")
Back to Journals » Infection and Drug Resistance » Volume 13
Current Aspects in the Biology, Pathogeny, and Treatment of Candida krusei, a Neglected Fungal Pathogen
Authors Gómez-Gaviria M, Mora-Montes HM
Received 1 February 2020
Accepted for publication 28 May 2020
Published 10 June 2020 Volume 2020:13 Pages 1673—1689
DOI https://doi.org/10.2147/IDR.S247944
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Joachim Wink
Manuela Gómez-Gaviria, Héctor M Mora-Montes
Departamento de Biología, División de Ciencias Naturales y Exactas, Campus Guanajuato, Universidad de Guanajuato, Guanajuato, Gto, México
Correspondence: Héctor M Mora-Montes
Departamento de Biología, División de Ciencias Naturales y Exactas, Campus Guanajuato, Universidad de Guanajuato, Noria Alta s/n, Col. Noria Alta, C.P. 36050, Guanajuato, Gto, México
Tel +52 473-7320006 Ext. 8193
Fax +52 473-7320006 Ext. 8153
Email [email protected]
Abstract: Fungal infections represent a constant and growing menace to human health, because of the emergence of new species as causative agents of diseases and the increment of antifungal drug resistance. Candidiasis is one of the most common fungal infections in humans and is associated with a high mortality rate when the fungi infect deep-seated organs. Candida krusei belongs to the group of candidiasis etiological agents, and although it is not isolated as frequently as other Candida species, the infections caused by this organism are of special relevance in the clinical setting because of its intrinsic resistance to fluconazole. Here, we offer a thorough revision of the current literature dealing with this organism and the caused disease, focusing on its biological aspects, the host-fungus interaction, the diagnosis, and the infection treatment. Of particular relevance, we provide the most recent genomic information, including the gene prediction of some putative virulence factors, like proteases, adhesins, regulators of biofilm formation and dimorphism. Moreover, C. krusei veterinary aspects and the exploration of natural products with anti-C. krusei activity are also included.
Keywords: virulence, candidiasis, host-fungus interplay, antifungal drug, immune sensing
Introduction
Candidiasis is the infection caused by members of the fungal genus Candida, which can be a superficial or a deep-seated disease. The latter is often associated with high morbidity and mortality rates, in particular in hospitalized or immunosuppressed patients. A rate of 3–28 patients out of 1000 intensive care unit admissions in European hospitals develop candidemia;1 and in the United States of America the scenario is similar, as the Transplant-Associated Infection Surveillance Network reported that 3.8% of solid organ transplant recipients developed invasive candidiasis.2 Studies that have enrolled cancer patients admitted into hospitals placed in Europe or the Middle East showed a 36–39% mortality rate after one month of hospitalization,3,4 and these figures suffer minor modifications when the mortality associated to systemic candidiasis among intensive care unit patients is analyzed, which has been calculated in 48%.5 This rate though can scale to figures in the range of 63–75%, depending on the hospital and the patient’s staying ward.6 Thus, there is no doubt that candidemia represents a global healthcare problem and a significant burden on patients.
Candida albicans is the most frequent etiological agent of candidiasis, although other Candida species are also relevant in the clinical setting, causing about 35–65% of candidemia cases.7,8 These include Candida tropicalis, Candida parapsilosis, Candida glabrata, Candida guilliermondii, Candida dubliniensis, Candida auris, and Candida krusei.6,9 Collectively, these species are the causative agents of non-albicans candidiasis and infections by C. krusei are characterized because of their high mortality rate (40–58%) and poor response to standard antifungal therapies.10,12
Due to the clinical relevance of this organism and the significant amount of information generated in recent years; here, we provide a literature revision on the C. krusei basic and clinical aspects.
Biological and Fungal Aspects
C. krusei produces cylindrical yeast cells that may have up 25 µm of length (Figure 1). They usually resemble long-grain rice, which contrasts with the spheric or ovoid shape of other Candida species.13 Like C. albicans, C. krusei shows thermodimorphism, producing hyphae when growing at 37°C and blastoconidia and pseudohyphae when incubated at lower temperatures (Figure 1).14,15 The colony morphology is the typical one of other Candida species, with no obvious features that could provide a hint about the species: round, creamy, and smooth whitish colonies of about 5–8 mm diameter when grown at 25–28°C in rich culturing media, such as malt yeast extract glucose agar, yeast extract peptone glucose agar, or Sabouraud agar (Figure 1). Even though colony morphology switching has been reported,13 no systematic attempt to classify the morphological variations has been reported, like those in C. parapsilosis, C. tropicalis, and C. albicans.16–18 One interesting contrast with other medically relevant Candida species is the presence of sexual reproduction in C. krusei, being Issatchenkia orientalis the teleomorph.13
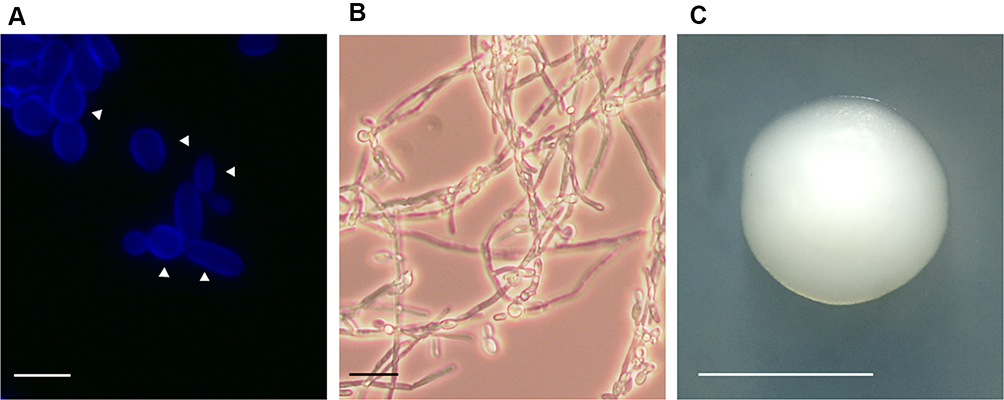 |
Figure 1 Candida krusei cell and colony morphology. (A) Yeast cells were grown in YPD broth until reach the exponential phase and then stained with calcofluor white, to label chitin. Scale bar = 10 µm. The arrowheads indicate the mother cells. (B) Cell filamentation was stimulated in RPMI medium incubated at 37°C. Scale bar = 20 µm. (C) A C. krusei colony grown on a YPD plate. Scale bar = 5.0 mm. Images from panels A and B were taken with a Zeiss Axioscope-40 microscope and an Axiocam MRc camera. |
Like other fungal cells, a cell wall, intracellular vesicles, endoplasmic reticulum, mitochondria, ribosomes, and intracellular glycogen-like granules have been described when cells are inspected under transmission electron microscopy.19 Importantly, the microscopical examination indicates that these are mononuclear cells.19 Thus far, the study of the C. krusei organelles has not been reported in detail, with the exception of the cell wall. This bias in the study of C. krusei components is likely to be related to the relevance of this structure during the interaction with the host and because it is a target of some antifungal drugs, as revised in the following sections. The early study of the C. krusei cell wall by transmission electron microscopy showed the presence of three major layers: the outermost is an electron-dense layer that includes flocculent material surrounding the cell, followed by an electron-transparent layer in the middle with the appearance to be composed of fluffy material and scatter granules, and an innermost electron-dense layer closer to the plasma membrane.19 Recently, our group characterized the basic components of the C. krusei cell wall and found that contains the same polysaccharides found in the C. albicans wall: chitin, β-glucan, and mannans.20 Even though both species have similar levels of cell wall β-glucan, the chitin content is a 4.1-fold higher in C. krusei than in C. albicans, and mannan is 34% less abundant in C. krusei when compared to the C. albicans mannan content.20 In agreement with this observation, the cell wall protein content, and O-linked and N-linked mannans decorating the C. krusei wall glycoproteins were lower than those found in C. albicans.20 The structural study of the C. krusei N-linked mannans indicated that the outer chain is short and lightly branched with α-1,2-mannose units,21 which supports our observations and contrasts with the structure of the C. albicans N-linked mannans, where the outer chain is highly branched with α-1,2-mannose units and capped with either α-1,3-mannose or β-1,2-mannose residues.22 In regard to the C. krusei O-linked mannans, these are oligosaccharides composed of α-1,2-mannose units that can contain from two to four sugar residues,23 which again contrasts with the O-linked mannans found on the C. albicans surface, that may contain up to seven α-1,2-mannose residues.24 Like other Candida species, the C. krusei mannans are modified with mannose residues bound via phosphodiester links, named phosphomannan, although the content of this is about the half of the phosphomannan found in the C. albicans cell wall.20
Like in other Candida species, the C. krusei structural polysaccharides chitin and β-1,3-glucan are localized underneath other cell wall components, and this impairs the proper sensing of these polysaccharides by the host immunity.20
Thus far, only one report dealing with the C. krusei cell wall proteome has been reported, but this was performed with walls from cells growing in the presence of oxidative stressors.25 Interestingly, only moonlighting proteins were identified, which could be a result of contaminants from intracellular compartments, since cells were disrupted with an ultrasonic homogenizer.25 Nonetheless, the presence and abundance of canonical cell wall proteins found in other Candida species remain to be established.
The metabolism of this fungal species is another aspect poorly studied to date. This yeast cell is capable of using exclusively glucose as carbon source,13 which is a trait exploited in its identification by zymograms and chromogenic culturing media.26 This has also been taken in advantage to produce and accumulate glycerol in fermentative processes with potential industrial applications,27 and to prepare traditional meals and alcoholic beverages used by some African communities.28,29 The fermentation process involving C. krusei is positively affected by the presence of lactic acid bacteria, which promote tolerance to short-term changes in the extracellular pH.30 Interestingly, and contrary to this restricted carbohydrate assimilation, C. krusei has been isolated from the decaying wood from Ficus religiosa; suggesting this could be an environmental niche of this fungal species.31 Even though arabinitol is produced by many yeast-like cells and the presence of this metabolite has been reported in serum from patients with invasive candidiasis, C. krusei is incapable of producing this five-carbon polyol.32
In regard to the C. krusei genome, it has been reported this species does not belong to the CUG clade of the Candida genus, it is a diploid and heterozygous organism, with the genetic information distributed in five chromosomes.33–35 The C. krusei genome sequence of a clinical isolate (strain 81-B-5) showed that the nuclear genome sequence size is 10.9 Mbp, the mitochondrial genome contains 51.3 kbp, the single nucleotide polymorphism rate was calculated in 1/340 bases, being higher than that reported for C. albicans isolates.36 Moreover, it was reported a GC content of 38.42%, a 2.15% of repeat content with no significant similarity to the repeat sequences found in C. albicans, and a total of 4949 protein-encoding genes.33,35 The number of intron-containing genes in the coding regions was calculated in 205.35 Different from other Candida species where high variability in the mating loci has been reported,36 these subtelomeric loci are complete in C. krusei.33 A recent study reported the sequencing of 32 strains of C. krusei, Pichia kudriavzevii, Issatchenkia orientalis, and Candida glycerinogenes and found that they are the same species with more of 99% identical DNA sequences.35 Since the analysis of single nucleotide polymorphisms could not segregate between clinical and environmental strains, it was suggested that infections by C. krusei are acquired from the environment.35 Finally, the analysis of the genome sequences supported the re-classification of these organisms in the Pichia genus, being a distant relative of the Candida species.35
Recent Understanding of the Host-Pathogen Interaction
Virulence Factors
Since C. krusei and C. albicans belong to the same taxonomical genus, it has been assumed they share biological traits that help them to interact with the host, a wrong rationale that applies not only to C. krusei but other medically relevant non-albicans Candida species.20,37-39 Therefore, the study of C. krusei virulence factors is a research area with limited information, if compared with the vast amount of reports dealing with C. albicans virulence. In this section, we will provide the most relevant information about C. krusei virulence factors and a genomic comparison to predict putative orthologs of well-known factors already described in C. albicans.
