")
Back to Journals » International Journal of General Medicine » Volume 15
CTHRC1 is a Potential Prognostic Biomarker and Correlated with Macrophage Infiltration in Breast Cancer
Authors Wang Z, Zhang S, Zheng C, Xia K, Sun L, Tang X, Zhou F, Ouyang Y, Tang F
Received 13 March 2022
Accepted for publication 13 June 2022
Published 20 June 2022 Volume 2022:15 Pages 5701—5713
DOI https://doi.org/10.2147/IJGM.S366272
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Zejun Wang,1,* Shichao Zhang,2,* Chaochao Zheng,3 Kaide Xia,4 Liangquan Sun,4 Xuejie Tang,4 Fulin Zhou,4 Yan Ouyang,2 Fuzhou Tang3
1Department of Gastrointestinal Surgery, The Affiliated Cancer Hospital of Guizhou Medical University, Guiyang, People’s Republic of China; 2Key Laboratory of Infectious Immune and Antibody Engineering in Guizhou Province/School of Biology and Engineering, Guizhou Medical University, Guiyang, People’s Republic of China; 3Immune Cells and Antibody Engineering Research Center of Guizhou province/School of Biology and Engineering, Guizhou Medical University, Guiyang, People’s Republic of China; 4Guiyang Maternal and Child Health Care Hospital, Guiyang Children’s Hospital, Guiyang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Fuzhou Tang, Immune Cells and Antibody Engineering Research Center of Guizhou province/School of Biology and Engineering, Guizhou Medical University, Guiyang, People’s Republic of China, Tel +86 085188174043, Email [email protected] Yan Ouyang, Key Laboratory of Infectious Immune and Antibody Engineering in Guizhou province/School of Biology and Engineering, Guizhou Medical University, Guiyang, People’s Republic of China, Email [email protected]
Background: Tumor immune cell infiltration is closely associated with the occurrence and development of tumors. Collagen triple helix repeats containing 1 (CTHRC1), a regulator of collagen expression and cell migration, is involved in the metastasis and invasion of tumors. However, the role of CTHRC1 in breast cancer remains unclear. This study aimed to investigate the prognostic value of CTHRC1, and further explore its association with immune infiltration in breast cancer.
Methods: CTHRC1 expression pattern and prognostic value were analyzed using ONCOMINE, PrognoScan, GEPIA, and Kaplan–Meier Plotter databases. We then detected CTHRC1 mRNA levels in breast cancer tissues and paired normal breast tissues by Q-PCR. Subsequently, the University of California Santa Cruz (UCSC) database was used to determine the methylation status of CTHRC1. Furthermore, CTHRC1 mutations were investigated using the Catalogue of Somatic mutations in Cancer (COSMIC) and cBioPortal databases. We also assessed the correlation between CTHRC1 expression and immune cell infiltration using TIMER. In addition, The relationship of CTHRC1 expression with the immune marker sets of various immune cells was evaluated using GEPIA and TIMER.
Results: CTHRC1 was highly expressed in a variety of tumors, including breast cancer. Elevated CTHRC1 expression was related to a poor prognosis. Notably, CTHRC1 expression was significantly associated with macrophage infiltration, especially the immune infiltration gene marker set of M2. Copy number variations, DNA mutations and methylation states might be potential mechanisms for regulating CTHRC1 expression. Protein digestion and absorption, human papillomavirus infection, ECM-receptor interaction, focal adhesion, and PI3K-Akt signaling pathways were identified as the potential CTHRC1-driven signaling pathways.
Conclusion: These findings suggest that CTHRC1 could be a promising immune-related biomarker for the treatment of breast cancer patients.
Keywords: CTHRC1, breast cancer, prognostic, macrophage, biomarker
Introduction
Breast cancer is the most commonly diagnosed tumor in women.1 Although there have been improvements in early clinical detection and therapy strategies, the prognosis for breast cancer patients remains unsatisfying.2 With the rapid development of high-throughput sequencing technology, researchers have identified a number of prognosis-related genes in breast cancer, including MUC1, COL12A1, CDK6, TXN, SEMA3B, and CCNB2.3–5 Recently, tumor immune cell infiltration has been reported to play an important role in tumor progression and to influence the immunotherapy effect.6 Researchers also found an adaptive immune response against breast tumors in either the patient’s peripheral blood or the tumor.2 Accurate biomarkers for the early clinical diagnosis and prognosis evaluation in breast cancer patients could improve the efficiency of current treatments and provide potential molecular markers for targeted therapies. Thus, identification of immune-related biomarkers plays a crucial role in the immunotherapy of breast cancers.
Collagen-containing triple helix repeat-1 (CTHRC1), a secreted 28-kDa protein, was first discovered in balloon-injured arteries of rats.7,8 CTHRC1 is expressed by fibroblasts, which reshapes the adventitia, and by smooth muscle cells of the neointima, which inhibits collagen expression, and increases cell migration.7 Subsequent studies found that CTHRC1 is anchored in the cell surface by the physical interaction between Wnt ligands and coiled receptors, and that these anchors also activated the planar cell polarity pathway in Wnt signaling to regulate cell motility and gliding.9 CTHRC1 is abnormally upregulated in a variety of tumor types (eg, lung, breast, thyroid, ovarian, cervical, liver, and pancreatic cancers), and it is associated with the tumor progression.10–12 Furthermore, CTHRC1 plays an important role in the regulation of the tumor microenvironment such as M2-like macrophage recruitment, and invasion in endometrial carcinoma.13 Although a significant correlation between CTHRC1 overexpression and poor prognosis has been reported in breast invasive ductal carcinomas,14 the mechanism of CTHRC1 overexpression and the relationship between CTHRC1 expression and immune infiltration remain unclear.
In this paper, we systematically analyzed the correlation between CTHRC1 expression and the prognosis of breast cancer patients based on ONCOMINE, PrognoScan, GEPIA, and Kaplan-Meier Plotter databases. Then, the methylation status and mutations of CTHRC1 were investigated using UCSC Cancer Genomics Browser as well as COSMIC and cBioPortal databases. Moreover, the association between CTHRC1 expression and immune cell infiltration in the tumor microenvironment was evaluated by TIMER and GEPIA, and potential CTHRC1-driven signaling pathways were analyzed by KEGG.
Materials and Methods
Oncomine Database Analysis
CTHRC1 expression levels in the different types of cancers were obtained from the Oncomine database (https://www.oncomine.org/resource/login.html).15 Parameters used were: P-value of 0.001, fold change of 1.5, and gene ranking of all.
UCSC Cancer Genomics Browser Analysis
The heat map of CTHRC1 expression was analyzed using the UCSC Cancer Genomics Browser (http://xena.ucsc.edu/).16–18 The regulatory mechanism of CTHRC1 expression was detected using TCGA Breast Cancer (dataset ID: TCGA.BRCA.sampleMap/HumanMethylation450).
