")
Back to Archived Journals » Research and Reports in Forensic Medical Science » Volume 9
DNA phenotyping: current application in forensic science
Received 26 October 2018
Accepted for publication 27 December 2018
Published 8 February 2019 Volume 2019:9 Pages 1—8
DOI https://doi.org/10.2147/RRFMS.S164090
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Henrik Druid
Leonardo Arduino Marano,1 Cintia Fridman2
1Laboratório de Genética Molecular Forense, Polícia Científica do Estado do Paraná, Curitiba, PR, Brazil; 2Departamento de Medicina Legal, Ética Médica e Medicina Social e do Trabalho, Faculdade de Medicina FMUSP, Universidade de São Paulo, São Paulo, SP, Brazil
Abstract: DNA analysis for forensic investigations is based on the idea that each individual is genetically unique, except in cases of monozygotic twins. DNA obtained from biological samples is able to individualize this material by direct comparison of short tandem repeats genetic profile, obtained from biological samples of unknown origin to a reference sample profile. One of the major limitations of this approach is the need for a reference sample for comparison. Numerous studies seeking to understand the relationship between certain polymorphisms and certain phenotypic characteristics are increasing and have generated promising results in aiding forensic sciences. The process of inferring externally visible characteristics (EVCs) with forensic purpose – eg, the color of skin, iris and hair, height, facial features, and male baldness pattern – from biological samples is known as forensic DNA phenotyping (FDP). Therefore, FDP provides more details about the subject to which a given biological sample belongs, without the need for a reference sample for comparative analysis. Some ethical and legal aspects should be taken into account so that this new technology does not promote segregation or ethnic persecution of certain population groups. Despite this, several real cases have benefited from these methods to orientate investigations to identify both suspects and victims.
Keywords: forensic DNA phenotyping, DNA, externally visible characteristics, forensic genetics
Introduction
Forensic human identification
Human identification based on genetic profiles obtained from DNA polymorphisms (short tandem repeats [STRs]) is considered to be the gold standard among forensic science techniques. STRs are polymorphisms generated by a sequence (in tandem) of copies of small DNA segments (ranging from 2 to 6 base pairs). The most informative STRs may present more than a dozen alleles, and thousands of these polymorphisms have already been identified in humans; some estimations point to the existence of about one million STRs distributed among the human genome.1,2
The biological samples collected at crime scenes are processed to obtain a DNA profile that will be compared to the suspect’s profile, assisting investigation to establish a connection between the offender and the crime scene, or even eliminating suspects.3 The same approach can be used to identify missing persons or unidentified bodies, comparing their profiles in familial searches.
The main advantage of STR markers is due to their high allele diversity, making such markers highly informative. The most polymorphic STRs have a high discriminating power (probability that two randomly selected individuals have distinct genotypes) and low probability of match (probability that two randomly selected individuals have identical genotypes).1,2 In practical terms, these values ensure that each individual in the world population (except for identical twins) may have a unique genetic profile.
The main disadvantage lies in the fact that this type of examination is uniquely comparative, requiring a pair of unknown/reference samples to be compared. In the absence of such a pair, the only possibility would be searching a DNA database containing suspects’ profiles. Another disadvantage is the structure of the STR markers, which are composed of repetitions from ~100 to 300 base pairs in length. There are situations in which biological material from the crime scene is so degraded that DNA samples obtained are not feasible of obtaining enough data for an accurate identification.4
In face of such difficulties using traditional STR markers protocols in some situations, various researches have allowed the use of genetic predictions of externally visible characteristics (EVCs) to be of assistance in police investigations, in both tracking suspects and identifying victims. Several advances in genetics appear to have identified potentially useful markers for the prediction of various physical characteristics.
Phenotype prediction from DNA markers
Several variations (Insertion/Deletion [InDels] and single nucleotide polymorphisms [SNPs]) located in DNA coding or regulatory regions can lead to amino acid substitutions, altering the functional properties of the translated protein and consequently being expressed in distinct phenotypes, some of them being the visible characteristics of the individual. In an attempt to obtain information about the physical features of individuals from DNA extracted from biological material, such as blood drops, hair strands, or small body fragments, the scientific community has intensively researched the association between genetic markers and physical traits. Some studies have already evaluated the existence of polymorphisms associated with skin, hair and eye color, facial forms, height, and baldness.
