")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 15
Comparative Proteomic Analysis of Membrane Vesicles from Clinical C. acnes Isolates with Differential Antibiotic Resistance
Authors Jiang M, Fan X , Jiang Z , Chen H , Liu Y, Yu T, Huang Q, Ma Y
Received 22 February 2022
Accepted for publication 12 April 2022
Published 17 April 2022 Volume 2022:15 Pages 703—712
DOI https://doi.org/10.2147/CCID.S363537
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Jeffrey Weinberg
Min Jiang,* Xiaoyao Fan,* Ziqi Jiang, Huyan Chen, Ye Liu, Tianze Yu, Qiong Huang, Ying Ma
Department of Dermatology, Huashan Hospital, Fudan University, Shanghai, 200040, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qiong Huang; Ying Ma, Department of Dermatology, Huashan Hospital, Fudan University, No. 12 Wulumuqizhong Road, Shanghai, 200040, People’s Republic of China, Email [email protected]; [email protected]
Purpose: Cutibacterium acnes (C. acnes) is closely associated with the pathogenesis of acne, and antibiotics targeting C. acnes have been widely used for decades. However, antibiotic resistance has been increasing rapidly. Membrane vesicles (MVs) have been found to play important roles in antibiotic resistance in some bacteria. We aimed to explore the mechanism of antibiotic resistance and the virulence components within C. acnes-derived MVs.
Materials and Methods: We isolated clinical C. acnes strains from the lesions of acne patients who were sensitive or resistant to the antibiotics erythromycin and clindamycin. We analyzed the proteome of MVs from four sensitive C. acnes isolates and three resistant isolates by LC-MS/MS.
Results: We identified 543 proteins within the MVs of clinical C. acnes strains. Several lipases, NlpC/P60, CAMP factor, and Hta domain protein were detected as virulence factors in the C. acnes-derived MVs. The levels of two lipases and FtsZ were significantly higher in resistant C. acnes-derived MVs compared with sensitive strains (p < 0.05).
Conclusion: According to the implications of this study, improper antibiotic use might not only increase antibiotic resistance in C. acnes but could also further alter the cutaneous lipid composition and aggravate host inflammation, thus resulting in worse clinical manifestations in acne patients. This study re-emphasizes that the improper use of antibiotics should be treated more seriously in clinical practice. Furthermore, to combat multidrug resistance in C. acnes, this study suggests that FtsZ inhibitors could be useful.
Keywords: Cutibacterium acnes, antibiotics, FtsZ inhibitor, membrane vesicles
Introduction
Membrane vesicles (MVs) serve important functions in the bacterial life cycle, participating in virulence factor secretion, biofilm formation, antibiotic stress reactions, microbiome homeostasis, etc.1,2 MVs were initially identified as products of gram-negative bacteria, composed of constituents of the outer membrane (lipopolysaccharide, phospholipids, and proteins) and the periplasm (cytosolic or inner membrane proteins and nucleic acids).3 Because gram-positive bacteria have a thick wall structure and no outer membrane, it was thought for decades that gram-positive bacteria do not secrete MVs.4,5 However, in 2009, Lee et al first found MV secretion in Staphylococcus aureus.6 Since then, an increasing number of gram-positive bacteria (Bacillus spp., Clostridium perfringens, Streptomyces coelicolor, Listeria monocytogenes, etc.) have been shown to produce MVs.7–10 These gram-positive MVs are 20–250 nm in diameter and play roles in biological processes similar to those of gram-negative counterparts.
Cutibacterium acnes (C. acnes, formerly Propionibacterium acnes), an anaerobic gram-positive bacterium, is considered to be closely associated with the pathogenesis of acne.11,12 As one of the main treatments of acne, antibiotics targeting C. acnes have been used widely for decades. However, antibiotic resistance, especially multidrug resistance (MDR), has been increasing rapidly.13 MV production has recently been observed in C. acnes.14 MVs can serve as a secretory system and protect bacteria from antibiotic stress.3,15 Whether C. acnes-derived MVs play roles in antibiotic resistance is unknown. Furthermore, C. acnes-derived MVs have shown strong pro-inflammatory effects on human keratinocytes and peripheral blood monocytes.14 However, the virulence components within the MVs of clinical C. acnes isolates have not been identified. Here, we explored the proteome of MVs from clinically sensitive and resistant isolates to identify virulence components and the possible mechanism of antibiotic resistance in C. acnes. To the best of our knowledge, we are the first to compare the proteome of MVs from different clinical C. acnes isolates.