Virulence is classically determined by the ability to damage cells, tissues, organs, or a whole organism, and in mycology, both the in vitro and in vivo systems are thoroughly used. In the murine model of systemic candidiasis, C. krusei was incapable of killing both female or male mice, contrasting with the high mortality rate associated with C. albicans.40,41 When the colony-forming units of these organisms were analyzed in infected spleen, liver, kidneys, and lungs, a gradual reduction in the C. krusei burden was observed during the observation period, with a fungal clearance at day 21 post-infection, contrasting again with the fungal loads in C. albicans-infected organs, which were constants or slightly reduced during the same observation period.40 Therefore, C. krusei displays lower virulence than C. albicans in the murine model of systemic candidiasis. This in vivo system is regarded as the gold standard to assess the virulence of Candida species and isolates, but in recent years, logistical issues to include large numbers of animals per experimental condition and the increased ethical concerns about the use of these animals in basic research have limited their inclusion in the experimental design and stimulated the search for alternative models to study fungal virulence. Caenorhabditis elegans is an invertebrate model that has been used as an alternative for studying the Candida species virulence.42 Upon administration of fungal cells by feeding, C. krusei and C. albicans showed similar ability to kill C. elegans and were ranked as the most lethal species in this experimental setting.42 Like C. albicans, C. krusei was capable of producing aspartyl proteinases, phospholipase, hemolysins, and to develop biofilms, providing a possible explanation to the lethal behavior in the C. elegans system.42 Moreover, this invertebrate model has been useful in establishing the effect of the antibacterial drugs cefepime, imipenem, meropenem, and vancomycin on Candida spp. virulence. C. albicans, C. parapsilosis, C. krusei, and C. tropicalis incremented the proteolytic activity and killing of C. elegans upon incubation with these drugs, whereas amoxicillin potentiated the virulence of C. krusei and C. tropicalis.43 The wax moth Galleria mellonella has been proved as a good model to study infections caused by C. krusei. Upon injection into the hemocele, fungal cells decreased hemocyte density, induced melanization and animal dead.44 The virulence in this host is similar to that observed in the murine model, with C. krusei showing low to moderate ability to kill G. mellonella (median survival of larvae was 7 days), which contrast with the high mortality associated with C. albicans infection (median survival of larvae was 2 days)45 In addition, these larvae have helped to propose that Lactobacillus paracasei, Lactobacillus fermentum, and Lactobacillus rhamnosus, acid bacteria used as probiotics, have a prophylactic effect on the larvae, increasing the animal survival upon administration of a C. krusei lethal dose.46 Drosophila melanogaster is another invertebrate model that has been used to evaluate the C. krusei virulence. Adult flies with mutations in the toll signaling pathway were highly susceptible to infection with either C. albicans or C. krusei, demonstrating this model is useful for virulence assessment.47 Moreover, these data strongly suggest that C. krusei lethality depends on the immunological status of the host, as in immunocompetent animals this Candida species was not capable of killing the host.40,41
Cell and tissue adhesion are part of the early stage of the Candida-host interaction and will lead to the establishment of both commensalism and pathogenesis. Adhesion is paramount to establish colonization and tissue invasion in the oral epithelium, as this tissue is in constant contact with saliva, which cleans the epithelial surface. C. krusei binds to human buccal epithelial cells but not as efficient as C. albicans and C. tropicalis.48 Accordingly, exposure of the epithelial cells to the minimum inhibitory concentration of nystatin affected the Candida species adhesive properties, with C. krusei showing a 64% reduction of adhesion to epithelial cells, a value higher to that found in C. albicans (54%).48 Similar to epithelial cells, endothelial cells are also a surface where C. krusei can adhere, but not as efficiently as C. albicans.49 In agreement with these observations, C. krusei showed 11-fold lower colonization potential of the rat oral surface than C. albicans.50 However, C. krusei adheres in great numbers to acrylic surfaces.50 Interestingly, when the C. krusei phenotypical switching was induced with phloxine B, a 30-fold increment in adhesion to saliva-coated glass surface was observed.51 The Eap1, Iff4, Mp65, Phr1, Int1, Ecm33, and ALS gene family members are the major C. albicans adhesins.52–58 The putative functional orthologs of the genes encoding for Phr1 and Int1 were identified within the C. krusei genome but no those encoding for Eap1, Iff4, Ecm3 or any of the Als family members (Table 1). Interestingly, three putative orthologs of C. albicans Mp65 were identified (Table 1).
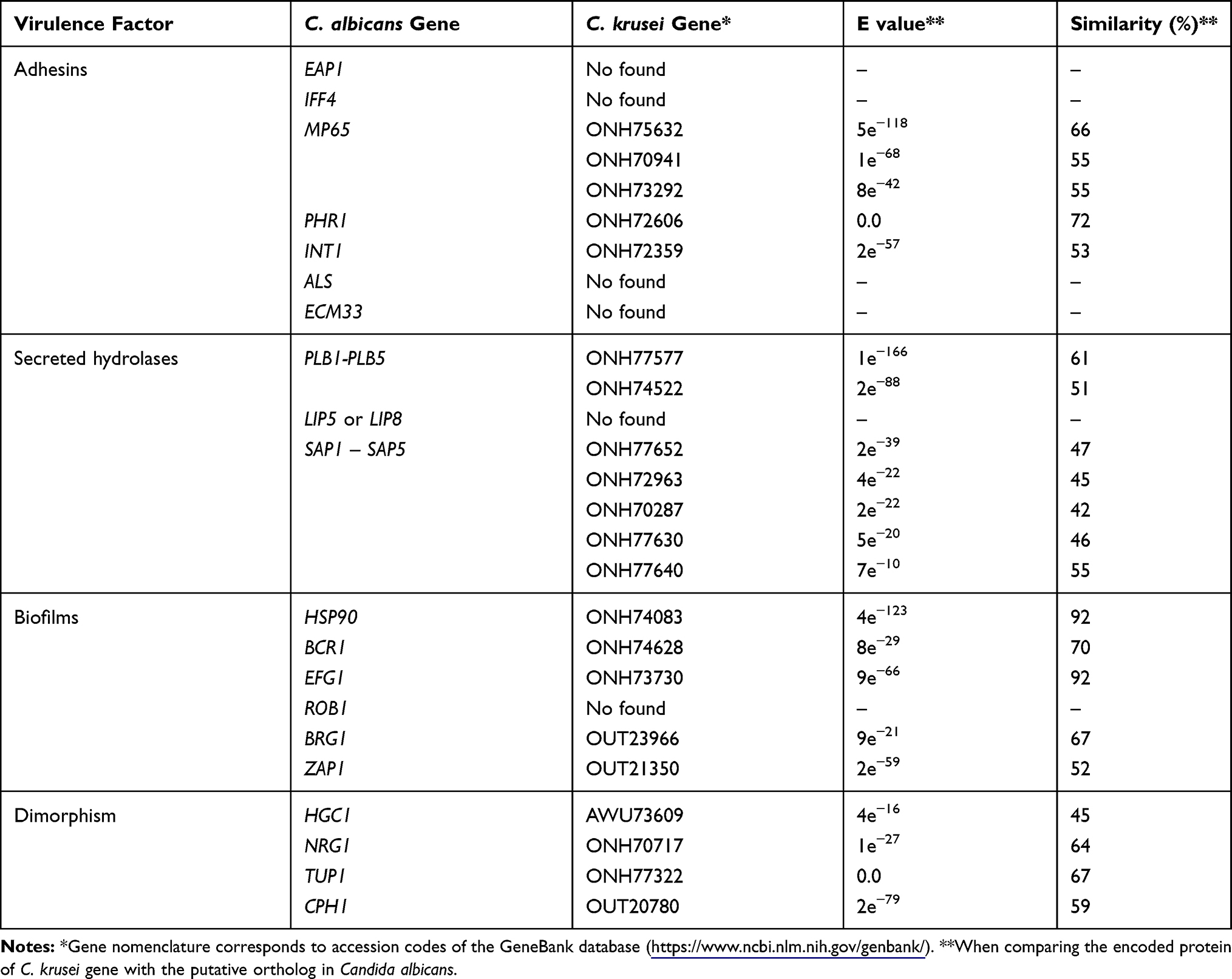 |
Table 1 Prediction of Some Virulence Factors in Candida krusei |
The cell wall hydrophobicity is an important aspect of interaction with the host components. A study that involved 20 C. krusei isolates showed these had higher wall hydrophobicity than C. albicans cells, and there was a correlation between hydrophobicity and adhesion to HeLa cells, but not to acrylic surfaces.59
Several studies have demonstrated that C. krusei, like other Candida species, is capable of secreting hydrolytic enzymes that could degrade host macromolecules, contributing to nutrient acquisition, to degrade immune effectors, or in the dissemination within the host tissues. In two independent studies, using clinical isolates, C. krusei did not show phospholipase activity, contrasting with C. albicans strains that had a strong presence of this enzyme activity.60 However, other studies have shown that this enzyme activity is found in C. krusei.61–63 Whether this discrepancy is due to different methodologies used to measure phospholipase activity or reflects the phenotypical plasticity of the species remains to be addressed. Nonetheless, the C. krusei genome contains two putative orthologs of the PLB gene family that encodes for the major C. albicans secreted phospholipase activity (Table 1).64,65 Interestingly though, no putative orthologs of the genes encoding for secreted lipases belonging to the C. albicans LIP gene family66 were found within the C. krusei genome (Table 1).
Other hydrolytic activities like proteinase, hemolytic factors, and DNase have been reported in C. krusei.60,67,68 A study that included clinical isolates from Turkish patients diagnosed with candidiasis, found that about half of the C. krusei isolates formed biofilms, 22% showed coagulase activity and all the isolates were capable of hemolyzing red blood cells.69 In C. albicans, most of the secreted proteolytic activity is associated with members of the SAP gene family.70,71 A search for putative orthologs of members of this gene family in C. krusei identified SAP1-SAP5, but not SAP6-SAP10 (Table 1). These genes are likely to account for the secreted proteolytic activity reported in C. krusei clinical isolates.
C. krusei is capable of forming biofilms on polyethylene, polyvinylchloride, and glass.67,72 These fungal biofilms are particularly sensitive to fluconazole when generated on polystyrene surfaces.73 This represents a promising observation that remains to be confirmed in vivo. In agreement with the reported ability to generate biofilms, the C. krusei genome contains key genes involved in the establishment of these multicellular communities. The chaperone Hsp90, besides contributing to the establishment of apical growth during the C. albicans dimorphism, is required to neutralize the cellular stress generated during biofilm formation;74 while Efg1, Brg1, Zap1, and Bcr1 are transcriptional factors required for biofilm formation in both in vitro and in vivo conditions.75–77 Putative orthologs for these genes were found within the C. krusei genome (Table 1). However, for the case of Rob1, a transcriptional factor required for biofilm formation in C. albicans,76 no putative ortholog was found within the C. krusei genome (Table 1), suggesting that the regulatory network of biofilm formation in both organisms could share master regulators but their contribution might be species-specific.
As mentioned, C. krusei belongs to the members of the genus capable of forming true hyphae. The C. albicans dimorphism has been associated with tissue invasion and the expression of several virulence factors that are morphology specific.78 The Hgc1 is a hypha-specific G1 cyclin essential for the establishment of the apical growth and is negatively regulated by the transcriptional repressors Nrg1 and Tup1; while Cph1 and Efg1 are transcriptional factors required to sustain hyphal growth.79–81 The C. krusei genome contains putative orthologs of these genes (Table 1), suggesting the central regulatory network that controls dimorphism is similar in both species.
In this regard, it is noteworthy to mention that C. krusei is capable of inhibiting the C. albicans filamentation and biofilm formation.82 Even though there is no formal explanation yet for these observations, the production of signaling molecules by C. krusei to avoid polarized growth in C. albicans, competition for nutrients, adherent surfaces, and space between the biofilms are the main hypotheses currently under study.82
Gliotoxin is an immunosuppressive compound from the fungal metabolism and has been suggested as a fungal virulence factor, including in C. albicans.83 However, a chemical analysis based on highly sensitive HPLC and tandem mass spectrometry of 100 clinical isolates of Candida spp, including C. albicans and C. krusei did not detect intracellular or extracellular gliotoxin production, suggesting this compound does not participate in the pathogenesis of Candida spp.83 In the same line, the C. albicans cytolytic peptide toxin named candidalysin, which is essential for mucosal infection and encoded by ECE1,84 has no putative ortholog within the C. krusei genome.
