")
Back to Journals » Infection and Drug Resistance » Volume 13
Discovery of a Novel Hypervirulent Acinetobacter baumannii Strain in a Case of Community-Acquired Pneumonia
Authors Zhou H , Larkin PMK , Huang J, Yao Y, Zhu B, Yang Q , Hua X , Zhou J , Yang S , Yu Y
Received 28 December 2019
Accepted for publication 2 April 2020
Published 21 April 2020 Volume 2020:13 Pages 1147—1153
DOI https://doi.org/10.2147/IDR.S244044
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Hua Zhou,1,* Paige MK Larkin,2,* Jinwei Huang,3,* Yake Yao,1 Bingquan Zhu,4 Qing Yang,5 Xiaoting Hua,6,7 Jianying Zhou,1 Shangxin Yang,2,8 Yunsong Yu6,7
1Department of Respiratory Diseases, First Affiliated Hospital of Zhejiang University, Hangzhou, Zhejiang, People’s Republic of China; 2Department of Pathology & Laboratory Medicine, University of California, Los Angeles, Los Angeles, CA, USA; 3Department of Respiratory Diseases, Sixth Affiliated Hospital of Wenzhou Medical University, Lishui, Zhejiang, People’s Republic of China; 4Department of Child Health Care, Zhejiang University Children’s Hospital, Hangzhou, Zhejiang, People’s Republic of China; 5State Key Laboratory for Diagnostic and Treatment of Infectious Diseases, First Affiliated Hospital of Zhejiang University, Hangzhou, People’s Republic of China; 6Department of Infectious Diseases, Sir Run Run Shaw Hospital of Zhejiang University, Hangzhou, People’s Republic of China; 7Key Laboratory of Microbial Technology and Bioinformatics of Zhejiang Province, Hangzhou, People’s Republic of China; 8Zhejiang-California International Nanosystems Institute, Zhejiang University, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shangxin Yang; Yunsong Yu
Department of Infectious Diseases, Sir Run Run Shaw Hospital of Zhejiang University, Hangzhou 310016, People’s Republic of China
Tel +86 571 87236873
Email [email protected]; [email protected]
Purpose: Acinetobacter baumannii is associated with both hospital-acquired infections and community-acquired pneumonia (CAP). Here, we describe a novel strain of A. baumannii in a case of CAP in a previously healthy rural villager from Central Eastern China.
Materials and Methods: A. baumannii isolated from the patient (LS01) was compared to well-characterized pathogenic strain (AB5075), nosocomial circulating strain in China (ZJ06), and wild-type strain (ATCC17978). Growth rate studies were conducted under different environmental stressors, and virulence studies were performed using Galleria mellonella larvae. Whole genome sequencing (WGS) was performed using MinIon and MiSeq. Center for Genomic Epidemiology, CLCbio, Geneious, and Virulence Factors of Pathogenic Bacteria database were used for genomic analysis.
Results: LS01 grew significantly faster at 37°C and 42°C and in the presence of zinc compared to other strains. LS01 was more virulent in G. mellonella, killing all larvae within 8 h. Although WGS revealed 44 virulence genes, these genes were also present in the other strains. While two chromosomally encoded β-lactamases were identified, there were no plasmids identified and LS01 was pan-susceptible to all antibiotics tested. Phylogenetic analysis revealed that the closest related strains were only 72.552% identical, supporting a novel strain.
Conclusion: LS01 is a novel strain of hypervirulent yet pan-drug susceptible A. baumannii isolated from a patient with no prior hospitalizations, sick contacts, or any of the typical risk factors. This raises concerns for an emerging pathogen, and more epidemiological studies should be conducted to assess the prevalence of this A. baumannii strain.