Materials and Methods
Isolation of C. acnes
We isolated C. acnes clinical strains from the lesions of acne patients in the dermatology department of Huashan Hospital Fudan University (Shanghai, China) according to the methods of our previous study.16 The Ethics Committee of Huashan Hospital Fudan University approved the research, and we conducted the research according to the principles of the Declaration of Helsinki. Informed consent was signed by all the participants. After being sterilized with iodine tincture, the acne lesions were compressed with a comedone extractor. Then the contents were transported immediately with a sample smear and inoculated on Brucella agar (Becton-Dickinson, USA) containing 5% (v/v) lysed defibrinated sheep’s blood (Zhuzhai Blood Reagent Supply Station, Shanghai, China), supplemented with vitamin K1 (Sigma-Aldrich, USA) and incubated in an atmosphere of 80% (v/v) N2, 10% (v/v) CO2, and 10% (v/v) H2 at 37°C for 48–72 h. After two purification cycles were completed, the cultured microorganisms were identified as C. acnes with MALDI-TOF MS (VITEK2, bioMérieux, Marcy I’Etoile, France). The isolated C. acnes strains were then placed in 40% glycerin broth and stored at −70°C until use.
Culture of C. acnes
We cultivated the isolated strains anaerobically. Minimum inhibitory concentrations (MICs) of erythromycin and clindamycin were measured by agar dilution as recommended by the Clinical and Laboratory Standards Institute (CLSI). We collected four sensitive strains that were sensitive to both erythromycin and clindamycin, with MICs ≤0.5 mg/L for erythromycin and ≤2 mg/L for clindamycin. We also collected three strains that were resistant to both erythromycin and clindamycin, with MICs >128 mg/L for erythromycin and clindamycin.
All C. acnes strains were grown in brain-heart infusion (BHI) broth (Difco Laboratories, Detroit, MI) at 37 °C in an atmosphere of 80% (v/v) N2, 10% (v/v) CO2, and 10% (v/v) H2. Cells were cultured for 96 h to reach the mid-exponential phase. We diluted the cell suspensions to adjust the optical density at 600 nm (OD600) to 1.0. Cells were then inoculated into another 100 mL of BHI broth at a 1/100 dilution and cultured for another 96 h. The cell-free culture supernatant was collected for further isolation of MVs.
Isolation of MVs
C. acnes-derived MVs were isolated according to literature methods with some modifications.14 The cell-free culture supernatant was sequentially centrifuged at 2000×g for 10 min and 10,000×g for 30 min at 4 °C. The supernatant was collected and filtered with a 0.22-μm membrane filter (Merck Millipore, Darmstadt, Germany). The supernatant was further ultracentrifuged at 100,000×g for 70 min at 4 °C (Beckman Coulter, Fullerton, CA). The pellet was washed once with PBS and ultracentrifuged again at 100,000×g for 70 min at 4 °C. The MV pellet was finally resuspended in PBS and stored at −80 °C until use.
Identification of MVs
Transmission electron microscopy (TEM) was used to identify the structure of MVs. MVs suspended in 2% paraformaldehyde were loaded on a copper grid and negatively stained with uranyl acetate solution for 5 min. The grid was then examined using a JEOL transmission electron microscope (JEM 2100, Tokyo, Japan) at 100 kV. The size distribution and particle concentration of MVs were measured by high-sensitivity flow cytometry (HSFCM) instrumentation (Flow NanoAnalyzer, Xiamen, China). As previously described, the HSFCM was equipped with a 200 mW 532 nm continuous-wave solid-state.17–19 The Nd:YAG laser was attenuated to 16 mW and used as the excitation source. The light emitted by individual MVs was collected.
Protein Digestion
Proteins were redissolved in 500 mM triethylammonium bicarbonate (TEAB). The protein concentration of the supernatant was measured using the BCA protein assay. One hundred micrograms of protein per condition was transferred into a new tube and adjusted to a final volume of 100 μL with 8 M urea. We added 11 μL of 1 M DTT, incubated the sample at 37 °C for 1 hour, and then transferred the samples into a 10K ultrafiltration tube (Millipore, Bedford, MA). To remove urea, samples were centrifuged by adding 100 mM TEAB three times. Then, 120 μL of 55 mM iodoacetamide was added to the sample and incubated for 20 minutes protected from light at room temperature. Then, the proteins were tryptically digested with sequencing-grade modified trypsin (Promega, Madison, WI). The tryptic peptides were further cleaned by removing sodium deoxycholate and desalted for basic reversed-phase fractionation.
Label-Free Quantitative LC-MS/MS Analysis
Two micrograms of peptide was separated and analyzed with nano-UPLC (EASY-nLC1200, Thermo Scientific, MA, USA) coupled to Q-Exactive mass spectrometry (Thermo Scientific, MA, USA). Separation was performed using a reversed-phase column (100 µm, ID×15 cm, Reprosil-Pur 120 C18-AQ, 1.9 µm, Dr. Math). The mobile phases were H2O with 0.1% FA and 2% ACN (phase A) and 80% ACN and 0.1% FA (phase B). Separation of the sample was executed with a 120-min gradient at 300 nL/min flow. Phase B was ramped from 8 to 30% for 92 min, 30 to 40% for 20 min, 40 to 100% for 2 min, 100% for 2 min, 100 to 2% for 2 min and 2% for 2 min. Data-dependent acquisition was performed in profile and positive mode with an Orbitrap analyzer at a resolution of 70,000 (@200 m/z) and a m/z range of 350–1600 for MS1. For MS2, the resolution was set to 17,500 with a dynamic first mass. The automatic gain control (AGC) target for MS1 was set to 3.0 E+6 with max IT 50 ms, and 5.0 E+4 for MS2 with max IT 100 ms. The top 20 most intense ions were fragmented by HCD with a normalized collision energy of 27% and an isolation window of 2 m/z. The dynamic exclusion time window was 30s.
