COX2) and strongly associated with M2 (protumor)54 macrophage markers (CD163, VSIG4, and MS4A4A) (Table 1). These results revealed that DOCK4 participates in macrophage polarization.
A high level of immune infiltration of Tregs is related to poor prognosis of cancer patients. FOXP3 participates in the regulation of Treg differentiation, suppressing the antitumor immune response.55 PD-1, CTLA4, Tim-3, and LAG3 are vital markers of exhausted T cells, representing a poor response to antigen-mediated T cell receptor (TCR) stimulation.56 There was a significant correlation between DOCK4 expression and Treg markers (FOXP3, TGFβ, CCR8, and STAT5B) as well as T cell exhaustion markers (PD-1, CTLA4, TIM-3, and LAG3) (Table 1). These results provide evidence that elevated DOCK4 levels suppress the antitumor immune response in clinical patients with STAD. NRP1, a DC marker, was strongly associated with DOCK4 and was determined to be a prognostic factor for STAD.57
T helper cell infiltration is conducive to prognostic prediction58 and the significant link between DOCK4 expression and several markers of T helper cells (Th1, Th2, Tfh, Th17) increased the predictive ability of prognosis in patients with STAD (Table 1). Based on the homologous data from TCGA, we analyzed the above markers in the GEPIA database and obtained similar results (Figure 5). Furthermore, we found that DOCK4 expression had an impact on OS at different stages of immune infiltration (Table 2). The decreased number of basophils, CD8+ T cells, macrophages, mesenchymal stem cells, natural killer T cells, and Th1 cells is consistent with our conclusion. However, overexpression of DOCK4 was also correlated with poor prognosis in enriched status of B cells, CD4+ memory T cells, CD8+ T cells, eosinophils, macrophages, Tregs, and Th2 cells, demonstrating the complexity of the immune system.
Immune infiltration, possibly as an upstream regulator of DOCK4, participates in the development of STAD. Furthermore, according to the analysis of TCGA genomic subtypes, Epstein-Barr virus (EBV)-positive patients with STAD show rich CD8+ T cell infiltration14 DOCK4 is related to the OS of patients with STAD with enriched CD8+ T cells; thus, decreasing DOCK4 expression enables individualized treatment of STAD. The prognosis of patients with STAD in the same classification and staging remains significantly different,4 and the molecular subtypes determined by TCGA can effectively classify patients to help provide personalized therapy. Moreover, evaluating DOCK4 expression may enhance the predictive ability for the prognosis of patients with STAD.
Immunotherapy has been shown to prolong the life of patients with multiple types of cancer.59,60 Similarly, decreasing DOCK4 expression improves prognosis of patients with STAD, and the combination with other immune checkpoint inhibitors may prove to be a new treatment strategy for some patients, such as those who experience adverse events from PD-1 inhibitors.
Finally, we analyzed the correlation between DOCK4 levels and clinical characteristics of patients with STAD (Figure 6). By combining the current and previous research, we made significant observations. The TP53-mutated subtype shows a lower degree of immune infiltration than the TP53-wildtype in gastric cancer patients,61 and DOCK4 expression is low in the TP53-mutated subtype. High DOCK4 expression is associated with poor prognosis in the low mutation burden group of patients with STAD. Moreover, a high mutation burden may indicate a good response to cancer treatment.62,63 Additionally, renal clear cell carcinoma is an immunotherapeutically responsive tumor with a moderate mutation burden.64 In our study, the prognostic significance of low mutation burden may be related to the regulation of TP53 mutations.
The most common genetic mutations in cancer are DNA copy number alterations. Evaluation of genetic alterations in DOCK4 showed that the copy gain frequency of DOCK4 accounted for 2.7% of STAD samples, which may explain the significant upregulation of DOCK4 in STAD tissues from the UALCAN database.
