")
Back to Journals » OncoTargets and Therapy » Volume 13
Circular RNA circ_0006282 Contributes to the Progression of Gastric Cancer by Sponging miR-155 to Upregulate the Expression of FBXO22
Authors He Y, Wang Y, Liu L, Liu S, Liang L , Chen Y, Zhu Z
Received 22 August 2019
Accepted for publication 2 December 2019
Published 31 January 2020 Volume 2020:13 Pages 1001—1010
DOI https://doi.org/10.2147/OTT.S228216
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
Yiren He, 1,* Yinfeng Wang, 2,* Liu Liu, 1 Shaojun Liu, 1 Lichuan Liang, 1 Yinan Chen, 3 Zhiqiang Zhu 1
1Department of General Surgery, The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei, Anhui 230001, People’s Republic of China; 2Department of Critical Care Medicine, The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei, Anhui 230001, People’s Republic of China; 3Department of Gastrointestinal Surgery, Cancer Hospital, The First Affiliated Hospital of Xiamen University, Teaching Hospital of Fujian Medical University, Xiamen 361000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiqiang Zhu
Department of General Surgery, The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, No. 17, Lujiang Road, Luyang District, Anhui, Hefei 230001, People’s Republic of China
Email [email protected]
Yinan Chen
Department of Gastrointestinal Surgery, Cancer Hospital, The First Affiliated Hospital of Xiamen University, Teaching Hospital of Fujian Medical University, 55 Zhenhai Rd, Siming, Xiamen 361000, People’s Republic of China
Email [email protected]
Background: There is increasing evidence that circular RNAs (circRNAs) play an important role in human cancers. As a newly identified human circular RNA, circ_0006282 is abnormally expressed in several types of cancers and promotes the development of cancers. However, the expression and function of circ_0006282 in gastric cancer (GC) remain unclear.
Methods: The expression of circ_0006282 in cancer tissues and adjacent non-cancer tissues was detected by quantitative real-time polymerase chain reaction (qRT-PCR) method, and the relationship between circ_0006282 expression and clinicopathological parameters was analyzed. After knockdown of circ_0006282 by RNA interference in GC cells, CCK-8 assay, colony formation and transwell assays were conducted to examine the effects of circ_0006282 on GC cells. The influence of circ_0006282 on tumor growth in vivo was assessed in a xenograft model. Furthermore, regulatory relationship between circ_0006282, miR-155 and FBXO22 was detected by luciferase assay, qRT-PCR and Western blot.
Results: The expression of circ_0006282 in GC tissues was significantly higher than its adjacent non-cancer tissues and over-expression of circ_0006282 was associated with tumor size, lymph nodes metastasis and TNM stage, but no obvious links with other pathological parameters. Knockdown of circ_0006282 inhibited the proliferation and metastasis ability of GC cells in vitro and suppressed the tumor growth in vivo. Furthermore, mechanistic investigations suggested that circ_0006282 served as a competing endogenous RNA (ceRNA) of miR-155. Moreover, FBXO22 was identified as the functional target of miR-155 and down-expression of circ_0006282 inhibited FBXO22 expression. Rescue assays also demonstrated that the oncogenic function of circ_0006282 is partly attributed to its regulation on miR-155/FBXO22 axis.
Conclusion: Our findings indicated that over-expression of circ_0006282 down‑regulated miR-155 to activate the expression of FBXO22, thus promoting proliferation and metastasis of GC cells, which provides a promising therapeutic target for GC treatment.
Keywords: circ_0006282, gastric cancer, proliferation, invasion, miR-155, FBXO22
Background
Gastric cancer (GC) is a digestive tract malignant tumor originating from gastric mucosal epithelial cells and its morbidity and mortality have been at the forefront of the world’s malignant tumors. In recent years, the incidence and mortality of gastric cancer have decreased globally, but it is still the second leading cause of death in cancer, second only to lung cancer, and it is an important cause of threats to people’s health and quality of life.1 The occurrence and development of gastric cancer is a multi-factor, multi-step complex process, closely related to the regulation of multiple genes. At present, the treatment of gastric cancer is still based on surgery, combined with radiotherapy, chemotherapy and drug treatment. However, the treatment effect is not optimistic, and the patient’s 5-year survival rate is less than 30%.2,3 Thus, improving the early diagnosis of GC and developing new therapeutic strategies based on a greater understanding of the biogenesis of GC are urgent research goals.
