")
Back to Journals » Journal of Pain Research » Volume 12
CSF levels of apolipoprotein C1 and autotaxin found to associate with neuropathic pain and fibromyalgia
Authors Lind AL, Just D , Mikus M , Fredolini C, Ioannou M, Gerdle B , Ghafouri B , Bäckryd E , Tanum L , Gordh T, Månberg A
Received 11 May 2019
Accepted for publication 1 August 2019
Published 15 October 2019 Volume 2019:12 Pages 2875—2889
DOI https://doi.org/10.2147/JPR.S215348
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Katherine Hanlon
Anne-Li Lind,1 David Just,2 Maria Mikus,2 Claudia Fredolini,2 Marina Ioannou,2 Björn Gerdle,3 Bijar Ghafouri,3 Emmanuel Bäckryd,3 Lars Tanum,4 Torsten Gordh,1 Anna Månberg2
1Department of Surgical Sciences, Uppsala University, Uppsala, Sweden; 2Division of Affinity Proteomics, SciLifeLab, Deptartment of Protein Science, KTH Royal Institute of Technology, Stockholm, Sweden; 3Pain and Rehabilitation Center, and Department of Medical and Health Sciences, Linköping University, Linköping, Sweden; 4Department of R&D in Mental Health, Akershus University Hospital, Lørenskog, Norway
Correspondence: Anna Månberg
Division of Affinity Proteomics, SciLifeLab, Department of Protein Science, KTH - Royal Institute of Technology, Tomtebodvägen 23A, Solna, Stockholm, Sweden
Tel +46 8 790 9803
Email [email protected]
Objective: Neuropathic pain and fibromyalgia are two common and poorly understood chronic pain conditions that lack satisfactory treatments, cause substantial suffering and societal costs. Today, there are no biological markers on which to base chronic pain diagnoses, treatment choices or to understand the pathophysiology of pain for the individual patient. This study aimed to investigate cerebrospinal fluid (CSF) protein profiles potentially associated with fibromyalgia and neuropathic pain.
Methods: CSF samples were collected from 25 patients with neuropathic pain (two independent sets, n=14 patients for discovery, and n=11 for verification), 40 patients with fibromyalgia and 134 controls without neurological disease from two different populations. CSF protein profiling of 55 proteins was performed using antibody suspension bead array technology.
Results: We found increased levels of apolipoprotein C1 (APOC1) in CSF of neuropathic pain patients compared to controls and there was a trend for increased levels also in fibromyalgia patients. In addition, levels of ectonucleotide pyrophosphatase family member 2 (ENPP2, also referred to as autotaxin) were increased in the CSF of fibromyalgia patients compared to all other groups including patients with neuropathic pain.
Conclusion: The increased levels of APOC1 and ENPP2 found in neuropathic pain and fibromyalgia patients may shed light on the underlying mechanisms of these conditions. Further investigation is required to elucidate their role in maintaining pain and other main symptoms of these disorders.
Keywords: cerebrospinal fluid, neuropathic pain, fibromyalgia, antibody suspension bead arrays, APOC1, ENPP2
Introduction
Pain conditions such as fibromyalgia and neuropathic pain cause substantial suffering,1 disability,2 and great societal costs.3 In addition, they are difficult to treat4,5 and sometimes difficult to diagnose.6–8 Progress has been made in clinical classification and diagnostic criteria for neuropathic pain 9,10 and fibromyalgia,11,12 but there is a need for better understanding of the pathophysiology and for more effective treatments.13–17 Currently, there are no biological tests on which to base pain diagnoses, treatment choices or to understand the pathophysiology of the individual pain patient. Such markers that reflect the pathophysiology of individual pain patients would be important tools for pain clinicians, scientists, and pharmaceutical companies to aid in diagnosis, treatment selection, and to guide and monitor the development of new treatments. Although cerebrospinal fluid (CSF) collection is an invasive procedure, CSF is in direct contact with the brain and spinal cord and changes in CSF protein levels may reflect pathological processes in the central nervous system.18
Neuropathic pain is often described as a particularly unpleasant form of pain19 with shooting, shock-like, aching, cramping, crushing, smarting, and burning features.20 Neuropathic pain is caused by lesion or disease of the somatosensory nervous system 21 and affects 1–10% of the general population.22–29 Previous investigations of CSF from neuropathic pain patients have typically analyzed one or a few interesting markers (mainly proteins) in small sample cohorts. Several studies have found differences in the levels of one or more proteins including inflammatory markers between neuropathic pain patients and controls30–35 while other studies showed no significant differences.36,37
Fibromyalgia affects around 2% of the population and is characterized by widespread pain and generalized hyperalgesia for mechanical pressure.38 Fibromyalgia patients often suffer from psychological distress, sleep and memory disturbances, and fatigue.38 There are many theories behind pathophysiology of FM, but the etiology is still uncertain. The current view is that the clinical presentation of fibromyalgia depends on central phenomena rather than peripheral dysfunction and substantial evidence exists for abnormalities in sensory signaling, including reduction of descending control and changes in key neurotransmitters associated with central sensitization.10 However, altered levels of cytokines, anti-inflammatory lipids, and prominent alterations both in muscle tissue and circulating proteins have been reported39–44 as well as small nerve fiber impairment in FM.45 A wide range of proteins including inflammatory markers were found altered in the CSF of fibromyalgia patients.46–53
Biomarker profiles that can be used to characterize similarities and differences between chronic pain conditions would be valuable for understanding the pathophysiological mechanisms and give new leads for treatment development. In a recent mass spectrometry (MS) investigation of CSF samples, we demonstrated altered levels of several proteins associated with satisfactory spinal cord stimulation (SCS) treatment.54 Here, we applied the antibody suspension bead array technology that offers a flexible platform for parallel protein detection using only 15 µL of crude biological sample. It has previously been used to study CSF and plasma within other neurological diseases such as multiple sclerosis, amyotrophic lateral sclerosis, and Alzheimer’s disease.55–57 In the present study, we identified CSF proteins associated with pain pathophysiology by comparing patients with neuropathic pain and fibromyalgia to CSF from controls without chronic pain.
Methods
Subjects
In this study, we analyzed a total of 199 CSF samples from neuropathic pain patients, fibromyalgia patients, and two types of controls (Table 1). The first set of neuropathic pain patients (denoted NP1) included 14 individuals that were recruited from Uppsala University Hospital (mean age 57 (47–68), four males). They suffered from long-lasting neuropathic pain (median 10 years, range 3–23 years) and had permanently implanted spinal cord stimulation (SCS) since more than 3 months (median 3 years, range 1–10 years) with self-reported good pain relief. For these patients, samples were collected both when the stimulation was turned on and off.54
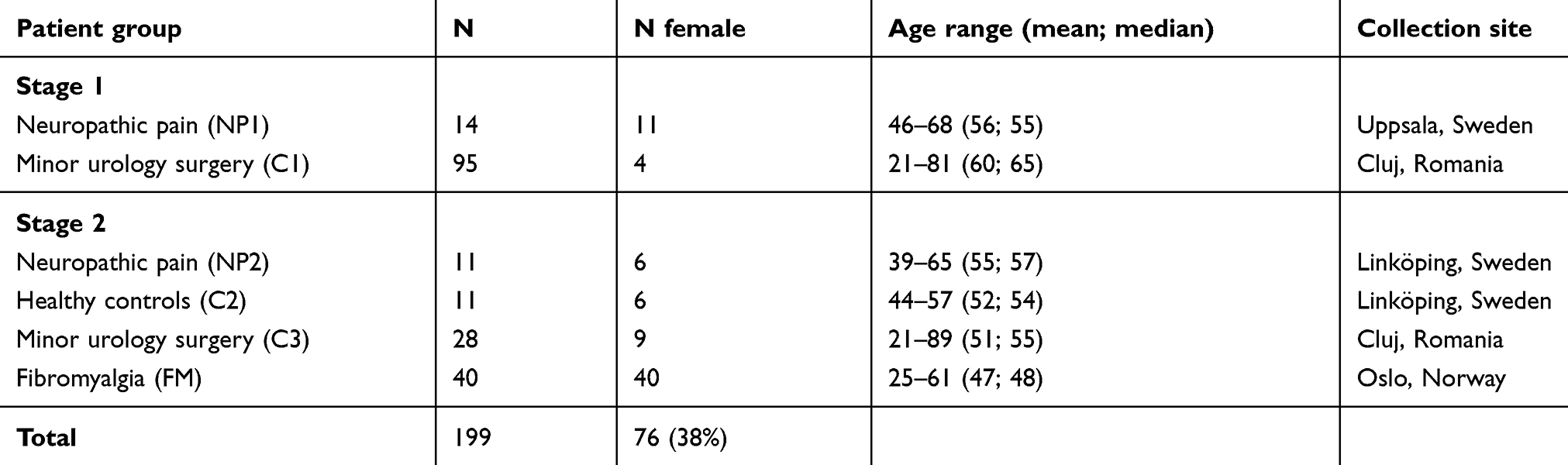 |
Table 1 Patient and experimental overview |
The second set of patients (denoted NP2) were 11 adults suffering from chronic (≥6 months) neuropathic pain due to trauma or surgery. These individuals participated in a clinical trial of intrathecal bolus injections of the analgesic ziconotide (ClinicalTrials.gov identifier NCT01373983), the mean age was 55 years (range 39–65) and six of the individuals were females. All samples were collected prior treatment at Linköping University Hospital in Sweden.
The fibromyalgia (FM) patients included in the study were 40 females (ages 20–60) suffering from FM as defined by the 1990 criteria of the American College of Rheumatology (ACR). Patient recruitment and sampling were performed at the privately operated Red Cross Hospital in Olso, Norway.