COSMIC Analysis
To analyze CTHRC1 mutations in breast cancer, the percentage of CTHRC1 mutations and the alteration frequency of CTHRC1 mutations were investigated by the COSMIC database (http://cancer.sanger.ac.uk) and cBioPortal database.19,20
RNA Extraction and Q-PCR
Breast cancer patients with distant metastases, and with a history of other solid tumors, radiotherapy, chemotherapy, or neoadjuvant chemotherapy, were excluded. A total of 24 samples (including 12 tumor tissues and 12 adjacent normal tissues) were acquired from 12 breast cancer patients. The study was conducted according to the criteria set in the Declaration of Helsinki and all patients provided informed consent. The study protocol was approved by the Ethics Committee of Guizhou Medical University [reference number 2022(105)].
Trizol (Takara, 9108, Japan) was used to extract total RNA from breast cancer and normal breast tissues. Then, CTHRC1 mRNA expression was analyzed by 2X Super SYBR Green qPCR Master Mix (ES Science, QP220, China). The primer sequences for CTHRC1 were: forward, 5’-GCGGAGTGTACATTTACAAAGATGCG-3’; reverse, 5’-GAAATACCAACTCTGACAGCATGC-3’; The primer sequences for GAPDH were: forward, 5’-GACCTGACCTGCCGTCTA-3’; reverse, 5’-AGGAGTGGGTGTCGCTGT-3’.
Kaplan-Meier Plotter Database Analysis
Kaplan-Meier curves (http://kmplot.com/analysis/) were plotted to distinguish the low- or high-risk of breast cancer patients based on the relationship between CTHRC1 expression and survival.21 The Log rank test was used. The hazard ratios (HR) were calculated with 95% confidence intervals, P-values were two-sided, and analyses were conducted using R/Bioconductor.
TIMER Database Analysis
To evaluate the association between CTHRC1 expression and tumor immune cell infiltration, the TIMER database was used (https://cistrome.shinyapps.io/timer/).22 According to the gene expression profiles, tumor infiltration immune cell can be identified via the deconvolution statistical method using The Cancer Genome Atlas (TCGA).23,24 We used the TIMER database to explore CTHRC1 expression in different types of cancers and the relationship between CTHRC1 expression and tumor immune cell infiltration. These immune cells contained dendritic cells, CD4+ T cells, CD8+ T cells, macrophages, B cells, and neutrophils. We also analyzed the association between CTHRC1 expression and tumor immune cell infiltration gene markers by correlation modules based on the TIMER database. These gene markers included markers of dendritic cells, M1 macrophages, M2 macrophages, T cells (general), monocytes, CD8+ T cells, TAMs, B cells, and neutrophils.25,26 In the expression scatter plots, Spearman correlation was used to evaluate statistical significance. Gene expression levels were presented in log2 TRM.
Gene Correlation Analysis in GEPIA
Gene Expression Profiling Interactive Analysis (GEPIA) (http://gepia.cancer-pku.cn/index.html) was used to further identify CTHRC1 related genes. Survival curves were obtained from GEPIA based on gene expression with the Log rank test and the Mantel-Cox test for tumors. Correlation analysis of gene expression was performed on the given TCGA expression data set. The correlation coefficient was analyzed by the Spearman method.
Functional Enrichment Analysis
The genes coexpressed with CTHRC1 were obtained from cBioPortal for Cancer Genomics (http://www.cbioportal.org/) based on |Spearman’s r| > 0.642. The enrichment analysis of these coexpressed genes included Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways with the false discovery rate (FDR) < 0.05.
Statistical Analysis
CTHRC1 expression differences between breast cancer and normal groups were evaluated using unpaired Welch’s t-test. The correlations between CTHRC1 expression and prognosis, CTHRC1 expression and tumor immune cell infiltration, and CTHRC1 expression and tumor immune cell infiltration gene markers were detected by Pearson’s correlation analysis. Gene expression levels were presented as log2 TRM. P-values < 0.05 were considered statistically significant.
Results
CTHRC1 Expression Levels in Cancers
CTHRC1 expression was detected using the Oncomine database, and higher CTHRC1 expression was shown in tumor tissues compared to normal tissues for brain, breast, cervical, colorectal, esophageal, gastric, head and neck, kidney, leukemia, liver, lung, lymphoma, melanoma, pancreatic, and prostate cancers, respectively (Figure 1A).
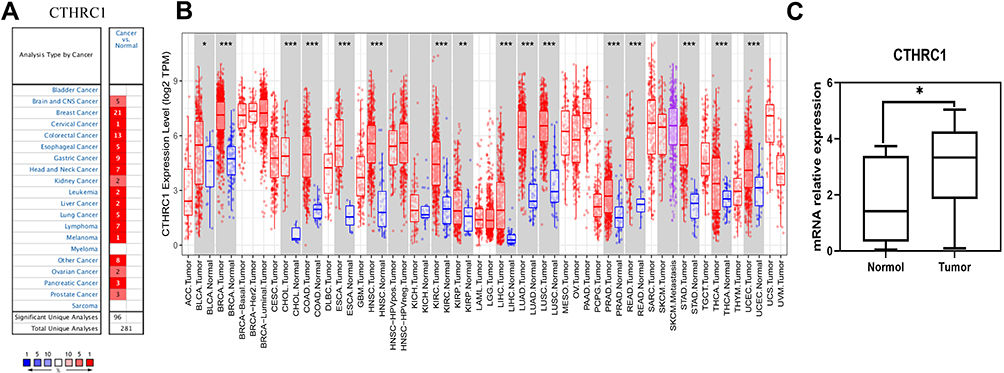 |
Figure 1 CTHRC1 expression status in various cancers. (A and B) CTHRC1 expression levels were assessed using the Oncomine (A), and TCGA databases (B). (C) The difference in CTHRC1 expression between breast cancer and normal tissues (n=12). (*P < 0.05, **P < 0.01, ***P < 0.001). |
To further explore CTHRC1 expression levels, CTHRC1 RNA sequence data from the tumors with their adjacent normal tissues in TCGA database were compared (Figure 1B). We discovered significantly high expression of CTHRC1 in many tumor types, including bladder urothelial carcinoma, BRCA (breast invasive carcinoma), CHOL (cholangiocarcinoma), COAD (colon adenocarcinoma), ECSA (Esophageal carcinoma), HNSC (head and neck cancer), KIRC (kidney renal clear cell carcinoma), KIRP (Kidney renal papillary cell carcinoma), LIHC (liver hepatocellular carcinoma), LUAD (lung adenocarcinoma), LUSC (Lung squamous cell carcinoma), PRAD (prostate adenocarcinoma), READ (rectum adenocarcinoma), STAD (stomach adenocarcinoma), STAD (Stomach adenocarcinoma), THCA (thyroid carcinoma), and uterine corpus endometrial carcinoma.
Moreover, we collected 12 paired cancer tissues and adjacent breast tissues, and verified that CTHRC1 expression was remarkably higher in cancer tissues than in adjacent nontumor tissues (Figure 1C).