In this context, it is expected that the genotyping of such genetic markers in crime scene evidence, or in an unidentified body, can contribute significantly to increase the accurate information on the physical characteristics of those involved.5,6 Although some following inferences may not present definitive value as forensic evidence, they may be an important factor leading police investigation and reducing the number of suspects to a small set.5
DNA phenotyping
Eye color
Eye color can be considered as one of the human traits with the most color variability, ranging from light shades of blue to dark shades of brown or black, through intermediate colors such as gray, hazel, yellow, and green. This color difference follows a pattern similar to the pigmentation of the skin and hair, being defined by the amount of melanin and number of melanosomes in the outer layer of the iris: blue eyes have less melanin/melanosomes than brown eyes, for example.7
One of the first phenotyping tools developed and validated was the Irisplex System8 consisting of six SNPs distributed among pigmentation genes (HERC2, OCA2, SLC24A4, SLC45A2, TYR, and IRF4). This tool allows the differentiation between blue and brown eyes with high accuracy (>90%), confirmed both in homogeneous and admixed populations.9–13 Despite these results, the tool did not demonstrate the same accuracy in samples from Asian populations, suggesting that more studies should be performed on more distinct population samples.14
However, intermediate eye colors are still a problem, requiring further research to identify new genetic variants, since the accuracy in their predictions is still much lower when compared with blue and brown eyes.15–17 Despite the difficulty in predicting these intermediate colors, a study by Pośpiech et al18 demonstrated gene–gene interaction between three of the main pigmentation genes (HERC2, OCA2, and TYRP1) related to green eye color, thus aiding in the elaboration of future prediction models.
Another debated aspect contemplates gender as a possible influencing factor in the determination of eye pigmentation. It has been observed that women tend to have darker eyes (predominantly brown and green) than men (predominantly blue and gray) in some European countries.11,19,20 However, no genetic factor has yet been found to explain this difference, and more studies will be needed to evaluate this correlation.
Hair color
Hair color, as well as eye and skin colors, is among the most noticeable EVCs with a wide range of phenotypes. The main differences observed in hair color are the result of two types of melanin: brown/black eumelanin and red/yellow pheomelanin.21 Individuals with red hair display a relative increase in the amount of pheomelanin compared to eumelanin, whereas in dark hair the amount of eumelanin prevails and in blond hair there is little of each kind of melanin.22 This variability in hair color has probably arisen in Europe, evolving from a dark ancestral color, resulting from human mating preferences.23
Among several genes involved in the melanogenesis process, MC1R was one of the first to demonstrate a strong discriminating power for red hair, fair skin, and freckles. Subsequently, associations were made with other genes, such as SLC45A2, SLC24A5, and HERC2 and a predictive model based on 22 SNPs was created, reaching 81%–93% accuracy for each hair color category.24
A new system was then developed in 2013, adding 18 hair color markers to the 6 preexisting Irisplex SNPs, called the HIrisplex System.16 HIrisplex encompasses markers from MC1R, HERC2, OCA2, SLC45A2, KITLG, EXOC2, TYR, SLC24A4, IRF4, ASIP, and TYRP1 genes and, even though it has fewer markers than the model previously created by Branicki et al,24 it can reach similar accuracy values (75%–92%).25
However, current hair prediction models face a challenge: the accurate prediction of hair colors from individuals who have had hair color changes throughout life (eg, darker hair after childhood). Most studies do not contemplate the sampling of younger individuals, or question adult subjects about distinct phenotypes in early childhood. Therefore, prediction models are only elaborated with phenotypic information observed in adults, without taking into account informative markers for age-dependent phenotypes, partially explaining the lower accuracy value for blond hair (only 69.5% against 78.5%, 80%, and 87.5% for brown, red, and black colors, respectively) prediction by HIrisplex. A study done with young individuals found that hair darkening usually occurs between 6 and 13 years of age and that HIrisplex model incorrectly predicts hair phenotypes for those individuals who were blond only during early childhood, advising for the need to identify new markers that could reduce this error rate.26
Skin color
Skin color has been one of the most complex pigmentation phenotypes studied. It is believed that the skin pigmentation variability emerged as an evolutionary response to the intensity of ultraviolet radiation among different planet regions. Regions closer to the Equator line, with higher luminous intensity (high UV), would present a higher selective pressure, keeping dark skin with high frequency, while more distant regions, with less luminous intensity, would exert less selective pressure, allowing the appearance of lighter skin tones.27
This evolutionary factor makes the genotype/phenotype associations in mapping studies difficult, as well as resulting in correlations that only apply to a specific population group. While associations found in admixed populations did not have the same discriminatory power in more homogeneous populations, such as Europeans,28 other studies developed using homogeneous populations could not differentiate skin colors between discrete groups of Asian, African, and Native Americans.29
Taking into account this evolutionary obstacle, a global prediction model was developed based on 36 markers distributed among 16 pigmentation genes.17 This model was created taking into account three (light, dark, dark–black) or five (very pale, pale, intermediate, dark, dark–black) skin tones, obtaining prediction accuracies ranging from 83%–97% for the three-category scale to 72%–97% for the five-category scale. Some of these associations have previously been described in admixed populations for some of these genes, such as SLC24A5,30,31 HERC2,31 and SLC45A2,32 making these seemingly promising for future applications.