To find disease-associated proteins, the samples from the patients were compared to samples from two sets of controls. The largest control group consisted of 123 individuals undergoing spinal anesthesia for minor urological surgery at Cluj Hospital, Romania (C1 and C3, see Study design). Samples were collected according to the same protocol as in Uppsala, Sweden and although comparable in age, these individuals were mostly males. The other control cohort (C2) consisted of healthy volunteers matched to the NP2 patients and was collected simultaneously as the patients in Linköping, Sweden.
The sample collection procedures of all samples, from patient recruitment via lumbar puncture to aliquoting and storage, are detailed in Supplementary methods.
Study design
The experiments in this study were performed in two stages. The first stage of analysis included the NP1 patients from Uppsala, Sweden (n=14) and a subset of the Romanian controls (denoted C1, n=95). In the second stage, we analyzed the independent cohort of neuropathic pain CSF samples (NP2, n=11) and the samples from fibromyalgia patients (FM, n=40). These were compared to control samples from the healthy volunteers (C2, n=11) and the 28 additional controls from the Romanian cohort (denoted C3).
Creating the bead array
A bead microarray was customized and created using 100 antibodies from the Human Protein Atlas project (www.proteinatlas.org). These antibodies targeted 55 unique proteins that were selected based on an earlier mass spectrometry investigation of protein levels associated with satisfactory SCS treatment,54 see Table S1. Immobilization of antibodies to beads was performed as previously described58 with one specific antibody assigned to a certain bead identity.
CSF analysis
CSF samples were processed and analyzed through a direct labeling approach as previously reported.55 In short, CSF samples were diluted 1/2 in a protein-containing buffer (37.5 mg/mL bovine serum albumin (Sigma-Aldrich) in PBS with addition of 15 mg/mL rIgG (Bethyl Laboratories Inc., Montgomery, Texas, USA)) and a labeling solution with 10x molar excess of biotin (NHS-PEG4-Biotin, Thermo Scientific) over total protein amount, to a final volume of 30 µL. The reaction was terminated by addition of 250x Tris-HCl over biotin amount. Before applying samples to the bead array, they were further diluted one-eighth in assay buffer (PVXCas) to an end volume of 50 µL then heat treated at 56°C for 30 mins in a water bath. Following cooling to ambient temperature for 15 mins, 45 µL of sample was combined with 5 µL of the bead array solution and incubation was performed overnight. After washing off unbound proteins with 3×100 µL PBST (0.05% Tween-20), the interacting proteins were cross-linked during 10 mins using paraformaldehyde (0.4% in PBS). A second wash was followed by addition of detection reagent (streptavidin-conjugated R-phycoerythrin, Invitrogen, 1:750 diluted in PBST) and incubation was performed for 20 mins. Finally, the interacting proteins were analyzed using a FlexMap3D instrument (Luminex Corp., Austin, Texas, USA) and the relative abundance reported as median fluorescent intensities per sample and bead identity.
Verification of protein identities by immunocapture mass spectrometry (IC-MS)
Assessment of antibody selectivity was performed by IC-MS as previously described.57 Briefly, antibodies coupled to magnetic beads were incubated with CSF from pooled human donors (Lee Biosolutions, Maryland Heights, Missouri, USA) diluted in assay buffer 1:10. After incubation, beads were washed and proteins captured were reduced, alkylated, and digested using trypsin and lysC (Promega, Madison, Wisconsin, USA). Peptides were re-suspended in buffer A (3% acetonitrile (ACN), 0.1% formic acid (FA) and separated on the Ultimate 3000 Rapid Separation Liquid Chromatography (RSLC) nanosystem (Dionex, Sunnyvale, California, USA). The analytical column was a 50 cm × 75 μm ID Easy spray (PepMap RSLC C18, Thermo Fisher). A linear gradient of Buffer B (90% ACN, 5% DMSO, 0.1% FA) was run in 50 mins at 250 nL/min (from 3% to 43%). MS analysis was performed using a Q-Exactive HF (Thermo Fisher) operated in a data-dependent mode with higher energy collision dissociation (HCD) on the top five most abundant ions. Full MS scan (300–1600 m/z) was performed at a resolution of 60,000. Database searching of raw data was performed on MaxQuant. Search parameters were set as in 57 allowing for match between runs and label-free quantification using the MaxLFQ algorithm integrated into MaxQuant.59 Proteins identified in each IC-MS were compared with IC-MS performed for unrelated antibodies. Z-scores were calculated to rank the proteins enriched by each antibody and plotted against LFQ intensities.
Statistical analysis
The intensities obtained from the antibody suspension bead array analysis were subjected to statistical analysis and visualization using R Studio (version 1.2.1335).60 Comparisons of protein levels were made within the same experimental stage only, to avoid any contribution to the results caused by inter-plate variation. For a general data overview, a principal component analysis (PCA, R package stats) of all groups was performed to assess general data quality and to remove potential outliers from further data analysis.
Multivariate data analysis was performed by projection (MVDA) using SIMCA-P+, version 13.0 (Umetrics AB, Umeå, Sweden).32,61 Data were log-transformed when needed (using the transform function in SIMCA-P+) and scaling was done according to the unit variance method. Outlier detection for the multivariate model was performed using Hotelling’s T2 (T2 Critical 99%) for strong outliers and by distance to model in X-space (DModX) for serious moderate outliers. Then, orthogonal partial least squares – discriminant analysis (OPLS-DA) was used to identify class-discriminating proteins. The significance of OPLS-DA models was assessed by analysis of variance of cross-validated predictive residuals (CV-ANOVA), and the relative contribution of the proteins for group discrimination was determined by the predictive loadings and by the corresponding loadings scaled as a correlation coefficient, denoted p(corr), between the model and original data. An absolute p(corr)≥0.5 was considered significant.62
For univariate analysis, statistical significance of group differences was evaluated by the Wilcoxon rank sum test (R package stats) and p-values below 0.05 after multiple testing correction (FDR) were regarded as significant. To select proteins of interest, the assays of the first stage were repeated twice and proteins of interest selected based on concordant significant differences.
Results
In this study, we have performed CSF profiling using an affinity proteomics approach in order to investigate 55 proteins with potential relevance for pain pathophysiology and pain treatment. Our sample set consisted of in total 199 CSF samples from 65 patients and 134 controls (Table 1). In the first experimental stage, we analyzed CSF from 14 patients with neuropathic pain on and off SCS treatment (NP1) and 95 controls without pain pathology that underwent minor urological surgery (C1). To validate the results, an independent set of CSF samples from patients with neuropathic pain (NP2) were analyzed together with matched controls (C2) as well as additional control subjects from the Romanian site (C3).
Multivariate protein signatures
To explore disease-associations in a multivariate manner, we performed OPLS-DA analysis of the complete datasets generated in both stage I and stage II. In the first stage, 10 outliers (eight controls and two patients) were removed after initial PCA, rendering a final PCA model with four latent variables, R2=0.60, and Q2=0.52. Then, an OPLS-DA model was computed to discriminate between NP1 and C1 (referred to as model 1, four latent variables, R2=0.85, Q2=0.67, and p<0.001) and the proteins with significant contribution to the model (|p(corr)| ≥0.5) were apolipoprotein C-I (APOC1), keratin 1 (KRT1), apolipoprotein A1 (APOA1) and neurexin-1 (NRXN1) (Figure 1 and Table S2A). It was not possible to generate an OPLS-DA model comparing neuropathic pain patients to healthy controls from the same center (NP2 vs C2), indicating subtle differences in protein levels between groups. Nevertheless, models were calculated for NP2 compared to C3 (referred to as model 2, five latent variables, R2=0.98, Q2=0.86, and p-value<0.001) with 14 proteins contributing significantly (Table S2B). When comparing the results from model 1 and 2, the protein with significant contribution in both models was APOA1. Although not significant, APOC1, KRT1, and NRXN1 were just under the significance threshold with |p(corr)| of 0.38, 0.44, and 0.49, respectively.
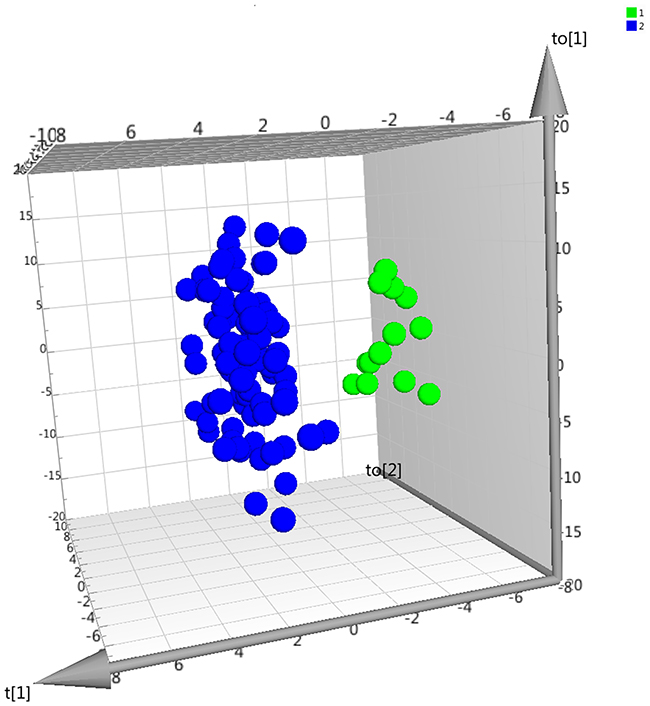 |
Figure 1 Score plot of multivariate model discriminating neuropathic pain patients and controls. Visualization of the first three latent variables from OPLS-DA model 1, neuropathic pain patients (NP1, green dots) vs controls (C1, blue dots). The t[1] axis represents interclass variation, whereas the two other two axes (to[1] and to[2]) represent intra-class variation. |
For fibromyalgia, one patient sample was identified as an outlier and removed. Two OPLS-DA models were then computed, one comparing the fibromyalgia patients to C2 (referred to as model 3 with five latent variables, R2=0.95, Q2=0.75, p-value <0.001 and C3 (referred to as model 4 with five latent variables, R2=0.94, Q2=0.80, p-value<0.001). Model 3 were based on seven significantly contributing proteins (Table S2C), but in model 4 only autotaxin (or ectonucleotide pyrophosphatase/phosphodiesterase family member 2, ENPP2) contributed significantly.