Interestingly, multiple breast cancer types showed increased CTHRC1 expression (P < 1×10−4), including invasive lobular breast carcinoma (P = 8.47E−15) (Figure 2A), invasive ductal breast carcinoma (P = 1.33E−20) (Figure 2B), mixed lobular and ductal breast carcinoma (P = 5.06E−4) (Figure 2C), and invasive breast carcinoma (P = 2.68E−26) (Figure 2D).
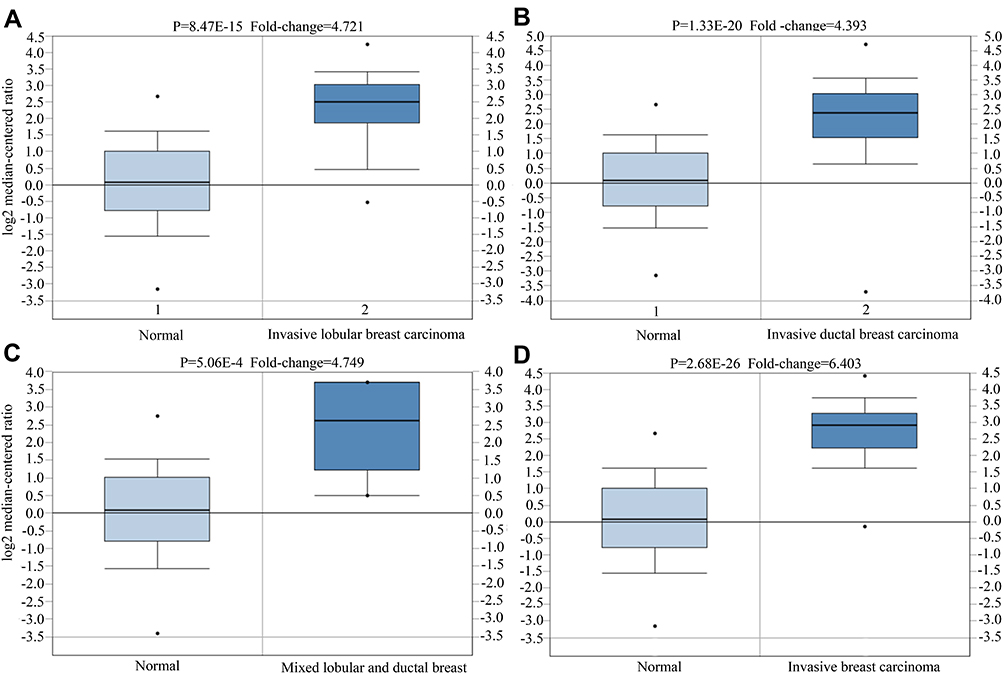 |
Figure 2 CTHRC1 expression status in different types of breast cancer. (A) Invasive lobular breast carcinoma, (B) invasive ductal breast carcinoma, (C) mixed lobular and ductal breast carcinoma, (D) invasive breast carcinoma. |
Methylation State, Mutation, and Copy Number Variation of CTHRC1 in Breast Cancer
In order to explore the regulatory mechanisms of AGR2 expression, CTHRC1 methylation states, mutations, and copy number variations were analyzed. We first found that DNA methylation of the promoter CTHRC1 was correlated with gene expression across PAM50 breast cancer subtypes (Figure 3A). Compared with normal tissues, methylation levels of CTHRC1 were significantly decreased in tumor tissues (P < 2.22E−16) (Figure 3B). Moreover, the mutant types are shown in Figure 4A, revealing that missense substitutions dominated (12.50%). Each C>G, G>A, and T>G mutation accounted for 33% of the CTHRC1 coding strand (Figure 4B). Finally, the amplified copy number of CTHRC1 (the alteration frequency of CTHRC1 was more than 7%) was observed in a variety of breast cancer cohorts, including Breast (METABRIC), Breast Invasive Carcinoma (TCGA; 2015 and Firehose Legacy), Metastatic Breast Cancer (INSERM), The metastatic Breast Cancer Project (Provisional), and Breast Invasive Carcinoma (TCGA; PanCaner Atlas and 2012). Together, these results suggest that DNA methylation and gene copy number variation were the potential mechanism for regulating the CTHRC1 expression.
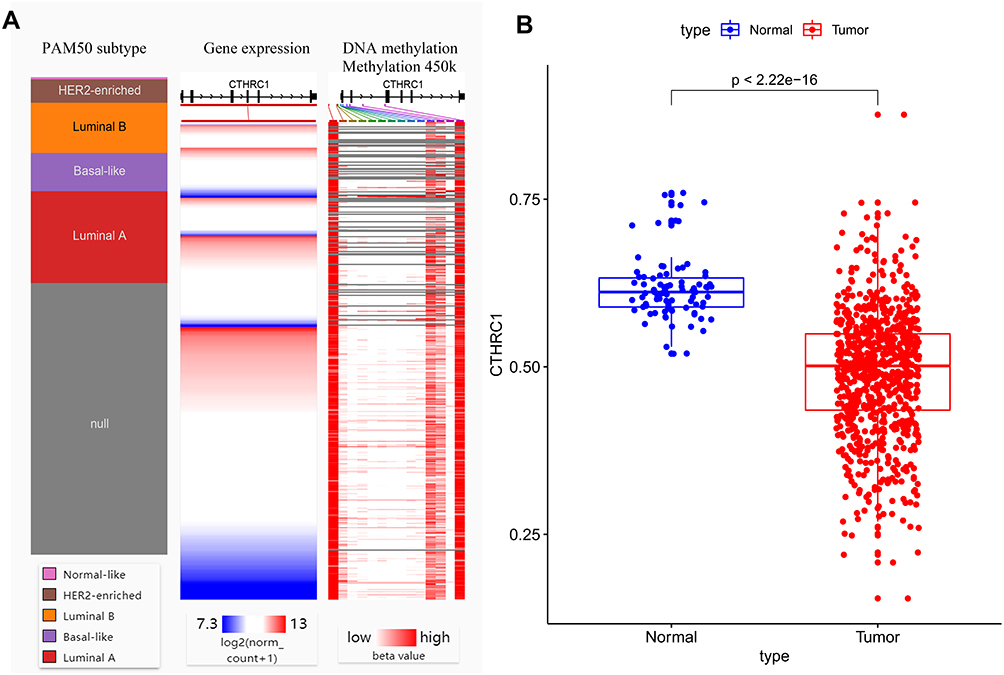 |
Figure 3 Association between CTHRC1 methylation status and its expression in breast cancer. (A) CTHRC1 expression heatmap and its DNA methylation status across PAM50 breast cancer subtypes. (B) Promoter methylation level of CTHRC1. |
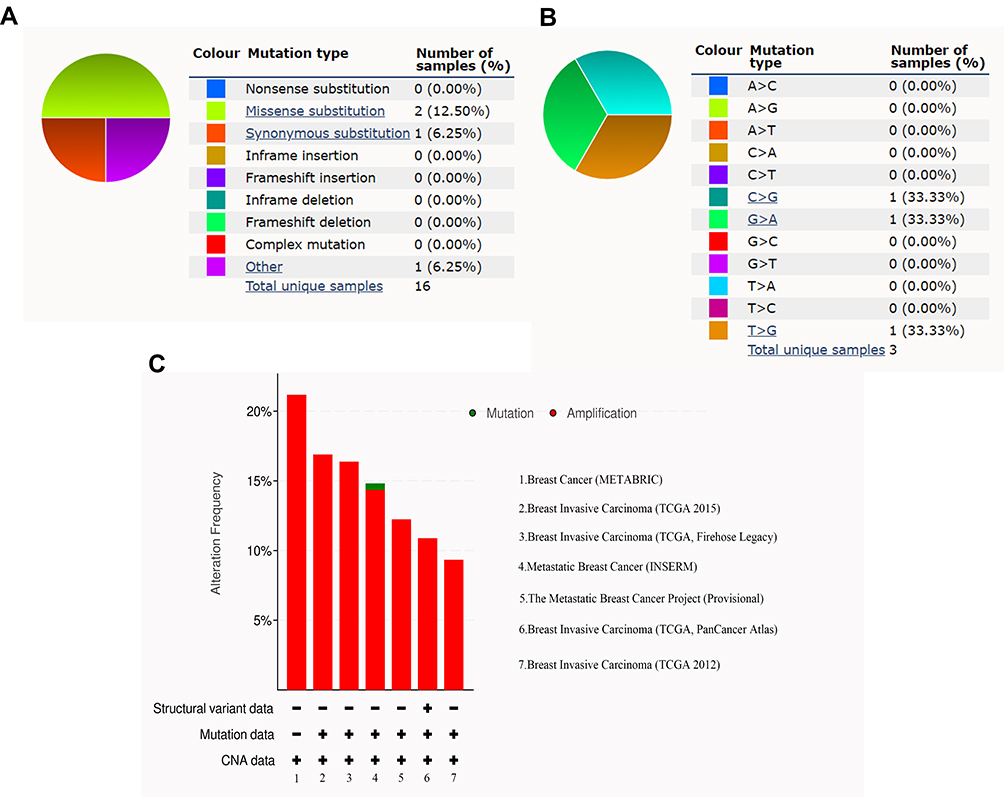 |
Figure 4 CTHRC1 mutations in breast cancer. (A and B) The percentage of CTHRC1 mutations based on COSMIC database. (C) The alteration frequency of CTHRC1 mutations based on cBioPortal. |