In recent years, studies have found that the abnormal expression of circular RNA (circRNA) is involved in the occurrence and development of a variety of tumors, and is expected to become a new target for tumor diagnosis and treatment.4,5 CircRNA is a closed, circular non-coding RNA molecule formed by covalent bonding between the 3ʹ end and the 5ʹ end by reverse splicing. It is widely present in organisms and is more stable than linear RNA.6 CircRNA can act as a “microRNA (miRNA) sponge” involved in the regulation of gene expression or by binding proteins to affect its function, and some circRNA can be translated into short peptides to function.7,8 Numerous reports indicate that circRNA is involved in the occurrence and development of tumors, including GC. For instance, circRNA-LARP4 can competitively bind to miR-424-5p through sponge action to regulate the expression of LATS1 gene and finally inhibit the proliferation and invasion of gastric cancer cells.9 Compared with the healthy control group, the serum of 104 patients with GC was detected. The expression level of circ-0000190 in serum was significantly down-regulated, and its specificity was significantly better than CEA and CA199, and the expression level of circ-0000190 in patients with CEA-positive serum was significantly elevated.10 At the same time, it was found that the overall survival and disease-free survival of patients with low expression of circPVTl were significantly shorter than those with high expression of circPVTl.11
Circ_0006282 was a newly identified circRNA. Shao et al reported that circ_0006282 was one of the top 10 upregulated circRNAs by circRNA microarray.12 However, to our best knowledge, the biological function and the mechanism of circ_0006282 in cancers including GC have not yet been investigated. We detected circ_0006282 expression in GC cell lines and GC tissues. Moreover, we identified the function of circ_0006282 in GC cells by applying loss-of-function approaches in vitro and in vivo.
Materials and Methods
Human Specimens
The tissue specimens used in this study were from the Department of General Surgery of The First Affiliated Hospital of USTC from October 2015 to March 2019. All gastric cancer tissue specimens and adjacent non-tumor tissues were confirmed by postoperative pathological examination and had complete clinical and pathological data. No chemotherapy or radiotherapy was given before surgery. All patients or guardians (a small number of illiterate patients designate their guardians to sign) provided written informed consent before enrollment, and the study protocol was approved by the Ethics Committee of the First Affiliated Hospital of USTC.
Cell Lines and Transfection
Human GC cell line (BGC-823, MKN-45, AGS, HGC-27) and human gastric mucosal epithelial immortalized cell line GES-1 were purchased from Cell Bank of Chinese Academy of Sciences (Shanghai, China). The cells were cultured in RPMI-l640 cell culture medium containing 10% calf serum at 37 ° C and 5% CO2, and cells in the logarithmic growth phase were used for the experiment.
BGC-823 and MKN-45 cells cultured on the six-well plate were transfected with siRNAs of si-circ_0006282 1# and 2#, si-FBXO22, miR-155 mimics/inhibitor, or negative control, which were obtained from RiboBio (Guangzhou, China). The siRNA sequences were as follows: si- circ_0006282 1#: 5′‐GCCCAGGAAACTACTAAAGTT‐3′; si-circ_0006282 2#: 5′‐GAGTGGCCCAGGAAACTACTA‐3′; siFBXO22: 5′‐GGUGGGAGCCAGUAAUUAUTT‐3′. Cell transfection was performed using Lipofectamine 2000 Reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions.
RNA Extraction and Quantitative Real-Time PCR
Total RNA was extracted from tissues and cells using Trizol (Invitrogen, Carlsbad, CA). The standard procedure for RT-qPCR was performed as previously described.13 We used GAPDH or U6 as internal controls. Relative transcription alteration was evaluated as 2−ΔΔCt. The primer sequences were as follows: hsa_circ_0006282 (Forward, 5ʹ-AGGCACGATAAAAGCCATGT-3ʹ; Reverse, 5ʹ-GGTCCTTCACAGCCACCATA-3ʹ), miR-155 (Forward, 5ʹ-AACTTGTAAACTCCCTCGACTG-3ʹ; Reverse, 5ʹ-CCTTACGTGACCTGGAGTCG-3ʹ), FBXO22 (Forward, 5ʹ- CGGAGCACCTTCGTGTTG-3ʹ; Reverse, 5ʹ- CACACACTCCCTCCATAAGCG-3ʹ), GAPDH (Forward, 5ʹ- GGACCTGACCTGCCGTCTAG-3ʹ; Reverse, 5ʹ- GTAGCCCAGGATGCCCTTGA-3ʹ) and U6 (Forward, 5ʹ-GCTTCGGCAGCACATATACTAAAAT-3ʹ; Reverse, 5ʹ-CGCTTCACGAATTTGCGTGTCAT-3ʹ).
circRNA Confirmatory Assay
Total RNA (5 μg) was kept in 3 U/μg of RNase R (Epicentre Biotechnologies, Madison, WI, USA) for 15 min at 37°C. Two RNase R digestion reactions were performed according to previously published methods.14 The agarose gels with TE buffer (Thermo Scientific, Waltham, MA, USA) were used to detect the genomic DNA (gDNA) and cDNA PCR products of circ_0006282.
Western Blot
Total protein of cells was extracted in RIPA buffer (Beyotime) containing 1% protease inhibitor cocktail and 1% PMSF. Protein samples were separated by 10% SDS-PAGE and transferred to polyvinylidene difluoride membrane (Millipore, USA). The membranes were blocked with 5% non-fat milk in Tris-buffered saline and then incubated with primary antibodies at 4°C overnight. After washing in PBST 3 times, the membranes were then incubated in HRP-conjugated secondary antibodies. Proteins were detected by an enhanced chemiluminescence detection system (Amersham Bioscience, Piscataway, NJ, USA).