It is noteworthy to mention that C. krusei has been isolated from bat feces in a Brazilian urban region, and showed the ability to secrete proteases, to form biofilms, and kill laboratory animals, suggesting the animal depositions could be an environmental source for C. krusei infections.85 Similarly, C. krusei has been isolated from droppings of healthy breeding rheas, chickens and hens,86,87 and from the vestibule and vagina of healthy female horses.88
The C. krusei-Immune System Interaction
Similar to other pathogens, the C. krusei recognition and interaction with components of the host immunity is required to establish a response that could protect against the infection. Both the innate and adaptive branches of immunity are essential to control fungal pathogens, including C. albicans and other causative agents of candidiasis.89,90
As part of the humoral factors that belong to the innate immunity, some cells produce antimicrobial peptides that show antifungal properties. The human β-defensin 2 is produced by epithelial cells, while the human neutrophil peptides 1–3 are α-defensins synthesized by circulating white blood cells.91 Both kinds of antimicrobial peptides were stimulated by C. albicans and C. krusei, being the former a stronger inductor than C. krusei cells.91 With no doubt, this study showed the ability of C. krusei to stimulate both local and systemic responses against this pathogen. Similarly, it was reported that C. krusei is 1.4 times more sensitive to lactoferrin, a secreted antimicrobial protein, than C. albicans cells; and this difference has been suggested to be relevant to modulate the fungal oral carriage.92
The peripheral blood mononuclear cells (PBMCs) are a heterogeneous group of immune cells with the ability to produce cytokines upon the interaction between pathogen-associated molecular patterns and their pattern recognition receptors.
Different to the human PBMCs-C. albicans interaction, where low levels of TNFα, IL-6, IL-1β, or IL-10 were stimulated, the immune cells interacting with C. krusei produced higher levels of these four cytokines.20 Both C. albicans and C. krusei heat-killed cells, which expose inner wall components like β-1,3-glucan and chitin at the cell surface, stimulated higher levels of TNFα, IL-6, IL-1β, or IL-10, when compared to live cells.20 Interestingly, a difference between C. krusei and C. albicans was observed when O-linked mannans were removed from the cell wall: C. albicans recognition by PBMCs was not affected, indicating this wall component is dispensable for cytokine stimulation; while in C. krusei, cells with no O-linked mannans on the surface were capable of stimulating higher cytokine levels, most likely because of the unmasking of β-1,3-glucan and increased recognition via dectin-1.20 Like in C. albicans, this immune receptor is essential to control C. krusei infections, as dectin-1 knock out mice are more susceptible to C. krusei and showed poor ability to establish a protective anti-C. krusei immunity.93
In contrast with C. albicans cells, C. krusei yeast cells induced lower levels of complement components C3 and factor B, and the granulocyte-macrophage colony-stimulating factor, but a significant amount of IL-12 (p70).94,95 This differential ability to stimulate IL-12 (p70) could be part of the C. albicans strategies to avoid the establishment of an effective type I immune response against this pathogen, a situation likely to occur fo the case of C. krusei.94
Contrary to this observation though, C. albicans is more readily phagocytosed by neutrophils than C. krusei cells (37% vs 9%),96 and more susceptible to the neutrophil-expressed antimicrobial protein S100A12 than C. krusei,97 underscorings that the differences in pathogenicity and control by the innate immune system are difficult to be reduced to the analysis of a handful of biological parameters.
Like other pathogens, C. krusei is capable of interacting with macrophages, but interestingly the outcome is variable. Rat alveolar macrophages phagocytosed C. glabrata and C. albicans in a similar rate, but this was significantly slower for the case of C. krusei, due to reduced attachment.98 This differential recognition was abrogated though when the fungal uptake was performed with opsonized cells.98 For the case of primary human PBMC-derived macrophages, the results are the opposite: C. krusei is more readily phagocytosed than C. albicans, C. auris, C. tropicalis, and C. guilliermondii.20 In mice, both a macrophage-like cell line and primary macrophages are capable of uptaking C. krusei yeast cells, with around 10 to 20% of the immune cells ingesting yeasts after 2 h, without opsonization.99 However, C. krusei was capable of surviving and undergoing filamentation inside the phagocytic cells, induced defects in the phagolysosome maturation, yeast transfer between infected macrophages, macrophage fusion, and death of the immune cells.99 These data clearly show that the origin of the immune cells has to be taken in to account before drawing general conclusions.
Pyroptosis, an inflammasome-mediated macrophage death process, is activated upon interaction with C. albicans cells.100 This caspase-1-, ASC-, and NLRP3-dependent pathway is triggered in lower levels by C. krusei cells and does not restrict the fungal replication.100
The interaction between C. krusei with dendritic cells has particular outcomes too. The C. krusei mannan but not the cell wall component isolated from C. albicans, C. tropicalis or C. glabrata induced strong cytokine production by these immune cells and led to apoptosis.101 These effects on dendritic cells were mediated by TLR2 and activation of a MyD88-dependent pathway, which controlled the production of the polarizing cytokines IL-12 and IL-6, and thus the Th1/Th17 switching.101
Interestingly, human PBMCs tend to proliferate in the presence of either voriconazole or caspofungin and produce increased levels of IL-2, IFN-γ, and IL-6 when stimulated with either C. albicans or C. krusei, with no effect on the stimulation of TGF-β and IL-10.102 These data suggest that antifungal therapy has a positive immunomodulatory effect on human PBMCs, an observation that should be further explored and taken into account during the treatment of candidiasis and other fungal infections.
Another promising study on new immunomodulatory approaches for the treatment of candidiasis caused by C. krusei involves chromogranin A, a mammalian-expressed soluble protein of the adrenal medullary chromaffin granules and neurons. Chromogranin A N-46, a 46 amino acid portion of the chromogranin A N terminal has shown to have antifungal properties.103 In line with this observation, treatment with this peptide (60 mg/kg/day) had positive effects on mice infected with C. krusei. Treated animals showed increments in the body weight and survival, along with higher counts of circulating monocytes, lymphocytes, and neutrophils.103
Candidiasis Caused by C. krusei
The list of the etiological agents of candidiasis is vast and new species have been added in recent years; however, most of the cases are caused only by five species, named C. albicans, C. glabrata, C. tropicalis, C. parapsilosis, and C. krusei.104 Even though C. albicans is the most common cause of candidemia worldwide, infections caused by C. krusei are an emergent public health threat. Data from the ARTEMIS DISK registry gathered from 1997 to 2007 indicated that the frequency of C. krusei-associated infections was stable, ranging from 1.7 to 3.2%.104
A report from a tertiary care academic hospital in Montreal, which included 190 cases of candidemia, found that C. krusei was the causative agent in 7% of the patients, but this increased up to 13% in the period of 2003–2006. 105 The most likely explanation for this observation in a specific period of time was the systematic prophylaxis with fluconazole in risk groups to develop candidemia.105 Similarly, a Cancer Center of Texas, USA, and a tertiary health care center localized in Haryana, India reported that 8% and 9% of candidemia cases were associated with C. krusei, respectively, but in these cases, the studies were conducted in a pediatric population.106,107 Another study from the USA, but in this case performed in Ihowa, reported that 3.4% of candidemia cases were due to infection with C. krusei, and these showed the lowest 90-day survival rate.108 A study that enrolled 26 short-stay university hospitals in the Paris area reported that the candidemia incidence caused by C. krusei in patients not admitted to ICU was 5.2% in patients with hematological diseases, 3.7% in patients with an oncological condition, and 1.2% in patients with no malignancy diagnosed; while in patients in ICU, the figures were 5.1%, 4.4%, and 2.3% for patients with a hematological disease, oncological conditions, and no malignancy, respectively. 109 Similarly, studies conducted in the Republic of Korea, Australia, Spain, the USA, India, Taiwan, Saudi Arabia, and Portugal reported that 2.6%, 4%, 6%, 5%. 3.3%, 4%, 6%, and 5% of candidemia cases were caused by C. krusei, respectively, with 25% of the Portuguese isolates resistant to posaconazole.110–118 Contrary to these figures though, a study based on a tertiary care hospital in North China found that only 0.9% of the candidemia cases were caused by C. krusei.110–118 Similar to this Chinese study, reports from two Greek, one Brazilian, one Swiss consortium of hospitals, and one Mexican tertiary hospital found that 1.8%, 0.2%, 1%, 2%, and 2.2% of candidemia cases were caused by C. krusei, respectively,119–123 and a study conducted in neonates admitted to ICU, participating in the National Nosocomial Infection Surveillance system from 1995 to 2004 in the USA reported only 0.15% of candidemia cases associated to C. krusei.124
As an outlier report, a study carried out in the University of Texas M. D. Anderson Cancer Center with clinical records of patients admitted from 1993 to 2003 found that C. krusei was the causative agent of 24% and 2% candidemia cases in patients with hematological malignancies and solid tumors, respectively.125 The authors of this study proposed that this prevalence in patients with hematological conditions is due to the prevalent use of fluconazole as a prophylactic antifungal agent, especially in patients with hematological malignancies and recipients of bone marrow transplantation.125 The figures above reported were similar in the period of 2001 to 2007 in the same cancer center, where 17% of candidemia cases were caused by C. krusei.126
The risk factors for fungemia due to C. krusei include the recent surgery report (< 30 days), artificial implants, splenectomy, neutropenia, the presence of oncological conditions such as solid tumors, acute leukemia, or lymphoma as an underlying disease; stem cell transplantation, preexposure to fluconazole, echinocandins or antibacterial agents, specifically vancomycin or piperacillin-tazobactam.104,109,110,125,127,128 At first glance, it is difficult to relate the use of antibacterial agents with the risk to acquire an infection caused by C. krusei or other Candida species. It has been proposed that vancomycin can alter the ecology of the normal skin microbiota, promoting colonization by Candida species and thus increasing the potential to develop a systemic infection; while anti-anaerobic antibacterial agents such piperacillin-tazobactam, may promote overpopulation of yeast species and colonization of the gastrointestinal tract.128 In neonatal patients, among the risk factors associated with C. krusei fungemia are parenteral nutrition, recent fluconazole exposure, use of broad-spectrum antimicrobials, and the presence of a percutaneous inserted central catheter.129,130
Besides the systemic disease, C. krusei is also associated with superficial infections. This organism can cause bronchopneumonia and vulvovaginal candidiasis but is a rare etiological agent in the latter, being isolated only in 0.1% of cases and has a good response to nystatin.131,132 C. krusei has been also found infecting the tonsils, where only surgical removal of the organ offered a permanent cure, causing septic arthritis, ulcers, urinary tract infections, and vasculitis133–136
In veterinary, this organism can also cause infections and deteriorate the health conditions in animals. C. krusei was reported as the causative agent of bovine bronchopneumonia and mastitis in Japan, China, Turkey, Algeria, Canada, Polony, and the United Kingdon;137–143 while in cats could be responsible for the failure of long-term gastrostomy tubes.144 For the case of bovine mastitis, it has been suggested that wheat silage, rather than unappropriated milking is the source of the C. krusei cells affecting the udder.145 Despite it is part of the normal microbiota of birds, C. krusei has been associated with gastrointestinal diseases in white-crowned parrots (Pionus senilis),146 and acute necrotizing ventriculitis in Eclectus parrot (Eclectus roratus). 147
C. krusei Identification
Since C. krusei belongs to a fungal genus that contributes with several species as etiological agents of candidiasis, the methods for identification of C. krusei have been originally developed to discriminate C. albicans from other species. Nonetheless, the following strategies have been refined to identify other Candida species, including C. krusei.