Keywords: Acinetobacter baumannii, hypervirulent, community-acquired pneumonia, novel strain, emerging pathogen
Introduction
Acinetobacter baumannii, a glucose-non-fermentative aerobic Gram-negative coccobacilli, causes up to 20% of intensive care unit (ICU) infections worldwide, including ventilator-associated and blood stream infections.1,2 A. baumannii has a plastic genome, allowing for rapid mutations, acquisition of new genes, and increased virulence in response to environmental stress,1,3-5 posing a difficult challenge for treatment and infection control. With the global rise in antibiotic resistance, multi-drug resistant (MDR) A. baumannii is an increasing threat for both nosocomial and community-acquired infections. Mechanisms for resistance include mobile genetic elements,6 efflux pumps, altered permeability, modification of target sites, and β-lactamases, with all four classes of β-lactamases identified in A. baumannii.2 Although both tigecycline and colistin have been used effectively in the treatment of A. baumannii previously, resistant isolates have been reported.7,8
While traditionally associated with nosocomial infections, A. baumannii is more frequently attributed to community-acquired pneumonia (CAP) in tropical and sub-tropical climates.4,9,10 Although CAP strains are typically more susceptible to antibiotics than strains implicated in hospital acquired infections (HAI), CAP caused by A. baumanni is characterized by a fulminant and severe infection with high mortality.9 Here we describe a novel virulent A. baumannii strain (designated LS01) in a case of CAP in Central Eastern China. We investigated its growth pattern, virulence, antimicrobial susceptibility, and genomic characteristics to better understand the microbiology and pathogenicity of this novel strain.
Materials and Methods
Bacterial Isolates and Antimicrobial Susceptibility Testing
In addition to LS01 isolated from this case, other A. baumannii strains used in this study included a well characterized pathogenic strain AB5075,11 a well characterized nosocomial strain ZJ06 circulating in China,6 and a wild-type control strain ATCC17978. LS01 was identified by the Vitek GNI card (BioMe´rieux, Marcy-l’Etoile, France) as members of the Acinetobacter calcoaceticus-A. baumannii complex. The bacteria were cultured in Luria–Bertani (LB) medium at 37°C. Susceptibility testing for strains was performed using the Etest strip according to the manufacturer’s instructions. The results were interpreted according to the CLSI breakpoints (Clinical and Laboratory Standards Institute, 28th Edition).
Efficiency-of-Plating Experiments
Overnight cultures of the indicated strains were diluted into fresh LB medium and grown for 4 h. Aliquots of the cultures were serially diluted, and 100-μL volumes were spotted onto Mueller–Hinton (MH) agar plates containing the indicated stressors as follows: for acidic pH, MH medium adjusted to pH 5.5 or pH 4.5 with HCl prior to autoclaving; zinc sulfate (ZnSO4) was added to achieve a concentration of 1.25 mM; 30 oC/42 oC were selected as low/high temperature condition. Efficiency of plating (EOP) was calculated by dividing the CFU recovered on stress medium by the CFU recovered on plain MH grown at 37°C. EOP experiments were repeated at least three times, with the most representative colonies shown (Supplementary Figure 1).
Bacterial Growth Curves and Fitness Cost Measurement
Four independent cultures per strain were grown over night in MH broth, diluted 1:1000 into fresh MH broth and aliquoted into a flat-bottom 100-well plate in four replicates. The pH 4.5/pH5.5/Zn2+ MH broth were prepared as described above. The normal/pH 4.5/pH 5.5/Zn2+ plates were agitated at 37 °C, and two normal plates were agitated at 30 °C/42 °C, respectively. The OD600 of each culture was determined every 5 min for 16 h using the Bioscreen C MBR machine (Oy Growth Curves Ab Ltd., Finland). The growth rate was estimated by an R script (http://dx.doi.org/10.3389/fmicb.2016.01715). Statistical analyses were performed using GraphPad Prim 5. Two-way analysis of variance (ANOVA) was used to assess differences between the means. A P value of <0.05 was considered as statistically significant.
Galleria mellonella Infection
Infection of G. mellonella was performed as previously described.11,12 A. baumannii strains were grown overnight in an orbital shaker (37°C, 200 rpm), and overnight cultures were then diluted 100-fold into fresh medium and grown for 4 h. Cells were collected by centrifugation (5 min, 5000×g), washed once in phosphate-buffered saline solution (PBS), and resuspended in PBS to a final OD600 of 1.0. Further dilutions were done in PBS. The number of bacterial cells in the injected sample was enumerated by plating 10-fold serial dilutions on MH agar plates and counting CFU after overnight incubation.