Cell Proliferation Assay and Colony Formation Assay
Different groups of GC cells at a concentration of 1.5 × 103 cells/200μL were seeded and cultured in 96-well plates. CCK-8 reagents (Dojindo, Kumamoto, Japan) were added on 1, 2, 3, 4, and 5 days, respectively, and their OD450 values were determined using a Microplate Autoreader (Bio-Rad, Hercules, CA, USA). For the colony formation assay, 1×103 GC cells were seeded into 6-well plates and cultured in medium containing 10% calf serum, and the culture medium was changed every 3 to 4 days. After 2 weeks of continuous culture, the cells were fixed with methanol and stained with crystal violet and washed for photographing.
Transwell Assay
For migration assay, 1×105 cells were suspended in serum-free RPMI-1640 and plated in a transwell chamber previously placed in a 24-well plate. For the invasion assay, the difference was that matrigel (BD Bioscience, CA, USA) was pre-coated on the bottom of the transwell chamber, and then 1×105 serum-free RPMI-1640 cells were added to the chamber. For both assays, culture medium containing 10% calf serum was added as a chemoattractant to the lower compartment of the chamber. After 24 hrs of incubation, the chamber was stained with a 0.5% crystal violet solution in methanol for 15 mins, and then the cells that did not penetrate to the lower layer of the chamber were wiped off with a cotton swab and soaked in PBS. Finally, the cells in the lower chamber were counted under an inverted microscope, and five views were randomly selected to calculate the average number of cells (200 x).
Xenograft Model
Animal experiment complied with the National Institutes of Health guide for the care and use of laboratory animals. The experiments were approved by the First Affiliated Hospital of USTC and conducted in accordance with the Institutional Animal Care and Use Committee guidelines of the First Affiliated Hospital of USTC. Cells (1×106/100 mL/site) were injected into the flank of 4-week-old female BALB/c nude mice. Tumor growth curve was determined by measuring tumor volumes every week. After the sacrifice, the animals were harvested, measured, and weighed and Ki67 expression was determined by the immunohistochemistry method as previously described.13
Nuclear-Cytoplasmic Fractionation
5×106 BGC-823 and MKN-45 cells were rinsed twice in PBS and suspended in cell fraction buffer. Followed by incubation on ice for 10 min, cells were centrifuged. The upper solution and the nuclear pellet were collected to extract RNA in cell disruption buffer. At last, the isolated RNA was measured by qRT-PCR, normalizing to GAPDH (cytoplasmic control) and U6 (nuclear control).
Luciferase Reporter Assay
Circular RNA Interactome (https://circinteractome.nia.nih.gov/) and Starbase (http://starbase.sysu.edu.cn) were used to predict binding sites between circular RNA, miRNA and target gene. The reporter plasmids (pGL3-Firefly_Luciferase-Renilla_Luciferase containing the circ_0006282 sequence or a mutant sequence and pGL3-Firefly_Luciferase-Renila_Luciferase containing the FBXO22 sequence or a mutant sequence) were designed by Shanghai Genechem company. Cells were seeded in 24-well plates 1 day prior to transfection with wild-type or mutant-type dual luciferase plasmid and the indicated miRNAs. Cells were then transfected with Lipofectamine 2000 according to the manufacturer’s instructions. After incubation for 48 h, luciferase activity was analyzed using a DualLuciferase Reporter Assay System (Promega, Madison, WI, USA).
Statistical Analysis
All statistical data analyses were analyzed with SPSS 20.0 software (IBM, Chicago, IL, USA). Results are shown as mean ± SD. The significance of the differences was determined by Student’s t-test or one-way ANOVA followed by Tukey’s post hoc test as appropriate. P < 0.05 was considered as significant.
Results
circ_0006282 Is Upregulated in GC Tissues and Cell Lines
circ_0006282 was one member of the circRNAs as its expression had no significant change when being treated by RNase R (Figure 1A). At the same time, the cDNA of circ_0006282 was amplified, while the gDNA did not produce amplification with divergent primers, confirming that circ_0006282 was a circRNA (Figure 1A). We first examined the expression of circ_0006282 in 84 pairs of GC tissues and adjacent non-tumor tissues using qRT-PCR. The results showed that the expression level of circ_0006282 in GC tissues was significantly higher than that in non-tumor tissues (P < 0.05, Figure 1B). We also analyzed the relationship between circ_0006282 expression and clinical pathological parameters (Table 1). We also found that the expression of circ_0006282 in patients with positive lymph node metastasis and late staging was higher than that in patients with negative lymph node metastasis and early stage (P < 0.05, Figure 1C and D). Besides, we analyzed the expression of circ_0006282 in GC cell lines and found that its expression increased significantly in GC cell lines (P < 0.05, Figure 1E), BGC-823 and MKN-45 cell lines were selected to down-regulate circ_0006282 expression and used for biological behavioral studies (P < 0.05, Figure 1F and G).