CTHRC1 Alterations and Clinicopathological Parameters in Breast Cancer Patients
Based on different clinicopathological indicators, the expression patterns of CTHRC1 were evaluated across PAM50 breast cancer subtypes. As shown in Figure 5, tumor patients with age ≤ 51 years had significantly high expression of CTHRC1 (P = 0.0481). CTHRC1 expression was also remarkably increased in the human epidermal growth factor receptor 2 (HER 2) (+) group (P < 0.0001). However, there was no significant differences between the statuses of estrogen receptors (ER) (+) and ER (-), the statuses of progesterone receptors (PR) (+) and PR (-), the statuses of non-TNBC and TNBC, and the statuses of non-basal-like and basal-like. It is worth notice that high CTHRC1 expression displayed in P53-mutant patients (P = 0.0023). In addition, the prognostic value of CTHRC1 expression was assessed by the Kaplan-Meier plotter database and UALCAN database, respectively. Patients with high CTHRC1 expression exhibited the decreased distant metastasis-free survival (DMFS) (Figure 6A), relapse-free survival (RFS) (Figure 6B), and overall survival (OS) time (Figure 6C). These findings indicate that CTHRC1 could be a potential prognostic biomarker for patients with breast cancer.
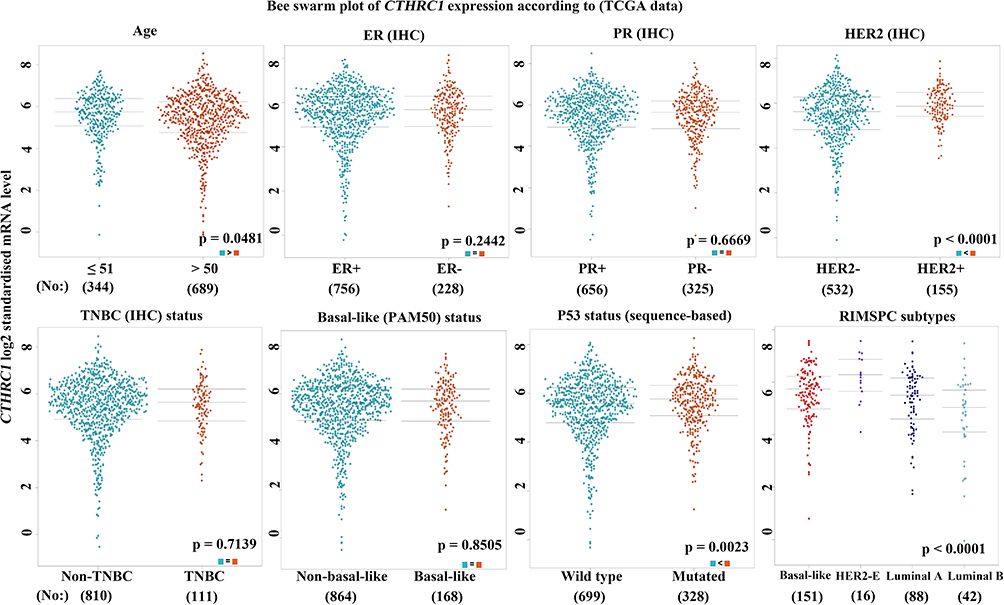 |
Figure 5 Association with CTHRC1 expression and clinicopathological parameters in breast cancer. |
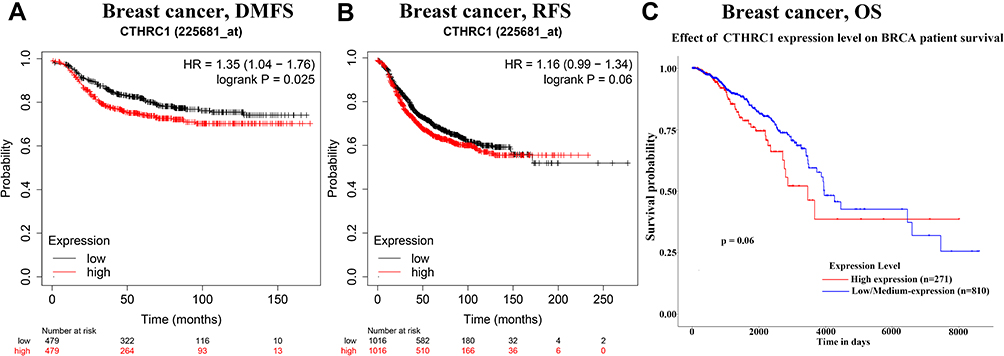 |
Figure 6 Prognostic signatures of CTHRC1 for breast cancer patients. (A) DMFS, (B) RFS, (C) OS. |
CTHRC1 Expression Related with Tumor Immune Cell Infiltration in Breast Cancers
Tumor immune cell infiltration plays an important role in modulating tumorigenesis, development, and cancer’s response to immunotherapy. Thus, we studied whether CTHRC1 expression was associated with tumor immune cell infiltration in breast cancer. The results showed that macrophage infiltration levels were significantly associated with CTHRC1 expression levels (Figure 7) (Partial.cor = 0.480, 0.386, 0.683, and 0.522 showed in BRCA, BRCA-Basal, BRCA-Her2, and BRCA-Luminal, respectively), which suggests that CTHRC1 might play an important role in inducing macrophage infiltration in breast cancer.