G. mellonella larvae weighing 200 to 300 mg were used in the survival assays. Briefly, 10 μL of the sample was injected into the last left proleg of the larvae using a 10-μL glass syringe (Hamilton, Reno, NV) fitted with a 30G needle (Novo Nordisk, Princeton, NJ). Each experiment included control groups of larvae injected with 10μL sterile PBS. Injected larvae were incubated at 37°C, assessing death at 2h intervals in the first 24h, and 8h intervals in the next 48 h, with a total monitor time of 72h. Larvae were considered dead if they did not respond to physical stimuli. Experiments in which 10% or more of the larvae in either of the control groups died were omitted from the statistical analysis. The survival experiments were repeated three times using 10 larvae per experimental group, and the most representative curves were presented. Significance of survival differences was assessed with the log rank test using GraphPad Prim 5. A P value of <0.05 was considered as statistically significant.
Whole Genome Sequencing and Analysis
Whole genome sequencing (WGS) by Hiseq (Illumina, Carlsbad, CA) were performed as described previously.8 Bacteria from a single colony were cultured overnight at 37 °C in MH broth. The genomic DNA was extracted via a QIAamp DNA minikit (Qiagen, Valencia, CA, USA) following the protocol of the manufacturer’s protocol. Agarose gel electrophoresis and a NanoDrop spectrophotometer (ND-1000, ThermoFisher, Waltham, MA, USA) were used to determine the quality and quantity of the extracted genomic DNA. The DNA library was prepared using Nextera XT kit (Illumina, Carlsbad, CA) and sequenced by using 2x150 bp paired-end sequencing protocol. WGS by MinION (Nanopore, Oxford, UK) was performed following the manufacturer’s protocols. The complete bacterial genome was generated by combining the Hiseq sequences and MinION sequences through a hybrid assembly using Unicycler.13
Determination of plasmid replicons and antimicrobial resistance (AMR) performed using online Center for Genomic Epidemiology (CGE) tools (http://www.genomicepidemiology.org/).14 Virulence genes were identified using the VF Analyzer part of the Virulence Factors of Pathogenic Bacteria (VFDB) database using the default settings.15 Only the results meeting the following criteria are considered valid: Score (bits) >100 and % Identity >90%. Closely related species were identified using CGE Kmer-Finder. Single nuclear polymorphism (SNP) tree was generated using CGE CSI-Phylogeny and visualized using Geneious (Biomatters, New Zealand). Geneious was also used to perform sequence alignment analysis. A Kmer tree was constructed using CLC Genomics Workbench v12 (Qiagen, Valencia, CA, USA).
Results
Case History
A previously healthy 52 years old male villager from Lishui, a town in the Southern Zhejiang Province was admitted to the Sixth affiliated Hospital of Wenzhou Medical University for severe CAP and respiratory failure. The patient presented with productive cough, high fever (39.1°C), chills, fatigue, anorexia, chest tightness, shortness of breath, and abdominal pain. Pulmonary CT showed lobar pneumonia in the right lobe and pleural effusion. Blood test results were consistent with sepsis (white blood cells: 17.7x109/L with 91.8% neutrophils, C-reactive protein: > 200mg/L and procalcitonin: 41.04 ng/mL). Both blood and sputum culture grew A. baumannii with the same antibiogram (Table 1). This patient was successfully treated with imipenem 1g IV q8h and levofloxacin 0.5g qd, with symptom improved after four weeks of treatment. He was discharged after five weeks of hospitalization and the follow-up CT exam showed the lung lesions were significantly resolved three months later. The patient had been a vegetable farmer in a small village of less than 100 people, with no known sick contacts. The patient had also no history of prior hospitalizations nor recent travel prior to the illness. The isolate from the patient’s blood (designated LS01) was used for all experiments and genomic analysis in this study.