Data Analysis
Raw MS files were processed with MaxQuant (Version 1.6.1.0). The protein sequence database (UniProt-proteome-Propionibacterium acnes. Fasta 2020_06) was downloaded from UNIPROT. This database and its reverse decoy were then searched against by MaxQuant software. Carbamidomethyl (C) was set as fixed modification. Both peptide and protein FDR should be less than 0.01. The significance analysis of microarrays (SAM) statistical approach was used to identify differentially expressed proteins. Differentially expressed proteins were filtered if their fold change (FC) was greater than 2 and contained at least 2 unique peptides with a statistical p value (SAM) below 0.05. Blast2GO version 4 was used for functional annotation. Statistically altered functions of differentially expressed proteins were calculated by Fisher’s exact test in Blast2GO.
Results
Physical Characterization of MVs from C. acnes
To investigate MVs secreted from C. acnes, MVs were isolated and examined using TEM and HSFCM. The TEM images of the resistant MVs and the sensitive MVs revealed bilayered spherical morphology with an approximate size range of 40 nm to 90 nm (Figure 1). HSFCM indicated that the size distribution of the sensitive MVs was 77.07 nm ± 29.24 nm on average, whereas that of the resistant MVs was 71.70 nm ± 30.61 nm (Figure 2). There were no significant differences between the two groups in morphology or size.
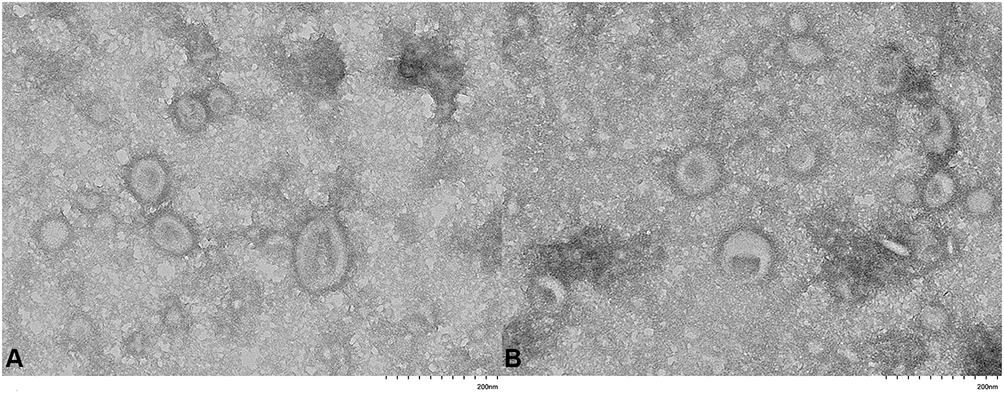 |
Figure 1 TEM image of MVs prepared from C. acnes. (A) Sensitive C. acnes-derived MVs. (B) Resistant C. acnes-derived MVs. |
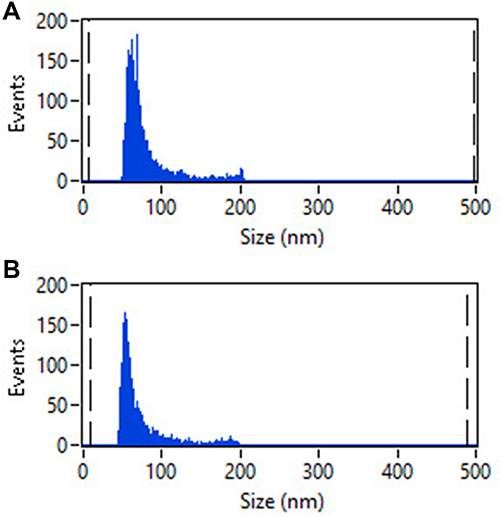 |
Figure 2 The size distribution of C. acnes-derived MVs as measured by HSFCM. (A) MVs from the sensitive C. acnes isolate. (B) MVs from the resistant C. acnes isolate. |
Proteomic Profile of MVs from Clinical C. acnes Strains
A total of 4081 peptides were identified from the MVs of clinical C. acnes strains. Overall, 543 individual proteins with ≥2 unique peptides were identified by a combined dataset. Information on protein properties and detailed peptide data for identified proteins is presented in S1 of the Supporting information. Proteins were classified according to their gene ontology (GO) terms, including biological process (BP), cellular component (CC), and molecular function (MF) (Figure 3). The detailed annotations of BP, CC, and MF are presented in S2 of the Supporting information. Of the total MV proteins from the clinical isolates related to BP, metabolic processes were the most abundant with 249 proteins (82 upregulated and 167 downregulated in the resistant MVs vs sensitive MVs), followed by cellular process with 210 proteins (71 upregulated and 139 downregulated in the resistant MVs vs sensitive MVs). Regarding CC, 152 of the total MV proteins belonged to the cell part (37 upregulated and 115 downregulated in the resistant MVs vs sensitive MVs), 75 proteins belonged to the membrane part (11 upregulated and 64 downregulated in the resistant MVs vs sensitive MVs), and 40 proteins belonged to the protein-containing complex (5 upregulated and 35 downregulated in the resistant MVs vs sensitive MVs). With respect to MF, the MV proteins from the clinical isolates included 277 proteins associated with catalytic activity (108 upregulated and 169 downregulated in the resistant MVs vs sensitive MVs) and 191 proteins associated with binding (60 upregulated and 131 downregulated in the resistant MVs vs sensitive MVs). Other molecular functions included transcription regulator activity, antioxidant activity, transporter activity, molecular carrier activity, and structural molecule activity.