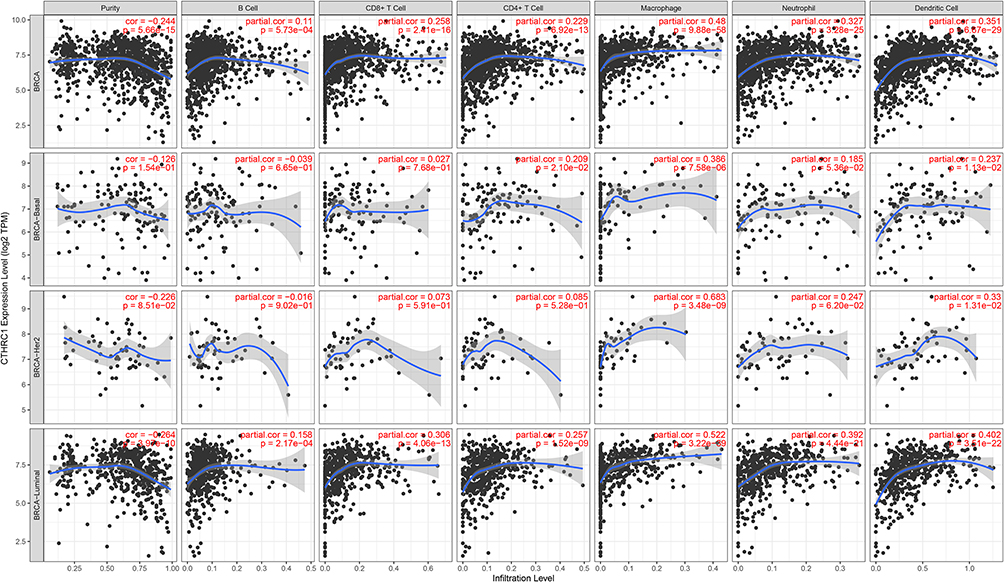 |
Figure 7 Association with CTHRC1 expression and immune cell infiltration in breast cancer. |
CHTCR1 Expression Associated with Immune Marker Sets
To further investigate the association between CTHRC1 expression and macrophage infiltration, the relationship between CTHRC1 expression and immune marker sets of immune cells (Monocyte, tumor-associated macrophage (TAM), M1 macrophage, and M2 macrophage) was assessed using the TIMER and GEPIA databases. The results revealed that CTHRC1 expression levels were significantly correlated with most of the tested immune marker sets in breast cancer, including CSF1R and CD86 of monocyte, CD68 and IL10 of TAMs, and CD163 and MS4A4A of the M2 phenotype (P < 0.001; Figure 8).
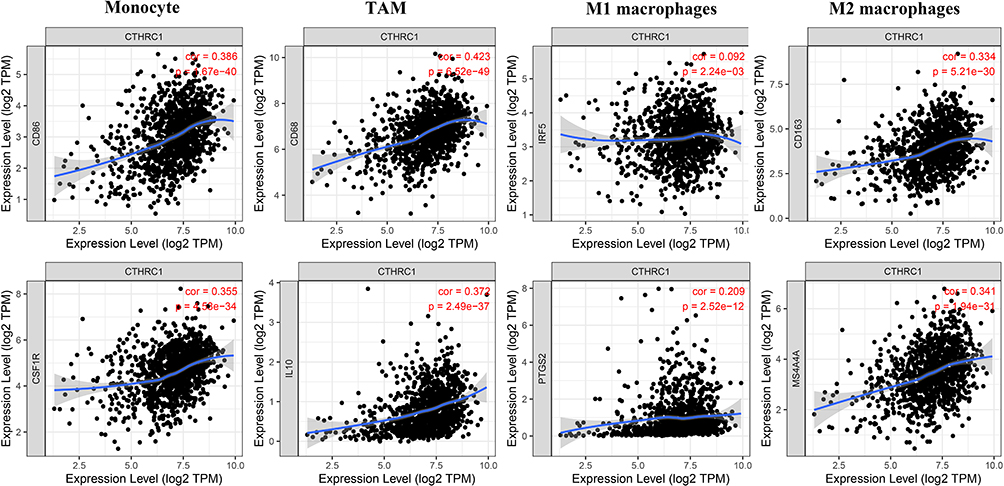 |
Figure 8 Correlation of CTHRC1 expression with macrophage polarization in breast cancer. Marker genes of monocytes (CD86 and CSF1R), TAMs (tumor-associated macrophages) (CD68 and IL10), M1 macrophages (IRF5 and PTGS2), M2 macrophages (CD163 and MS4A4A). |
Functional Enrichment Analysis of the Genes Co-Expressed with CTHRC1
The 100 genes were identified as co-expressed genes with CTHRC1 by using cBioPortal for Cancer Genomics in breast cancer with |Spearman’s r| > 0.642. The GO analysis results showed that in biological processes (BP) these genes were mainly enriched in extracellular matrix organization, extracellular structure organization, and ossification; in cellular components (CC) these genes were mainly enriched in collagen-containing extracellular matrix, endoplasmic reticulum lumen, and collagen trimer; in molecular functions (MF) these genes were mainly enriched in extracellular matrix structural constituent, glycosaminoglycan binding, and collagen binding (Figure 9A). The pathway analysis using the KEGG pathway database showed these genes were mainly enriched in protein digestion and absorption, human papillomavirus infection, ECM-receptor interaction, focal adhesion, and PI3K-Akt signaling pathway (Figure 9B).