The results from the IrisPlex, HIrisPlex, and HIrisPlex-S systems were compiled into a publically available interactive tool used to predict eye, hair, and skin color from DNA data. From https://hirisplex.erasmusmc.nl/, one can use the tool to insert genotype data from the 41 markers available and obtain probabilities for three eye, four hair, and five skin color categories to get individual probabilities through their prediction model.
Height
Until 2008, only a few genes have been described as associated with human height. Subsequent association studies were performed in 2008 (in which 54 loci with direct correlation with height variation were observed) and in 2010, increasing the number of genetic markers to 180 and subsequently reaching almost 700 markers in 2014.33–37 Most of these genes are involved with growth-signaling pathways, such as the fibroblast growth factor, as well as genes expressed in important tissues such as the growth plate38 – oddly though, many of these markers are not directly involved in human growth pathways.
Even with significant increase in the number of height-related variants, there are still no significant values for prediction tests. While the initial studies obtained accuracy values of ~65%, the most current studies failed to raise this value >75%, demonstrating the large number of SNPs still to be discovered and how complex this trait may be.39 Moreover, human height may have a different etiology other than genetic aspects, such as gestational (placental features and maternal health aspects such as nutrition, pathologies, and drugs), hormonal, and environmental factors (nutrition and lifestyle) mainly during childhood.40
Facial features
Among all EVCs, the facial shape prediction is one of the major objectives when studying phenotyping, glimpsing the final “DNA facial composite”. The face morphology is studied from the distances between facial landmarks, as nostrils width, lips width, distance between eyes and face height.
Some of the genetic markers associated with facial features are initially found in syndromes and facial deformities diseases studies (such as cleft palate, cleft lip, and other craniofacial dysplasias). Some of these markers are then correlated to craniofacial development and consequently linked to the normal variation of facial shape.41 For example, PAX3 gene encodes a transcription factor present in neural crest cells, which was also related to Waardenburg syndrome and was later associated with the nasion position.42 Other candidate genes have been identified following patterns similar to PAX3, such as PRDM16 and TP63. However, similar to height determination, each of these genetic markers seems to have a small contribution toward the total face morphology.41
The approach used by Claes et al,43 based primarily on data obtained from admixed populations, employs a first step in which the sample ancestry and gender are used to create a base-face, in which data from 24 SNPs will subsequently be used, to convey nose, lips, face roundness, jaw, chin, and supraorbital crest information to this primary face. Other studies also found significant associations with facial width, eyebrow width, distance between eyes, columella inclination, nose bridge width, nostril width, and mouth shape.44,45
Baldness
It is empirically known that male pattern of baldness or androgenic alopecia has a strong hereditary factor, displaying a heritability of ~80%.46 Among the various loci possibly involved, the major ones are those on q12 region on X chromosome, containing AR/EDA2R genes directly linked to the production of androgen receptor and ectodysplasin A2 receptor, respectively, on the 20p11 region, and on genes EBF1, TARDBP, and HDAC9 with predictive potential. Together, these five SNPs have the best association values to date, with 76.2% accuracy, reaching 86.4% if other 15 markers are added (rs1041668, rs6625163, rs6625150, rs962458, rs12007229, rs2180439, rs913063, rs1160312, rs6113491, rs6461387, rs6945541, rs7349332, rs4679955, rs9668810, and rs10502861), demonstrating that even low prediction markers can have high accuracy when added to stronger ones.47
Age estimation
Using a different approach from those previously shown here (SNP typing mostly), epigenetics studies using DNA methylation detection technologies have been shown to be useful to the age estimation of an individual. The age estimation has great importance in the forensic context, since it is complementary to the data obtained by the EVCs mentioned here: besides reducing the number of suspects by predicting the age group of the sample donor, it can also complement the obtained facial composite.48
DNA methylation changes throughout an individual’s life – levels of methylation increase in childhood and then decrease after reaching adulthood.49 These changes can be measured and used to calculate an age estimate of an individual from biological samples of diverse origins (various tissues and body fluids) and various situations (either from human remains or from a crime scene) with high accuracy (deviation of 3.15 years in relation to the real chronological age) with as few as seven markers.50
Ancestry
Some specific DNA markers can bring information about the ancestral composition of an individual, allowing their biogeographic contributions to be detailed (Africa, Europe, Asia, Amerindian). Thus, the use of ancestry informative markers (AIMs) allows the inference of an individual’s ancestry, providing data to reinforce potential witnesses, or even bringing new information about crime scene evidence.51