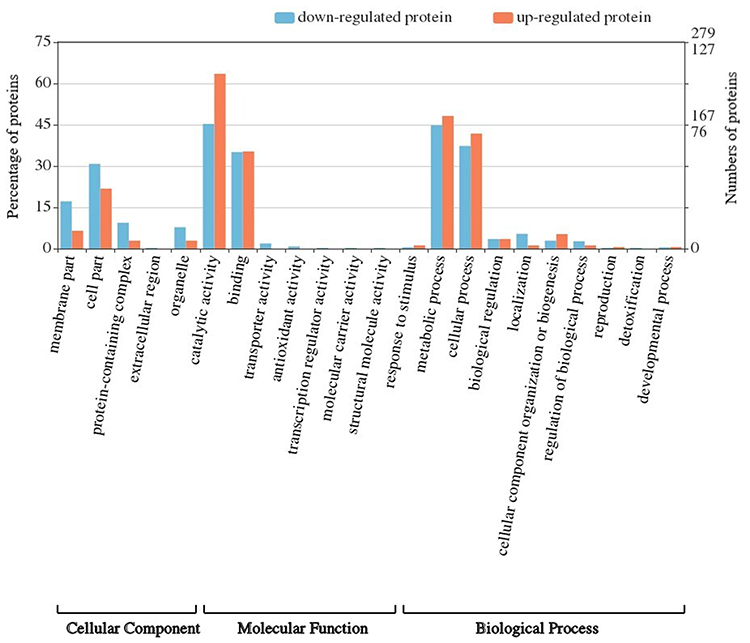 |
Figure 3 GO annotation of all proteins from the C. acnes-derived MVs. Proteins were classified according to BP, CC, and MF. Blue columns indicate proteins downregulated in resistant MVs compared to sensitive MVs. Red columns indicate proteins upregulated in resistant MVs compared to sensitive MVs. |
Differentially Expressed Proteins Between the Resistant MVs and the Sensitive MVs
We further compared the protein components of MVs from the resistant isolates and the sensitive isolates. Twenty-two proteins (unique peptides ≥ 2, FC > 2 or < 0.5, p<0.05) were significantly differentially expressed between the sensitive and resistant isolates (Table 1). Three proteins, D4HCT5 (FtsZ), D4HEV8, and D4H9R7, were upregulated, and another 19 proteins were downregulated. All the significantly differential proteins were enriched with respect to their BP, CC, and MF (Figure 4). From the BP perspective, DNA-templated transcription/elongation, alpha-glucan biosynthetic process, and protein polymerization were the top 3 significantly different processes between the sensitive and resistant strains (p<0.05). In the CC group, the cell division site was the most significantly different component (p<0.01), implicating altered cell division capacity between the sensitive and resistant strains. In the MF group, IMP dehydrogenase activity, serine O-acetyltransferase activity, serine O-acyltransferase activity, transferase activity transferring hexosyl groups, and 1-deoxy-D-xylulose-5-phosphate reductoisomerase activity were the top 5 significantly different functions between the sensitive and resistant strains. The detailed annotation of each differential protein was also obtained from the KEGG (www.kegg.jp) and UNIPROT (www.uniprot.org) websites and is presented in Table 1.