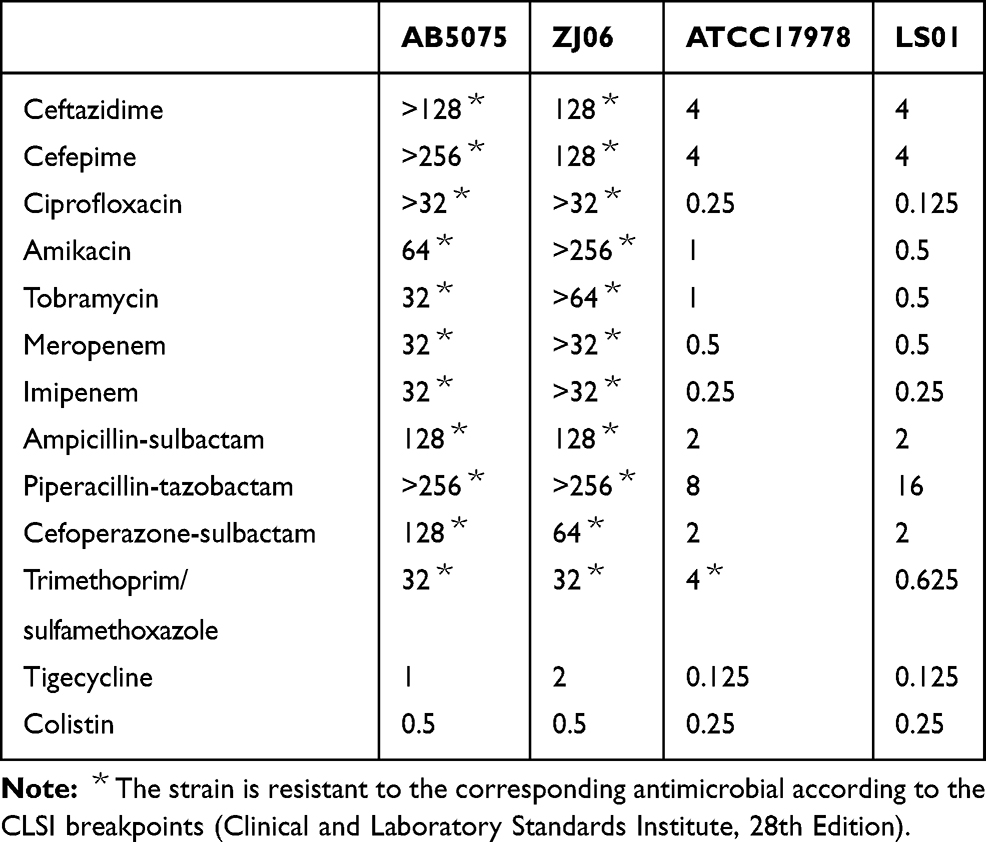 |
Table 1 Antimicrobial Susceptibility Profile of A. baumannii Strains |
Characterization of Growth and Virulence of the LS01 Strain
LS01 strain grew significantly faster than other strains AB5075, ZJ06, and ATCC17978 at 37°C, 42°C, and when Zn2+ was added (P<0.001) (Figure 1). LS01 strain was also significantly more virulent than the other strains tested as it killed 100% of G. mellonera larvae within 8 h, while no other strains killed 100% of larvae within 72 h (Figure 2). WGS revealed multiple virulence genes including 14 genes associated with biofilm formation, 1 gene associated with adherence, 2 phospholipase genes, 18 genes associated with iron uptake, and 2 signal transduction genes (Supplementary Table S1). However, none of these virulence genes were unique to LS01 as they are also found in other strains included in this study (data not shown).