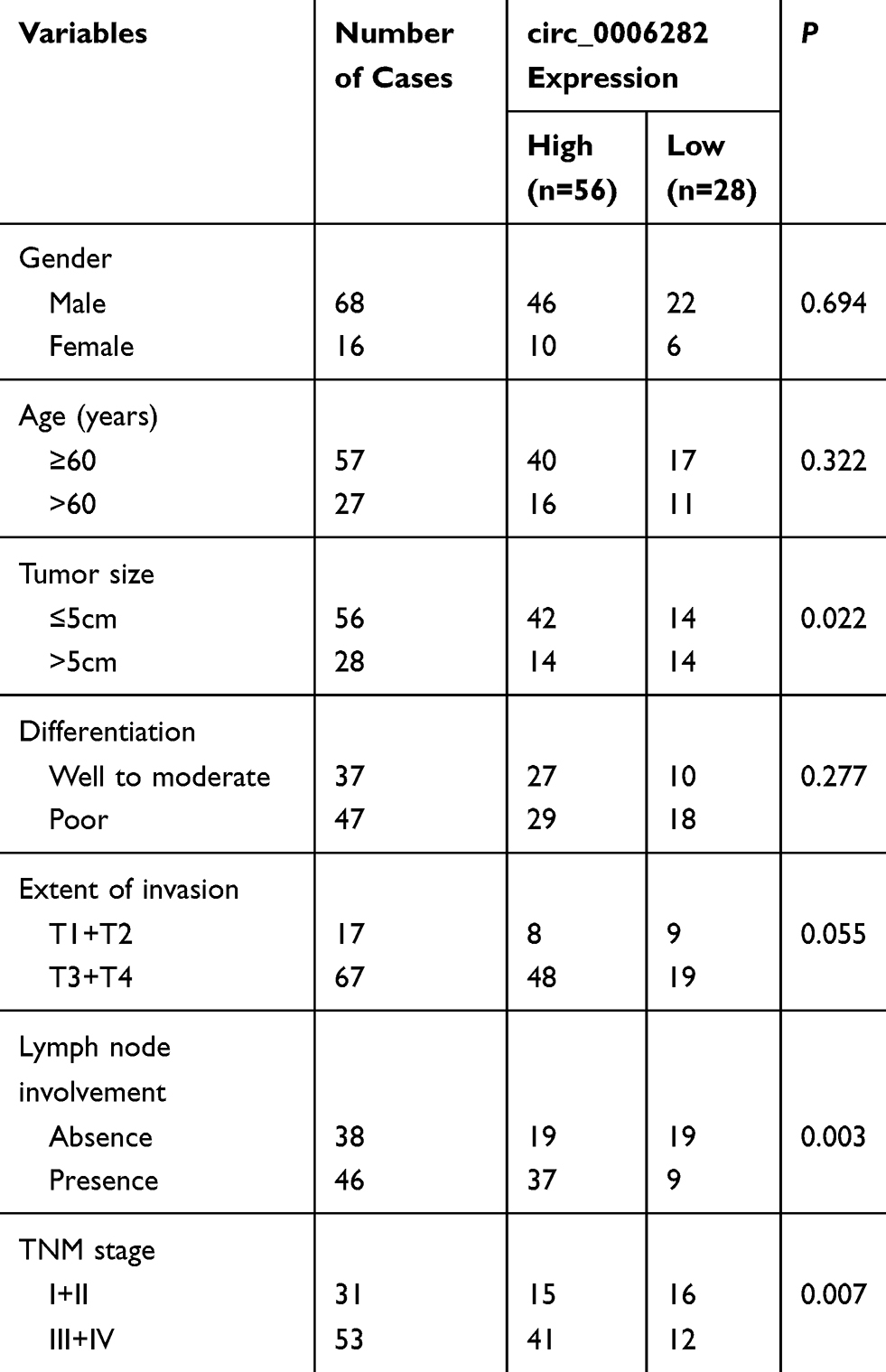 |
Table 1 Association Between circ_0006282 Expression and Clinicopathological Factors of GC Patients |
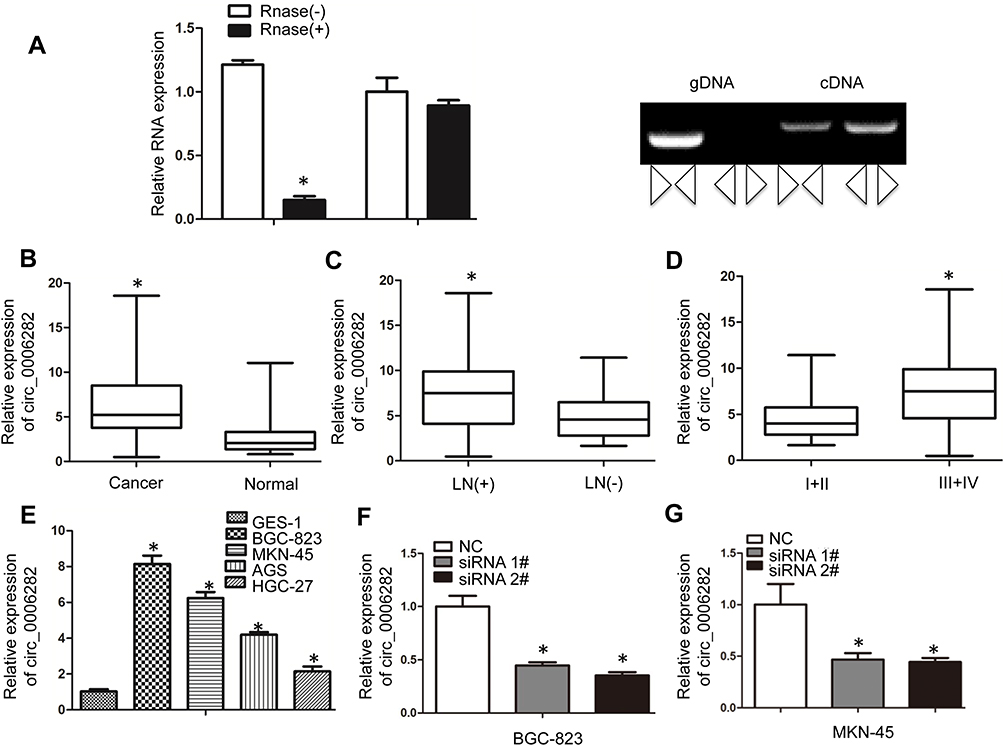 |
Figure 1 Circ_0006282 is upregulated in GC tissues and cell lines. (A) The relative expression of circ_0006282 had no significant change after RNase R digestion compared with RNase R(−). The cDNA of circ_0006282 was amplified with divergent primers, while the gDNA was not amplified. (B) qRT-PCR analysis was used to test circ_0006282 expression in 84 GC tissues and their paired non-tumor tissues. (C) qRT-PCR analysis showed increased expression of circ_0006282 in lymph node metastasis GC tissues compared to non-metastatic tissues. (D) The upregulated circ_0006282 indicated the advanced tumor stage of GC. (E) circ_0006282 expression was upregulated in GC cell lines. (F and G) circ_0006282 expression was decreased by transfection with two independent siRNAs in BGC-823 cells and MKN-45 cells. *P < 0.05. |
circ_0006282 Promotes the Malignant Phenotype of GC Cells in vitro and in vivo
CCK8 assay was performed to investigate the effect of circ_0006282 on GC cells proliferation. As shown in Figure 2A and B, down-regulation of circ_0006282 in BGC-823 and MKN-45 cells can significantly inhibit the proliferation of gastric cancer cells. Knockdown of circ_0006282 led to decreased colonies in colony formation assay (P < 0.05, Figure 2C–F). We performed a transwell assay to examine the effect of circ_0006282 on the motility of GC cells and found that circ_0006282 under-expression significantly inhibited the migration and invasion ability of BGC-823 and MKN-45 cells (P < 0.05, Figure 3A–D). Furthermore, the effects of circ_0006282 dysregulation on tumorigenicity were tested