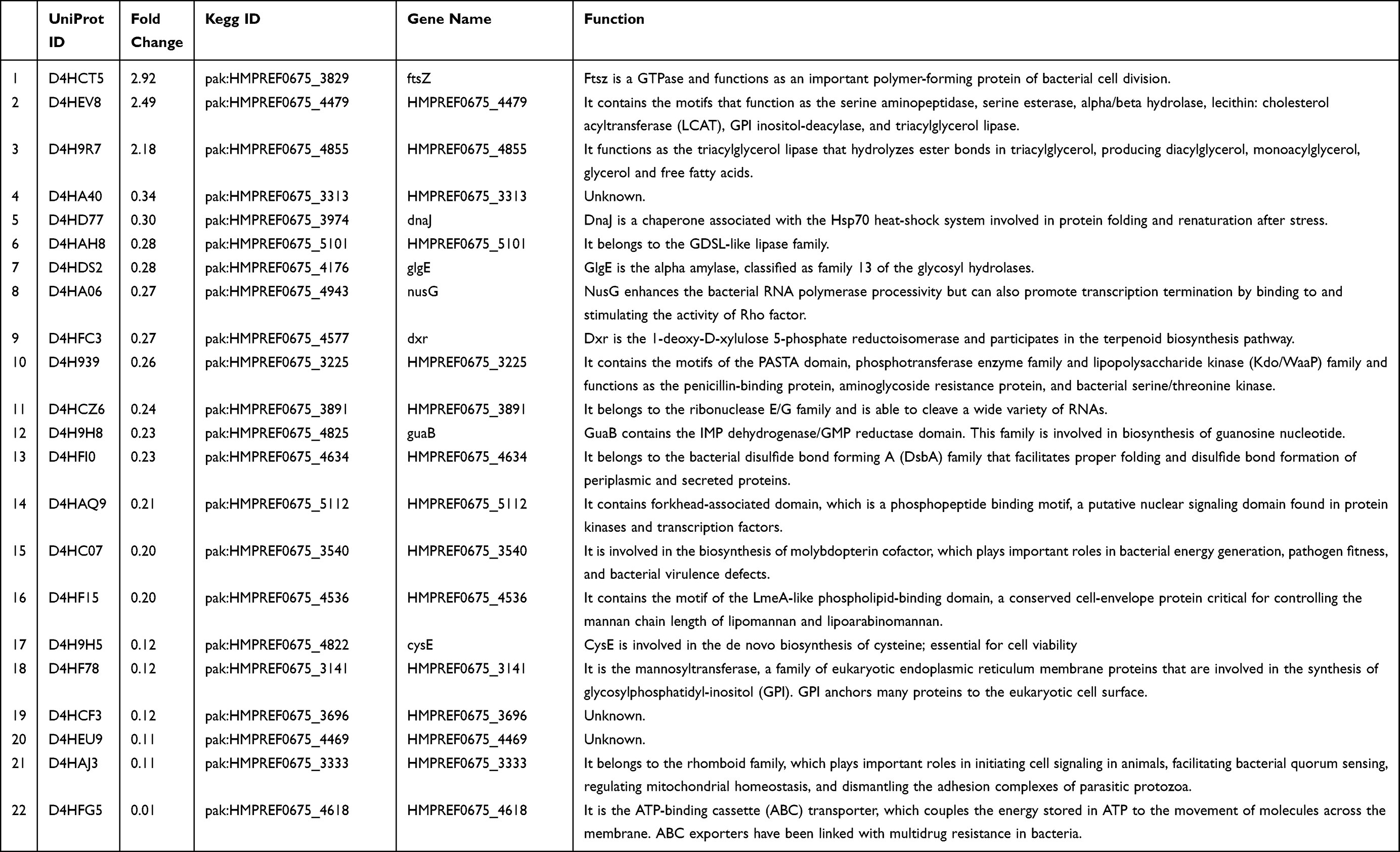 |
Table 1 Detailed Annotation of Each Differentially Expressed Protein |
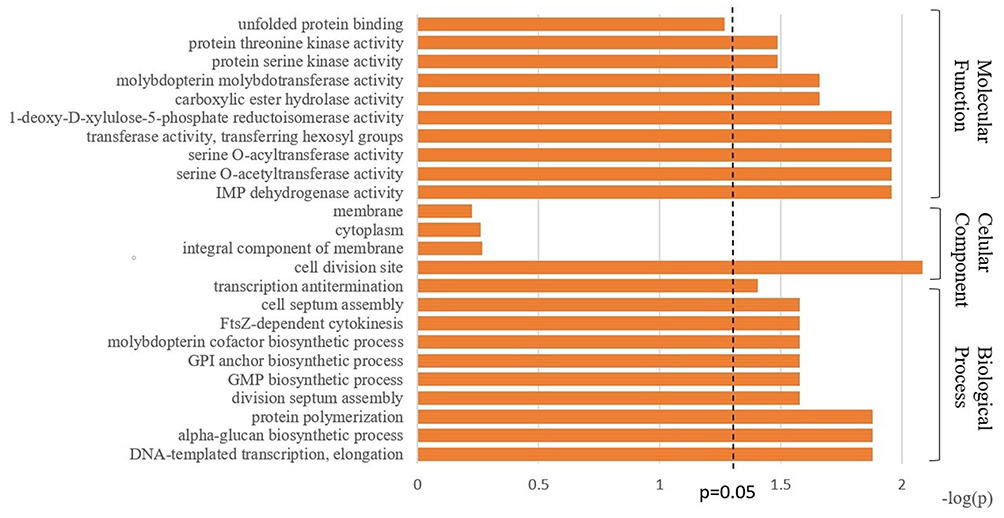 |
Figure 4 GO enrichment based on BP, CC, and MF terms for differentially expressed proteins between the sensitive and resistant MVs. |
Discussion
C. acnes-derived MVs contain many membrane and cytoplasmic proteins, including different kinds of transferases, hydrolases, peptidases, ion binding proteins, DNA polymerases, ATP binding proteins, ribosomal proteins, and other metabolic enzymes. Taken together, the results of this study suggest that C. acnes-derived MVs might facilitate the transfer of proteins involved in bacterial physiology, bacterial virulence, and host inflammation.