Increased levels of APOC1 in neuropathic pain patients
In the first analysis stage, we observed no significant differences between samples collected from patients on and off SCS treatment using univariate statistics (data not shown). In the comparison between patients and controls, the two proteins APOC1 and secretogranin-1 (CHGB also referred to as SCG1) were found at significantly higher levels in the CSF of neuropathic pain patients off SCS treatment (NP1) compared to controls (C1). In addition, NRX1, APOA1, dickkopf-related protein 3 (DKK3), superoxide dismutase 1 (SOD1), complement factor H (CFH), prostaglandin-H2 D-isomerase (PTGDS), complement component C7 (C7), and hemopexin (HPX) were observed at lower levels in the patients (p-value<0.05, Table 2).
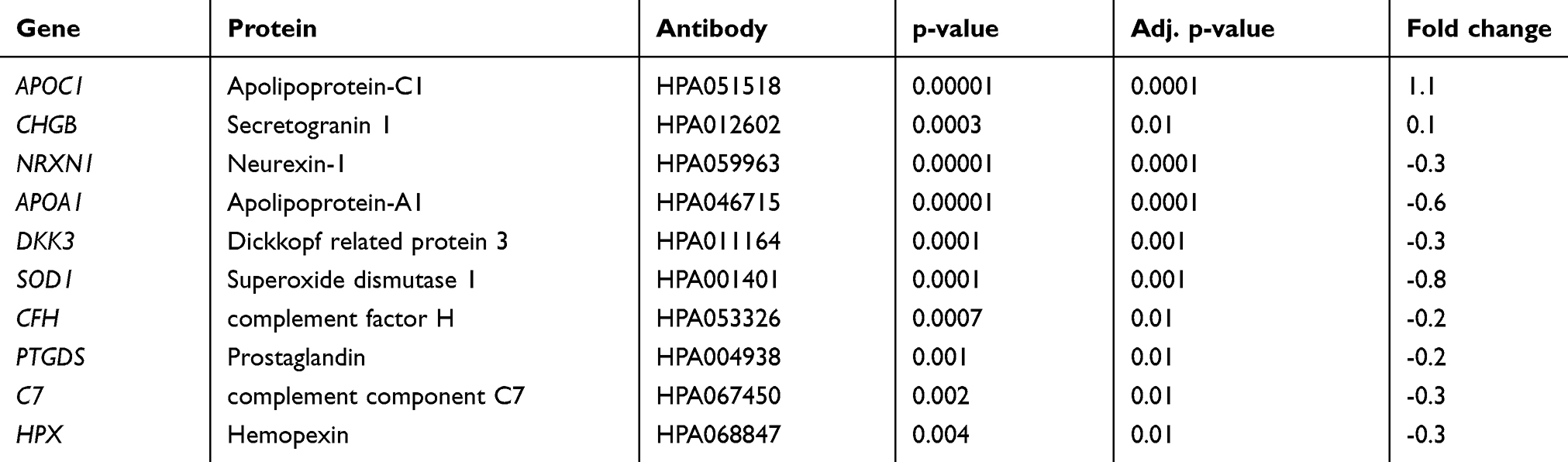 |
Table 2 Proteins with significantly altered levels in neuropathic pain patients |
In the second stage analysis, none of the 10 proteins showed significant differences between the NP2 patients and both control groups although APOC1 (detected by antibody HPA051518) showed near significant and concordant trends with higher CSF levels in patients compared to both C2 and C3 (Figure 2, uncorrected p-values 0.3 and 0.05, respectively). When comparing the pain patients and controls collected at the same site (NP2 and C2), the protein osteoglycin (OGN) was significantly altered before correction for multiple testing (uncorrected p-value 0.006). However, this protein was found at similar levels in patients and controls in the comparison of NP1 and C1 (p-value 0.8).
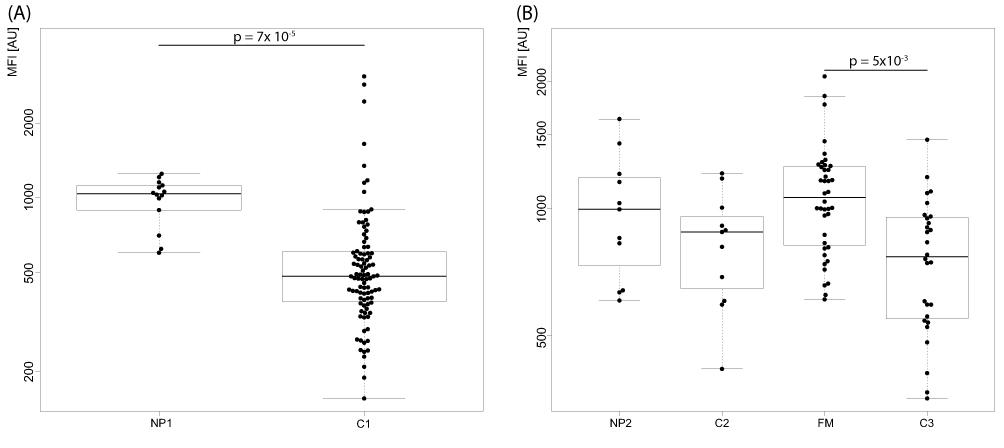 |
Figure 2 CSF levels of APOC1 in (A) neuropathic pain patients (NP1) compared to controls (C1) in the first stage and (B) for neuropathic pain patients (NP2) and fibromyalgia (FM) compared to controls (C2 and C3) in the second analysis stage. Significant differences are indicated with p-values. MFI – median fluorescence intensity. |
Increased levels of ENPP2 in fibromyalgia patients
To explore proteins with potential association to fibromyalgia pathology, profiles of fibromyalgia patients (FM) were compared to the two separate control groups analyzed in the second stage (C2 and C3). Summarizing the results, APOC1 and ENPP2 (antibody HPA053652) were the two proteins with concordant differences in patients compared to the control groups, both observed with higher levels in the patients (Figures 2 and 3). For APOC1, the difference was statistically significant in the comparison with C3 (p-value 0.005) while comparison to C2 was not significant after p-value correction (uncorrected p-value 0.03). The levels of ENPP2, on the other hand, were significantly higher in patients compared to both control groups (0.0001 for FM vs C2 and 0.000003 for FM vs C3). Levels of ENPP2 were also increased in fibromyalgia patients compared to those with neuropathic pain (NP2, p-value 0.0000003)
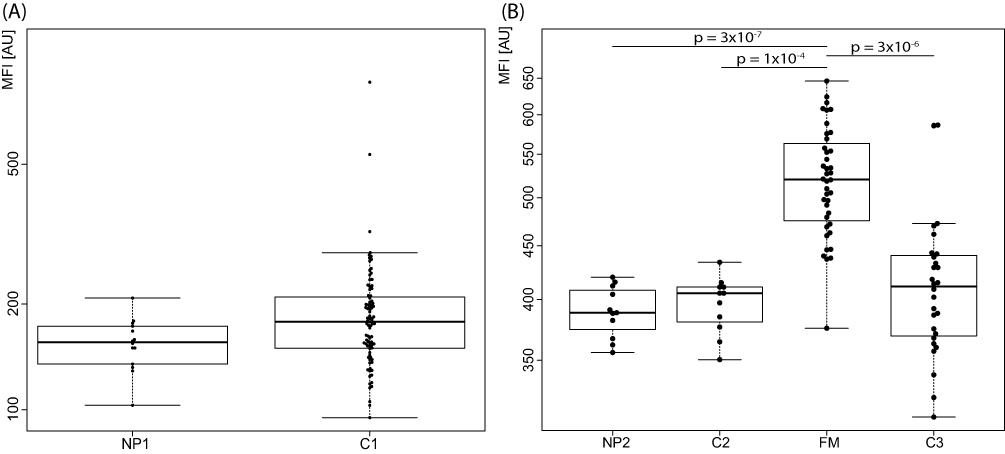 |
Figure 3 CSF levels of ENPP2 in (A) neuropathic pain patients (NP1) compared to controls (C1) in the first stage and (B) for neuropathic pain patients (NP2) and fibromyalgia (FM) compared to controls (C2 and C3) in the second analysis stage. Significant differences are indicated with p-values. MFI – median fluorescence intensity. |
Antibody validation by IC-MS
We applied immunocapture mass spectrometry (IC-MS) in order to verify the selectivity of the antibodies targeting APOC1 (HPA051518) and ENPP2 (HPA053652). Both antibodies captured and enriched their intended target when incubated with CSF from healthy donors (Figure S1). APOC1 was the protein mostly enriched by HPA051518 (z-score 2.59, LFQ intensity 1,765,450,000) but also APOA1 and apolipoprotein E (APOE) were detected. ENPP2 was identified as captured by the HPA053652 antibody (z-score 2.60, LFQ Intensity: 54,462,500) together with the additional proteins Collagen alpha-3(VI) chain (COL6A3), Myosin heavy chain 7 (MYH7), and prothrombin (F2) (Figure S1). See Table S3 for all Z-scores and LFQ intensities.
Discussion
In this study, we performed protein profiling of human CSF samples. We measured levels of 55 proteins previously identified to have a potential role in pain treatment. Our study material consisted of samples from 199 individuals of which 25 were patients with neuropathic pain, 40 patients with fibromyalgia and 134 controls.
Increased levels of APOC1 in CSF of neuropathic pain patients
The analysis of the first neuropathic pain cohort and controls demonstrated that APOC1 levels were significantly increased in patients. We also found a non-significant increase of CSF APOC1 in the verification sample set as well as an increase of APOC1 in fibromyalgia patient samples compared to both control groups.