in nude mice. As illustrated in Figure 4A–C, circ_0006282 silencing dramatically delayed GC growth as indicated by reduced tumor’s weights and volume. Moreover, we also found that Ki-67 staining percentage was less in circ_0006282 silencing tumor samples compared with the control group (P < 0.05, Figure 4D and E).
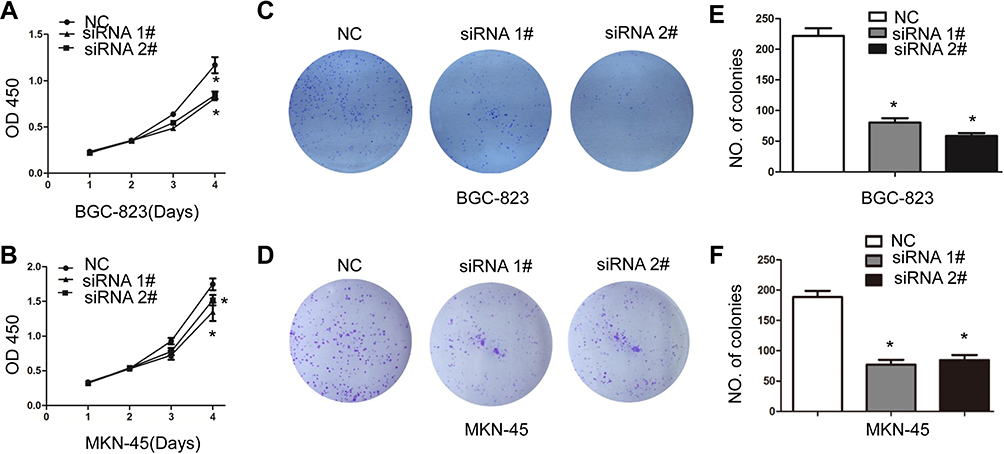 |
Figure 2 Circ_0006282 silencing inhibits the proliferation of GC cells. (A and B) circ_0006282 silencing inhibited the proliferation of BGC-823 and MKN-45 cells shown by CCK8. (C and D) Representative photographs of plate colony formation of BGC-823 and MKN-45 cells infected with circ_0006282 siRNA and control vector. (E and F) Quantitative analysis of plate colony formation of BGC-823 and MKN-45 cells infected with circ_0006282 siRNA and control vector. *P < 0.05. |
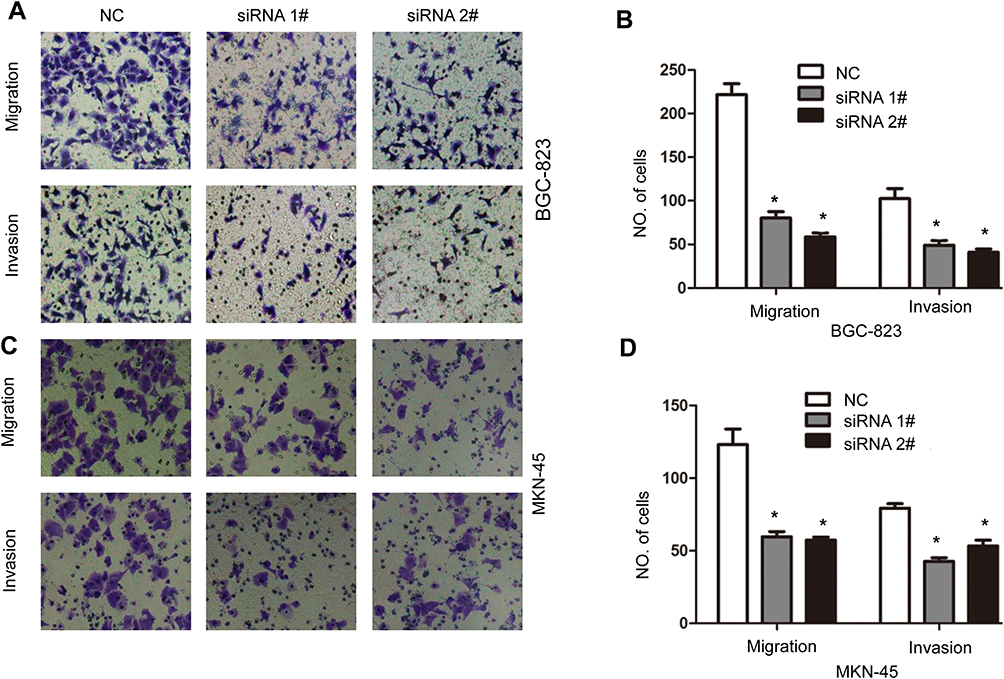 |
Figure 3 Circ_0006282 silencing inhibits the migration and invasion of GC cells. (A) Representative photographs of migration and invasion of BGC-823 infected with circ_0006282 siRNA and control vector. (B) Quantitative analysis of migration and invasion of BGC-823 cells infected with circ_0006282 siRNA and control vector. (C) Representative photographs of migration and invasion of MKN-45 infected with circ_0006282 siRNA and control vector. (D) Quantitative analysis of migration and invasion of MKN-45 cells infected with circ_0006282 siRNA and control vector. *P < 0.05. |
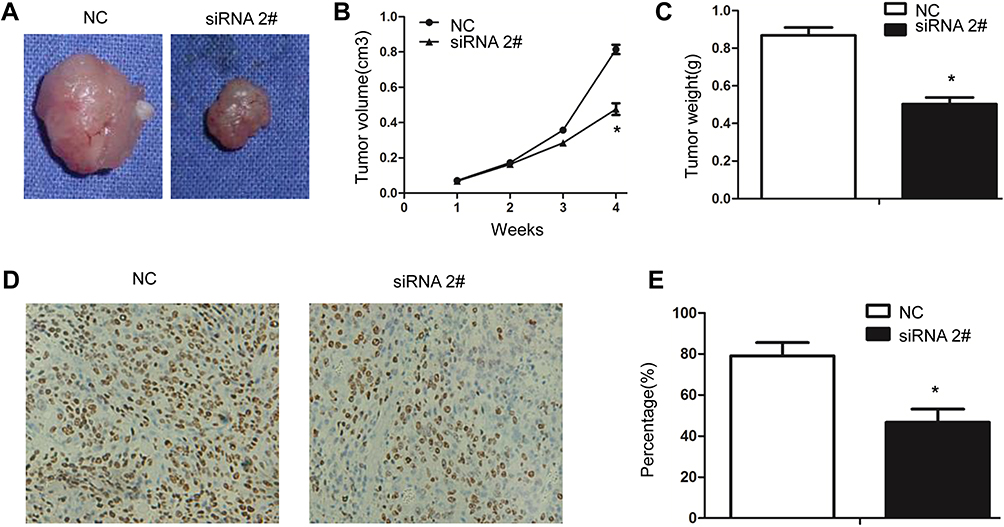 |
Figure 4 Circ_0006282 silencing inhibits the subcutaneous tumor formation in vivo. (A) Xenograft tumor models showed that tumors grown from circ_0006282 down-regulated BGC-823 cells were smaller than control groups. (B) Tumor volume was determined every week for total 4 weeks. (C) Tumor weight was measured when mice were sacrificed at week 4. (D and E) The tumor sections from circ_0006282 silencing group and control group underwent IHC staining using antibodies against Ki-67. *P < 0.05. |