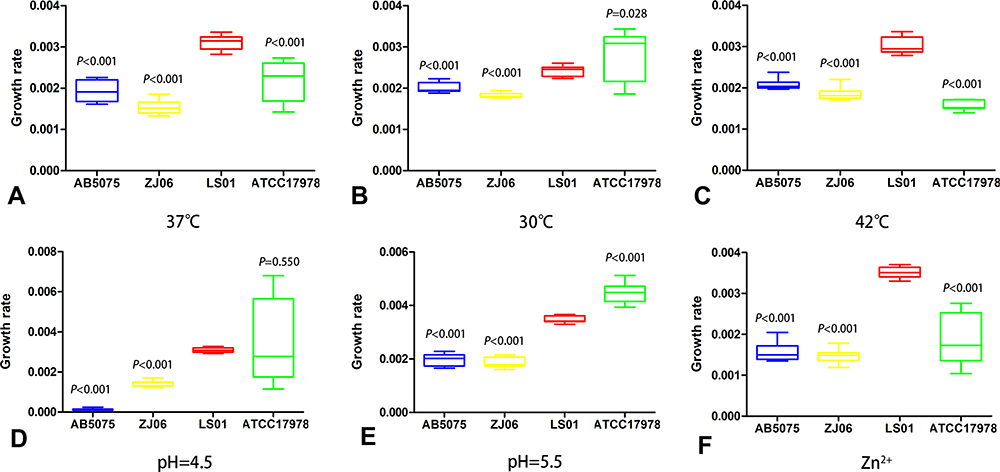 |
Figure 1 Growth rate study. Growth rate of AB5075, ZJ06, LS01, and ATCC17978 strains were measured at (A) 37°C, (B) 30°C, (C) 42°C, (D) pH=4.5, (E) pH=5.5, and (F) Zn2+. Two-way analysis of variance (ANOVA) was used to assess differences between the means. |
 |
Figure 2 G. mellonera killing study. The percent survival of G. mellonera after infection with AB5075, ZJ06, LS01, and ATCC17978 was measured for 72 h after infection. Statistical significance was calculated using log rank test. The survival experiments were repeated three times (n=10 larvae per experimental group), and the most representative curves were presented. |
Characterization of Drug Susceptibility and AMR Genes
LS01 was susceptible to all antibiotics tested and its susceptibility pattern was most similar to the ATCC17978 strain (Table 1). Whole genome sequencing data matched this as no plasmid replicon was detected and there were no acquired AMR genes except for chromosome carrying β-lactamase genes (blaADC-25 and blaOXA-91). When comparing to the reference resistance genes, LS01 had 99.88% identity and 96.18% identity to blaOXA-91 (DQ519086) and blaADC-25 (EF016355), respectively. There was a single silent mutation in blaOXA-91 (G>A at nucleotide position 720). However, ISAba1 insertion sequence was not found upstream of blaOXA-91, which has been shown to be necessary to turn on the expression of this gene.21 Six non-synonymous mutations were identified for blaADC-25 including E119V, K150Q, S167P, A270T, V347I, and N379S, but it is not clear how these mutations affect the enzymatic function.
Phylogenetic Analysis of the Novel A. baumanii Strain
LS01 species identification was confirmed by NCBI BLAST based on the full-length 16S ribosomal RNA gene sequence extracted from the WGS data, with > 99.8% identity to A. baumannii and <99.0% identity to other Acinetobacter species. The A. baumanii strains associated with the following names and NCBI accession numbers were identified as the most closely related: XH858 (NZ_CP014528.1), MRSN15313 (NZ_CP033869), ATCC 17978-mff (NZ_CP012004.1), ZW85-1 (NC_023028.1), CA-17 (NZ_CP012587.1), Ab04-mff (NZ_CP012006.1), IOMTU 433 (NZ_AP014649.1), and AB042 (NZ_CP019034.1). A pathogenic strain AB5075-UW (NZ_CP008706.1) and the main nosocomial strain circulating in China MDR-ZJ06 (CP001937.2) were also included for comparison. The most closely related A. baumanii strains were AB04-mff and XH858 (Figure 3). However, the percentage identity between LS01 and those two strains was only 72.5% (Supplementary Table S2). LS01 was even less related to the predominant circulating A. baumanii strain in China MDR-ZJ06 and a highly pathogenic strain AB5075-UW with percentage identify of 65.1% and 64.4%, respectively (Supplementary Table S2).