The source of alterations in APOC1 levels may be central or peripheral. Peripheral APOC1 is mainly synthesized in the liver, circulates in the blood bound to lipoprotein particles and is involved in cholesterol and lipid homeostasis.63 APOC1 deriving from the central nervous system is produced by astrocytes, microglia, and endothelial cells.64 Less is known about the role of apolipoproteins in the central nervous system (CNS).65 The blood-brain barrier (BBB) and the blood-spinal cord barrier (BSCB) limit the exchange of lipoprotein particles between the blood and the CNS, but HDL-like particles are able to cross. Thus, APOC1 CSF level alterations such as reported here may be caused by APOC1 having traversed CNS barriers in HDL-like particles as part of a normal cross-barrier communication or through “leakier” brain or spinal cord barriers induced by nerve injury.66,67 A recent study of rodents suggested that the lumbar spinal cord is the source of increased serum levels of apolipoproteins after nerve injury.68 This suggests that even local spinal cord increases of apolipoproteins can be reflected not only in CSF but also in the blood a long time after injury. This further suggests that apolipoprotein levels in serum or plasma could be suitable as markers of ongoing pain relevant processes in the spinal cord.
We have recently found signs of inflammation in the CSF of neuropathic pain patients 69 as well as in the serum of patients with sciatic pain one year after disc hernia.70 APOC1 can modulate inflammatory processes71 and may play a role in chronic inflammation.72–75 APOC1 enhances the toll-like receptor 4 (TLR4)-dependent inflammatory response75,76 that induce production of pro-inflammatory cytokines and reactive oxygen species,77,78 and is an important part of glial activation in rodent models of neuropathic pain.77,79–82 Neuropathic pain-like behaviors can be reversed by intrathecal injection of TLR4 antagonists.83 TLR4 can be activated by endogenous ligands,84 cell products released upon injury85 and in neuropathic pain models, additional cofactors are required to enhance TLR4-dependent signaling.86 Since the BSCB is more permeable after nerve injury 67 it may allow entry of several such ligands. Thus, we hypothesize that the high levels of APOC1 we find in neuropathic pain patient CSF in this study may contribute to increased TLR4 activity which can explain parts of the prolonged glial activation and signs of central neuroinflammation seen in neuropathic pain conditions.53 Interestingly, TLR4 antagonists, as well as small molecule modulators, are being investigated as drug candidates for pain treatment.87 but paradoxically, APOC1 has also been found to be immunosuppressive.88 These apparently contradicting results are not fully understood.
Another interpretation is that the increased levels of APOC1 observed reflect an ongoing CNS degeneration and regeneration in persistent neuropathic pain conditions. Apolipoproteins are known to contribute to nerve cell regeneration by local delivery of lipids and other unknown mechanisms.68 Lipid metabolism is central to CNS homeostasis, especially due to myelin which is required for optimal neuronal and network function. Several studies have shown that APOC1 and APOE are closely linked both genetically 89,90 and functionally.91 Levels of other apolipoproteins have recently been shown to be altered in neuropathic and discogenic pain patient CSF compared to controls32,92 as well as after nerve injury in rat models.68,93 In the present investigation, APOE levels were not found to be significantly altered in fibromyalgia or neuropathic pain, although there are at least four previous studies linking APOE gene variants and APOE levels to risk of persistent pain.94–97 To our knowledge, there are no previously published results directly linking APOC1 to persistent pain or its treatments.
Increased levels of ENPP2 in CSF of patients with fibromyalgia
The ENPP2 protein was found to be significantly increased in CSF from patients with fibromyalgia when compared to both groups of controls as well as patients with neuropathic pain. To our knowledge, a direct relation between ENPP2 and fibromyalgia has not been reported previously, but ENPP2 inhibitors have been patented for treatment of fibromyalgia.98 ENPP2 has also been demonstrated to increase in CSF from patients with chronic fatigue syndrome (CFS),99 a condition that shares characteristics with fibromyalgia. There are also reports of important pain modulating the activity of two of the main products of ENPP2.100
ENPP2 is an enzyme with lysophospholipase D activity that catalyzes the formation of the lipid mediator lysophosphatidic acid (LPA) from lysophosphatidylcholine.101 A recent publication states that LPA signaling is the mechanism behind neuropathic pain and also refers to unpublished observations that LPA signaling appears to play a crucial role in fibromyalgia.102 Under normal conditions, LPA promotes healing after tissue trauma and negatively regulates ENPP2 transcription. In a high cytokine environment, LPA can induce ENPP2 expression to increase overall LPA concentrations in a positive feedback loop.103 LPA is an important pro-inflammatory mediator in the spinal cord. Neuroinflammation can cause neurodegeneration104 and it has been reported that blocking LPA can improve recovery after spinal cord injury as LPA has been shown to cause nerve demyelination.105,106 Distal demyelinating polyneuropathy has been reported in a subset of fibromyalgia patients107 and at least six studies have reported peripheral small fiber abnormalities in fibromyalgia patients.108–113 Heterozygous mutant mice (ENPP2+/−) display a 50% decrease in ENPP2 activity, reduced LPA production and, interestingly, better recovery from neuropathic pain-like behaviors after injury.114–116 Therefore, ENPP2 has been of interest as an upstream target of LPA and for the treatment of neuropathic pain.117 It is noteworthy that we observed no increase of ENPP2 in neuropathic pain patients; however, the present investigation does not contain any information of potential changes in ENPP2 activity levels or production of its metabolites.
ENPP2 has a functional domain118 which regulates oligodendrocyte process outgrowth and branching.119–121 ENPP2 can also act on sphingosylphosphorylcholine to produce sphingosine-1-phosphate (S1P)122 which has been found to modulate spinal nociceptive processing123 and excitability of dorsal root ganglion sensory neurons.124,125 Concentrations of CSF S1P have also been reported as decreased in an animal model of acute and inflammatory pain.126 Neuronal S1P1 receptors play an important role in nociceptor regulation and S1P signaling is a suspected mediator of the onset of thermal hypersensitivity and hyperalgesia associated with inflammation.127 Central S1P has been shown to have anti-nociceptive effects, but there appear to be many points of regulation and the interactions are not fully understood as reviewed by others.128 An untargeted metabolomics investigation of a rodent neuropathic pain model suggests that altered levels of sphingolipids are part of the chronic pain pathophysiology.129
A hypothesis that can be generated from our result is that the high levels of ENPP2 indicate elevated levels of LPA or S1P, mediators which were not measured in this study. If so, these inflammatory agents could damage or alter activity of components of the patient’s nervous tissue and contribute to their pain and other symptoms. Because CFS patients were also reported to have high ENPP2 levels,99 another hypothesis is that ENPP2 levels somehow reflect the impaired sleep quality commonly described in both CFS and fibromyalgia. It could also be so that increased levels of ENPP2 are a secondary effect due to alterations in other factors regulating the clearance of proteins.130
Target identification by IC-MS
IC-MS analysis was performed to evaluate the selectivity of antibodies targeting APOC1 and ENPP2 and in both cases, the intended targets were detected. The additional proteins present may represent competitors (off-target interactors) or proteins in a complex with the target and so co-immunoprecipitated with it. The alignment of the three sequences APOC1, APOA, APOE performed by CLUSTALO shows a low identity 1.52%. APOC1-APOA1: 7.72%. APOC1-APOE: 4.42% between APOC1 and the two apolipoproteins. This would poorly support an off-target binding while the hypothesis that APOC1 may be captured in a complex with APOA and APOE is partially supported by literature.131–133
Even though several other proteins appear to be enriched by the ENPP2 antibody with similar z-score values, only ENPP2 and PLG were exclusively identified in the IC-MS experiment performed using HPA053652. The low LFQ intensity associated with PLG and also to the other proteins indicate that they may represent contaminant proteins. As shown in other studies,134 several contaminant proteins are identified in immunoprecipitation experiments and only the screening of a higher number of negative control and replicates allows discrimination of co-immunoprecipitating proteins from background. In this experimental setting, results from only 16 different IC-MS experiments have been compared (data not shown).
In conclusion, the data of IC-MS are supportive for the validation of the two antibodies targeting APOC1 and ENPP2 used in the bead array screening, and further studies would be required to investigate if they are captured in a complex with other proteins.
Study limitations
CSF more closely mirrors the microenvironment of the central nervous system than plasma. However, lumbar puncture involves a medical risk for study subjects and access to both patient and healthy control CSF material is limited.
We have analyzed samples from multiple clinical locations and countries (Sweden, Norway, and Romania). Pre-analytical parameters such as collection tube type, sample volume, time from collection to centrifugation, freezing cycles, and storage temperature can affect the detectable proteome135,136 as can geographical differences and population lifestyle. In order to reduce the contribution of these factors to variation, the same CSF sample collection and handling protocol were used in Sweden and Romania and the person performing the lumbar punctures for the control samples in Cluj, Romania was trained with the clinicians in Uppsala, Sweden. Despite this, we did observe differences in protein profiles between samples from the different clinics. Therefore, we included additional control groups, compared the control groups to each other and filtered out differences derived from sampling site. There was a difference in both centrifugation protocol and sample freezer storage time between the Norwegian sample set and the others of 15 years. Enroth et al, analyzed plasma samples with a difference of 26 years in sample storage time and reported that for 10% of the proteins, sample storage time could explain 5–35% of the observed variance in protein level.137 It is not clear if ENPP2 belongs to the group of proteins that are affected by sample storage time, or how storage time effects in CSF differ from that of plasma, as plasma contains 100–200 times more protein than CSF.