Biochemical reactions are the most common strategies for speciation of Candida isolates, and these have been taken into account to develop chromogenic media that easily differentiate Candida species, depending on the color and morphology of the colonies growing on the plates. C. krusei generates purple fuzzy, large rough colonies with flat pale edges when grown on HiCrome Candida; while the color change to pink fuzzy when cultured on CHROMagar Candida (CHROMagar) or CHROMagar-Pal’s plates.148 In Brilliance™ Candida Agar (formerly Oxoid Chromogenic Candida Agar, OCCA) this species grows like dry, irregular pink-brown colonies.149 Even though this colony color could be informative for species identification, it could be mistaken with the one generated by other species that develop pinkish colonies, such as C. parapsilosis, Candida kefyr, and Candida haemulonii.148,149 Another medium for quick detection of Candida species is CandiSelect™ 4 (BioRad), where C. krusei generates large turquoise-blue colonies with a characteristically rough morphotype, a dry appearance, and an irregular outline. However, C. tropicalis and C. glabrata also grow like smooth, turquoise colonies, making the species identification troublesome.150 Due to these limitations, these media are used for preliminary species identification, and additional phenotypic or genotypic tests should be included for the proper identification of C. krusei and other Candida species. The API ID32C method is currently the gold standard for phenotypic characterization of these organisms, but another alternative is Micronaut-Candida (Bornheim), a microplate-based system that contains 21 biochemical reactions, and 14 carbohydrate assimilation tests (melibiose, D-xylose, L-rhamnose, gentibiose, D-glucose, inositol, cellobiose, saccharose, trehalose, galactose, maltose, lactose, raffinose, and a control reaction), and urease test with its control. Results are generated in 24 h and interpretation assisted by the Micronaut software.151 This strategy proved to be as good as the API ID32C method for the C. krusei identification.151 The Vitek 2 system (bioMérieux) is an automated alternative for C. krusei identification based also on biochemical reactions and has 100% specificity to identify this species.152
Another alternative for Candida identification based on phenotypic traits is the analysis of volatiles using offline gas chromatography and mass spectrometry. The p-xylene, 2-octanone, 2-heptanone, and n-butyl acetate are signature volatiles of the C. krusei presence in in vitro analyses.153
Among the molecular strategies, PCR is one of the techniques thoroughly analyzed and applied for the identification of C. krusei and other Candida species. It was reported that a single primer pair aiming to amplify a fragment of L1A1 gene, which encodes for a cytochrome P-450 lanosterol-14α-demethylase, was capable of detecting fungal DNA in clinical specimens with a sensitivity of 200 fg of DNA but the amplicon size was not useful for Candida species discrimination, as this was in the range of 336 to 350 bp for all the analyzed species.154 However, combined with restriction enzyme analysis using HincII, NsiI, and Sau3A it was generated a species-specific pattern of restriction fragments.154 Similarly, the amplification of the gene coding for the small ribosomal subunit 18S-rRNA and restriction with AluI, BanI, BbsI, DraII, Eco147I, and NheI generated a species-specific profile able to discriminate C. krusei from other Candida species.155 The PCR-RFLP method amplifying the ITS1-5.8S-ITS2 rDNA region followed by restriction with MspI was reported as another alternative to identify this organism.156
Alternatively, a PCR method using a primer pair that amplifies the polymorphic species-specific repetitive sequence C. krusei repeated sequence 1 (CKRS-1) of the non-transcribed intergenic regions of rRNA genes showed 100% specificity and a sensitivity to detect 10 to 100 fg of purified DNA.157 Another alternative for C. krusei identification by PCR is the amplification of part of the gene encoding for topoisomerase II. A nested PCR reaction using a combination of degenerate and specific primers was reported to amplify a characteristic 227 bp DNA fragment from the C. krusei genome, with 100% specificity and a sensitivity of 40 fg of genomic DNA.158
The real-time PCR has been also adapted for C. krusei identification. The amplification of the 5.8 rRNA gene demonstrated that was possible to obtain positive reactions with a detection limit of 10 CFU/mL blood, with 100% specificity.159
More recently, it has been developed the CanTub-simplex PCR based on the amplification of the gene encoding for β-tubulin in a real-time platform, where the amplicons melting temperature is species-specific.160 Following the same rationale, amplification of the internally transcribed spacer region 2 and analysis of melting peaks and curves, allowed the discrimination of C. krusei from the other 15 Candida species included in the study.161
The current molecular alternatives developed for C. krusei identification also include microarrays. Based on the nucleotide sequences of the internal transcribed spacer regions (ITS1 and ITS2) of the rRNA gene a chip to identify 32 fungal pathogens was recently reported.162 Even though the results of the microarray and the automated system Vitek 2 were concordant in 96.7% of cases for all the pathogens tested, for the case of C. krusei the specificity was of 100%.162
The multianalyte profiling system has also been adapted for the identification of Candida species. This consists of mixed polystyrene beads covalently linked to specific DNA probes for C. albicans, C. glabrata, C. tropicalis, C. parapsilosis, C. krusei, and C. dubliniensis, which are incubated with amplicons containing the ITS2 region of Candida species rRNA gene previously amplified with universal fungal primers. The beads conjugated with the corresponding amplicons are analyzed in the multianalyte profiling system flow cytometer that measures the fluorescence produced by the different pairs of amplicons and beads.163 This approach was 100% specific and showed a sensitivity limit of 0.5 pg of DNA.163
The strategies for C. krusei identification also include immunological tests, although these are not as specific as the molecular methods. The Krusei color test (Fumouze) is a latex beads agglutination assay performed with red particles coated with a monoclonal antibody that specifically reacts with a C. krusei antigen found on the cell surface. Although all the C. krusei strains used in the study agglutinated the latex beads, false-positive reactions were observed with C. famata, C. glabrata, C. guilliermondii, C. kefyr, C. parapsilosis, and C. tropicalis.164
Therapy
The treatment of Candida infections includes the use of several kinds of family compounds, named polyenes, azoles, echinocandins, nucleoside analogs, and allylamines. Fluconazole is one of the most common antifungal drugs used for empirical treatment of candidiasis; however, C. krusei is a species intrinsically resistant to this drug, with more than 95% of clinical and veterinary isolates being fluconazole-resistant.141,165 The mechanisms behind this observation are not fully understood yet, but the flux pump activity of the ATP-binding cassette transporter Abc1 and reduced fluconazole affinity to Erg11 have been associated with this phenotypic trait.165,166 It has been also proposed that both proteins could be part of the resistance mechanisms observed in some itraconazole-resistant strains.165 The in vitro acquisition of resistance to voriconazole has been reported after exposing daily C. krusei to 1 µg/mL of the drug. In these cells, drug resistance was associated with increased expression of the ABC1 gene and point mutations within ERG11.167
Several studies conducted with clinical samples have shown that most of the C. krusei strains are susceptible to voriconazole, itraconazole, posaconazole, anidulafungin, micafungin, 5-flucytosine, and amphotericin B; but intermediate resistance to caspofungin has been reported in some isolates.168–170 Although C. krusei is a rare etiological agent of vaginitis, the use of local clotrimazole, ciclopirox olamine, terconazole, and boric acid is recommended.171,172
In veterinary infections though, isolates resistant to fluorocytosine, itraconazole, ketoconazole, and amphotericin B have been reported.141 The pharmacological alternatives to treat candidiasis caused by C. krusei in animals include sulphamethoxypyridazine in cases of bovine mastitis.173
There is a vast amount of studies addressing the search and design of compounds with antifungal activity, as well as the use of herbal derivatives with anti-Candida activity, with the potential of being explored as new alternatives to control candidiasis. Among the most relevant new alternatives are VT-1161 and VT-1129, a new generation of CYP51 inhibitors, a lanosterol 14-α-demethylase that belongs to the cytochrome P450 family and has a role in ergosterol biosynthesis, which showed the inhibition of C. krusei growth at concentrations of ≤ 2 μg/mL after 24 h of incubation.174 Another alternative that is currently under investigation is the use of nanoparticles to deliver antifungal drugs into the fungal cells. It has been recently demonstrated that lipid core nanocapsules containing fluconazole were capable of reducing the effective dose of this antifungal drug and reverted the resistance to fluconazole observed in several C. krusei strains.175 The use of palmatine, in combination with either fluconazole or itraconazole, has shown antifungal synergism, in a mechanism that inhibits the efflux pumps, with the consequent increment in the intracellular azole content.176
A trypsin inhibitor from Tecoma stans (yellow elder) leaves has been isolated, characterized, and shown to have anti-Candida activity.177 The minimal inhibitory concentration for this compound against C. krusei cells was 100 µg/mL, whereas the minimal fungicidal concentration was 200 µg/mL. ATP depletion and lipid peroxidation are thought to be the mechanisms behind its antifungal activity. In addition, it showed no cytotoxicity against human PBMCs,177 being a promising candidate to move forward in the search for new antifungal compounds to treat candidiasis. Flavonoid and tannic fractions from Psidium guajava L. contain high levels of phenolic compounds and showed anti-C. krusei activity that synergizes with fluconazole and affects the morphological transition.178 Similarly, flavonoids from Plinia cauliflora leaves, which mainly contain glycosylated quercetin and myricetin showed inhibitory activity against C. krusei (minimal inhibitory concentration of 19 µg/mL) and low cytotoxicity effect on human cells.179 The water-insoluble fraction from Uncaria tomentosa (cat’s claw) showed a synergistic effect with either terbinafine or fluconazole in a mechanism that involves the action of the plant proanthocyanidins on the fungal cell wall.180
These studies show that traditional medicine, mainly based on herbology, could be a source of a new generation of antifungal drugs.
Animals are also a source of molecules with antifungal activity. The 2-lysophosphatidylcholines isolated from deer antler extracts showed fungistatic activity, suppressing the morphological transition in C. albicans, C. krusei, C. guilliermondii, and C. parapsilosis, in a mechanism mediated through the mitogen-activated protein kinase pathway.181
Concluding Remarks
In recent years, there is a significant amount of information gathered about C. krusei biological and clinical aspects, underscoring the relevance of this organism as an emergent species, most likely because of its intrinsic resistance to fluconazole. The C. krusei genomic sequencing has opened new doors for basic research in this organism that could be translated into clinical applications. The genes prediction, along with their organization within the genome, the proteomic, transcriptomic, and metabolomic analyses could unveil species-specific genes related to virulence or drug resistance, information that could be later exploited in the diagnosis or treatment of the infection. The isolation of this organism from vegetal material and animal dropping points out to the environment as the source of candidiasis caused by C. krusei and provides the background to expand this kind of analysis to get a closer panorama of the C. krusei ecological distribution. The further expansion of our current knowledge on the C. krusei-host interaction would discover singularities in this species, which might be exploited for the design of alternative strategies to control the disease caused by this and other Candida species. This is of particular interest because thus far no vaccine is currently available to prevent candidiasis.182 However, new immunotherapeutic approaches, and the ultimate development of a vaccine against Candida species will rely on the deep knowledge of the immunity against these organisms.
Even though there are phenotypical and molecular strategies to identify C. krusei available in the clinical setting, faster, cheaper and more accurate alternatives are desirable for the early diagnosis of C. krusei and other Candida species. We provided some examples of natural products that could have antifungal activity, and these efforts should be replicated and look into the mechanisms behind the antifungal effect, as these compounds could be part of a new generation of drugs to treat candidiasis and other fungal infections. Finally, the search for the mechanisms behind the resistance to fluconazole in this species would provide useful information for the design of new treatment alternatives.