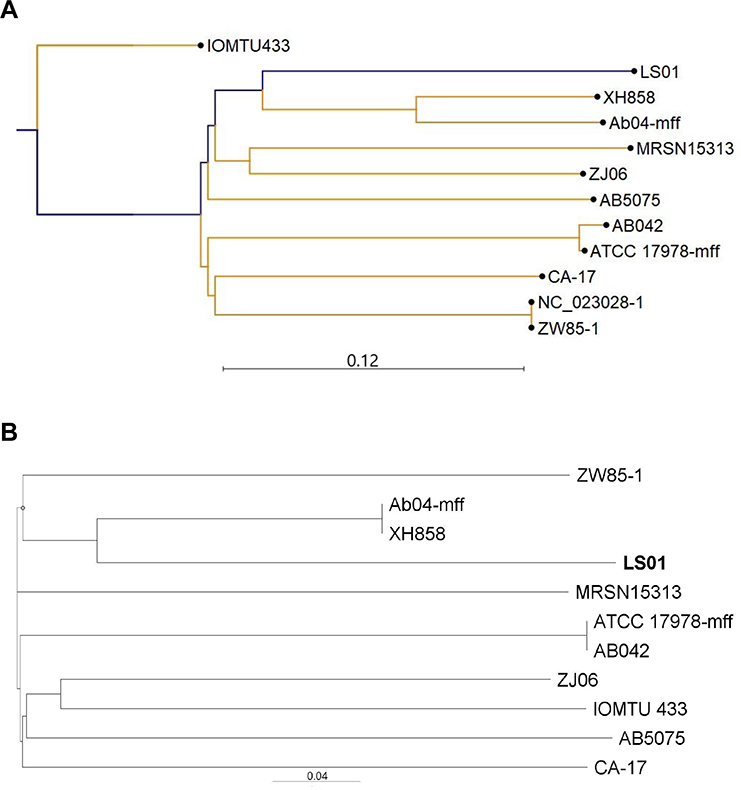 |
Figure 3 Phylogenetic analysis of the novel A. baumannii strain LS01. (A) K-mer tree; (B) single nuclear polymorphism (SNP) tree. |
Discussion
This was an unusual case of CAP caused by a novel wild-type A. baumannii strain (LS01). It remains uncertain how this patient became infected with this virulent strain as the patient was a vegetable grower in a rural village of less than 100 people, with no history of previous hospitalizations or sick contacts. Individuals with chronic pulmonary disease, renal disease, heavy smoking, diabetes mellitus, or excessive alcohol consumption have an increased risk for developing A. baumannii CAP.4 Our patient met none of these criteria, further posing the question of the source of infection. One potential hypothesis for acquisition of this hypervirulent A. baumannii strain is through contact with the environment. Although there has not been a natural source of A. baumannii identified,16 Acinetobacter has been found in water and food sources in wet climates, with antibiotic resistant strains isolated from these sources.4 In fact, MDR A. baumannii isolates from urban sewage and river water matched strains in hospital-acquired pneumonia,17 supporting a connection between clinical and environmental strains.
CAP due to A. baumannii is typically more fulminant compared to HAP. This is evidenced by an overall higher mortality,18 with mortality occurring within 48–72 h after hospital admission.9 In one study, all patients with CAP A. baumannii developed an acute clinical presentation, including septic shock, acute kidney injury, or respiratory failure and over half of the patients died.10 Consistent with these studies, our patient presented with severe pneumonia and bacteremia.
The virulence of this strain, as demonstrated by the moth killing experiment (Figure 2) is striking. Increased virulence in A. baumannii has been shown to be attributed to porins, capsular polysaccharides, protein secretion systems, biofilm formation, and metal acquisition systems.2,12 The thick capsular polysaccharide of A. baumannii reduces the susceptibility to complement-mediated killing, while the ability to form biofilms promotes persistent growth in the healthcare setting.1 A. baumannii actively pumps out chlorhexidine, a common hospital antiseptic that targets cell membranes, using the Acinetobacter chlorhexidine (AceI) efflux protein, rendering the disinfectant less effective.1 One study found that nearly a quarter of air samples from patient rooms were contaminated with carbapenem-resistant A. baumannii,4 demonstrating the potential airborne spread of the bacteria. Furthermore, virulence is maintained despite desiccation and starvation.5
In the case of this isolate, LS01, multiple virulence factors were identified including genes associated with biofilm formation, adherence, phospholipase, and iron uptake. Interestingly, many of these virulence genes have also been identified in MDR A. baumannii including ompA, bap, bfmS, csgA csuE, and pgaB19 as well as the ATCC 17978-mff, AB5075-UW, and MDR-ZJ06 strains (data not shown). Despite the prevalence of these genes in MDR A. baumannii, one study found that there was no significant difference in the presence of ompA and bap genes between environmental and clinical isolates.20 This poses the question of why LS01 is so virulent despite no known unique virulence factors identified. This will be the focus of future studies. One limitation of this study is that the database used to identify virulence genes does not provide information regarding chromosomal mutations or structural changes that may impact virulence. The other limitation is that we did not include more clinical A. baumannii strains in this study, which may help elucidate the reasons for this hypervirulent phenotype.