DOCK4 interacted with ribosomal-related proteins (Figure 8B) which have been reported to participate in the development of cancers. The mutation of ribosomal-related proteins is related to the deregulation of the p53 tumor suppressor network,65 providing further evidence for the correlation between DOCK4 and p53 in STAD. We also observed that DOCK4 expression was higher in patients with advanced cancer stages and grades. Additionally, both, the TGF-β/Smad pathway, vital for metastasis, and the transcription factor c-MAF, a key factor of bone metastasis, are related to DOCK4 expression.28,66 Moreover, the genes correlated with DOCK4 expression were enriched in regulating cell migration pathways (Figure 8A). This process might be the mechanism by which DOCK4 participates in metastasis of STAD; however, experimental verification is warranted. Reducing the mortality of patients with advanced/metastatic STAD proves to be a challenge.67 However, DOCK4 expression had an impact on OS in patients with STAD with advanced clinical stages (Table 3). Moreover, systemic chemotherapy is an effective method for treating advanced gastric cancer,68 and DOCK4 is associated with immune infiltration as a platinum-chemosensitive gene in ovarian cancer patients.29 Finally, DOCK4 has an unfavorable effect on survival outcomes as it regulates the immune escape of the TME of STAD. And it potentially targets platinum chemosensitivity and provides new treatment strategies for advanced/metastatic patients.
Conclusions
Our study revealed the crucial role of DOCK4 in tumorigenesis and STAD development. Overexpression of DOCK4 has a negative impact on prognosis, which could be due to the promotion of immune escape of STAD. A newly discovered prognostic factor for STAD, DOCK4 plays a vital role in immune infiltration and is a valuable molecule for immunotherapy.
Abbreviations
DOCK4, dedicator for cytokinesis 4; STAD, stomach adenocarcinoma; GEPIA, Gene Expression Profiling Interactive Analysis; PPI, protein-protein interaction; TME, tumor microenvironment; CTLA4, cytotoxic T-lymphocyte antigen; PD-1, programmed death-1; EMT, Epithelial-mesenchymal transition; OS, overall survival; HR, hazard ratio; TAM, tumor-associated macrophage; DC, dendritic cell; Th1, T helper 1 cells; Th2, T helper 2 cells; Th17, T helper 17 cells; Tfh, follicular helper T cells; Treg, regulatory T cells.
Ethics Approval and Informed Consent
This study has been reviewed by the Ethics Committee of Tianjin Medical University General Hospital. All data utilized in our study were obtained through public databases and our study did not involve any experiment on humans or animals; therefore, the Ethics Committees required no ethical approval.
Acknowledgments
We would like to thank the Natural Science Foundation of Tianjin (17JCYBJC25600) for supporting our work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Van Cutsem E, Sagaert X, Topal B, et al. Gastric cancer. Lancet. 2016;388(10060):2654–2664. doi:10.1016/S0140-6736(16)30354-3
3. Hu B, El Hajj N, Sittler S, et al. Gastric cancer: classification, histology and application of molecular pathology. J Gastrointest Oncol. 2012;3(3):251–261. doi:10.3978/j.issn.2078-6891.2012.021
4. Johnston FM, Beckman M. Updates on management of gastric cancer. Curr Oncol Rep. 2019;21(8):67. doi:10.1007/s11912-019-0820-4
5. Ning F-L, Zhang NN, Wang J, et al. Prognostic value of modified Lauren classification in gastric cancer. World J Gastrointest Oncol. 2021;13(9):1184–1195. doi:10.4251/wjgo.v13.i9.1184
6. Zhu MH, Zhang K-C, Yang Z-L, et al. Comparing prognostic values of the 7th and 8th editions of the American Joint Committee on Cancer TNM staging system for gastric cancer. Int J Biol Markers. 2019;35(1):26–32. doi:10.1177/1724600819891585