However, information about ancestry cannot be used solely as a criterion for determining the appearance of an individual. One must understand the difference between ancestry and the mistaken concept of race: the percentage of an individual’s ancestral contribution will not necessarily reflect their appearance. This is especially noticeable in admixed population samples in which AIMs demonstrate that there is no direct correlation between appearance (ethnicity) and ancestral biogeographic origin.52
Legal and ethical aspects
In view of the various legal issues surrounding the sample collection from suspects for comparison, an advantage of DNA phenotyping approach is that it is more focused on obtaining genetic profiles from crime scene samples, thus not harming dignity or integrity rights. In addition, EVCs are publicly available features and therefore would not involve privacy issues.53
However, all those individuals who share the characteristics of a facial composite may be interviewed and required to donate samples for comparison to the crime sample in question. In these cases, it is necessary to raise questions about harassment that certain groups with a determined physical characteristic could suffer from the moment a phenotype is obtained from evidence. One must question whether such individuals will receive any protection, since they will now belong to a group of suspects solely by their physical appearance. Therefore, it should be noted if safety of such groups is being preserved to the detriment of investigation and public safety objectives, and if new legal and ethical regulations should be created to preserve the integrity and intimacy of people involved in DNA phenotyping-based investigations. Attention should also be drawn to the use of markers that are neutral in relation to ancestry, since information from some AIMs may be erroneously associated to certain phenotypes, resulting in ethnic persecution.54
Application and real cases
In 2010, a woman was sexually assaulted in broad daylight in Florida (USA) and no match was found in DNA databases. Seven years later, police contracted a private DNA phenotype company, which obtained a facial composite predicting a male subject with light brown skin, brown-hazel eyes, and black hair. The new DNA facial composite led investigation to the crime scene neighboring property, a wildlife sanctuary, where police found a suspect with matching characteristics. After a voluntary sample donation, Hugo Giron-Polanco was arrested since a DNA comparison showed him and the semen sample found on the victim shared the same STR profile with a 1:400,000,000,000 match probability.55 However, Parabon Nanolabs (the private DNA phenotype company contracted in this case) has not scientifically published its methodology or any validation tests so far.
Toronto Police Service has already submitted several cold case samples to private DNA phenotyping services, assisting to change the original direction of the investigations, although no actual arrests occurred.56 Several other cold cases across the world are counting on facial composites obtained from DNA profiles, with hope some new information will arise from this new data added to the investigation, hoping to identify both suspects and human remains.57–60
On May 15, 2018, the German state of Bavaria approved a law for police to analyze DNA samples and predict hair, eye, and skin color in addition to ancestry. These same DNA predictions have already been used in the Netherlands, France, UK, Canada, and several US states, although some of these countries have no precise laws regarding the practice.61
EVCs prediction can also be very useful in disaster victim identification and identification of missing persons, when no close relatives or antemortem samples exist for comparison. Forty-nine bone and teeth samples from World War II victims found in Slovenia had their DNA extracted and were typed for hair and eye pigmentation genetic markers. All 49 samples had a successful prediction of eye and hair color, two of the predictions being confirmed by the living sister of two of the victims.62
In the future, computers may even be able to search composites generated from DNA obtained from biological evidences against government databases or even photos from social media platforms. Even if this system does not find a perfect match, at least it will be able to narrow down the subjects of interest.56
Evaluation and validation
A recent initiative was done by a group of scientists to create a project aiming to establish new scientific knowledge, to develop, validate, and implement analysis tools that allow the prediction of an individual’s appearance from DNA samples for use in forensic routine. In 2017, the VISible Attributes Through GEnomics (VISAGE) Consortium was created, composed by members of academic, police, and justice institutions from eight European countries, with the goal to expand the use of DNA as a tool to quickly obtain facial composites from evidence. The limitation of current DNA forensic techniques is expected to be overcome and new technologies to be developed allowing police investigations to find unknown suspects more quickly through DNA information from crime scenes. The VISAGE Consortium intends to use massive parallel sequencing techniques to obtain higher quantitative data about suspect’s appearance, age, and ancestry predictions, as well as to create software that facilitates the interpretation of the generated data.63
Furthermore, the European DNA Profiling (EDNAP) Group is responsible for promoting meetings and developing collaborative comparison exercises to promote critical evaluations between laboratories around the world, aiming to test the reliability and consistency of new forensic DNA technologies. So far, EDNAP has tested the IrisPlex System reproducibility between 21 laboratories, considering it to be successful64 and has recently been evaluating the age prediction through DNA methylation analysis (data not published).