There was also a gender bias among the included subjects. The sample set from minor urology surgery patients naturally contains more males than females. All the fibromyalgia CSF samples were from female patients, yet they were compared to samples from mixed sex groups. Sex has not proven to influence CSF proteins in our previous investigations with the exception of the expected difference in prostate-specific antigen.138 Nevertheless, to address this potential bias we performed group comparison analysis with only female participants and the data generated in stage II. The results showed that the levels of ENPP2 were significantly higher also in patients compared to both the female neuropathic pain patients and the female controls (Figure S2). As is typical in persistent pain, most patients in the study were on medications.2 Although interference cannot be ruled out, there are indications from other studies that drugs do not necessarily interfere with the protein profile in CSF50 or on glial activation in patients.139
Conclusion
We found increased levels of APOC1 in CSF of both patients with neuropathic pain and fibromyalgia while increased levels of ENPP2 were found only in the CSF of patients with fibromyalgia. Available literature supports the involvement of lipids in both neuropathic pain and fibromyalgia pathophysiology and supports further investigation of these proteins and their mediators to determine their potential role in human pain pathophysiology.
Ethics approval and informed consent
The study was conducted in accordance with the Declaration of Helsinki and approved by the Regional Ethical Review Boards of Uppsala and Linköping (Dnr M136-06 and Dnr 2012/94-32) Sweden, Regional Committee for Medical Research Ethics, South-East Health Region, Norway, and Cluj, Romania, respectively, and undertaken with the informed written consent of the individual donors.
Abbreviations
ACN, acetonitrile; APOA1, apolipoprotein A1; APOC1, apolipoprotein C-I; APOE apolipoprotein E; BBB, blood-brain barrier; BSCB, blood-spinal cord barrier; C7, complement component C7; CFH, complement factor H; CFS, chronic fatigue syndrome; CNS, central nervous system; COL6A3, collagen alpha-3(VI) chain; CSF, cerebrospinal fluid; DKK3, dickkopf related protein 3; ENPP2, autotaxin; F2, prothrombin; FA, formic acid; HPA, Human Protein Atlas; HPX, hemopexin; IC-MS, immunocapture mass spectrometry; LPA; lysophosphatidic acid; MS, mass spectrometry; MYH7, myosin heavy chain 7; NRX1, neurexin-1; OGN, osteoglycin; OPLS-DA, orthogonal partial least squares – discriminant analysis; PCA, principal component analysis; PTGDS, prostaglandin-H2 D-isomerase; S1P, sphingosine-1-phosphate; SCG1, secretogranin-1; SCS, spinal cord stimulation; SOD1, superoxide dismutase 1; TLR4, toll-like receptor 4.
Data sharing statement
Access to data as well as other documents will be considered on a case-by-case basis through the corresponding author.
Acknowledgment
This work was performed as a collaboration between Uppsala Berzelii Technology Centre for Neurodiagnostics and SciLifeLab Stockholm and was financed by the Swedish Governmental Agency for Innovation Systems, the Swedish Research Council (Vetenskapsrådet), Uppsala University, KTH Center for Applied Precision Medicine (KCAP), the Marie-Curie fellowship from the INSENS/FP7-PEOPLE-2013 (607616) framework, a grant from NEURO Sweden, AFA Insurance and ALF Grants, Region Östergötland. Constantin Bodolea and Lenka Katila are acknowledged for contributions in sample collection. Anders Wåhlstedt, Eva Leljewahl and co-workers are acknowledged for SCS patient recruitment and Peter Nilsson for many fruitful discussions. Finally, the authors would like to thank the entire staff of the Human Protein Atlas Project for their efforts. Anne-Li Lind and David Just are shared first co-authors.
Author contributions
TG, ALL planned and designed the clinical part of the SCS study. LK, LT, BG, and CB collected clinical data, pain, and control samples. ALL biobanked and aliquoted the samples. AM, DJ, and PN designed antibody suspension bead array experiments. DJ and MM designed and performed the multiplex antibody suspension bead assay preparation and sample analysis with the participation of ALL, CF, and MI performed the IC-MS experiments and corresponding data analysis. DJ, AM, and EB performed statistical analysis and multivariate modeling. ALL, DJ, and AM drafted the manuscript. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
Prof. Dr. Björn Gerdle reports grants from the Swedish Research Council, grants from County Council of Östergötland (Research-ALF), grants from AFA Insurance, during the conduct of the study. Dr. Emmanuel Bäckryd reports grants from ALF Grants, Region Östergötland, grants from NEURO Sweden, during the conduct of the study. The authors declare no other conflicts of interest in this work.
References
1. Meyer-Rosberg K, Burckhardt CS, Huizar K, Kvarnström A, Nordfors LO, Kristofferson A. A comparison of the SF-36 and Nottingham health profile in patients with chronic neuropathic pain. Eur J Pain. 2001;5(4):391–403. doi:10.1053/eujp.2001.0260
2. Hans G, Masquelier E, De Cock P. The diagnosis and management of neuropathic pain in daily practice in Belgium: an observational study. BMC Public Health. 2007;7:170. doi:10.1186/1471-2458-7-170
3. Berger A, Dukes EM, Oster G. Clinical characteristics and economic costs of patients with painful neuropathic disorders. J Pain. 2004;5(3):143–149. doi:10.1016/j.jpain.2003.12.004
4. Truini A, Cruccu G. How diagnostic tests help to disentangle the mechanisms underlying neuropathic pain symptoms in painful neuropathies. Pain. 2016;157(Suppl 1):S53–S59. doi:10.1097/j.pain.0000000000000367
5. Ablin J, Fitzcharles MA, Buskila D, Shir Y, Sommer C, Häuser W. Treatment of fibromyalgia syndrome: recommendations of recent evidence-based interdisciplinary guidelines with special emphasis on complementary and alternative therapies. Evid Based Complement Alternat Med. 2013;2013:485272. doi:10.1155/2013/485272
6. Walitt B, Katz RS, Bergman MJ, Wolfe F. Three-quarters of persons in the us population reporting a clinical diagnosis of fibromyalgia do not satisfy fibromyalgia criteria: the 2012 National Health Interview Survey. PLoS One. 2016;11(6):e0157235. doi:10.1371/journal.pone.0157235
7. Wolfe F, Egloff N, Häuser W. Widespread pain and low widespread pain index scores among fibromyalgia-positive cases assessed with the 2010/2011 fibromyalgia criteria. J Rheumatol. 2016;43(9):1743–1748. doi:10.1007/s00134-008-1077-9
8. Mick G, Baron R, Correa-Illanes G, et al. Is an easy and reliable diagnosis of localized neuropathic pain (LNP) possible in general practice? Development of a screening tool based on IASP criteria. Curr Med Res Opin. 2014;30(7):1357–1366. doi:10.1185/03007995.2014.907562
9. Treede RD, Jensen TS, Campbell JN, et al. Neuropathic pain: redefinition and a grading system for clinical and research purposes. Neurology. 2008;70(18):1630–1635. doi:10.1212/01.wnl.0000282763.29778.59
10. Finnerup NB, Haroutounian S, Kamerman P, et al. Neuropathic pain: an updated grading system for research and clinical practice. Pain. 2016;157(8):1599–1606. doi:10.1097/j.pain.0000000000000492
11. Wolfe F, Clauw DJ, Fitzcharles MA, et al. The American College of Rheumatology preliminary diagnostic criteria for fibromyalgia and measurement of symptom severity. Arthritis Care Res (Hoboken). 2010;62(5):600–610. doi:10.1002/acr.20140
12. Wolfe F, Clauw DJ, Fitzcharles MA, et al. Fibromyalgia criteria and severity scales for clinical and epidemiological studies: a modification of the ACR preliminary diagnostic criteria for fibromyalgia. J Rheumatol. 2011;38(6):1113–1122. doi:10.3899/jrheum.100594
13. Baron R. Mechanisms of disease: neuropathic pain–a clinical perspective. Nat Clin Pract Neurol. 2006;2(2):95–106. doi:10.1038/ncpneuro0113
14. Dworkin RH, O’Connor AB, Backonja M, et al. Pharmacologic management of neuropathic pain: evidence-based recommendations. Pain. 2007;132(3):237–251. doi:10.1016/j.pain.2007.08.033
15. Derry S, Cording M, Wiffen PJ, Law S, Phillips T, Moore RA. Pregabalin for pain in fibromyalgia in adults. Cochrane Database Syst Rev. 2016;9:CD011790.
16. Cording M, Derry S, Phillips T, Moore RA, Wiffen PJ. Milnacipran for pain in fibromyalgia in adults. Cochrane Database Syst Rev. 2015;(10):CD008244.
17. Theoharides TC, Tsilioni I, Arbetman L, et al. Fibromyalgia syndrome in need of effective treatments. J Pharmacol Exp Ther. 2015;355(2):255–263. doi:10.1124/jpet.115.227298
18. Yuan X, Desiderio DM. Proteomics analysis of human cerebrospinal fluid. J Chromatogr B Analyt Technol Biomed Life Sci. 2005;815(1–2):179–189. doi:10.1016/j.jchromb.2004.06.044
19. Smith BH, Lee J, Price C, Baranowski AP. Neuropathic pain: a pathway for care developed by the British Pain Society. Br J Anaesth. 2013;111(1):73–79. doi:10.1093/bja/aet206
20. Jensen TS, Gottrup H, Sindrup SH, Bach FW. The clinical picture of neuropathic pain. Eur J Pharmacol. 2001;429(1–3):1–11. doi:10.1016/s0014-2999(01)01302-4
21. TWG IASP. Classification of Chronic Pain. In: Descriptions of Chronic Pain Syndromes and Definitions of Pain Terms [webpage].
22. Bouhassira D, Lanteri-Minet M, Attal N, Laurent B, Touboul C. Prevalence of chronic pain with neuropathic characteristics in the general population. Pain. 2008;136(3):380–387. doi:10.1016/j.pain.2007.08.013
23. Dworkin RH. An overview of neuropathic pain: syndromes, symptoms, signs, and several mechanisms. Clin J Pain. 2002;18(6):343–349.