7. Zhang H, Liu H, Shen Z, et al. Tumor-infiltrating neutrophils is prognostic and predictive for postoperative adjuvant chemotherapy benefit in patients with gastric cancer. Ann Surg. 2018;267(2):311–318. doi:10.1097/SLA.0000000000002058
8. Jiang X, Wang J, Deng X, et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol Cancer. 2019;18(1):10. doi:10.1186/s12943-018-0928-4
9. Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19(11):1423–1437. doi:10.1038/nm.3394
10. Hinshaw DC, Shevde LA. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019;79(18):4557–4566. doi:10.1158/0008-5472.CAN-18-3962
11. Barbee MS, Ogunniyi A, Horvat TZ, et al. Current status and future directions of the immune checkpoint inhibitors ipilimumab, pembrolizumab, and nivolumab in oncology. Ann Pharmacother. 2015;49(8):907–937. doi:10.1177/1060028015586218
12. Garon EB, Rizvi NA, Hui R, et al. Pembrolizumab for the treatment of non-small-cell lung cancer. N Engl J Med. 2015;372(21):2018–2028. doi:10.1056/NEJMoa1501824
13. Rini BI, Plimack ER, Stus V, et al. Pembrolizumab plus axitinib versus sunitinib for advanced renal-cell carcinoma. N Engl J Med. 2019;380(12):1116–1127. doi:10.1056/NEJMoa1816714
14. Joshi SS, Badgwell BD. Current treatment and recent progress in gastric cancer. CA Cancer J Clin. 2021;71(3):264–279. doi:10.3322/caac.21657
15. Raufi AG, Klempner SJ. Immunotherapy for advanced gastric and esophageal cancer: preclinical rationale and ongoing clinical investigations. J Gastrointest Oncol. 2015;6(5):561–569. doi:10.3978/j.issn.2078-6891.2015.037
16. Gadea G, Blangy A. Dock-family exchange factors in cell migration and disease. Eur J Cell Biol. 2014;93(10–12):466–477. doi:10.1016/j.ejcb.2014.06.003
17. Cote JF, Vuori K. GEF what? Dock180 and related proteins help Rac to polarize cells in new ways. Trends Cell Biol. 2007;17(8):383–393. doi:10.1016/j.tcb.2007.05.001
18. Harada Y, Tanaka Y, Terasawa M, et al. DOCK8 is a Cdc42 activator critical for interstitial dendritic cell migration during immune responses. Blood. 2012;119(19):4451–4461. doi:10.1182/blood-2012-01-407098
19. Ham H, Guerrier S, Kim J, et al. Dedicator of cytokinesis 8 interacts with talin and Wiskott-Aldrich syndrome protein to regulate NK cell cytotoxicity. J Immunol. 2013;190(7):3661–3669. doi:10.4049/jimmunol.1202792
20. Dun B, Sharma A, Teng Y, et al. Mycophenolic acid inhibits migration and invasion of gastric cancer cells via multiple molecular pathways. PLoS One. 2013;8(11):e81702. doi:10.1371/journal.pone.0081702
21. Chang H, Rha SY, Jeung H-C, et al. Identification of genes related to a synergistic effect of taxane and suberoylanilide hydroxamic acid combination treatment in gastric cancer cells. J Cancer Res Clin Oncol. 2010;136(12):1901–1913. doi:10.1007/s00432-010-0849-0
22. Chi HC, Tsai C-Y, Wang C-S, et al. DOCK6 promotes chemo- and radioresistance of gastric cancer by modulating WNT/beta-catenin signaling and cancer stem cell traits. Oncogene. 2020;39(37):5933–5949. doi:10.1038/s41388-020-01390-0
23. Li X, Jiang M, Chen D, et al. miR-148b-3p inhibits gastric cancer metastasis by inhibiting the Dock6/Rac1/Cdc42 axis. J Exp Clin Cancer Res. 2018;37(1):71. doi:10.1186/s13046-018-0729-z
24. Guo D, Peng Y, Wang L, et al. Autism-like social deficit generated by Dock4 deficiency is rescued by restoration of Rac1 activity and NMDA receptor function. Mol Psychiatry. 2021;26(5):1505–1519. doi:10.1038/s41380-019-0472-7
25. Li H, Wang M, Zhou H, et al. Long noncoding RNA EBLN3P promotes the progression of liver cancer via alteration of microRNA-144-3p/DOCK4 signal. Cancer Manag Res. 2020;12:9339–9349. doi:10.2147/CMAR.S261976
26. Wang YY, Yan L, Yang S, et al. Long noncoding RNA AC073284.4 suppresses epithelial-mesenchymal transition by sponging miR-18b-5p in paclitaxel-resistant breast cancer cells. J Cell Physiol. 2019;234(12):23202–23215. doi:10.1002/jcp.28887
27. Kobayashi M, Harada K, Negishi M, et al. Dock4 forms a complex with SH3YL1 and regulates cancer cell migration. Cell Signal. 2014;26(5):1082–1088. doi:10.1016/j.cellsig.2014.01.027