Although several studies cited here have shown significant phenotype–genotype association (Table 1), it should be noted that each target population has its own genetic background, and extrapolation of these data should be done with caution. Each set of genetic markers associated to a certain phenotype should be carefully evaluated in additional populations with a different genetic background from those originally studied.
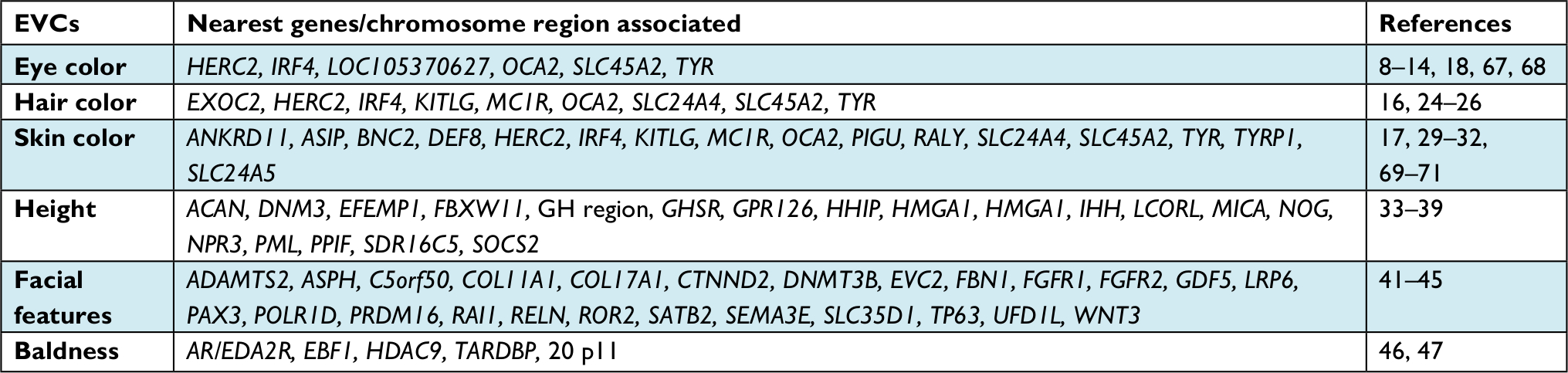 | Table 1 Summary of main EVCs and some associated genetic markers and references Abbreviation: EVCs, externally visible characteristics. |
As most studies are conducted in more genetically homogeneous populations, these found associations should be confirmed in admixed populations when possible, since these populations may present several challenging factors to already established associations, such as population stratification.
When association studies are done in multiple populations, it is possible to elucidate similarities and differences in genetic structure, limiting candidate regions for new variants identification.65 Thus, each population evolutionary history (eg, recombination, mutation, and drift) can influence the traceability of a variant, favoring its detection in one population than in another, confirming the importance of association studies in admixed and homogeneous populations combined.66
Conclusion
With all research done in genetic phenotyping presented here, it can be stated that obtaining a set of genetic markers that accurately predict most of human EVCs for forensic use is closer than ever, and a complete “DNA facial composite” is already at sight for forensic routine. Research still needs to be performed to confirm data obtained in diverse global populations and check if none of the associations found are due to ancestry or other populational background in addition to discovering new associations of physical characteristics. However, despite the many ethical and legal issues that still permeate this subject, the high statistical accuracy of most of these studies makes them viable for practical use in forensic routine.
Disclosure
The authors report no conflicts of interests in this work.