24. Adoukonou T, Gnonlonfoun D, Kpozehouen A, et al. [Prevalence and characteristics of chronic pain with neuropathic component at Parakou in northern Benin in 2012]. Rev Neurol (Paris). 2014;170(11):703–711. doi:10.1016/j.neurol.2014.07.013
25. Ferreira K, Schwalbach MT, Schwalbach J, Speciali J. Chronic pain in Maputo, Mozambique: new insights. Pain Med. 2013;14(4):551–553. doi:10.1111/j.1526-4637.2012.01429.x
26. de Moraes Vieira EB, Garcia JB, Da Silva AA, Mualem Araújo RL, Jansen RC. Prevalence, characteristics, and factors associated with chronic pain with and without neuropathic characteristics in São Luís, Brazil. J Pain Symptom Manage. 2012;44(2):239–251. doi:10.1016/j.jpainsymman.2011.08.014
27. Harifi G, Amine M, Ait Ouazar M, et al. Prevalence of chronic pain with neuropathic characteristics in the Moroccan general population: a national survey. Pain Med. 2013;14(2):287–292. doi:10.1111/pme.12009
28. Yawn BP, Wollan PC, Weingarten TN, Watson JC, Hooten WM, Melton LJ. The prevalence of neuropathic pain: clinical evaluation compared with screening tools in a community population. Pain Med. 2009;10(3):586–593. doi:10.1111/j.1526-4637.2009.00588.x
29. Torrance N, Smith BH, Bennett MI, Lee AJ. The epidemiology of chronic pain of predominantly neuropathic origin. Results from a general population survey. J Pain. 2006;7(4):281–289. doi:10.1016/j.jpain.2005.11.008
30. Conti A, Ricchiuto P, Iannaccone S, et al. Pigment epithelium-derived factor is differentially expressed in peripheral neuropathies. Proteomics. 2005;5(17):4558–4567. doi:10.1002/pmic.200402088
31. Bäckryd E, Ghafouri B, Larsson B, Gerdle B. Do low levels of beta-endorphin in the cerebrospinal fluid indicate defective top-down inhibition in patients with chronic neuropathic pain? A cross-sectional, comparative study. Pain Med. 2014;15(1):111–119. doi:10.1111/pme.12248
32. Bäckryd E, Ghafouri B, Carlsson AK, Olausson P, Gerdle B. Multivariate proteomic analysis of the cerebrospinal fluid of patients with peripheral neuropathic pain and healthy controls - a hypothesis-generating pilot study. J Pain Res. 2015;8:321–333. doi:10.2147/JPR.S82970
33. Cannistraci CV, Ravasi T, Montevecchi FM, Ideker T, Alessio M. Nonlinear dimension reduction and clustering by minimum curvilinearity unfold neuropathic pain and tissue embryological classes. Bioinformatics. 2010;26(18):i531–i539. doi:10.1093/bioinformatics/btq376
34. Guieu R, Peragut JC, Roussel P, et al. Adenosine and neuropathic pain. Pain. 1996;68(2–3):271–274. doi:10.1016/s0304-3959(96)03214-9
35. Zhao W, Wang Y, Fang Q, et al. Changes in neurotrophic and inflammatory factors in the cerebrospinal fluid of patients with postherpetic neuralgia. Neurosci Lett. 2017;637:108–113. doi:10.1016/j.neulet.2016.11.041
36. Ludwig J, Binder A, Steinmann J, Wasner G, Baron R. Cytokine expression in serum and cerebrospinal fluid in non-inflammatory polyneuropathies. J Neurol Neurosurg Psychiatry. 2008;79(11):1268–1273. doi:10.1136/jnnp.2007.134528
37. Eisenach JC, Thomas JA, Rauck RL, Curry R, Li X. Cystatin C in cerebrospinal fluid is not a diagnostic test for pain in humans. Pain. 2004;107(3):207–212. doi:10.1016/j.pain.2003.09.007
38. Clauw DJ. Fibromyalgia: a clinical review. JAMA. 2014;311(15):1547–1555. doi:10.1001/jama.2014.3266
39. Olausson P, Gerdle B, Ghafouri N, Sjostrom D, Blixt E, Ghafouri B. Protein alterations in women with chronic widespread pain–an explorative proteomic study of the trapezius muscle. Sci Rep. 2015;5:11894. doi:10.1038/srep11894
40. Ernberg M, Christidis N, Ghafouri B, et al. Plasma cytokine levels in fibromyalgia and their response to 15 weeks of progressive resistance exercise or relaxation therapy. Mediators Inflamm. 2018;2018:14. doi:10.1155/2018/3985154
41. Gerdle B, Ghafouri B, Ghafouri N, Backryd E, Gordh T. Signs of ongoing inflammation in female patients with chronic widespread pain: a multivariate, explorative, cross-sectional study of blood samples. Medicine (Baltimore). 2017;96(9):e6130. doi:10.1097/MD.0000000000006130
42. Olausson P, Gerdle B, Ghafouri N, Larsson B, Ghafouri B. Identification of proteins from interstitium of trapezius muscle in women with chronic myalgia using microdialysis in combination with proteomics. PLoS One. 2012;7(12):e52560. doi:10.1371/journal.pone.0052560
43. Stensson N, Ghafouri B, Gerdle B, Ghafouri N. Alterations of anti-inflammatory lipids in plasma from women with chronic widespread pain - a case control study. Lipids Health Dis. 2017;16(1):112. doi:10.1186/s12944-017-0565-8
44. Rodriguez-Pintó I, Agmon-Levin N, Howard A, Shoenfeld Y. Fibromyalgia and cytokines. Immunol Lett. 2014;161(2):200–203. doi:10.1016/j.imlet.2014.01.009
45. Üçeyler N, Sommer C. Small nerve fiber pathology. In: Häuser W, Perrot S, editors. Fibromylagia Syndrome and Widespread Pain - from Construction to Relevant Recognition. Philadelphia: Wolters Kluwer; 2018:204–214.
46. Giovengo SL, Russell IJ, Larson AA. Increased concentrations of nerve growth factor in cerebrospinal fluid of patients with fibromyalgia. J Rheumatol. 1999;26(7):1564–1569.
47. Sarchielli P, Mancini ML, Floridi A, et al. Increased levels of neurotrophins are not specific for chronic migraine: evidence from primary fibromyalgia syndrome. J Pain. 2007;8(9):737–745. doi:10.1016/j.jpain.2007.05.002
48. Kadetoff D, Lampa J, Westman M, Andersson M, Kosek E. Evidence of central inflammation in fibromyalgia-increased cerebrospinal fluid interleukin-8 levels. J Neuroimmunol. 2012;242(1–2):33–38. doi:10.1016/j.jneuroim.2011.10.013
49. Kosek E, Altawil R, Kadetoff D, et al. Evidence of different mediators of central inflammation in dysfunctional and inflammatory pain–interleukin-8 in fibromyalgia and interleukin-1 β in rheumatoid arthritis. J Neuroimmunol. 2015;280:49–55. doi:10.1016/j.jneuroim.2015.02.002
50. Sarchielli P, Alberti A, Candeliere A, Floridi A, Capocchi G, Calabresi P. Glial cell line-derived neurotrophic factor and somatostatin levels in cerebrospinal fluid of patients affected by chronic migraine and fibromyalgia. Cephalalgia. 2006;26(4):409–415. doi:10.1111/j.1468-2982.2005.01048.x
51. Peres MF, Zukerman E, Senne Soares CA, Alonso EO, Santos BF, Faulhaber MH. Cerebrospinal fluid glutamate levels in chronic migraine. Cephalalgia. 2004;24(9):735–739. doi:10.1111/j.1468-2982.2004.00750.x
52. McLean SA, Williams DA, Stein PK, et al. Cerebrospinal fluid corticotropin-releasing factor concentration is associated with pain but not fatigue symptoms in patients with fibromyalgia. Neuropsychopharmacology. 2006;31(12):2776–2782. doi:10.1038/sj.npp.1301200
53. Bäckryd E, Tanum L, Lind AL, Larsson A, Gordh T. Evidence of both systemic inflammation and neuroinflammation in fibromyalgia patients, as assessed by a multiplex protein panel applied to the cerebrospinal fluid and to plasma. J Pain Res. 2017;10:515–525. doi:10.2147/JPR.S128508
54. Lind AL, Emami Khoonsari P, Sjödin M, et al. Spinal cord stimulation alters protein levels in the cerebrospinal fluid of neuropathic pain patients: a proteomic mass spectrometric analysis. Neuromodulation. 2016;19(6):549–562. doi:10.1111/ner.12473
55. Haggmark A, Bystrom S, Ayoglu B, et al. Antibody-based profiling of cerebrospinal fluid within multiple sclerosis. Proteomics. 2013;13(15):2256–2267. doi:10.1002/pmic.201200580
56. Häggmark A, Mikus M, Mohsenchian A, et al. Plasma profiling reveals three proteins associated to amyotrophic lateral sclerosis. Ann Clin Transl Neurol. 2014;1(8):544–553. doi:10.1002/acn3.83
57. Remnestål J, Just D, Mitsios N, et al. CSF profiling of the human brain enriched proteome reveals associations of neuromodulin and neurogranin to Alzheimer’s disease. Proteomics Clin Appl. 2016;10(12):1242–1253. doi:10.1002/prca.201500150
58. Schwenk JM, Lindberg J, Sundberg M, Uhlen M, Nilsson P. Determination of binding specificities in highly multiplexed bead-based assays for antibody proteomics. Mol Cell Proteomics. 2007;6(1):125–132. doi:10.1074/mcp.T600035-MCP200
59. Cox J, Mann M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol. 2008;26(12):1367–1372. doi:10.1038/nbt.1511
60. Ihaka R, Gentleman RR. A language for data analysis and graphics. J Comput Graph Stat. 1996;5(3):299–314.
61. Eriksson L, Byrne T, Johansson E, TJaV C. Multi- and Megavariate Data Analysis: Basic Principles and Applications. Malmö, Sweden: MKS Umetrics AB.; 2013.