circ_0006282 Promoted FBXO22 Expression via Serving as a ceRNA of miR-155
Based on bioinformatics analysis, we found that circ_0006282 may regulate miR-155 by sponge action. At the same time, further predictions suggest that miR-155 may directly target FBXO22 (Figure 5B). To demonstrate this hypothesis, we first examined the expression of circ_0006282 in the cytoplasm and nucleus and found that circ_0006282 is mainly expressed in the cytoplasm of cells (Figure 5A). Then, we constructed a dual-luciferase reporter plasmid system. The luciferase reporter assay showed that the miR-155 mimics significantly inhibited the luciferase activity of circ_0006282-WT and FBXO22-WT. However, this inhibitory effect apparently disappeared after mutation of its binding site, demonstrating the interaction between miR-155 and circ_0006282 or FBXO22 (Figure 5C and D). qRT-PCR results showed that knockdown of circ_0006282 expression in BGC-823 cells promoted the expression of miR-155 (Figure 5E), while miR-155 mimics inhibited FBXO22 expression and we found that circ_0006282 silencing can inhibit the expression of FBXO22, and this inhibition can be alleviated by miR-155/inhibitor (Figure 5F and G). Western blotting also demonstrated that circ_0006282 can regulate FBXO22 expression through miR-155 in GC cells (Figure 5H and I). Moreover, we found that miR-155 expression was negatively correlated with circ_0006282 or FBXO22 expression in GC tissues (Figure 5J and K). These data reveal that circ_0006282 interacts with miR-155/FBXO22 axis in gastric cancer.