References
Butler JM. Forensic DNA Typing: Biology, Technology, and Genetics of STR Markers. 2nd ed. Amsterdam, Boston: Elsevier Academic Press; 2005. | ||
Goodwin W, Linacre A, Hadi S. An Introduction to Forensic Genetics. 2nd ed. Chichester, West Sussex, UK: Wiley-Blackwell; 2011. | ||
Bond JW. Value of DNA evidence in detecting crime. J Forensic Sci. 2007;52(1):128–136. | ||
Budowle B, Bieber FR, Eisenberg AJ. Forensic aspects of mass disasters: strategic considerations for DNA-based human identification. Leg Med (Tokyo). 2005;7(4):230–243. | ||
Walsh SJ. Recent advances in forensic genetics. Expert Rev Mol Diagn. 2004;4(1):31–40. | ||
Branicki W, Brudnik U, Kupiec T, Wolañska-Nowak P, Wojas-Pelc A. Determination of phenotype associated SNPs in the MC1R gene. J Forensic Sci. 2007;52(2):349–354. | ||
Sturm RA, Frudakis TN. Eye colour: portals into pigmentation genes and ancestry. Trends Genet. 2004;20(8):327–332. | ||
Walsh S, Liu F, Ballantyne KN, van Oven M, Lao O, Kayser M. IrisPlex: a sensitive DNA tool for accurate prediction of blue and brown eye colour in the absence of ancestry information. Forensic Sci Int Genet. 2011;5(3):170–180. | ||
Dembinski GM, Picard CJ. Evaluation of the IrisPlex DNA-based eye color prediction assay in a United States population. Forensic Sci Int Genet. 2014;9:111–117. | ||
Kastelic V, Pośpiech E, Draus-Barini J, Branicki W, Drobnič K. Prediction of eye color in the Slovenian population using the IrisPlex SNPs. Croat Med J. 2013;54(4):381–386. | ||
Pietroni C, Andersen JD, Johansen P, et al. The effect of gender on eye colour variation in European populations and an evaluation of the IrisPlex prediction model. Forensic Sci Int Genet. 2014;11:1–6. | ||
Pośpiech E, Karłowska-Pik J, Ziemkiewicz B, et al. Further evidence for population specific differences in the effect of DNA markers and gender on eye colour prediction in forensics. Int J Legal Med. 2016;130(4):923–934. | ||
Ruiz Y, Phillips C, Gomez-Tato A, et al. Further development of forensic eye color predictive tests. Forensic Sci Int Genet. 2013;7(1):28–40. | ||
Yun L, Gu Y, Rajeevan H, Kidd KK. Application of six IrisPlex SNPs and comparison of two eye color prediction systems in diverse Eurasia populations. Int J Legal Med. 2014;128(3):447–453. | ||
Walsh S, Lindenbergh A, Zuniga SB, et al. Developmental validation of the IrisPlex system: determination of blue and brown iris colour for forensic intelligence. Forensic Sci Int Genet. 2011;5(5):464–471. | ||
Walsh S, Liu F, Wollstein A, et al. The HIrisPlex system for simultaneous prediction of hair and eye colour from DNA. Forensic Sci Int Genet. 2013;7(1):98–115. | ||
Chaitanya L, Breslin K, Zuñiga S, et al. The HIrisPlex-S system for eye, hair and skin colour prediction from DNA: introduction and forensic developmental validation. Forensic Sci Int Genet. 2018;35:123–135. | ||
Pośpiech E, Draus-Barini J, Kupiec T, Wojas-Pelc A, Branicki W. Gene-gene interactions contribute to eye colour variation in humans. J Hum Genet. 2011;56(6):447–455. | ||
Sulem P, Gudbjartsson DF, Stacey SN, et al. Genetic determinants of hair, eye and skin pigmentation in Europeans. Nat Genet. 2007;39(12):1443–1452. | ||
Martinez-Cadenas C, Peña-Chilet M, Ibarrola-Villava M, Ribas G. Gender is a major factor explaining discrepancies in eye colour prediction based on HERC2/OCA2 genotype and the IrisPlex model. Forensic Sci Int Genet. 2013;7(4):453–460. | ||
Gerstenblith MR, Goldstein AM, Fargnoli MC, Peris K, Landi MT. Comprehensive evaluation of allele frequency differences of MC1R variants across populations. Hum Mutat. 2007;28(5):495–505. | ||
Rees JL. Genetics of hair and skin color. Annu Rev Genet. 2003;37(1):67–90. | ||
Frost P. European hair and eye color: a case of frequency-dependent sexual selection? Evol Hum Behav. 2006;27(2):85–103. | ||
Branicki W, Liu F, van Duijn K, et al. Model-based prediction of human hair color using DNA variants. Hum Genet. 2011;129(4):443–454. | ||