62. Wheelock Å, Wheelock CE. Trials and tribulations of ‘omics data analysis: assessing quality of SIMCA-based multivariate models using examples from pulmonary medicine. Mol Biosyst. 2013;9(11):2589–2596. doi:10.1039/c3mb70194h
63. Lauer SJ, Walker D, Elshourbagy NA, Reardon CA, Levy-Wilson B, Taylor JM. Two copies of the human apolipoprotein C-I gene are linked closely to the apolipoprotein E gene. J Biol Chem. 1988;263(15):7277–7286.
64. Abildayeva K, Berbée JF, Blokland A, et al. Human apolipoprotein C-I expression in mice impairs learning and memory functions. J Lipid Res. 2008;49(4):856–869. doi:10.1194/jlr.M700518-JLR200
65. Elliott DA, Weickert CS, Garner B. Apolipoproteins in the brain: implications for neurological and psychiatric disorders. Clin Lipidol. 2010;51(4):555–573. doi:10.2217/CLP.10.37
66. Gordh T, Sharma HS. Chronic spinal nerve ligation induces microvascular permeability disturbances, astrocytic reaction, and structural changes in the rat spinal cord. Acta Neurochir Suppl. 2006;96:335–340.
67. Gordh T, Chu H, Sharma HS. Spinal nerve lesion alters blood-spinal cord barrier function and activates astrocytes in the rat. Pain. 2006;124(1–2):211–221. doi:10.1016/j.pain.2006.05.020
68. Bellei E, Vilella A, Monari E, et al. Serum protein changes in a rat model of chronic pain show a correlation between animal and humans. Sci Rep. 2017;7:41723. doi:10.1038/srep41723
69. Backryd E, Lind AL, Thulin M, Larsson A, Gerdle B, Gordh T. High levels of cerebrospinal fluid chemokines point to the presence of neuroinflammation in peripheral neuropathic pain: a cross-sectional study of 2 cohorts of patients compared with healthy controls. Pain. 2017;158(12):2487–2495. doi:10.1097/j.pain.0000000000001061
70. Moen A, Lind AL, Thulin M, et al. Inflammatory serum protein profiling of patients with lumbar radicular pain one year after disc herniation. Int J Inflam. 2016;2016:3874964. doi:10.1155/2016/3874964
71. Schippers EF, Berbée JF, van Disseldorp IM, et al. Preoperative apolipoprotein CI levels correlate positively with the proinflammatory response in patients experiencing endotoxemia following elective cardiac surgery. Intensive Care Med. 2008;34(8):1492–1497. doi:10.1007/s00134-008-1077-9
72. Curocichin G, Wu Y, McDade TW, et al. Single-nucleotide polymorphisms at five loci are associated with C-reactive protein levels in a cohort of Filipino young adults. J Hum Genet. 2011;56(12):823–827. doi:10.1038/jhg.2011.106
73. Elliott P, Chambers JC, Zhang W, et al. Genetic Loci associated with C-reactive protein levels and risk of coronary heart disease. JAMA. 2009;302(1):37–48. doi:10.1001/jama.2009.954
74. Westerterp M, Berbée JF, Pires NM, et al. Apolipoprotein C-I is crucially involved in lipopolysaccharide-induced atherosclerosis development in apolipoprotein E-knockout mice. Circulation. 2007;116(19):2173–2181. doi:10.1161/CIRCULATIONAHA.107.693382
75. Berbée JF, Coomans CP, Westerterp M, Romijn JA, Havekes LM, Rensen PC. Apolipoprotein CI enhances the biological response to LPS via the CD14/TLR4 pathway by LPS-binding elements in both its N- and C-terminal helix. J Lipid Res. 2010;51(7):1943–1952. doi:10.1194/jlr.M006809
76. Berbée JF, van der Hoogt CC, Kleemann R, et al. Apolipoprotein CI stimulates the response to lipopolysaccharide and reduces mortality in gram-negative sepsis. Faseb J. 2006;20(12):2162–2164. doi:10.1096/fj.05-5639fje
77. Tanga FY, Nutile-McMenemy N, DeLeo JA. The CNS role of Toll-like receptor 4 in innate neuroimmunity and painful neuropathy. Proc Natl Acad Sci U S A. 2005;102(16):5856–5861. doi:10.1073/pnas.0501634102
78. Hutchinson MR, Bland ST, Johnson KW, Rice KC, Maier SF, Watkins LR. Opioid-induced glial activation: mechanisms of activation and implications for opioid analgesia, dependence, and reward. ScientificWorldJournal. 2007;7:98–111. doi:10.1100/tsw.2007.230
79. Bettoni I, Comelli F, Rossini C, et al. Glial TLR4 receptor as new target to treat neuropathic pain: efficacy of a new receptor antagonist in a model of peripheral nerve injury in mice. Glia. 2008;56(12):1312–1319. doi:10.1002/glia.20699
80. Wang X, Grace PM, Pham MN, et al. Rifampin inhibits Toll-like receptor 4 signaling by targeting myeloid differentiation protein 2 and attenuates neuropathic pain. Faseb J. 2013;27(7):2713–2722. doi:10.1096/fj.12-222992
81. Milligan ED, Watkins LR. Pathological and protective roles of glia in chronic pain. Nat Rev Neurosci. 2009;10(1):23–36. doi:10.1038/nrn2533
82. Buchanan MM, Hutchinson M, Watkins LR, Yin H. Toll-like receptor 4 in CNS pathologies. J Neurochem. 2010;114(1):13–27. doi:10.1111/j.1471-4159.2010.06736.x
83. Hutchinson MR, Zhang Y, Brown K, et al. Non-stereoselective reversal of neuropathic pain by naloxone and naltrexone: involvement of toll-like receptor 4 (TLR4). Eur J Neurosci. 2008;28(1):20–29. doi:10.1111/j.1460-9568.2008.06321.x
84. Osterloh A, Breloer M. Heat shock proteins: linking danger and pathogen recognition. Med Microbiol Immunol. 2008;197(1):1–8. doi:10.1007/s00430-007-0055-0
85. Smiley ST, King JA, Hancock WW. Fibrinogen stimulates macrophage chemokine secretion through toll-like receptor 4. J Immunol. 2001;167(5):2887–2894. doi:10.4049/jimmunol.167.5.2887
86. Hutchinson MR, Ramos KM, Loram LC, et al. Evidence for a role of heat shock protein-90 in toll like receptor 4 mediated pain enhancement in rats. Neuroscience. 2009;164(4):1821–1832. doi:10.1016/j.neuroscience.2009.09.046
87. Li J, Csakai A, Jin J, Zhang F, Yin H. Therapeutic developments targeting toll-like receptor-4-mediated neuroinflammation. ChemMedChem. 2016;11(2):154–165. doi:10.1002/cmdc.201500188
88. Cudaback E, Li X, Yang Y, et al. Apolipoprotein C-I is an APOE genotype-dependent suppressor of glial activation. J Neuroinflammation. 2012;9:192. doi:10.1186/1742-2094-9-192
89. Huang R, Hughes M, Mobley S, Lanham I, Poduslo SE. APOE genotypes in African American female multiple sclerosis patients. Neurosci Lett. 2007;414(1):51–56. doi:10.1016/j.neulet.2006.12.049
90. Xu Y, Berglund L, Ramakrishnan R, et al. A common Hpa I RFLP of apolipoprotein C-I increases gene transcription and exhibits an ethnically distinct pattern of linkage disequilibrium with the alleles of apolipoprotein E. J Lipid Res. 1999;40(1):50–58.
91. Jong MC, van Dijk KW, Dahlmans VE, et al. Reversal of hyperlipidaemia in apolipoprotein C1 transgenic mice by adenovirus-mediated gene delivery of the low-density-lipoprotein receptor, but not by the very-low-density-lipoprotein receptor. Biochem J. 1999;338(Pt 2):281–287.
92. Lim TKY, Anderson KM, Hari P, et al. Evidence for a role of nerve injury in painful intervertebral disc degeneration: a cross-sectional proteomic analysis of human cerebrospinal fluid. J Pain. 2017. doi:10.1016/j.jpain.2017.06.002
93. Jiménez CR, Stam FJ, Li KW, et al. Proteomics of the injured rat sciatic nerve reveals protein expression dynamics during regeneration. Mol Cell Proteomics. 2005;4(2):120–132. doi:10.1074/mcp.M400076-MCP200
94. Miao J, Wang F, Zheng W, Zhuang X. Association of the apolipoprotein E polymorphism with migraine: a meta-analysis. BMC Neurol. 2015;15:138. doi:10.1186/s12883-015-0385-2
95. Reeser JC, Payne E, Kitchner T, McCarty CA. Apolipoprotein e4 genotype increases the risk of being diagnosed with posttraumatic fibromyalgia. Pm R. 2011;3(3):193–197. doi:10.1016/j.pmrj.2010.12.009
96. de Knegt NC, Schuengel C, Evenhuis HM, Lobbezoo F, Scherder EJ. Apolipoprotein E ɛ4, cognitive function, and pain experience in down syndrome: a pilot study. Arch Clin Neuropsychol. 2016;31(5):389–400. doi:10.1093/arclin/acw022
97. Cameron BM, VanderPutten DM, Merril CR. Preliminary study of an increase of a plasma apolipoprotein E variant associated with peripheral nerve damage. A finding in patients with chronic spinal pain. Spine (Phila Pa 1976). 1995;20(5):
98. Ueda H
99. Baraniuk JN, Casado B, Maibach H, Clauw DJ, Pannell LK, Hess SS. A chronic fatigue syndrome - related proteome in human cerebrospinal fluid. BMC Neurol. 2005;5:22. doi:10.1186/1471-2377-5-22
100. Juárez-Contreras R, Rosenbaum T, Morales-Lázaro SL, Acid L. Ion channels as molecular mediators of pain. Front Mol Neurosci. 2018;11:462. doi:10.3389/fnmol.2018.00462
101. van Meeteren LA, Ruurs P, Christodoulou E, et al. Inhibition of autotaxin by lysophosphatidic acid and sphingosine 1-phosphate. J Biol Chem. 2005;280(22):21155–21161. doi:10.1074/jbc.M413183200
102. Ueda H. Lysophosphatidic acid signaling is the definitive mechanism underlying neuropathic pain. Pain. 2017. doi:10.1097/j.pain.0000000000000813
103. Benesch MG, Zhao YY, Curtis JM, McMullen TP, Brindley DN. Regulation of autotaxin expression and secretion by lysophosphatidate and sphingosine 1-phosphate. J Lipid Res. 2015;56(6):1134–1144. doi:10.1194/jlr.M057661
104. Kempuraj D, Thangavel R, Natteru PA, et al. Neuroinflammation induces neurodegeneration. J Neurol Neurosurg Spine. 2016;1(1):1003.