Acknowledgments
This work was supported by Consejo Nacional de Ciencia y Tecnología (ref. PDCPN2014-247109, and FC 2015-02-834), Universidad de Guanajuato (ref. CIIC 087/2019), and Red Temática Glicociencia en Salud (CONACYT-México).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Klingspor L, Tortorano AM, Peman J, et al. Invasive Candida infections in surgical patients in intensive care units: a prospective, multicentre survey initiated by the European Confederation of Medical Mycology (ECMM) (2006–2008). Clin Microbiol Infect. 2015;21(1):
2. Andes DR, Safdar N, Baddley JW, et al. The epidemiology and outcomes of invasive Candida infections among organ transplant recipients in the United States: results of the Transplant-Associated Infection Surveillance Network (TRANSNET). Transpl Infect Dis. 2016;18(6):921–931. doi:10.1111/tid.12613
3. Cornely OA. on behalf of the EIDG, Gachot B, et al. Epidemiology and outcome of fungemia in a cancer cohort of the infectious diseases group (IDG) of the european organization for research and treatment of cancer (EORTC 65031). Clin Infect Dis. 2015;61(3):324–331. doi:10.1093/cid/civ293
4. Viscoli C, Girmenia C, Marinus A, et al. Candidemia in cancer patients: a prospective, multicenter surveillance study by the Invasive Fungal Infection Group (IFIG) of the European Organization for Research and Treatment of Cancer (EORTC). Clin Infect Dis. 1999;28(5):1071–1079. doi:10.1086/514731
5. Blot SI, Vandewoude KH, Hoste EA, Colardyn FA. Effects of nosocomial candidemia on outcomes of critically ill patients. Am J Med. 2002;113(6):480–485. doi:10.1016/S0002-9343(02)01248-2
6. Eliakim-Raz N, Babaoff R, Yahav D, Yanai S, Shaked H, Bishara J. Epidemiology, microbiology, clinical characteristics, and outcomes of candidemia in internal medicine wards—a retrospective study. Int J Infect Dis. 2016;52:49–54. doi:10.1016/j.ijid.2016.09.018
7. Lass-Flörl C. The changing face of epidemiology of invasive fungal disease in Europe. Mycoses. 2009;52(3):197–205. doi:10.1111/j.1439-0507.2009.01691.x
8. Tan TY, Tan AL, Tee NWS, Ng LSY, Chee CWJ. The increased role of non-albicans species in candidaemia: results from a 3-year surveillance study. Mycoses. 2010;53(6):515–521. doi:10.1111/j.1439-0507.2009.01746.x
9. Pfaller MA, Boyken L, Hollis RJ, Messer SA, Tendolkar S, Diekema DJ. In vitro activities of anidulafungin against more than 2,500 clinical isolates of Candida spp., including 315 isolates resistant to fluconazole. J Clin Microbiol. 2005;43(11):5425–5427. doi:10.1128/JCM.43.11.5425-5427.2005
10. Pfaller Michael A, Pappas Peter G, Wingard John R. Invasive fungal pathogens: current epidemiological trends. Clin Infect Dis. 2006;43(S1):S3–S14. doi:10.1086/504490
11. Tortorano AM, Kibbler C, Peman J, Bernhardt H, Klingspor L, Grillot R. Candidaemia in Europe: epidemiology and resistance. Int J Antimicrob Agents. 2006;27(5):359–366. doi:10.1016/j.ijantimicag.2006.01.002
12. Wisplinghoff H, Bischoff T, Tallent Sandra M, Seifert H, Wenzel Richard P, Edmond Michael B. Nosocomial bloodstream infections in US hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin Infect Dis. 2004;39(3):309–317. doi:10.1086/421946
13. Samaranayake YH, Samaranayake LP. Candida krusei: biology, epidemiology, pathogenicity and clinical manifestations of an emerging pathogen. J Med Microbiol. 1994;41(5):295–310. doi:10.1099/00222615-41-5-295
14. Samaranayake YH, Wu PC, Samaranayake LP, PL HO. The relative pathogenicity of Candida Krusei and C. albicans in the rat oral mucosa. J Med Microbiol. 1998;47(12):1047–1057. doi:10.1099/00222615-47-12-1047
15. Fleischmann J, Broeckling CD, Lyons S. Candida krusei form mycelia along agar surfaces towards each other and other Candida species. BMC Microbiol. 2017;17(1):60. doi:10.1186/s12866-017-0972-z
16. Toth R, Nosek J, Mora-Montes HM, et al. Candida parapsilosis: from genes to the bedside. Clin Microbiol Rev. 2019;32:2.
17. Zheng Q, Zhang Q, Bing J, Ding X, Huang G. Environmental and genetic regulation of white-opaque switching in. Candida Tropicalis. Mol Microbiol. 2017;106(6):999–1017. doi:10.1111/mmi.13862
18. Soll DR. The role of phenotypic switching in the basic biology and pathogenesis of Candida albicans. J Oral Microbiol. 2014;6(1):22993. doi:10.3402/jom.v3406.22993
19. Joshi KR, Wheeler EE, Gavin JB. The ultrastructure of Candida krusei Candida krusei. Mycopathologia. 1975;56(1):5–8. doi:10.1007/BF00493575
20. Navarro-Arias MJ, Hernandez-Chavez MJ, Garcia-Carnero LC, et al. Differential recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by human innate immune cells. Infect Drug Resist. 2019;12:783–794. doi:10.2147/IDR.S197531
21. Kogan G, Pavliak V, Sandula J, Masler L. Novel structure of the cellular mannan of the pathogenic yeast Candida krusei. Carbohydr Res. 1988;184:171–182. doi:10.1016/0008-6215(88)80015-6
22. Mora-Montes HM, Ponce-Noyola P, Villagómez-Castro JC, Gow NAR, Flores-Carreón A, López-Romero E. Protein glycosylation in Candida. Future Microbiol 2009;4(9):1167–1183. doi:10.2217/fmb.09.88
23. Kuraoka T, Ishiyama A, Oyamada H, Ogawa Y, Kobayashi H. Presence of O-glycosidically linked oligosaccharides in the cell wall mannan of Candida krusei purified with Benanomicin A. FEBS Open Bio. 2018;9(1):129–136. doi:10.1002/2211-5463.12558
24. Diaz-Jimenez DF, Mora-Montes HM, Hernandez-Cervantes A, Luna-Arias JP, Gow NA, Flores-Carreon A. Biochemical characterization of recombinant Candida albicans mannosyltransferases Mnt1, Mnt2 and Mnt5 reveals new functions in O- and N-mannan biosynthesis. Biochem Biophys Res Commun. 2012;419(1):77–82. doi:10.1016/j.bbrc.2012.01.131
25. Serrano-Fujarte I, Lopez-Romero E, Cuellar-Cruz M. Moonlight-like proteins of the cell wall protect sessile cells of Candida from oxidative stress. Microb Pathog. 2016;90:22–33. doi:10.1016/j.micpath.2015.10.001
26. Vecchione A, Florio W, Celandroni F, Barnini S, Lupetti A, Ghelardi E. Comparative evaluation of six chromogenic media for presumptive yeast identification. J Clin Pathol. 2017;70(12):1074–1078. doi:10.1136/jclinpath-2017-204396
27. Liu HJ, Liu DH, Zhong JJ. Novel fermentation strategy for enhancing glycerol production by Candida krusei. Biotechnol Prog. 2003;19(5):1615–1619. doi:10.1021/bp034097f
28. Greppi A, Rantisou K, Padonou W, et al. Yeast dynamics during spontaneous fermentation of mawè and tchoukoutou, two traditional products from Benin. Int J Food Microbiol. 2013;165(2):200–207. doi:10.1016/j.ijfoodmicro.2013.05.004
29. Omemu AM, Oyewole OB, Bankole MO. Significance of yeasts in the fermentation of maize for ogi production. Food Microbiol. 2007;24(6):571–576. doi:10.1016/j.fm.2007.01.006
30. Halm M, Hornbæk T, Arneborg N, Sefa-Dedeh S, Jespersen L. Lactic acid tolerance determined by measurement of intracellular pH of single cells of Candida krusei and Saccharomyces cerevisiae isolated from fermented maize dough. Int J Food Microbiol. 2004;94(1):97–103. doi:10.1016/j.ijfoodmicro.2003.12.019
31. Randhawa HS, Mussa AY, Khan ZU. Decaying wood in tree trunk hollows as a natural substrate for Cryptococcus neoformans and other yeast-like fungi of clinical interest. Mycopathologia. 2001;151(2):63–69. doi:10.1023/A:1010906220888
32. Bernard EM, Christiansen KJ, Tsang SF, Kiehn TE, Armstrong D. Rate of arabinitol production by pathogenic yeast species. J Clin Microbiol. 1981;14(2):189–194. doi:10.1128/JCM.14.2.189-194.1981
33. Cuomo CA, Shea T, Yang B, Rao R, Forche A. Whole genome sequence of the heterozygous clinical isolate Candida krusei 81-B-5. G3 (Bethesda). 2017;7(9):2883–2889.
34. Jacobsen MD, Gow NAR, Maiden MCJ, Shaw DJ, Odds FC. Strain typing and determination of population structure of Candida krusei by multilocus sequence typing. J Clin Microbiol. 2007;45(2):317–323. doi:10.1128/JCM.01549-06
35. Douglass AP, Offei B, Braun-Galleani S, et al. Population genomics shows no distinction between pathogenic Candida krusei and environmental Pichia kudriavzevii: one species, four names. PLoS Pathog. 2018;14(7):e1007138–e1007138. doi:10.1371/journal.ppat.1007138
36. Butler G, Rasmussen MD, Lin MF, et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature. 2009;459(7247):657–662. doi:10.1038/nature08064
37. Estrada-Mata E, Navarro-Arias MJ, Perez-Garcia LA, et al. Members of the Candida parapsilosis complex and Candida albicans are differentially recognized by human peripheral blood mononuclear cells. Front Microbiol. 2015;6:1527.