Although hospital-acquired A. baumannii strains are typically resistant to most drugs, CAP strains are usually less resistant, as is the case with this CAP strain, LS01. LS01 was susceptible to all drugs tested (Table 1) despite possessing naturally occurring β-lactamases blaOXA-91 and blaADC-25. BlaOXA-91 is a blaOXA-51-type β-lactamase, which does not contribute to intrinsic β-lactam resistance unless an ISAba1 insertion sequence is located upstream.21,22 BlaADC-25 is a chromosomally encoded cephalosporinase that typically confers resistance to all β-lactams.23 Our results showed that there are no identified mutations that could account for change in function of blaOXA-91, matching our susceptibility testing. However, six non-synonymous mutations identified in blaADC-25 could potentially change the enzymatic activity of this β-lactamase. Further studies are needed to understand the mechanism behind the pan-susceptible nature of LS01.
Taken together, we describe a novel community-acquired hypervirulent strain of A. baumannii causing a severe pneumonia in a previously healthy individual. The discovery of this strain raises concerns for an emerging pathogen, which is more than 25% genetically different from all other known A. baumannii strains. It should be noted that A. baumannii is known to have a very plastic genome with 25-46% of the genes unique to each strain.3 The introduction of this virulent strain from the environment into a hospital setting is concerning since the bacteria could possibly spread to other areas and rapidly gain antibiotic resistance due to its genomic plasticity. Further epidemiological studies are needed to understand the degree of spread of this pathogenic strain among the community in China.
Ethics Approval
This study has been reviewed and approved by the Research Ethics Committee of the First Affiliated Hospital of Zhejiang University (ref#20191002). The patient had given his consent for publication of clinical details, and written informed consent was obtained.
Funding
This work was supported by a research grant from the Natural Science Foundation of Zhejiang Province (LQ18H190001, LQ20H0006) and a grant from the National Natural Science Foundation of China (81971897). The research funding was provided to Dr. Hua Zhou, Dr. Bingquan Zhu and Dr. Yake Yao.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Harding CM, Hennon SW, Feldman MF. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat Rev Microbiol. 2018;16(2):91–102. doi:10.1038/nrmicro.2017.148
2. Lee CR, Lee JH, Park M, et al. Biology of acinetobacter baumannii: pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front Cell Infect Microbiol. 2017;7:55.