28. Yu JR, Tai Y, Jin Y, et al. TGF-beta/Smad signaling through DOCK4 facilitates lung adenocarcinoma metastasis. Genes Dev. 2015;29(3):250–261. doi:10.1101/gad.248963.114
29. Zhao Q, Zhong J, Lu P, et al. DOCK4 is a platinum-chemosensitive and prognostic-related biomarker in ovarian cancer. PPAR Res. 2021;2021:6629842. doi:10.1155/2021/6629842
30. Chandrashekar DS, Bashel B, Balasubramanya SAH, et al. UALCAN: a portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia. 2017;19(8):649–658. doi:10.1016/j.neo.2017.05.002
31. Lanczky A, Munkácsy G, Szabó A, et al. miRpower: a web-tool to validate survival-associated miRNAs utilizing expression data from 2178 breast cancer patients. Breast Cancer Res Treat. 2016;160(3):439–446. doi:10.1007/s10549-016-4013-7
32. Li T, Trousdale MD, LaBadie DR, et al. TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017;77(21):e108–e110. doi:10.1158/0008-5472.CAN-17-0307
33. Tang Z, Li C, Kang B, et al. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
34. Gao J, Aksoy BA, Dogrusoz U, et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013;6(269):pl1–pl1. doi:10.1126/scisignal.2004088
35. Szklarczyk D, Gable AL, Lyon D, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D607–D613. doi:10.1093/nar/gky1131
36. Franz M, Rodriguez H, Lopes C, et al. GeneMANIA update 2018. Nucleic Acids Res. 2018;46(W1):W60–W64. doi:10.1093/nar/gky311
37. Sawant A, Hensel JA, Chanda D, et al. Depletion of plasmacytoid dendritic cells inhibits tumor growth and prevents bone metastasis of breast cancer cells. J Immunol. 2012;189(9):4258–4265. doi:10.4049/jimmunol.1101855
38. Suzuki H, Oda I, Abe S, et al. High rate of 5-year survival among patients with early gastric cancer undergoing curative endoscopic submucosal dissection. Gastric Cancer. 2016;19(1):198–205. doi:10.1007/s10120-015-0469-0
39. Kim JW, Nam KH, Ahn S-H, et al. Prognostic implications of immunosuppressive protein expression in tumors as well as immune cell infiltration within the tumor microenvironment in gastric cancer. Gastric Cancer. 2016;19(1):42–52. doi:10.1007/s10120-014-0440-5
40. Shitara K, Özgüroğlu M, Bang Y-J, et al. Pembrolizumab versus paclitaxel for previously treated, advanced gastric or gastro-oesophageal junction cancer (KEYNOTE-061): a randomised, open-label, controlled, Phase 3 trial. Lancet. 2018;392(10142):123–133. doi:10.1016/S0140-6736(18)31257-1
41. Ralph C, Elkord E, Burt DJ, et al. Modulation of lymphocyte regulation for cancer therapy: a Phase II trial of tremelimumab in advanced gastric and esophageal adenocarcinoma. Clin Cancer Res. 2010;16(5):1662–1672. doi:10.1158/1078-0432.CCR-09-2870
42. Boureux A, Vignal E, Faure S, et al. Evolution of the Rho family of ras-like GTPases in eukaryotes. Mol Biol Evol. 2007;24(1):203–216. doi:10.1093/molbev/msl145
43. Namekata K, Kimura A, Kawamura K, et al. Dock GEFs and their therapeutic potential: neuroprotection and axon regeneration. Prog Retin Eye Res. 2014;43:1–16. doi:10.1016/j.preteyeres.2014.06.005
44. Murray DW, Didier S, Chan A, et al. Guanine nucleotide exchange factor Dock7 mediates HGF-induced glioblastoma cell invasion via Rac activation. Br J Cancer. 2014;110(5):1307–1315. doi:10.1038/bjc.2014.39
45. Xiao Y, Peng Y, Wan J, et al. The atypical guanine nucleotide exchange factor Dock4 regulates neurite differentiation through modulation of Rac1 GTPase and actin dynamics. J Biol Chem. 2013;288(27):20034–20045. doi:10.1074/jbc.M113.458612
46. Pagnamenta AT, Bacchelli E, de Jonge MV, et al. Characterization of a family with rare deletions in CNTNAP5 and DOCK4 suggests novel risk loci for autism and dyslexia. Biol Psychiatry. 2010;68(4):320–328. doi:10.1016/j.biopsych.2010.02.002
47. Park N, Kang H. BMP-Induced MicroRNA-101 expression regulates vascular smooth muscle cell migration. Int J Mol Sci. 2020;21(13):4764.