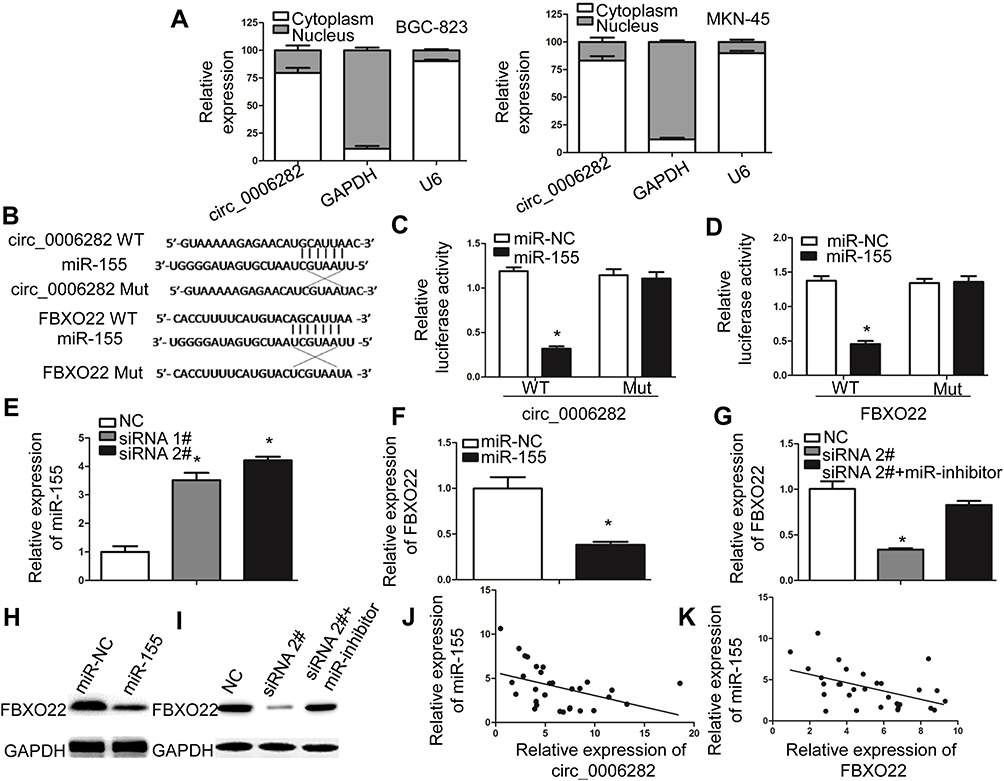 |
Figure 5 Circ_0006282 regulates miR-155/FBXO22 axis in GC cells. (A) The nuclear or cytoplasmic circ_0006282 in BGC-823 and MKN-45 cells. circ_0006282 was mainly expressed in the cytoplasm of cells. (B) Schematic graph of the putative binding sites of miR-155 in the circ_0006282 or FBXO22 3’UTR. circ_0006282 mut or FBXO22 mut indicates the mutation of circ_0006282 or FBXO22 in miR-155 binding sites. (C and D) Luciferase reporter assay showed that the luciferase activity of either circ_0006282-WT or FBXO22 3’-UTR-WT was inhibited by miR-155 mimics. (E) miR-155 expression was upregulated in BGC-823 cells after circ_0006282 silencing. (F and H) FBXO22 mRNA and protein expression was downregulated in BGC-823 cells after transfection with miR-155 mimics. (G and I) circ_0006282 regulated FBXO22 mRNA and protein expression in BGC-823 cells through inhibiting miR-155. (J) The expression of circ_0006282 was reversely correlated with miR-155 expression in GC tissues. (K) The expression of miR-155 was negatively correlated with FBXO22 expression in GC tissues. *P < 0.05. |
circ_0006282/miR-155/FBXO22 Axis Regulates GC Progression
To further confirm the role of circ_0006282/miR-155/FBXO22 signaling in the progression of gastric cancer, we knocked down the expression of circ_0006282 in BGC-823 cells and simultaneously transfected miR-155 inhibitor or (and) FBXO22 siRNA. Then, FBXO22 expression was tested by qRT-PCR and western-blot, and the results showed that circ_0006282 can affect the expression of FBXO22 by the regulation of miR-155 (Figure 6A and B). Then, CCK8, colony formation and transwell assay were conducted to analyze proliferation and metastasis ability after GC cells transfected with described plasmids. As shown in Figure 6C–F, addition of miR-155 inhibitor significantly reversed the inhibitory effect of circ_0006282 siRNA on proliferation, colony formation, migration and invasion of GC cells. In addition, knockdown of FBXO22 further reversed the biological effects of the simultaneous transfection of circ_0006282 siRNA and miR-155 inhibitor on GC cells. Taken together, circ_0006282/miR-155/FBXO22 signaling plays a critical role in regulating GC progression.