3. Antunes LC, Visca P, Towner KJ. Acinetobacter baumannii: evolution of a global pathogen. Pathog Dis. 2014;71(3):292–301. doi:10.1111/2049-632X.12125
4. Wong D, Nielsen TB, Bonomo RA, Pantapalangkoor P, Luna B, Spellberg B. Clinical and pathophysiological overview of acinetobacter infections: a century of challenges. Clin Microbiol Rev. 2017;30(1):409–447. doi:10.1128/CMR.00058-16
5. Chapartegui-Gonzalez I, Lazaro-Diez M, Bravo Z, Navas J, Icardo JM, Ramos-Vivas J. Acinetobacter baumannii maintains its virulence after long-time starvation. PLoS One. 2018;13(8):e0201961. doi:10.1371/journal.pone.0201961
6. Zhou H, Zhang T, Yu D, et al. Genomic analysis of the multidrug-resistant Acinetobacter baumannii strain MDR-ZJ06 widely spread in China. Antimicrob Agents Chemother. 2011;55(10):4506–4512. doi:10.1128/AAC.01134-10
7. Li X, Liu L, Ji J, et al. Tigecycline resistance in Acinetobacter baumannii mediated by frameshift mutation in plsC, encoding 1-acyl-sn-glycerol-3-phosphate acyltransferase. Eur J Clin Microbiol Infect Dis. 2015;34(3):625–631. doi:10.1007/s10096-014-2272-y
8. Hua X, Liu L, Fang Y, et al. Colistin resistance in Acinetobacter baumannii MDR-ZJ06 revealed by a multiomics approach. Front Cell Infect Microbiol. 2017;7:45. doi:10.3389/fcimb.2017.00045
9. Dexter C, Murray GL, Paulsen IT, Peleg AY. Community-acquired Acinetobacter baumannii: clinical characteristics, epidemiology and pathogenesis. Expert Rev Anti Infect Ther. 2015;13(5):567–573. doi:10.1586/14787210.2015.1025055
10. Ong CW, Lye DC, Khoo KL, et al. Severe community-acquired Acinetobacter baumannii pneumonia: an emerging highly lethal infectious disease in the Asia-Pacific. Respirology. 2009;14(8):1200–1205. doi:10.1111/j.1440-1843.2009.01630.x
11. Jacobs AC, Thompson MG, Black CC, et al. AB5075, a highly virulent isolate of Acinetobacter baumannii, as a model strain for the evaluation of pathogenesis and antimicrobial treatments. MBio. 2014;5(3):e01076–14. doi:10.1128/mBio.01076-14
12. Gebhardt MJ, Shuman HA. GigA and GigB are master regulators of antibiotic resistance, stress responses, and virulence in Acinetobacter baumannii. J Bacteriol. 2017;199:10. doi:10.1128/JB.00066-17
13. Wick RR, Judd LM, Gorrie CL, Holt KE. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. 2017;13(6):e1005595. doi:10.1371/journal.pcbi.1005595
14. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
15. Liu B, Zheng D, Jin Q, Chen L, Yang J. VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019. 47(D1):D687–D92. doi:10.1093/nar/gky1080.
16. Visca P, Seifert H, Towner KJ. Acinetobacter infection–an emerging threat to human health. IUBMB Life. 2011;63(12):1048–1054. doi:10.1002/iub.534
17. Seruga Music M, Hrenovic J, Goic-Barisic I, Hunjak B, Skoric D, Ivankovic T. Emission of extensively-drug-resistant Acinetobacter baumannii from hospital settings to the natural environment. J Hosp Infect. 2017;96(4):323–327. doi:10.1016/j.jhin.2017.04.005
18. Chen CT, Wang YC, Kuo SC, et al. Community-acquired bloodstream infections caused by Acinetobacter baumannii: a matched case-control study. J Microbiol Immunol Infect. 2018;51(5):629–635. doi:10.1016/j.jmii.2017.02.004
19. Zeighami H, Valadkhani F, Shapouri R, Samadi E, Haghi F. Virulence characteristics of multidrug resistant biofilm forming Acinetobacter baumannii isolated from intensive care unit patients. BMC Infect Dis. 2019;19(1):629. doi:10.1186/s12879-019-4272-0
20. Bardbari AM, Arabestani MR, Karami M, Keramat F, Alikhani MY, Bagheri KP. Correlation between ability of biofilm formation with their responsible genes and MDR patterns in clinical and environmental Acinetobacter baumannii isolates. Microb Pathog. 2017;108:122–128. doi:10.1016/j.micpath.2017.04.039
21. Koh TH, Sng LH, Wang GC, Hsu LY, Zhao Y. IMP-4 and OXA beta-lactamases in Acinetobacter baumannii from Singapore. J Antimicrob Chemother. 2007;59(4):627–632. doi:10.1093/jac/dkl544
22. Turton JF, Woodford N, Glover J, Yarde S, Kaufmann ME, Pitt TL. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J Clin Microbiol. 2006;44(8):2974–2976. doi:10.1128/JCM.01021-06
23. Dortet L, Bonnin RA, Bernabeu S, et al. First occurrence of OXA-72-producing Acinetobacter baumannii in Serbia. Antimicrob Agents Chemother. 2016;60(10):5724–5730. doi:10.1128/AAC.01016-16
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.