48. Sundaravel S, Duggan R, Bhagat T, et al. Reduced DOCK4 expression leads to erythroid dysplasia in myelodysplastic syndromes. Proc Natl Acad Sci USA. 2015;112(46):E6359–68. doi:10.1073/pnas.1516394112
49. Makihara S, Morin S, Ferent J, et al. Polarized dock activity drives Shh-Mediated axon guidance. Dev Cell. 2018;46(4):410–425 e7. doi:10.1016/j.devcel.2018.07.007
50. Yang W-H, Lan H-Y, Huang C-H, et al. RAC1 activation mediates Twist1-induced cancer cell migration. Nat Cell Biol. 2012;14(4):366–374. doi:10.1038/ncb2455
51. Zhu X, Gao G, Chu K, et al. Inhibition of RAC1-GEF DOCK3 by miR-512-3p contributes to suppression of metastasis in non-small cell lung cancer. Int J Biochem Cell Biol. 2015;61:103–114. doi:10.1016/j.biocel.2015.02.005
52. Alsum Z, Hawwari A, Alsmadi O, et al. Clinical, immunological and molecular characterization of DOCK8 and DOCK8-like deficient patients: single center experience of twenty five patients. J Clin Immunol. 2013;33(1):55–67. doi:10.1007/s10875-012-9769-x
53. Shulman Z, Pasvolsky R, Woolf E, et al. DOCK2 regulates chemokine-triggered lateral lymphocyte motility but not transendothelial migration. Blood. 2006;108(7):2150–2158. doi:10.1182/blood-2006-04-017608
54. Noy R, Pollard JW. Tumor-associated macrophages: from mechanisms to therapy. Immunity. 2014;41(1):49–61. doi:10.1016/j.immuni.2014.06.010
55. Tanaka A, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017;27(1):109–118. doi:10.1038/cr.2016.151
56. Ando M, Ito M, Srirat T, et al. Memory T cell, exhaustion, and tumor immunity. Immunol Med. 2020;43(1):1–9. doi:10.1080/25785826.2019.1698261
57. Kang JY, Gil M, Kim KE. Neuropilin 1 expression acts as a prognostic marker in stomach adenocarcinoma by predicting the infiltration of treg cells and M2 macrophages. J Clin Med. 2020;9(5):1430. doi:10.3390/jcm9051430
58. Gu-Trantien C, Loi S, Garaud S, et al. CD4(+) follicular helper T cell infiltration predicts breast cancer survival. J Clin Invest. 2013;123(7):2873–2892. doi:10.1172/JCI67428
59. Hodi FS, O’Day SJ, McDermott DF, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010;363(8):711–723. doi:10.1056/NEJMoa1003466
60. Brahmer JR, Tykodi SS, Chow LQM, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med. 2012;366(26):2455–2465. doi:10.1056/NEJMoa1200694
61. Jiang Z, Liu Z, Li M, et al. Immunogenomics analysis reveals that TP53 mutations inhibit tumor immunity in gastric cancer. Transl Oncol. 2018;11(5):1171–1187. doi:10.1016/j.tranon.2018.07.012
62. Rizvi NA, Hellmann MD, Snyder A, et al. Cancer immunology. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science. 2015;348(6230):124–128. doi:10.1126/science.aaa1348
63. Eroglu Z, Zaretsky JM, Hu-Lieskovan S, et al. High response rate to PD-1 blockade in desmoplastic melanomas. Nature. 2018;553(7688):347–350.
64. Senbabaoglu Y, Gejman RS, Winer AG, et al. Tumor immune microenvironment characterization in clear cell renal cell carcinoma identifies prognostic and immunotherapeutically relevant messenger RNA signatures. Genome Biol. 2016;17(1):231. doi:10.1186/s13059-016-1092-z
65. Goudarzi KM, Lindstrom MS. Role of ribosomal protein mutations in tumor development (Review). Int J Oncol. 2016;48(4):1313–1324. doi:10.3892/ijo.2016.3387
66. Pavlovic M, Arnal-Estapé A, Rojo F, et al. Enhanced MAF Oncogene expression and breast cancer bone metastasis. J Natl Cancer Inst. 2015;107(12):djv256. doi:10.1093/jnci/djv256
67. Sano T, Coit DG, Kim HH, et al. Proposal of a new stage grouping of gastric cancer for TNM classification: international Gastric Cancer Association staging project. Gastric Cancer. 2017;20(2):217–225. doi:10.1007/s10120-016-0601-9
68. Wagner AD, Syn NL, Moehler M, et al. Chemotherapy for advanced gastric cancer. Cochrane Database Syst Rev. 2017;8:CD004064. doi:10.1002/14651858.CD004064.pub4
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.