We detected several lipases within the C. acnes-derived MVs. According to the GO annotation, three detected proteins (D4HDE2, D4HDE3, D4HFE5) participate in free fatty acid (FFA) metabolism. These three proteins were expressed in both sensitive MVs and resistant MVs, with no significant difference between the two groups. Among the 22 significantly differentially expressed proteins, two (D4HEV8 and D4H9R7) were associated with lipid metabolism based on the KEGG and UNIPROT annotations of the protein motifs. D4H9R7 is a triglyceride lipase, and D4HEV8 is thought to be an ester hydrolase. These two proteins were expressed at significantly higher levels in resistant MVs compared with sensitive MVs (p<0.05). Bacterial metabolism is believed to influence the cutaneous lipid composition. The lipase activity of C. acnes can liberate FFAs from triglycerides (TGs), and the released FFAs are much more viscous than TGs. FFAs lead to ductal hypercornification and increase adhesion between C. acnes and keratinocytes, promoting colonization of C. acnes and the formation of biofilms.20,21 Furthermore, FFAs are thought to be inflammatory.22 Mono-unsaturated acids among the released FFAs are known as the triggering signal of the inflammasome response, which plays important roles in the pathogenesis of acne.23,24 In a previous study, it was reported that a type IA clinical strain shows a higher level of secreted triglyceride lipase protein than a clinical IB strain.25,26 In our study, triglyceride lipase (D4H9R7) and ester hydrolase (D4HEV8) levels were both significantly higher in resistant C. acnes-derived MVs than in sensitive MVs, which implies that resistant strains have different lipase activities from sensitive strains, thus eliciting different inflammatory responses and cutaneous lipid compositions among patients refractory to antibiotic treatment.
Previous studies by Choi have shown that C. acnes-derived MVs induce epidermal deformation and increase the expression of proinflammatory cytokines in keratinocytes, indicating the important role of C. acnes-derived MVs in the pathogenesis of acne.14 Here, our proteome analysis confirmed the pathogenic potential of MVs by detecting NlpC/P60, CAMP factors, and Hta domain proteins in C. acnes-derived MVs. D4HAA4 is a peptidoglycan DL-endopeptidase that belongs to the NlpC/P60 family protein. NlpC/P60 family proteins can mediate the release of peptidoglycan (PGN) fragments that have been shown to modulate host inflammatory responses.27,28 PGN fragments can activate intracellular immune signaling by binding nucleotide-binding oligomerization domain (Nod) 1 and Nod2 receptors.29 Nod1 is activated by meso-diaminopimelic acid (mDAP)-containing muropeptides, which are found in the cell walls of most gram-negative bacteria and some gram-positive bacteria. Nod2 usually recognizes muramyl dipeptide (MDP), which is found in all PGN-containing bacteria. Downstream signaling of Nod receptors activates the NF-kB pathway and leads to the release of a variety of proinflammatory cytokines and antimicrobial compounds.30,31 Protein D4HAV7 is a type of Christie-Atkins-Munch-Petersen (CAMP) factor. CAMP factor is considered a pore-forming toxin. It binds to immunoglobulin G and M classes and then degrades and invades host cells.32 The CAMP factor can trigger inflammation in keratinocytes and macrophages and induce cell death of sebocytes in sebaceous glands. Protein D4HCW2 belongs to HtaA domain protein. HtaA, a heme-binding protein, participates in the iron uptake system in C. acnes.33 Iron is involved in many important metabolic processes and is a key component of pathogenicity. Iron participates in single electron transfer and is capable of generating toxic reactive oxygen species (ROS), which may cause cell damage and inflammation in the host environment.34 Interestingly, although the fold changes were not statistically significant, the expression levels of NlpC/P60, CAMP factor, and Hta domain protein were all uniformly higher in the resistant strains compared with the sensitive strains. These findings suggest that the resistant strains have higher potential to cause host inflammation and elicit a worse clinical acne manifestation compared with sensitive strains. We need to increase the experimental sample numbers and conduct in vitro and in vivo experiments to further confirm this hypothesis.
To identify the multidrug resistance mechanism of C. acnes, we further analyzed D4HCT5, which was significantly higher in the resistant C. acnes-derived MVs than in the sensitive MVs (p<0.05). D4HCT5 belongs to FtsZ, which is highly conserved in bacteria. FtsZ is considered an essential GTPase and structurally belongs to the cytoskeletal family.34,35 During cell division, FtsZ forms a Z-ring structure at the mid-cell and functions as a scaffold to recruit other cell division proteins to form the division polymer. FtsZ promotes cell division and leads to the proliferation of bacteria, thus reducing antibiotic efficacy. Cells depleted of functional FtsZ cannot divide and will eventually lyse.36 To address antibiotic resistance, many inhibitors targeting FtsZ have been studied extensively in recent years. Further studies are needed to clarify the role of FtsZ in the multidrug resistance of C. acnes and to explore the effect of FtsZ inhibitors in treating resistant C. acnes.