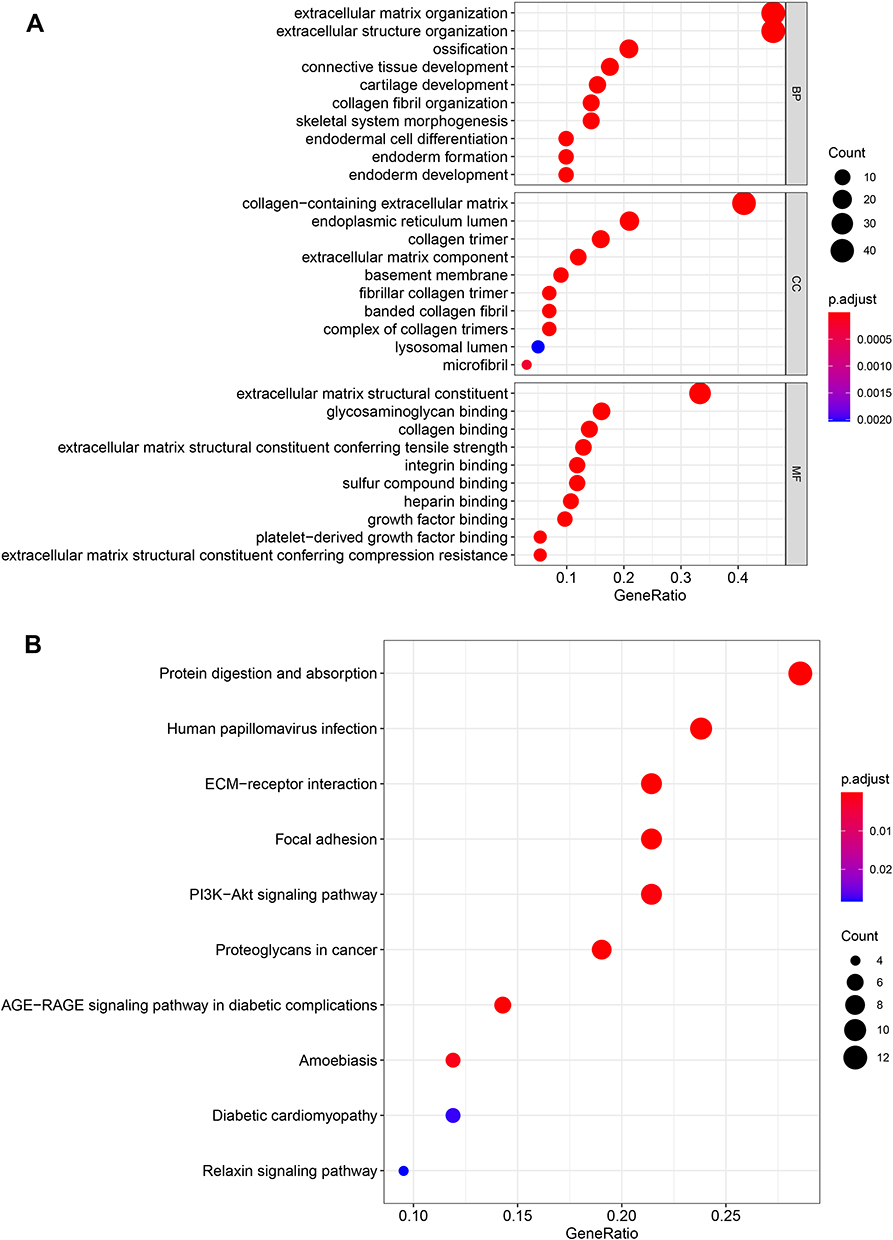 |
Figure 9 Functional enrichment analysis of the genes co-expressed with CTHRC1. (A) GO analysis, (B) KEGG pathway analysis. |
Discussion
CTHRC1 is a secretory glycoprotein that was discovered while differentially expressed genes were screened after severe arterial injury in rats.6,7 Subsequently, researchers reported that CTHRC1 is abnormally expressed in various tumor types, including breast cancer, and it is associated with tumor invasion and metastasis.12,27–30 Furthermore, CTHRC1 plays important roles in promoting M2-like macrophage recruitment, and invasion in endometrial carcinoma.13 Although the correlation between CTHRC1 expression and prognosis has been identified in the invasive ductal carcinoma of the breast,14 the role of CTHRC1 in breast cancer remains unclear.
In this study, CTHRC1 expression levels were evaluated using independent datasets from Oncomine, and 33 types of cancers with TCGA data in GEPIA. We found that CTHRC1 was highly expressed in various cancers. The high expression was confirmed by detecting the relative RNA expression of CTHRC1 in paired breast cancer and nontumor samples. The CTHRC1 transcriptional levels were obviously upregulated in invasive breast carcinoma, invasive lobular breast carcinoma, invasive ductal and mixed lobular, and ductal breast cancer. In addition, the overexpression of CTHRC1 was associated with increasing age and HER2 (+). These results suggest that high CTHRC1 expression might play a significant role in the development of breast cancer. The results of the survival analysis further revealed that the high CTHRC1 expression levels were related with bad prognosis, which was consistent with previous studies.14 Such poor prognosis have also been found in colorectal cancer and renal cell carcinomas.31,32 Thus, CTHRC1 could be used as a prognostic biomarker for breast cancer.
The infiltration of immune cells in tumors is closely related to the clinical results of tumor patients.5 Tumor-associated macrophages (TAMs) promote cell proliferation and invasion, which is associated with the poor prognosis of patients, including breast cancer.33,34 Thus, we analyzed the relationship between CTHRC1 expression and immunocyte infiltration. After cell purity correction, the macrophage infiltration was correlated with CTHRC1 expression and its subtypes (BRCA-Basal, BRCA-Her2, and BRCA-Luminal). The evaluation of CTHRC1 expression correlation with macrophage marker genes showed that CD68 and IL10 of TAM were positively associated with CTHRC1 expression. The results further supported that CTHRC1 was related with macrophage infiltration in breast cancer. Thus, high expression of CTHRC1 could play an important role in modulating macrophage infiltration, resulting in a poor prognosis.
Previous studies have shown that the recruitment of macrophages and M2 macrophage polarization correlate with a poor prognosis in breast cancer patients.27 TAMs inhibit the infiltration and function of anti-tumor CD8+ T cells (CTL), stimulate angiogenesis in the tumor, and promote the proliferation, and metastasis of tumor cells.28,29 M0 macrophages display strong plasticity and can change their phenotype based on environmental signals.27,28 Macrophage populations within tumors can be divided into M1 and M2 macrophages according to functional scales.35,36 M1-like macrophages are called classic activated macrophages, which show anti-tumor abilities by releasing pro-inflammatory cytokines (such as TNF and interleukin IL-2), active nitrogen, and oxygen intermediates.30,37 In contrast, M2-like macrophages, also known as alternative activated macrophages, promote tumor proliferation, and metastasis by producing tumor cytokines.38–40 CTHRC1 expression correlation with macrophage marker genes showed that CD86 and CSF1R of monocytes, CD68 and IL10 of TMAs, CD163 and MS4A4A genes of M2 macrophages were positively associated with CTHRC1 expression, respectively. Significantly, PTGS2 and IRF5 of the M1 macrophages were not correlated with CTHRC1 expression. These results revealed that high CTHRC1 expression might lead to poor prognosis by promoting the polarization role of the recruited macrophages to M2 macrophages.