Walsh S, Chaitanya L, Clarisse L, et al. Developmental validation of the HIrisPlex system: DNA-based eye and hair colour prediction for forensic and anthropological usage. Forensic Sci Int Genet. 2014;9:150–161. | ||
Kukla-Bartoszek M, Pośpiech E, Spólnicka M, et al. Investigating the impact of age-depended hair colour darkening during childhood on DNA-based hair colour prediction with the HIrisPlex system. Forensic Sci Int Genet. 2018;36:26–33. | ||
Jablonski NG, Chaplin G. The colours of humanity: the evolution of pigmentation in the human lineage. Philos Trans R Soc Lond B Biol Sci. 2017;372(1724):20160349. | ||
Maroñas O, Phillips C, Söchtig J, et al. Development of a forensic skin colour predictive test. Forensic Sci Int Genet. 2014;13:34–44. | ||
Liu F, Visser M, Duffy DL, et al. Genetics of skin color variation in Europeans: genome-wide association studies with functional follow-up. Hum Genet. 2015;134(8):823–835. | ||
de Araújo Lima F, de Toledo Gonçalves F, Fridman C. SLC24A5 and ASIP as phenotypic predictors in Brazilian population for forensic purposes. Leg Med (Tokyo). 2015;17(4):261–266. | ||
Cerqueira CC, Hünemeier T, Gomez-Valdés J, et al. Implications of the admixture process in skin color molecular assessment. PLoS One. 2014;9(5):e96886. | ||
Fracasso NCA, de Andrade ES, Wiezel CEV, et al. Haplotypes from the SLC45A2 gene are associated with the presence of Freckles and eye, hair and skin pigmentation in Brazil. Leg Med (Tokyo). 2017;25:43–51. | ||
Wood AR, Esko T, Yang J, et al. Defining the role of common variation in the genomic and biological architecture of adult human height. Nat Genet. 2014;46(11):1173–1186. | ||
Lango Allen H, Estrada K, Lettre G, et al. Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature. 2010;467(7317):832–838. | ||
Gudbjartsson DF, Walters GB, Thorleifsson G, et al. Many sequence variants affecting diversity of adult human height. Nat Genet. 2008;40(5):609–615. | ||
Lettre G, Jackson AU, Gieger C, et al. Identification of ten loci associated with height highlights new biological pathways in human growth. Nat Genet. 2008;40(5):584–591. | ||
Weedon MN, Lango H, Lindgren CM, et al. Genome-wide association analysis identifies 20 loci that influence adult height. Nat Genet. 2008;40(5):575–583. | ||
Guo MH, Hirschhorn JN, Dauber A. Insights and implications of genome wide association studies of height. J Clin Endocrinol Metab. 2018;103(9):3155–3168. | ||
Liu F, Hendriks AE, Ralf A, et al. Common DNA variants predict tall stature in Europeans. Hum Genet. 2014;133(5):587–597. | ||
Argente J. Challenges in the management of short stature. Horm Res Paediatr. 2016;85(1):2–10. | ||
Liu F, van der Lijn F, Schurmann C, et al. A genome-wide association study identifies five loci influencing facial morphology in Europeans. PLoS Genet. 2012;8(9):e1002932. | ||
Paternoster L, Zhurov AI, Toma AM, et al. Genome-wide association study of three-dimensional facial morphology identifies a variant in PAX3 associated with nasion position. Am J Hum Genet. 2012;90(3):478–485. | ||
Claes P, Hill H, Shriver MD. Toward DNA-based facial composites: preliminary results and validation. Forensic Sci Int Genet. 2014;13:208–216. | ||
Fagertun J, Wolffhechel K, Pers TH, et al. Predicting facial characteristics from complex polygenic variations. Forensic Sci Int Genet. 2015;19:263–268. | ||
Adhikari K, Fuentes-Guajardo M, Quinto-Sánchez M, et al. A genome-wide association scan implicates DCHS2, Runx2, Gli3, Pax1 and EDAR in human facial variation. Nat Commun. 2016;7(1):11616. | ||
Nyholt DR, Na G, Heath AC, Martin NG. Genetic basis of male pattern baldness. J Invest Dermatol. 2003;121(6):1561–1564. | ||
Marcińska M, Pośpiech E, Abidi S, et al. Evaluation of DNA variants associated with androgenetic alopecia and their potential to predict male pattern baldness. PLoS One. 2015;10(5):e0127852. | ||
Vidaki A, Kayser M. Recent progress, methods and perspectives in forensic epigenetics. Forensic Sci Int Genet. 2018;37:180–195. | ||
Jones MJ, Goodman SJ, Kobor MS. DNA methylation and healthy human aging. Aging Cell. 2015;14(6):924–932. | ||