Conclusion
We analyzed the proteome of MVs from four clinically sensitive C. acnes isolates and three resistant isolates. According to the results of the proteome study, C. acnes-derived MVs are widely involved in bacterial physiology, virulence, and host inflammation. In our study, we detected several lipases, NlpC/P60, CAMP factor, and Hta domain protein, that serve as virulence factors in C. acnes-derived MVs. The levels of two lipases (D4HEV8, D4H9R7) and FtsZ were significantly higher in the resistant C. acnes-derived MVs compared with sensitive clusters (p<0.05). These observations give us important suggestions for the factors in C. acnes-derived MVs that might contribute to altered cutaneous lipid compositions and influence distinct inflammatory responses in acne patients; FtsZ, serving as an essential cell division protein, might be a promising target for treating antibiotic-resistant C. acnes.
Prior Presentation
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD025460.
Reviewer accountant details:
username: [email protected]; password: 0ftQcnu3
Acknowledgments
The authors gratefully acknowledge the financial support from the National Natural Science Foundation of China (Grant No. 81803154 and Grant No. 82173444).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Haurat MF, Elhenawy W, Feldman MF. Prokaryotic membrane vesicles: new insights on biogenesis and biological roles. Biol Chem. 2015;396:95–109. doi:10.1515/hsz-2014-0183
2. Kim SW, Park SB, Im SP, et al. Outer membrane vesicles from β-lactam-resistant Escherichia coli enable the survival of β-lactam-susceptible E. coli in the presence of β-lactam antibiotics. Sci Rep. 2018;8:5402. doi:10.1038/s41598-018-23656-0
3. Kulkarni HM, Nagaraj R, Jagannadham MV. Protective role of E. coli outer membrane vesicles against antibiotics. Microbiol Res. 2015;181:1–7. doi:10.1016/j.micres.2015.07.008
4. Klimentová J, Stulík J. Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiol Res. 2015;170:1–9. doi:10.1016/j.micres.2014.09.006
5. Toyofuku M, Nomura N, Eberl L. Types and origins of bacterial membrane vesicles. Nat Rev Microbiol. 2019;17:13–24. doi:10.1038/s41579-018-0112-2
6. Lee EY, Choi DY, Kim DK, et al. Gram-positive bacteria produce membrane vesicles: proteomics-based characterization of Staphylococcus aureus-derived membrane vesicles. Proteomics. 2009;9:5425–5436. doi:10.1002/pmic.200900338
7. Jiang Y, Kong Q, Roland KL, Curtiss R
8. Lee JH, Choi CW, Lee T, Kim SI, Lee JC, Shin JH. Transcription factor σB plays an important role in the production of extracellular membrane-derived vesicles in Listeria monocytogenes. PLoS One. 2013;8:e73196. doi:10.1371/journal.pone.0073196
9. Rivera J, Cordero RJ, Nakouzi AS, Frases S, Nicola A, Casadevall A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc Natl Acad Sci U S A. 2010;107:19002–19007. doi:10.1073/pnas.1008843107
10. Schrempf H, Koebsch I, Walter S, Engelhardt H, Meschke H. Extracellular Streptomyces vesicles: amphorae for survival and defence. Microb Biotechnol. 2011;4:286–299. doi:10.1111/j.1751-7915.2011.00251.x
11. Dréno B, Pécastaings S, Corvec S, Veraldi S, Khammari A, Roques C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: a brief look at the latest updates. J Eur Acad Dermatol Venereol. 2018;32(Suppl 2):5–14. doi:10.1111/jdv.15043
12. Lee YB, Byun EJ, Kim HS. Potential role of the microbiome in acne: a comprehensive review. J Clin Med. 2019;8(7):987.