Gene mutation plays an essential role in the onset and progression of cancer. Breast cancer had a missense mutation in CTHRC1 and a low frequency of CTHRC1 changes. In addition, CTHRC1 expression showed a strong association with DNA methylation status and copy number variations. DNA methylation is the main form of epigenetics, which acts as a transcriptional repressor regulating gene expression and playing an important role in tumorigenesis and development. Previous studies reported that CTHRC1 was downregulated by hypermethylation in hepatocellular carcinoma and renal carcinoma, but up-regulated by promoter demethylation in gastric cancer.17,27,32 DNA copy number variations refer to the deletion, insertion, duplication, and complex multi locus variations (from 1000 bp to millions of bp). This variation causes the activation of oncogenes and the inactivation of tumor suppressor genes, which eventually leads to tumorigenesis. Thus, DNA hymomethylation and copy number variation could be important causes of CTHRC1 upregulation in breast cancer.
Although recent researches have reported that CTHRC1 upregulation in breast cancer promoting cell proliferation, invasion and migration could be through activating GSK-3β/β-catenin signaling,41 the exact mechanisms of CTHRC1 involvement in breast cancer remains unclear. Therefore, high expression of CTHRC1 might promoting the polarization role of recruited macrophages to M2 macrophages, resulting in poor prognosis. The KEGG analysis of CTHRC1 coexpressed genes showed that signaling pathway candidates were driven by CTHRC1, including protein digestion and absorption, human papillomavirus infection, ECM-receptor interaction, focal adhesion, and PI3K-Akt signaling pathways. CTHRC1 promotes macrophage recruitment in endometrial carcinoma by integrin-AKT signaling pathway.13 Furthermore, several studies have revealed activating the PI3K/AKT signaling pathway induced M2 macrophage polarization in a variety of tumors, including colorectal cancer, esophageal squamous cell carcinoma and gastric cancer.42–45 Thus, CTHRC1 may induce macrophage recruitment and M2 macrophage polarization by regulating the PI3K-Akt signaling pathway.
Conclusion
In summary, we found that CTHRC1 expression was upregulated in breast cancer, and it was associated with increased immune infiltration, the polarization of macrophages to M2, and poor prognosis. Protein digestion and absorption, human papillomavirus infection, ECM-receptor interaction, focal adhesion, and PI3K-Akt signaling pathway were the potential CTHRC1-driven signaling pathways. Thus, this work could provide insights for understanding the role of CTHRC1 in influencing the prognosis of breast cancer patient’s tumor immunology, and the potential application of CTHRC1 as an immunotherapy biomarker.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We acknowledge TCGA database for providing their platforms and contributors for uploading their meaningful datasets, and acknowledge Yue Chang (Lanzhou University), Weili Guan (Guizhou Medical University), Ya Wei (Guizhou Medical University), Yu Xiong (Guizhou Medical University), and Wei Qiu (Guizhou Medical University) for theirs guidance on bioinformatics analysis.
Author Contributions
All authors met the following conditions:
1. Made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation. 2. Took part in drafting, revising or critically reviewing the article. 3. Agreed on the journal to which the article has been submitted and agreed to be accountable for all aspects of the work. 4. Agreed on the final approval of the version to be published. 5. Agreed to take responsibility and be accountable for the contents of the article.
Funding
This work was supported by the [the National Natural Science Foundation of China] under Grant [number 31860244, 31760264, 32100442, and 31960139]; [the Science and Technology Foundation of Guizhou Province] under Grant [[2020]1Z016, [2021]172, ZK[2021]025, [2021]431, [2020]1Y087, [2019]1275 and 19NSP002]; [Excellent Young Talents Plan of Guizhou Medical University] under Grant [2020(105)]; and [the Science and Technology foundation of Guizhou Health Committee] under Grant [gzwjkj2019-1-037].
Disclosure
The authors have declared that no competing interests exist.
References
1. Wilcock P, Webster RM. The breast cancer drug market. Nat Rev Drug Discov. 2021;20(5):339–340. doi:10.1038/d41573-021-00018-6
2. Disis ML, Stanton SE. Immunotherapy in breast cancer: an introduction. Breast. 2018;37:196–199. doi:10.1016/j.breast.2017.01.013
3. Ahirwar R, Khan N, Kumar S. Aptamer-based sensing of breast cancer biomarkers: a comprehensive review of analytical figures of merit. Expert Rev Mol Diagn. 2021;21(7):703–721. doi:10.1080/14737159.2021.1920397
4. Bostanabad SY, Noyan S, Dedeoglu BG, et al. Overexpression of β-Arrestins inhibits proliferation and motility in triple negative breast cancer cells. Sci Rep. 2021;11(1):1539. doi:10.1038/s41598-021-80974-6
5. Zhou L, Rueda M, Alkhateeb A. Classification of breast cancer nottingham prognostic index using high-dimensional embedding and residual neural network. Cancers. 2022;14(4):934. doi:10.3390/cancers14040934
6. Zhang S, Zhang E, Long J, et al. Immune infiltration in renal cell carcinoma. Cancer Sci. 2019;110(5):1564–1572. doi:10.1111/cas.13996
7. Pyagay P, Heroult M, Wang Q, et al. Collagen triple helix repeat containing 1, a novel secreted protein in injured and diseased arteries, inhibits collagen expression and promotes cell migration. Circ Res. 2005;96(2):261–268. doi:10.1161/01.RES.0000154262.07264.12
8. Durmus T, LeClair RJ, Park KS, et al. Expression analysis of the novel gene collagen triple helix repeat containing-1 (Cthrc1). Gene Expr Patterns. 2006;6(8):935–940. doi:10.1016/j.modgep.2006.03.008
9. Yamamoto S, Nishimura O, Misaki K, et al. Cthrc1 selectively activates the planar cell polarity pathway of Wnt signaling by stabilizing the Wnt-receptor complex. Dev Cell. 2008;15(1):23–36. doi:10.1016/j.devcel.2008.05.007
10. Turashvili G, Bouchal J, Baumforth K, et al. Novel markers for differentiation of lobular and ductal invasive breast carcinomas by laser microdissection and microarray analysis. BMC Cancer. 2007;7:55. doi:10.1186/1471-2407-7-55
11. Ip W, Wellman-Labadie O, Tang L, et al. Collagen triple helix repeat containing 1 promotes melanoma cell adhesion and survival. J Cutan Med Surg. 2011;15(2):103–110. doi:10.2310/7750.2011.10014
12. Tang L, Dai DL, Su M, et al. Aberrant expression of collagen triple helix repeat containing 1 in human solid cancers. Clin Cancer Res. 2006;12(12):3716–3722. doi:10.1158/1078-0432.CCR-06-0030
13. Li LY, Yin KM, Bai YH, et al. CTHRC1 promotes M2-like macrophage recruitment and myometrial invasion in endometrial carcinoma by integrin-Akt signaling pathway. Clin Exp Metastasis. 2019;36(4):351–363. doi:10.1007/s10585-019-09971-4
14. Kim JH, Baek TH, Yim HS, et al. Collagen triple helix repeat containing-1 (CTHRC1) expression in invasive ductal carcinoma of the breast: the impact on prognosis and correlation to clinicopathologic features. Pathol Oncol Res. 2013;19(4):731–737. doi:10.1007/s12253-013-9636-y
15. Rhodes DR, Kalyana-Sundaram S, Mahavisno V, et al. Oncomine 3.0: genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia. 2007;9(2):166–180. doi:10.1593/neo.07112
16. Zhang X, Lv QL, Huang YT, et al. Akt/FoxM1 signaling pathway-mediated upregulation of MYBL2 promotes progression of human glioma. J Exp Clin Cancer Res. 2017;36(1):105. doi:10.1186/s13046-017-0573-6
17. Qu LP, Zhong YM, Zheng Z, et al. CDH17 is a downstream effector of HOXA13 in modulating the Wnt/β-catenin signaling pathway in gastric cancer. Eur Rev Med Pharmacol Sci. 2017;21(6):1234–1241.