38. Navarro-Arias MJ, Defosse TA, Dementhon K, et al. Disruption of protein mannosylation affects Candida guilliermondii cell wall, immune sensing, and virulence. Front Microbiol. 2016;7:1951. doi:10.3389/fmicb.2016.01951
39. Perez-Garcia LA, Csonka K, Flores-Carreon A, et al. Role of protein glycosylation in Candida parapsilosis cell wall integrity and host interaction. Front Microbiol. 2016;7:306. doi:10.3389/fmicb.2016.00306
40. Koga-Ito CY, Komiyama EY, de Paiva Martins CA, et al. Experimental systemic virulence of oral Candida dubliniensis isolates in comparison with Candida albicans, Candida tropicalis and Candida krusei. Mycoses. 2011;54(5):e278–e285. doi:10.1111/j.1439-0507.2010.01899.x
41. Anaissie E, Hachem R, Tin-U C, Stephens LC, Bodey GP. Experimental hematogenous candidiasis caused by Candida krusei and Candida albicans: species differences in pathogenicity. Infect Immun. 1993;61(4):1268–1271. doi:10.1128/IAI.61.4.1268-1271.1993
42. Ortega-Riveros M, De-la-Pinta I, Marcos-Arias C, Ezpeleta G, Quindós G, Eraso E. Usefulness of the non-conventional Caenorhabditis elegans model to assess Candida virulence. Mycopathologia. 2017;182(9):785–795. doi:10.1007/s11046-017-0142-8
43. Aguiar Cordeiro R, de Jesus Evangelista AJ, Serpa R, et al. β-lactam antibiotics & vancomycin increase the growth & virulence of Candida spp. Future Microbiol. 2018;13(8):869–875. doi:10.2217/fmb-2018-0019
44. Silva LN, Campos-Silva R, Ramos LS, et al. Virulence of Candida haemulonii complex in Galleria mellonella and efficacy of classical antifungal drugs: a comparative study with other clinically relevant non-albicans Candida species. FEMS Yeast Res. 2018;18(7):foy082. doi:10.1093/femsyr/foy082
45. Marcos-Zambrano LJ, Bordallo-Cardona MÁ, Borghi E, et al. Candida isolates causing candidemia show different degrees of virulence in Galleria mellonella. Med Mycol. 2020;58(1):83–92. doi:10.1093/mmy/myz027
46. Santos RB, Scorzoni L, Namba AM, Rossoni RD, Jorge AOC, Junqueira JC. Lactobacillus species increase the survival of Galleria mellonella infected with Candida albicans and non–albicans Candida clinical isolates. Med Mycol. 2018;57(3):391–394. doi:10.1093/mmy/myy032
47. Chamilos G, Lionakis MS, Lewis RE, et al. Drosophila melanogaster as a facile model for large-scale studies of virulence mechanisms and antifungal drug efficacy in Candida species. J Infect Dis. 2006;193(7):1014–1022. doi:10.1086/500950
48. Ellepola ANB, Panagoda GJ, Samaranayake LP. Adhesion of oral Candida species to human buccal epithelial cells following brief exposure to nystatin. Oral Microbiol Immunol. 1999;14(6):358–363. doi:10.1034/j.1399-302X.1999.140605.x
49. Klotz SA, Drutz DJ, Harrison JL, Huppert M. Adherence and penetration of vascular endothelium by Candida yeasts. Infect Immun. 1983;42(1):374–384. doi:10.1128/IAI.42.1.374-384.1983
50. Samaranayake YH, Wu PC, Samaranayake LP, So M, Yuen KY. Adhesion and colonisation of Candida krusei on host surfaces. J Med Microbiol. 1994;41(4):250–258. doi:10.1099/00222615-41-4-250
51. Arzmi MH, Abdul Razak F, Yusoff Musa M, Wan Harun WHA. Effect of phenotypic switching on the biological properties and susceptibility to chlorhexidine in Candida krusei ATCC 14243. FEMS Yeast Res. 2012;12(3):351–358. doi:10.1111/j.1567-1364.2011.00786.x
52. Li F, Palecek SP. EAP1, a Candida albicans gene involved in binding human epithelial cells. Eukaryot Cell. 2003;2(6):1266–1273. doi:10.1128/EC.2.6.1266-1273.2003
53. Kempf M, Apaire-Marchais V, Saulnier P, et al. Disruption of Candida albicans IFF4 gene involves modifications of the cell electrical surface properties. Colloids Surf B: Biointerfaces. 2007;58(2):250–255. doi:10.1016/j.colsurfb.2007.03.017
54. Sandini S, La Valle R, De Bernardis F, Macrì C, Cassone A. The 65 kDa mannoprotein gene of Candida albicans encodes a putative β-glucanase adhesin required for hyphal morphogenesis and experimental pathogenicity. Cell Microbiol. 2007;9(5):1223–1238. doi:10.1111/j.1462-5822.2006.00862.x
55. Calderon J, Zavrel M, Ragni E, Fonzi WA, Rupp S, Popolo L. PHR1, a pH-regulated gene of Candida albicans encoding a glucan-remodelling enzyme, is required for adhesion and invasion. Microbiology. 2010;156(8):2484–2494. doi:10.1099/mic.0.038000-0
56. Gale CA, Bendel CM, McClellan M, et al. Linkage of adhesion, filamentous growth, and virulence in Candida albicans; to a single gene INT1. Science. 1998;279(5355):1355–1358. doi:10.1126/science.279.5355.1355
57. Martinez-Lopez R, Park H, Myers CL, Gil C, Filler SG. Candida albicans Ecm33p is important for normal cell wall architecture and interactions with host cells. Eukaryot Cell. 2006;5(1):140–147. doi:10.1128/EC.5.1.140-147.2006
58. Hoyer LL. The ALS gene family of Candida albicans. Trends Microbiol. 2001;9(4):176–180. doi:10.1016/S0966-842X(01)01984-9
59. Samaranayake YH, Wu PC, Samaranayake LP, So M. Relationship between the cell surface hydrophobicity and adherence of Candida krusei and Candida albicans to epithelial and denture acrylic surfaces. APMIS. 1995;103(7‐8):707–713. doi:10.1111/j.1699-0463.1995.tb01427.x
60. Riceto É, Menezes R, Penatti MPA, Pedroso R. Enzymatic and hemolytic activity in different Candida species. Rev Iberoam Micol. 2015;32(2):79–82. doi:10.1016/j.riam.2013.11.003
61. Pandey N, Gupta MK, Tilak R. Extracellular hydrolytic enzyme activities of the different Candida spp. isolated from the blood of the intensive care unit-admitted patients. J Lab Physicians. 2018;10(4):392–396. doi:10.4103/JLP.JLP_81_18
62. Furlaneto-Maia L, Specian AF, Bizerra FC, de Oliveira MT, Furlaneto MC. In vitro evaluation of putative virulence attributes of oral isolates of Candida spp. obtained from elderly healthy individuals. Mycopathologia. 2008;166(4):209. doi:10.1007/s11046-008-9139-7
63. Portela MB, Lima de Amorim E, Santos AM, et al. Candida species from oral cavity of HIV-infected children exhibit reduced virulence factors in the HAART era. Microb Pathog. 2017;102:74–81. doi:10.1016/j.micpath.2016.11.020
64. Leidich SD, Ibrahim AS, Fu Y, et al. Cloning and disruption of caPLB1, a phospholipase B gene involved in the pathogenicity of Candida albicans. J Biol Chem. 1998;273(40):26078–26086. doi:10.1074/jbc.273.40.26078
65. Sugiyama Y, Nakashima S, Mirbod F, et al. Molecular cloning of a second phospholipase B gene, caPLB2 from Candida albicans. Medical Mycology. 1999;37(1):61–67.
66. Hube B, Stehr F, Bossenz M, Mazur A, Kretschmar M, Schäfer W. Secreted lipases of Candida albicans: cloning, characterisation and expression analysis of a new gene family with at least ten members. Arch Microbiol. 2000;174(5):362–374. doi:10.1007/s002030000218
67. Noumi E, Snoussi M, Noumi I, Saghrouni F, Aouni M, Valentin E. Phenotypic characterization and adhesive properties of vaginal Candida spp. strains provided by the CHU Farhat Hached (Sousse, Tunisia). Rev Iberoam Micol. 2015;32(3):170–179. doi:10.1016/j.riam.2014.06.006
68. Şeker E. Identification of Candida species isolated from bovine mastitic milk and their in vitro hemolytic activity in western Turkey. Mycopathologia. 2010;169(4):303–308.
69. Yigit N, Aktas E, Dagistan S, Ayyildiz A. Investigating biofilm production, coagulase and hemolytic activity in Candida species isolated from denture stomatitis patients. Eurasian J Med. 2011;43(1):27–32. doi:10.5152/eajm.2011.06
70. Magee BB, Hube B, Wright RJ, Sullivan PJ, Magee PT. The genes encoding the secreted aspartyl proteinases of Candida albicans constitute a family with at least three members. Infect Immun. 1993;61(8):3240–3243. doi:10.1128/IAI.61.8.3240-3243.1993
71. White TC, Miyasaki SH, Agabian N. Three distinct secreted aspartyl proteinases in Candida albicans. J Bacteriol. 1993;175(19):6126–6133. doi:10.1128/JB.175.19.6126-6133.1993
72. Sánchez-Vargas LO, Estrada-Barraza D, Pozos-Guillen AJ, Rivas-Caceres R. Biofilm formation by oral clinical isolates of Candida species. Arch Oral Biol. 2013;58(10):1318–1326. doi:10.1016/j.archoralbio.2013.06.006
73. Salari S, Sadat Seddighi N, Ghasemi Nejad Almani P. Evaluation of biofilm formation ability in different Candida strains and anti-biofilm effects of Fe3O4-NPs compared with fluconazole: an in vitro study. J Mycol Med. 2018;28(1):23–28. doi:10.1016/j.mycmed.2018.02.007
74. Cao -Y-Y, Cao Y-B, Xu Z, et al. cDNA microarray analysis of differential gene expression in Candida albicans biofilm exposed to farnesol. Antimicrob Agents Chemother. 2005;49(2):584–589. doi:10.1128/AAC.49.2.584-589.2005
75. Nobile CJ, Mitchell AP. Regulation of cell-surface genes and biofilm formation by the C. albicans transcription factor Bcr1p. Curr Biol. 2005;15(12):1150–1155. doi:10.1016/j.cub.2005.05.047
76. Nobile CJ, Fox EP, Nett JE, et al. A recently evolved transcriptional network controls biofilm development in Candida albicans. Cell. 2012;148(1–2):126–138.
77. Nobile CJ, Nett JE, Hernday AD, et al. Biofilm matrix regulation by Candida albicans Zap1. PLoS Biol. 2009;7(6):e1000133–e1000133. doi:10.1371/journal.pbio.1000133
78. Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence. 2013;4(2):119–128. doi:10.4161/viru.22913
79. Zheng X, Wang Y, Wang Y. Hgc1, a novel hypha-specific G1 cyclin-related protein regulates Candida albicans hyphal morphogenesis. EMBO J. 2004;23(8):1845–1856. doi:10.1038/sj.emboj.7600195
80. García-Sánchez S, Mavor AL, Russell CL, et al. Global roles of Ssn6 in Tup1- and Nrg1-dependent gene regulation in the fungal pathogen, Candida albicans. Mol Biol Cell. 2005;16(6):2913–2925. doi:10.1091/mbc.e05-01-0071
81. Liu H, Kohler J, Fink G. Suppression of hyphal formation in Candida albicans by mutation of a STE12 homolog. Science. 1994;266(5191):1723–1726. doi:10.1126/science.7992058
82. de Barros PP, Freire F, Rossoni RD, Junqueira JC, Jorge AOC. Candida krusei and Candida glabrata reduce the filamentation of Candida albicans by downregulating expression of HWP1 gene. Folia Microbiol (Praha). 2017;62(4):317–323. doi:10.1007/s12223-017-0500-4
83. Kupfahl C, Ruppert T, Dietz A, Geginat G, Hof H. Candida species fail to produce the immunosuppressive secondary metabolite gliotoxin in vitro. FEMS Yeast Res. 2007;7(6):986–992. doi:10.1111/j.1567-1364.2007.00256.x
84. Moyes DL, Wilson D, Richardson JP, et al. Candidalysin is a fungal peptide toxin critical for mucosal infection. Nature. 2016;532(7597):64–68. doi:10.1038/nature17625
85. Botelho NS, de Paula SB, Panagio LA, Pinge-Filho P, Yamauchi LM, Yamada-Ogatta SF. Candida species isolated from urban bats of Londrina-Paraná, Brazil and their potential virulence. Zoonoses Public Health. 2012;59(1):16–22. doi:10.1111/j.1863-2378.2011.01410.x
86. Subramanya SH, Sharan NK, Baral BP, et al. Diversity, in-vitro virulence traits and antifungal susceptibility pattern of gastrointestinal yeast flora of healthy poultry, Gallus gallus domesticus. BMC Microbiol. 2017;17(1):113. doi:10.1186/s12866-017-1024-4
87. Brilhante RSN, de Alencar LP, Cordeiro RA, et al. Detection of Candida species resistant to azoles in the microbiota of rheas (Rhea americana): possible implications for human and animal health. J Med Microbiol. 2013;62(Pt 6):889–895. doi:10.1099/jmm.0.055566-0
88. Azarvandi A, Khosravi AR, Shokri H, et al. Presence and distribution of yeasts in the reproductive tract in healthy female horses. Equine Vet J. 2017;49(5):614–617. doi:10.1111/evj.12657
89. Martinez-Alvarez JA, Perez-Garcia LA, Flores-Carreon A, Mora-Montes HM. The immune response against Candida spp. and Sporothrix schenckii. Rev Iberoam Micol. 2014;31(1):62–66. doi:10.1016/j.riam.2013.09.015
90. Hernandez-Chavez MJ, Perez-Garcia LA, Nino-Vega GA, Mora-Montes HM. Fungal strategies to evade the host immune recognition. J Fungi (Basel). 2017;3(4):51. doi:10.3390/jof3040051
91. Gácser A, Tiszlavicz Z, Németh T, Seprényi G, Mándi Y. Induction of human defensins by intestinal Caco-2 cells after interactions with opportunistic Candida species. Microbes Infect. 2014;16(1):80–85. doi:10.1016/j.micinf.2013.09.003
92. Samaranayake YH, Samaranayake LP, Wu PC, So M. The antifungal effect of lactoferrin and lysozyme on Candida krusei and Candida albicans. APMIS. 1997;105(7‐12):875–883. doi:10.1111/j.1699-0463.1997.tb05097.x
93. Chen SM, Zou Z, Qiu XR, et al. The critical role of dectin-1 in host controlling systemic Candida krusei infection. Am J Transl Res. 2019;11(2):721–732.