13. Xu H, Li H. Acne, the skin microbiome, and antibiotic treatment. Am J Clin Dermatol. 2019;20:335–344. doi:10.1007/s40257-018-00417-3
14. Choi EJ, Lee HG, Bae IH, et al. Propionibacterium acnes-derived extracellular vesicles promote acne-like phenotypes in human epidermis. J Invest Dermatol. 2018;138:1371–1379. doi:10.1016/j.jid.2018.01.007
15. Schaar V, Nordström T, Mörgelin M, Riesbeck K. Moraxella catarrhalis outer membrane vesicles carry β-lactamase and promote survival of Streptococcus pneumoniae and Haemophilus influenzae by inactivating amoxicillin. Antimicrob Agents Chemother. 2011;55:3845–3853. doi:10.1128/AAC.01772-10
16. Ma Y, Zhang N, Wu S, Huang H, Cao Y. Antimicrobial activity of topical agents against Propionibacterium acnes: an in vitro study of clinical isolates from a hospital in Shanghai, China. Front Med. 2016;10:517–521. doi:10.1007/s11684-016-0480-9
17. Tian Y, Ma L, Gong M, et al. Protein profiling and sizing of extracellular vesicles from colorectal cancer patients via flow cytometry. ACS Nano. 2018;12:671–680. doi:10.1021/acsnano.7b07782
18. Zhang Y, Jin X, Liang J, et al. Extracellular vesicles derived from ODN-stimulated macrophages transfer and activate Cdc42 in recipient cells and thereby increase cellular permissiveness to EV uptake. Sci Adv. 2019;5:eaav1564. doi:10.1126/sciadv.aav1564
19. Zhu Q, Ling X, Yang Y, et al. Embryonic stem cells-derived exosomes endowed with targeting properties as chemotherapeutics delivery vehicles for glioblastoma therapy. Adv Sci. 2019;6:1801899. doi:10.1002/advs.201801899
20. Bek-Thomsen M, Lomholt HB, Scavenius C, Enghild JJ, Brüggemann H, McDowell A. Proteome analysis of human sebaceous follicle infundibula extracted from healthy and acne-affected skin. PLoS One. 2014;9(9):e107908. doi:10.1371/journal.pone.0107908
21. Josse G, Mias C, Le Digabel J, et al. High bacterial colonization and lipase activity in microcomedones. Exp Dermatol. 2020;29:168–176. doi:10.1111/exd.14069
22. Snodgrass RG, Huang S, Choi IW, Rutledge JC, Hwang DH. Inflammasome-mediated secretion of IL-1β in human monocytes through TLR2 activation; modulation by dietary fatty acids. J Immunol. 2013;191:4337–4347. doi:10.4049/jimmunol.1300298
23. Coenye T, Peeters E, Nelis HJ. Biofilm formation by Propionibacterium acnes is associated with increased resistance to antimicrobial agents and increased production of putative virulence factors. Res Microbiol. 2007;158:386–392. doi:10.1016/j.resmic.2007.02.001
24. Kistowska M, Gehrke S, Jankovic D, et al. IL-1β drives inflammatory responses to Propionibacterium acnes in vitro and in vivo. J Invest Dermatol. 2014;134:677–685. doi:10.1038/jid.2013.438
25. Higaki S, Kitagawa T, Kagoura M, Morohashi M, Yamagishi T. Correlation between Propionibacterium acnes biotypes, lipase activity and rash degree in acne patients. J Dermatol. 2000;27:519–522. doi:10.1111/j.1346-8138.2000.tb02219.x
26. Kim HJ, Lee BJ, Kwon AR. The grease trap: uncovering the mechanism of the hydrophobic lid in Cutibacterium acnes lipase. J Lipid Res. 2020;61:722–733. doi:10.1194/jlr.RA119000279
27. Davis KM, Weiser JN, Maurelli AT. Modifications to the peptidoglycan backbone help bacteria to establish infection. Infect Immun. 2011;79(2):562–570. doi:10.1128/IAI.00651-10
28. Humann J, Lenz LL. Bacterial peptidoglycan degrading enzymes and their impact on host muropeptide detection. J Innate Immun. 2009;1:88–97. doi:10.1159/000181181
29. Mukherjee T, Hovingh ES, Foerster EG, Abdel-Nour M, Philpott DJ, Girardin SE. NOD1 and NOD2 in inflammation, immunity and disease. Arch Biochem Biophys. 2019;670:69–81. doi:10.1016/j.abb.2018.12.022
30. Caruso R, Warner N, Inohara N, Núñez G. NOD1 and NOD2: signaling, host defense, and inflammatory disease. Immunity. 2014;41:898–908. doi:10.1016/j.immuni.2014.12.010
31. Frirdich E, Gaynor EC. Peptidoglycan hydrolases, bacterial shape, and pathogenesis. Curr Opin Microbiol. 2013;16:767–778. doi:10.1016/j.mib.2013.09.005
32. Wang Y, Hata TR, Tong YL, et al. The anti-inflammatory activities of Propionibacterium acnes CAMP factor-targeted acne vaccines. J Invest Dermatol. 2018;138:2355–2364. doi:10.1016/j.jid.2018.05.032
33. Nairz M, Weiss G. Iron in infection and immunity. Mol Aspects Med. 2020;75:100864. doi:10.1016/j.mam.2020.100864
34. Haranahalli K, Tong S, Ojima I. Recent advances in the discovery and development of antibacterial agents targeting the cell-division protein FtsZ. Bioorg Med Chem. 2016;24:6354–6369. doi:10.1016/j.bmc.2016.05.003
35. Kusuma KD, Payne M, Ung AT, Bottomley AL, Harry EJ. FtsZ as an antibacterial target: status and guidelines for progressing this avenue. ACS Infect Dis. 2019;5:1279–1294. doi:10.1021/acsinfecdis.9b00055
36. Hurley KA, Santos TM, Nepomuceno GM, Huynh V, Shaw JT, Weibel DB. Targeting the bacterial division protein FtsZ. J Med Chem. 2016;59:6975–6998. doi:10.1021/acs.jmedchem.5b01098
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.