18. Liu J, Li T, Liu XL. DDA1 is induced by NR2F6 in ovarian cancer and predicts poor survival outcome. Eur Rev Med Pharmacol Sci. 2017;21(6):1206–1213.
19. Forbes SA, Beare D, Boutselakis H, et al. COSMIC: somatic cancer genetics at high-resolution. Nucleic Acids Res. 2017;45(1):777–783. doi:10.1093/nar/gkw1121
20. Forbes SA, Beare D, Bindal N, et al. COSMIC: high-resolution cancer genetics using the catalogue of somatic mutations in cancer. Curr Protoc Hum Genet. 2016;91:10. doi:10.1002/cphg.21
21. Lánczky A, Nagy Á, Bottai G, et al. miRpower: a web-tool to validate survival-associated miRNAs utilizing expression data from 2178 breast cancer patients. Breast Cancer Res Treat. 2016;160(3):439–446. doi:10.1007/s10549-016-4013-7
22. Li T, Fan J, Wang B, et al. TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017;77(21):108–110. doi:10.1158/0008-5472.CAN-17-0307
23. Li B, Severson E, Pignon JC, et al. Comprehensive analyses of tumor immunity: implications for cancer immunotherapy. Genome Biol. 2016;17(1):174. doi:10.1186/s13059-016-1028-7
24. Aran D, Sirota M, Butte AJ. Systematic pan-cancer analysis of tumour purity. Nat Commun. 2015;6:8971. doi:10.1038/ncomms9971
25. Siemers NO, Holloway JL, Chang H, et al. Genome-wide association analysis identifies genetic correlates of immune infiltrates in solid tumors. PLoS One. 2017;12(7):e0179726. doi:10.1371/journal.pone.0179726
26. Danaher P, Warren S, Dennis L, et al. Gene expression markers of tumor infiltrating leukocytes. J Immunother Cancer. 2017;5:18. doi:10.1186/s40425-017-0215-8
27. Tameda M, Sugimoto K, Shiraki K, et al. Collagen triple helix repeat containing 1 is overexpressed in hepatocellular carcinoma and promotes cell proliferation and motility. Int J Oncol. 2014;45(2):541–548. doi:10.3892/ijo.2014.2445
28. Kim HC, Kim YS, Oh HW, et al. Collagen triple helix repeat containing 1 (CTHRC1) acts via ERK-dependent induction of MMP9 to promote invasion of colorectal cancer cells. Oncotarget. 2014;5(2):519–529. doi:10.18632/oncotarget.1714
29. Yang XM, You HY, Li Q, et al. CTHRC1 promotes human colorectal cancer cell proliferation and invasiveness by activating Wnt/PCP signaling. Int J Clin Exp Pathol. 2015;8(10):12793–12801.
30. Xu M, Liu M, Du X, et al. Intratumoral delivery of IL-21 overcomes anti-Her2/Neu resistance through shifting tumor-associated macrophages from M2 to M1 phenotype. J Immunol. 2015;194(10):4997–5006. doi:10.4049/jimmunol.1402603
31. Ni S, Ren F, Xu M, et al. CTHRC1 overexpression predicts poor survival and enhances epithelial-mesenchymal transition in colorectal cancer. Cancer Med. 2018;7(11):5643–5654. doi:10.1002/cam4.1807
32. Zhou F, Shen D, Xiong Y, et al. CTHRC1 is a prognostic biomarker and correlated with immune infiltrates in kidney renal papillary cell carcinoma and kidney renal clear cell carcinoma. Front Oncol. 2021;10:570819. doi:10.3389/fonc.2020.570819
33. Yuan H, Lin Z, Liu Y, et al. Intrahepatic cholangiocarcinoma induced M2-polarized tumor-associated macrophages facilitate tumor growth and invasiveness. Cancer Cell Int. 2020;20(1):586. doi:10.1186/s12935-020-01687-w
34. Gajeton J, Krukovets I, Muppala S, et al. Hyperglycemia-induced miR-467 drives tumor inflammation and growth in breast cancer. Cancers. 2021;13(6):1346. doi:10.3390/cancers13061346
35. Jiang N, Cui Y, Liu J, et al. Multidimensional roles of collagen triple helix repeat containing 1 (CTHRC1) in malignant cancers. J Cancer. 2016;7(15):2213–2220. doi:10.7150/jca.16539
36. Zhang SC, Hu ZQ, Long JH, et al. Clinical implications of tumor-infiltrating immune cells in breast cancer. J Cancer. 2019;10(24):6175–6184. doi:10.7150/jca.35901
37. Ruffell B, Coussens LM. Macrophages and therapeutic resistance in cancer. Cancer Cell. 2015;27(4):462–472. doi:10.1016/j.ccell.2015.02.015
38. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8(12):958–969. doi:10.1038/nri2448
39. Mantovani A, Sozzani S, Locati M, et al. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002;23(11):549–555. doi:10.1016/s1471-4906(02)02302-5
40. Martinez FO, Gordon S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep. 2014;6:13. doi:10.12703/P6-13
41. Lai YH, Chen J, Wang XP, et al. Collagen triple helix repeat containing-1 negatively regulated by microRNA-30c promotes cell proliferation and metastasis and indicates poor prognosis in breast cancer. J Exp Clin Cancer Res. 2017;36(1):92. doi:10.1186/s13046-017-0564-7
42. Zhao S, Mi Y, Guan B, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13(1):156. doi:10.1186/s13045-020-00991-2
43. Wang Y, Lyu Z, Qin Y, et al. FOXO1 promotes tumor progression by increased M2 macrophage infiltration in esophageal squamous cell carcinoma. Theranostics. 2020;10(25):11535–11548. doi:10.7150/thno.45261
44. Wang X, Jiao X, Meng Y, et al. Methionine enkephalin (MENK) inhibits human gastric cancer through regulating tumor associated macrophages (TAMs) and PI3K/AKT/mTOR signaling pathway inside cancer cells. Int Immunopharmacol. 2018;65:312–322. doi:10.1016/j.intimp.2018.10.023
45. Mei D, Zhu Y, Zhang L, et al. The role of CTHRC1 in regulation of multiple signaling and tumor progression and metastasis. Mediators Inflamm. 2020;2020:9578701. doi:10.1155/2020/9578701
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.