94. Xiong J, Kang K, Liu L, Yoshida Y, Cooper KD, Ghannoum MA. Candida albicans and Candida krusei differentially induce human blood mononuclear cell interleukin-12 and gamma interferon production. Infect Immun. 2000;68(5):2464–2469. doi:10.1128/IAI.68.5.2464-2469.2000
95. H⊘gÅsen AKM, Abrahamsen TG, Gaustad P. Various Candida and Torulopsis species differ in their ability to induce the production of C3, factor B and granulocyte-macrophage colony-stimulating factor (GM-CSF) in human monocyte cultures. J Med Microbiol. 1995;42(4):291–298. doi:10.1099/00222615-42-4-291
96. Richardson MD, Donaldson F. Interaction of Candida krusei with human neutrophils in vitro. J Med Microbiol. 1994;41(6):384–388. doi:10.1099/00222615-41-6-384
97. Cunden LS, Gaillard A, Nolan EM. Calcium ions tune the zinc-sequestering properties and antimicrobial activity of human S100A12. Chem Sci. 2016;7(2):1338–1348. doi:10.1039/C5SC03655K
98. Nessa K, Johansson A, Jarstrand C, Camner P. Alveolar macrophage reaction to Candida species. Lett Appl Microbiol. 1997;25(3):181–185. doi:10.1046/j.1472-765X.1997.00200.x
99. García-Rodas R, González-Camacho F, Rodríguez-Tudela JL, Cuenca-Estrella M, Zaragoza O. The interaction between Candida krusei and murine macrophages results in multiple outcomes, including intracellular survival and escape from killing. Infect Immun. 2011;79(6):2136–2144. doi:10.1128/IAI.00044-11
100. Wellington M, Koselny K, Sutterwala FS, Krysan DJ. Candida albicans triggers NLRP3-mediated pyroptosis in macrophages. Eukaryot Cell. 2014;13(2):329–340. doi:10.1128/EC.00336-13
101. Nguyen TNY, Padungros P, Wongsrisupphakul P, et al. Cell wall mannan of Candida krusei mediates dendritic cell apoptosis and orchestrates Th17 polarization via TLR-2/MyD88-dependent pathway. Sci Rep. 2018;8(1):17123. doi:10.1038/s41598-018-35101-3
102. Fidan I, Yesilyurt E, Kalkanci A, et al. Immunomodulatory effects of voriconazole and caspofungin on human peripheral blood mononuclear cells stimulated by Candida albicans and Candida krusei. Am J Med Sci. 2014;348(3):219–223. doi:10.1097/MAJ.0000000000000236
103. Li R, Zhang L, Zhang H, et al. Protective effect of a novel antifungal peptide derived from human chromogranin a on the immunity of mice infected with Candida krusei. Exp Ther Med. 2017;13(5):2429–2434. doi:10.3892/etm.2017.4290
104. Guinea J. Global trends in the distribution of Candida species causing candidemia. Clin Microbiol Infect. 2014;20:5–10. doi:10.1111/1469-0691.12539
105. Labbé A-C, Pépin J, Patiño C, Castonguay S, Restieri C, Laverdiere M. A single-centre 10-year experience with Candida bloodstream infections. Can J Infect Dis Med Microbiol. 2009;20(2):45–50. doi:10.1155/2009/731070
106. Mullen CA, Abd. El-Baki H, Samir H, Tarrand JJ, Rolston KV. Non-albicans Candida is the most common cause of candidemia in pediatric cancer patients. Support Care Cancer. 2003;11(5):321–325. doi:10.1007/s00520-003-0453-9
107. Chaudhary U, Goel S, Mittal S. Changing trends of Candidemia and antifungal susceptibility pattern in a tertiary health care centre. Infect Disord Drug Targets. 2015;15(3):171–176. doi:10.2174/1871526515666150928115307
108. Pfaller M, Neofytos D, Diekema D, et al. Epidemiology and outcomes of candidemia in 3648 patients: data from the Prospective Antifungal Therapy (PATH Alliance®) registry, 2004–2008. Diagn Microbiol Infect Dis. 2012;74(4):323–331. doi:10.1016/j.diagmicrobio.2012.10.003
109. Lortholary O, Renaudat C, Sitbon K, et al. The risk and clinical outcome of candidemia depending on underlying malignancy. Intensive Care Med. 2017;43(5):652–662. doi:10.1007/s00134-017-4743-y
110. Ding X, Yan D, Sun W, Zeng Z, Su R, Su J. Epidemiology and risk factors for nosocomial Non-Candida albicans candidemia in adult patients at a tertiary care hospital in North China. Med Mycol. 2015;53(7):684–690. doi:10.1093/mmy/myv060
111. Marco F, Danés C, Almela M, et al. Trends in frequency and in vitro susceptibilities to antifungal agents, including voriconazole and anidulafungin, of Candida bloodstream isolates. results from a six-year study (1996–2001). Diagn Microbiol Infect Dis. 2003;46(4):259–264. doi:10.1016/S0732-8893(03)00086-5
112. Pfaller MA, Jones RN, Messer SA, Edmond MB, Wenzel RP. National surveillance of nosocomial blood stream infection due to species of Candida other than Candida albicans: frequency of occurrence and antifungal susceptibility in the SCOPE program. Diagn Microbiol Infect Dis. 1998;30(2):121–129. doi:10.1016/S0732-8893(97)00192-2
113. Sabino R, Verissimo C, Brandao J, et al. Epidemiology of candidemia in oncology patients: a 6-year survey in a Portuguese central hospital. Med Mycol. 2010;48(2):346–354. doi:10.3109/13693780903161216
114. Xess I, Jain N, Hasan F, Mandal P, Banerjee U. Epidemiology of candidemia in a tertiary care centre of North India: 5-year study. Infection. 2007;35(4):256–259. doi:10.1007/s15010-007-6144-6
115. Chen C-Y, Huang S-Y, Tsay W, et al. Clinical characteristics of candidaemia in adults with haematological malignancy, and antimicrobial susceptibilities of the isolates at a medical centre in Taiwan, 2001–2010. Int J Antimicrob Agents. 2012;40(6):533–538. doi:10.1016/j.ijantimicag.2012.07.022
116. Bukharie HA. Nosocomial candidemia in a tertiary care hospital in Saudi Arabia. Mycopathologia. 2002;153(4):195–198. doi:10.1023/A:1014945517790
117. Playford EG, Marriott D, Nguyen Q, et al. Candidemia in nonneutropenic critically ill patients: risk factors for non-albicans Candida spp. Crit Care Med. 2008;36(7):2034–2039. doi:10.1097/CCM.0b013e3181760f42
118. Ko J-H, Jung DS, Lee JY, et al. Changing epidemiology of non albicans candidemia in Korea. J Infect Chemother. 2019;25(5):388–391. doi:10.1016/j.jiac.2018.09.016
119. Bonfietti LX, Szeszs MW, Chang MR, et al. Ten-year study of species distribution and antifungal susceptibilities of Candida bloodstream isolates at a Brazilian tertiary hospital. Mycopathologia. 2012;174(5):389–396. doi:10.1007/s11046-012-9566-3
120. Dimopoulos G, Ntziora F, Rachiotis G, Armaganidis A, Falagas ME. Candida albicans versus non-albicans intensive care unit-acquired bloodstream infections: differences in risk factors and outcome. Anesth Analg. 2008;106(2):523–529. doi:10.1213/ane.0b013e3181607262
121. Papadimitriou-Olivgeris M, Spiliopoulou A, Kolonitsiou F, et al. Increasing incidence of candidaemia and shifting epidemiology in favor of Candida non-albicans in a 9-year period (2009–2017) in a university Greek hospital. Infection. 2019;47(2):209–216. doi:10.1007/s15010-018-1217-2
122. Treviño-Rangel R, Peña-López CD, Hernández-Rodríguez PA, Beltrán-Santiago D, González GM. Association between Candida biofilm-forming bloodstream isolates and the clinical evolution in patients with candidemia: an observational nine-year single center study in Mexico. Rev Iberoam Micol. 2018;35(1):11–16. doi:10.1016/j.riam.2017.01.005
123. Marchetti O, Bille J, Fluckiger U, et al. Epidemiology of candidemia in Swiss tertiary care hospitals: secular trends, 1991–2000. Clin Infect Dis. 2004;38(3):311–320. doi:10.1086/380637
124. Fridkin SK, Kaufman D, Edwards JR, Shetty S, Horan T. Changing incidence of Candida bloodstream infections among NICU patients in the United States: 1995–2004. Pediatrics. 2006;117(5):1680–1687. doi:10.1542/peds.2005-1996
125. Hachem R, Hanna H, Kontoyiannis D, Jiang Y, Raad I. The changing epidemiology of invasive candidiasis. Cancer. 2008;112(11):2493–2499. doi:10.1002/cncr.23466
126. Sipsas NV, Lewis RE, Tarrand J, et al. Candidemia in patients with hematologic malignancies in the era of new antifungal agents (2001-2007). Cancer. 2009;115(20):4745–4752. doi:10.1002/cncr.24507
127. Schuster MG, Meibohm A, Lloyd L, Strom B. Risk factors and outcomes of Candida krusei bloodstream infection: A matched, case-control study. J Infect. 2013;66(3):278–284. doi:10.1016/j.jinf.2012.11.002
128. Lin MY, Carmeli Y, Zumsteg J, et al. Prior antimicrobial therapy and risk for hospital-acquired Candida glabrata and Candida krusei fungemia: a case-case-control study. Antimicrob Agents Chemother. 2005;49(11):4555–4560. doi:10.1128/AAC.49.11.4555-4560.2005
129. Amaral-Lopes S, Moura A. Neonatal fungal sepsis by Candida krusei: A report of three cases and a literature review. Medical Mycol Case Rep. 2012;1(1):24–26. doi:10.1016/j.mmcr.2012.04.002
130. Prasad PA, Fisher BT, Coffin SE, et al. Pediatric risk factors for candidemia secondary to Candida glabrata and Candida krusei species. J Pediatric Infect Dis Soc. 2013;2(3):263–266. doi:10.1093/jpids/pis093
131. Fan SR, Liu XP, Li JW. Clinical characteristics of vulvovaginal candidiasis and antifungal susceptibilities of Candida species isolates among patients in southern China from 2003 to 2006. J Obstet and Gynaecol Res. 2008;34(4):561–566. doi:10.1111/j.1447-0756.2008.00817.x
132. Imtiaz T, Thomson F, Innes A, du Toit FC, Bal AM. Candida krusei bronchopneumonia with nodular infiltrates in a patient with chronic renal failure on haemodialysis – case report and review of literature. Mycoses. 2011;54(5):e611–e614. doi:10.1111/j.1439-0507.2010.01925.x
133. Rook GD, Brand D. Candida krusei as a pathogen; case report of an unusual infection of the tonsils. Pediatrics. 1950;6(4):638–643.
134. Lu H, Marengo MF, Mihu CN, Garcia-Manero G, Suarez-Almazor ME. Rare case of septic arthritis caused by Candida krusei: case report and literature review. J Rheumatol. 2012;39(6):1308–1309. doi:10.3899/jrheum.111348
135. Jud P, Valentin T, Regauer S, et al. Invasive Candida krusei infection and Candida vasculitis of a leg ulcer in an immunocompetent patient: a case report. Int J Infect Dis. 2017;55:96–98. doi:10.1016/j.ijid.2017.01.010
136. Multani A, Subramanian AK, Liu AY. Successful eradication of chronic symptomatic Candida krusei urinary tract infection with increased dose micafungin in a liver and kidney transplant recipient: case report and review of the literature. Transpl Infect Dis. 2019;21(4):e13118. doi:10.1111/tid.13118
137. Kano R, Konishi K, Nakata K, et al. Isolation of Candida krusei from a case of bovine bronchopneumonia in a one-year-old heifer. Vet Rec. 2001;148(20):636. doi:10.1136/vr.148.20.636
138. Gaudie CM, Wragg PN, Barber AM. Outbreak of disease due to Candida krusei in a small dairy herd in the UK. Vet Rec. 2009;165(18):535–537. doi:10.1136/vr.165.18.535
139. Sheena A, Sigler L. Candida krusei isolated from a sporadic case of bovine mastitis. Can Vet J. 1995;36(6):365.
140. Seker E. Identification of Candida species isolated from bovine mastitic milk and their in vitro hemolytic activity in Western Turkey. Mycopathologia. 2010;169(4):303–308.