105. Goldshmit Y, Matteo R, Sztal T, et al. Blockage of lysophosphatidic acid signaling improves spinal cord injury outcomes. Am J Pathol. 2012;181(3):978–992. doi:10.1016/j.ajpath.2012.06.007
106. McDougall JJ, Albacete S, Schuelert N, et al. Lysophosphatidic acid provides a missing link between osteoarthritis and joint neuropathic pain. Osteoarthritis Cartilage. 2017;25(6):926–934
107. Caro XJ, Winter EF, Dumas AJ. A subset of fibromyalgia patients have findings suggestive of chronic inflammatory demyelinating polyneuropathy and appear to respond to IVIg. Rheumatology (Oxford). 2008;47(2):208–211. doi:10.1093/rheumatology/kem345
108. Kim SH, Kim DH, Oh DH, Clauw DJ. Characteristic electron microscopic findings in the skin of patients with fibromyalgia–preliminary study. Clin Rheumatol. 2008;27(3):407–411.
109. Giannoccaro MP, Donadio V, Incensi A, Avoni P, Liguori R. Small nerve fiber involvement in patients referred for fibromyalgia. Muscle Nerve. 2014;49(5):757–759. doi:10.1002/mus.24156
110. Oaklander AL, Herzog ZD, Downs HM, Klein MM. Objective evidence that small-fiber polyneuropathy underlies some illnesses currently labeled as fibromyalgia. Pain. 2013;154(11):2310–2316. doi:10.1016/j.pain.2013.06.001
111. Ramírez M, Martínez-Martínez LA, Hernández-Quintela E, Velazco-Casapía J, Vargas A, Martínez-Lavín M. Small fiber neuropathy in women with fibromyalgia. An in vivo assessment using corneal confocal bio-microscopy. Semin Arthritis Rheum. 2015;45(2):214–219. doi:10.1016/j.semarthrit.2015.03.003
112. Serra J, Collado A, Solà R, et al. Hyperexcitable C nociceptors in fibromyalgia. Ann Neurol. 2014;75(2):196–208. doi:10.1002/ana.24065
113. Üçeyler N, Zeller D, Kahn AK, et al. Small fibre pathology in patients with fibromyalgia syndrome. Brain. 2013;136(Pt 6):1857–1867. doi:10.1093/brain/awt053
114. Inoue M, Xie W, Matsushita Y, Chun J, Aoki J, Ueda H. Lysophosphatidylcholine induces neuropathic pain through an action of autotaxin to generate lysophosphatidic acid. Neuroscience. 2008;152(2):296–298. doi:10.1016/j.neuroscience.2007.12.041
115. Inoue M, Ma L, Aoki J, Chun J, Ueda H. Autotaxin, a synthetic enzyme of lysophosphatidic acid (LPA), mediates the induction of nerve-injured neuropathic pain. Mol Pain. 2008;4:6. doi:10.1186/1744-8069-4-6
116. Inoue M, Ma L, Aoki J, Ueda H. Simultaneous stimulation of spinal NK1 and NMDA receptors produces LPC which undergoes ATX-mediated conversion to LPA, an initiator of neuropathic pain. J Neurochem. 2008;107(6):1556–1565. doi:10.1111/j.1471-4159.2008.05725.x
117. Velasco M, O’Sullivan C, Sheridan GK. Lysophosphatidic acid receptors (LPARs): potential targets for the treatment of neuropathic pain. Neuropharmacology. 2017;113(Pt B):608–617.
118. Wheeler NA, Fuss B. Extracellular cues influencing oligodendrocyte differentiation and (re)myelination. Exp Neurol. 2016;283(Pt B):512–530. doi:10.1016/j.expneurol.2016.03.019
119. Dennis J, Morgan MK, Graf MR, Fuss B. P2Y12 receptor expression is a critical determinant of functional responsiveness to ATX’s MORFO domain. Purinergic Signal. 2012;8(2):181–190. doi:10.1007/s11302-011-9283-2
120. Dennis J, White MA, Forrest AD, et al. Phosphodiesterase-Ialpha/autotaxin’s MORFO domain regulates oligodendroglial process network formation and focal adhesion organization. Mol Cell Neurosci. 2008;37(2):412–424. doi:10.1016/j.mcn.2007.10.018
121. Fox MA, Colello RJ, Macklin WB, Fuss B. Phosphodiesterase-Ialpha/autotaxin: a counteradhesive protein expressed by oligodendrocytes during onset of myelination. Mol Cell Neurosci. 2003;23(3):507–519.
122. The UniProt Consortium. UniProt: the universal protein knowledgebase. Nucleic Acids Res. 2017;45(D1):D158–D169. doi:10.1093/nar/gkw1099.
123. Coste O, Brenneis C, Linke B, et al. Sphingosine 1-phosphate modulates spinal nociceptive processing. J Biol Chem. 2008;283(47):32442–32451. doi:10.1074/jbc.M806410200
124. Zhang YH, Fehrenbacher JC, Vasko MR, Nicol GD. Sphingosine-1-phosphate via activation of a G-protein-coupled receptor(s) enhances the excitability of rat sensory neurons. J Neurophysiol. 2006;96(3):1042–1052. doi:10.1152/jn.00120.2006
125. Zhang YH, Vasko MR, Nicol GD. Intracellular sphingosine 1-phosphate mediates the increased excitability produced by nerve growth factor in rat sensory neurons. J Physiol. 2006;575(Pt 1):101–113. doi:10.1113/jphysiol.2006.111575
126. Coste O, Pierre S, Marian C, et al. Antinociceptive activity of the S1P-receptor agonist FTY720. J Cell Mol Med. 2008;12(3):995–1004. doi:10.1111/j.1582-4934.2008.00160.x
127. Mair N, Benetti C, Andratsch M, et al. Genetic evidence for involvement of neuronally expressed S1P1 receptor in nociceptor sensitization and inflammatory pain. PLoS One. 2011;6(2):e17268. doi:10.1371/journal.pone.0017268
128. Selley DE, Welch SP, Sim-Selley LJ. Sphingosine lysolipids in the CNS: endogenous cannabinoid antagonists or a parallel pain modulatory system? Life Sci. 2013;93(5–6):187–193. doi:10.1016/j.lfs.2013.06.004
129. Patti GJ, Yanes O, Shriver LP, et al. Metabolomics implicates altered sphingolipids in chronic pain of neuropathic origin. Nat Chem Biol. 2012;8(3):232–234. doi:10.1038/nchembio.767
130. Hulens M, Rasschaert R, Dankaerts W, Stalmans I, Vansant G, Bruyninckx F. Spinal fluid evacuation may provide temporary relief for patients with unexplained widespread pain and fibromyalgia. Med Hypotheses. 2018;118:55–58. doi:10.1016/j.mehy.2018.06.017
131. Fukuhara T, Ono C, Puig-Basagoiti F, Matsuura Y. Roles of lipoproteins and apolipoproteins in particle formation of hepatitis C virus. Trends Microbiol. 2015;23(10):618–629. doi:10.1016/j.tim.2015.07.007
132. Zhou M, Lucas DA, Chan KC, et al. An investigation into the human serum “interactome”. Electrophoresis. 2004;25(9):1289–1298. doi:10.1002/elps.200405866
133. Miao Z, Xie Z, Miao J, Ran J, Feng Y, Xia X. Regulated entry of hepatitis C virus into hepatocytes. Viruses. 2017;9:5. doi:10.3390/v9050100
134. Mellacheruvu D, Wright Z, Couzens AL, et al. The CRAPome: a contaminant repository for affinity purification-mass spectrometry data. Nat Methods. 2013;10(8):730–736. doi:10.1038/nmeth.2557
135. Berven FS, Kroksveen AC, Berle M, et al. Pre-analytical influence on the low molecular weight cerebrospinal fluid proteome. Proteomics Clin Appl. 2007;1(7):699–711. doi:10.1002/prca.200700126
136. Vanderstichele H, Demeyer L, Janelidze S, et al. Recommendations for cerebrospinal fluid collection for the analysis by ELISA of neurogranin trunc P75, α-synuclein, and total tau in combination with Aβ(1-42)/Aβ(1-40). Alzheimers Res Ther. 2017;9(1):40. doi:10.1186/s13195-017-0265-7
137. Enroth S, Hallmans G, Grankvist K, Gyllensten U. Effects of long-term storage time and original sampling month on biobank plasma protein concentrations. EBioMedicine. 2016;12:309–314. doi:10.1016/j.ebiom.2016.08.038
138. Lind AL, Wu D, Freyhult E, et al. A multiplex protein panel applied to cerebrospinal fluid reveals three new biomarker candidates in ALS but none in neuropathic pain patients. PLoS One. 2016;11(2):e0149821. doi:10.1371/journal.pone.0149821
139. Loggia ML, Chonde DB, Akeju O, et al. Evidence for brain glial activation in chronic pain patients. Brain. 2015;138(Pt 3):604–615. doi:10.1093/brain/awu377
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.