Hong SR, Jung SE, Lee EH, Shin KJ, Yang WI, Lee HY. DNA methylation-based age prediction from saliva: high age predictability by combination of 7 CpG markers. Forensic Sci Int Genet. 2017;29:118–125. | ||
Phillips C. Forensic genetic analysis of bio-geographical ancestry. Forensic Sci Int Genet. 2015;18:49–65. | ||
Cardena MMSG, Ribeiro-dos-Santos A, Santos S, Mansur AJ, Pereira AC, Fridman C. Assessment of the relationship between self-declared ethnicity, mitochondrial haplogroups and genomic ancestry in Brazilian individuals. PLoS One. 2013;8(4):e62005. | ||
Kayser M. Forensic DNA phenotyping: predicting human appearance from crime scene material for investigative purposes. Forensic Sci Int Genet. 2015;18:33–48. | ||
Williams R, Wienroth M. Social and ethical aspects of forensic genetics: a critical review. Forensic Sci Rev. 2017;29(2):145–169. | ||
Rodriguez L. South Florida police bring new life to cold cases with DNA phenotyping; 2018. Available from: https://www.nbcmiami.com/news/local/South-Florida-Police-Bring-New-Life-to-Cold-Cases-With-DNA-Analysis-483093561.html. Accessed October 24, 2018. | ||
Pollack A. Building a face, and a case, on DNA; 2015. Available from: https://www.nytimes.com/2015/02/24/science/building-face-and-a-case-on-dna.html. Accessed October 24, 2018. | ||
KPLC News. Man arrested in 2009 killing of Sierra Bouzigard. 2017; Available from: http://www.kplctv.com/story/35955469/watch-live-at-130-pm-cpso-news-conference-regarding-cold-case-murder/. Accessed October 24, 2018. | ||
Kiger PJ. Composite faces from DNA help solve cold cases; 2018. Available from: https://science.howstuffworks.com/life/genetic/composite-faces-from-dna-help-solve-cold-cases.htm. Accessed October 24, 2018. | ||
Naham M. “DNA phenotyping” gives human skull found 41 years ago a face in cold case; 2018. Available from: https://lawandcrime.com/crazy/dna-phenotyping-gives-human-skull-found-41-years-ago-a-face-in-cold-case/. Accessed October 24, 2018. | ||
Butler C. It sounds like science fiction, but police created this sketch out of a suspect’s DNA; 2018. Available from: https://www.cbc.ca/news/canada/london/thera-dieleman-dna-composite-sketch-cold-case-1.4802027. Accessed October 24, 2018. | ||
Vogel G. German law allows use of DNA to predict suspects’ looks. Science. 2018;360(6391):841–842. | ||
Chaitanya L, Pajnič IZ, Walsh S, Balažic J, Zupanc T, Kayser M. Bringing colour back after 70 years: predicting eye and hair colour from skeletal remains of World War II victims using the HIrisPlex system. Forensic Sci Int Genet. 2017;26:48–57. | ||
Erasmus MC Press Release. EU horizon 2020-funded VISAGE project kicks off; 2017. Available from: https://www.erasmusmc.nl/perskamer/archief/2017/6234017/?lang=en. Accessed December 1, 2018. | ||
Chaitanya L, Walsh S, Andersen JD, et al. Collaborative EDNAP exercise on the IrisPlex system for DNA-based prediction of human eye colour. Forensic Sci Int Genet. 2014;11:241–251. | ||
Rosenberg NA, Huang L, Jewett EM, Szpiech ZA, Jankovic I, Boehnke M. Genome-wide association studies in diverse populations. Nat Rev Genet. 2010;11(5):356–366. | ||
McCarthy MI, Hirschhorn JN. Genome-wide association studies: potential next steps on a genetic journey. Hum Mol Genet. 2008;17(R2):R156–R165. | ||
Kayser M, Liu F, Janssens AC, et al. Three genome-wide association studies and a linkage analysis identify HERC2 as a human iris color gene. Am J Hum Genet. 2008;82(2):411–423. | ||
Mengel-From J, Børsting C, Sanchez JJ, Eiberg H, Morling N. Human eye colour and HERC2, OCA2 and MATP. Forensic Sci Int Genet. 2010;4(5):323–328. | ||
Donnelly MP, Paschou P, Grigorenko E, et al. A global view of the OCA2-HERC2 region and pigmentation. Hum Genet. 2012;131(5):683–696. | ||
Walsh S, Chaitanya L, Breslin K, et al. Global skin colour prediction from DNA. Hum Genet. 2017;136(7):847–863. | ||
Visser M, Palstra RJ, Kayser M. Human skin color is influenced by an intergenic DNA polymorphism regulating transcription of the nearby BNC2 pigmentation gene. Hum Mol Genet. 2014;23(21):5750–5762. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.