141. Du J, Wang X, Luo H, Wang Y, Liu X, Zhou X. Epidemiological investigation of non-albicans Candida species recovered from mycotic mastitis of cows in Yinchuan, Ningxia of China. BMC Vet Res. 2018;14(1):251. doi:10.1186/s12917-018-1564-3
142. Ksouri S, Djebir S, Hadef Y, Benakhla A. Survey of bovine mycotic mastitis in different mammary gland statuses in two north-eastern regions of Algeria. Mycopathologia. 2015;179(3–4):327–331. doi:10.1007/s11046-014-9845-2
143. Wawron W, Bochniarz M, Szczubiał M. Enzymatic activity of yeasts isolated from the inflamed mammary secretion in dairy cows. Pol J Vet Sci. 2011;14(1):65–68. doi:10.2478/v10181-011-0009-8
144. Boutilier P, Carr A. Fungal colonization and failure of a long-term gastrostomy tube in a cat. Can Vet J. 2005;46(8):709–710.
145. Elad D, Shpigel NY, Winkler M, et al. Feed contamination with Candida krusei as a probable source of mycotic mastitis in dairy cows. J Am Vet Med Assoc. 1995;207(5):620–622.
146. Donnelly KA, Wellehan JFX
147. Muir M, Raidal SR. Necrotising ventriculitis due to combined infection with Rhizopus microsporus var. chinensis and Candida krusei in an eclectus parrot (Eclectus roratus). Aust Vet J. 2012;90(7):277–280. doi:10.1111/j.1751-0813.2012.00929.x
148. Daef E, Moharram A, Eldin SS, Elsherbiny N, Mohammed M. Evaluation of chromogenic media and seminested PCR in the identification of Candida species. Braz J Microbiol. 2014;45(1):255–262. doi:10.1590/S1517-83822014005000040
149. Baixench MT, Taillandier A, Paugam A. Clinical and experimental evaluation of a new chromogenic medium (OCCA, Oxoid) for direct identification of Candida albicans, C. tropicalis and C. krusei. Mycoses. 2006;49(4):311–315. doi:10.1111/j.1439-0507.2006.01259.x
150. Zhao L, de Hoog GS, Cornelissen A, et al. Prospective evaluation of the chromogenic medium CandiSelect 4 for differentiation and presumptive identification of non-Candida albicans Candida species. Fungal Biol. 2016;120(2):173–178. doi:10.1016/j.funbio.2015.09.006
151. Szabó Z, Tóth B, Kovács M, et al. Evaluation of the new Micronaut-Candida system compared to the API ID32C method for yeast identification. J Clin Microbiol. 2008;46(5):1824–1825. doi:10.1128/JCM.02350-07
152. Meurman O, Koskensalo A, Rantakokko-Jalava K. Evaluation of Vitek 2 for identification of yeasts in the clinical laboratory. Clin Microbiol Infect. 2006;12(6):591–593. doi:10.1111/j.1469-0691.2006.01409.x
153. Hertel M, Hartwig S, Schütte E, et al. Identification of signature volatiles to discriminate Candida albicans, glabrata, krusei and tropicalis using gas chromatography and mass spectrometry. Mycoses. 2016;59(2):117–126. doi:10.1111/myc.12442
154. Morace G, Sanguinetti M, Posteraro B, Lo Cascio G, Fadda G. Identification of various medically important Candida species in clinical specimens by PCR-restriction enzyme analysis. J Clin Microbiol. 1997;35(3):667–672. doi:10.1128/JCM.35.3.667-672.1997
155. Maiwald M, Kappe R, Sonntag HG. Rapid presumptive identification of medically relevant yeasts to the species level by polymerase chain reaction and restriction enzyme analysis. J Med Vet Mycol. 1994;32(2):115–122. doi:10.1080/02681219480000161
156. Rezazadeh E, Moazeni M, Sabokbar A. Use of cost effective and rapid molecular tools for identification of Candida species, opportunistic pathogens. Curr Med Mycol. 2016;2(3):1–4. doi:10.18869/acadpub.cmm.2.3.1
157. Carlotti A, Chaib F, Couble A, Bourgeois N, Blanchard V, Villard J. Rapid identification and fingerprinting of Candida krusei by PCR-based amplification of the species-specific repetitive polymorphic sequence CKRS-1. J Clin Microbiol. 1997;35(6):1337–1343. doi:10.1128/JCM.35.6.1337-1343.1997
158. Kanbe T, Horii T, Arishima T, Ozeki M, Kikuchi A. PCR-based identification of pathogenic Candida species using primer mixes specific to Candida DNA topoisomerase II genes. Yeast. 2002;19(11):973–989. doi:10.1002/yea.892
159. Guo Y, Yang J-X, Liang G-W. A real-time PCR assay based on 5.8S rRNA gene (5.8S rDNA) for rapid detection of Candida from whole blood samples. Mycopathologia. 2016;181(5–6):405–413. doi:10.1007/s11046-015-9977-z
160. Fidler G, Leiter E, Kocsube S, Biro S, Paholcsek M. Validation of a simplex PCR assay enabling reliable identification of clinically relevant Candida species. BMC Infect Dis. 2018;18(1):393. doi:10.1186/s12879-018-3283-6
161. Decat E, Van Mechelen E, Saerens B, et al. Rapid and accurate identification of isolates of Candida species by melting peak and melting curve analysis of the internally transcribed spacer region 2 fragment (ITS2-MCA). Res Microbiol. 2013;164(2):110–117. doi:10.1016/j.resmic.2012.10.017
162. Sturaro LL, Gonoi T, Busso-Lopes AF, et al. Visible DNA microarray system as an adjunctive molecular test in identification of pathogenic fungi directly from a blood culture bottle. J Clin Microbiol. 2018;56(5):e01908–01917. doi:10.1128/JCM.01908-17
163. Das S, Brown TM, Kellar KL, Holloway BP, Morrison CJ. DNA probes for the rapid identification of medically important Candida species using a multianalyte profiling system. FEMS Immunol Med Microbiol. 2006;46(2):244–250. doi:10.1111/j.1574-695X.2005.00031.x
164. Freydiere AM, Buchaille L, Guinet R, Gille Y. Evaluation of latex reagents for rapid identification of Candida albicans and C. krusei colonies. J Clin Microbiology. 1997;35(4):877–880. doi:10.1128/JCM.35.4.877-880.1997
165. Whaley SG, Berkow EL, Rybak JM, Nishimoto AT, Barker KS, Rogers PD. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida species. Front Microbiol. 2017;7:2173. doi:10.3389/fmicb.2016.02173
166. Katiyar SK, Edlind TD. Identification and expression of multidrug resistance-related ABC transporter genes in. Candida Krusei. Med Mycol. 2001;39(1):109–116. doi:10.1080/mmy.39.1.109.116
167. Ricardo E, Miranda IM, Faria-Ramos I, Silva RM, Rodrigues AG, Pina-Vaz C. In vivo and in vitro acquisition of resistance to voriconazole by Candida krusei. Antimicrob Agents Chemother. 2014;58(8):4604–4611. doi:10.1128/AAC.02603-14
168. Gong J, Xiao M, Wang H, et al. Genetic differentiation, diversity, and drug susceptibility of Candida krusei. Front Microbiol. 2018;9:2717. doi:10.3389/fmicb.2018.02717
169. Nguyen KT, Ta P, Hoang BT, et al. Anidulafungin is fungicidal and exerts a variety of postantifungal effects against Candida albicans, C. glabrata, C. parapsilosis, and C. krusei isolates. Antimicrob Agents Chemother. 2009;53(8):3347–3352. doi:10.1128/AAC.01480-08
170. Forastiero A, Garcia-Gil V, Rivero-Menendez O, et al. Rapid development of Candida krusei echinocandin resistance during caspofungin therapy. Antimicrob Agents Chemother. 2015;59(11):6975–6982. doi:10.1128/AAC.01005-15
171. Mendling W. Guideline: vulvovaginal candidosis (AWMF 015/072), S2k (excluding chronic mucocutaneous candidosis). Mycoses. 2015;58(S1):1–15. doi:10.1111/myc.12292
172. Locke JB, Almaguer AL, Donatelli JL, Bartizal KF. Time-kill kinetics of rezafungin (CD101) in vagina-simulative medium for fluconazole-susceptible and fluconazole-resistant Candida albicans and non-albicans Candida species. Infect Dis Obstet Gynecol. 2018;2018:7040498. doi:10.1155/2018/7040498
173. Mackie DP, Neill SD, Rodgers SP, Logan EF. Treatment of Candida krusei mastitis with sulphamethoxypyridazine. Vet Rec. 1987;120(2):48. doi:10.1136/vr.120.2.48
174. Schell WA, Jones AM, Garvey EP, Hoekstra WJ, Schotzinger RJ, Alexander BD. Fungal CYP51 inhibitors VT-1161 and VT-1129 exhibit strong in vitro activity against Candida glabrata and C. krusei isolates clinically resistant to azole and echinocandin antifungal compounds. Antimicrob Agents Chemother. 2017;61(3):e01817–01816. doi:10.1128/AAC.01817-16
175. Domingues Bianchin M, Borowicz SM, da Rosa Monte Machado G, et al. Lipid core nanoparticles as a broad strategy to reverse fluconazole resistance in multiple Candida species. Colloids Surf B Biointerfaces. 2019;175:523–529. doi:10.1016/j.colsurfb.2018.12.011
176. Wang T, Shao J, Da W, et al. Strong synergism of palmatine and fluconazole/itraconazole against planktonic and biofilm cells of Candida species and efflux-associated antifungal mechanism. Front Microbiol. 2018;9:2892. doi:10.3389/fmicb.2018.02892
177. Patriota LL, Procópio TF, de Souza MF, et al. A Trypsin inhibitor from Tecoma stans leaves inhibits growth and promotes ATP depletion and lipid peroxidation in Candida albicans and Candida krusei. Front Microbiol. 2016;7:611. doi:10.3389/fmicb.2016.00611
178. Bezerra CF, Rocha JE, Nascimento Silva MKD, et al. Analysis by UPLC-MS-QTOF and antifungal activity of guava (Psidium guajava L.). Food Chem Toxicol. 2018;119:122–132. doi:10.1016/j.fct.2018.05.021
179. Souza-Moreira TM, Severi JA, Rodrigues ER, et al. Flavonoids from Plinia cauliflora (Mart.) Kausel (Myrtaceae) with antifungal activity. Nat Prod Res. 2019;33(17):2579–2582. doi:10.1080/14786419.2018.1460827
180. Moraes RC, Carvalho AR, Lana AJ, et al. In vitro synergism of a water insoluble fraction of Uncaria tomentosa combined with fluconazole and terbinafine against resistant non-Candida albicans isolates. Pharm Biol. 2017;55(1):406–415. doi:10.1080/13880209.2016.1242631
181. Min J, Lee YJ, Kim YA, et al. Lysophosphatidylcholine derived from deer antler extract suppresses hyphal transition in Candida albicans through MAP kinase pathway. Biochim Biophys Acta. 2001;1531(1–2):77–89. doi:10.1016/S1388-1981(01)00088-9
182. Nami S, Aghebati-Maleki A, Morovati H, Aghebati-Maleki L. Current antifungal drugs and immunotherapeutic approaches as promising strategies to treatment of fungal diseases. Biomed Pharmacother. 2019;110:857–868. doi:10.1016/j.biopha.2018.12.009
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.