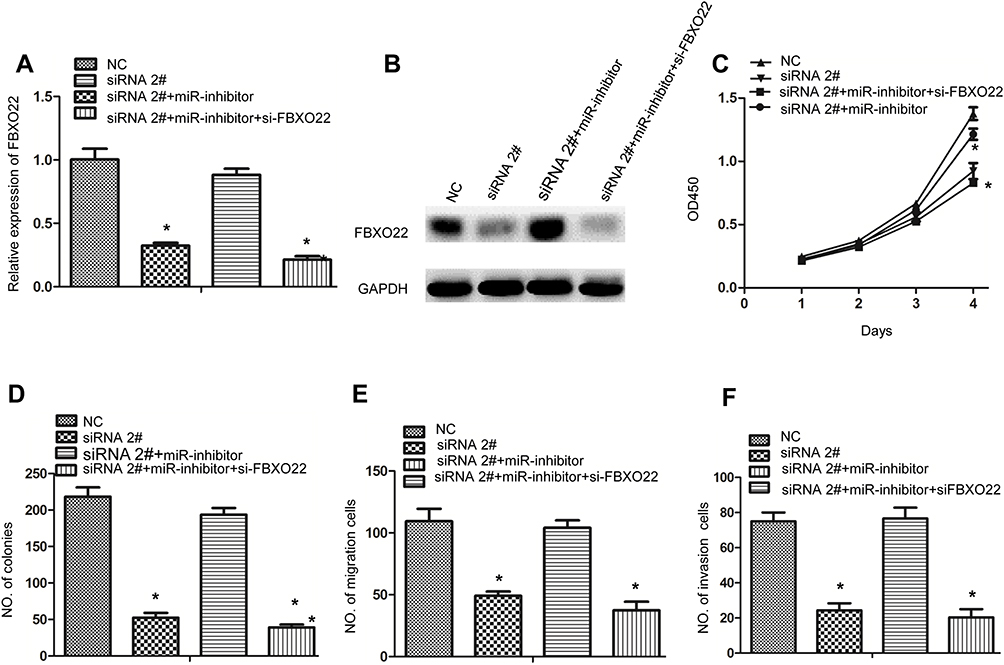 |
Figure 6 Circ_0006282 facilitates GC cell growth and invasion via upregulating FBXO22 expression by sponging miR-155. (A and B) Expression of FBXO22 was measured by qRT-PCR and Western blot after transfection with described vectors in BGC-823 cells. (C) CCK-8 assay was performed to analyze the cell proliferation of BGC-823 cells after transfection with described vectors. (D) Plate colony formation assay was performed to analyze the cell viability of BGC-823 cells after transfection with described vectors. (E and F) Transwell assay was performed to investigate cell migration and invasion of BGC-823 cells after with described vectors. *P < 0.05. |
Discussion
With the development of the economy, the diet structure has changed, the incidence of gastric cancer has also increased, and the mortality rate is high. Helicobacter pylori infection, gastric polyps, gastritis and other factors can lead to the onset of gastric cancer.15 According to epidemiological investigations, the risk factors for gastric cancer include both environmental and genetic aspects.16 Under the control of multiple genes, the gastric tissue undergoes malignant transformation and progresses to a tumor. CircRNA is a type of non-coding RNA widely present in eukaryotic cells. Unlike linear RNA, circRNA forms a covalently closed circular structure by trans-splicing, which makes it highly stable and biological conservation. Recent studies have shown that circRNA is abnormally expressed in a variety of tumor tissues, mainly through the regulation of proliferation, invasion and apoptosis of cancer cells as a sponge of miRNA, and is expected to become a new diagnostic marker and a potential therapeutic target.4,17 However, its role in gastric cancer is poorly understood. Circ_0006282 is a new type of circRNA that has never been studied. Not long ago, Shao Y et al found that the expression of circ_0006282 in gastric cancer tissue samples was significantly up-regulated compared with non-tumor tissue adjacent to the cancer by circRNA microarray.12 In this study, we found circ_0006282 was not only upregulated in GC tissues but also in a panel of GC cell lines compared with adjacent non-tumor tissues and GES-1, respectively. Furthermore, our results indicated that circ_0006282 could promote proliferation, migration and invasion in GC, suggesting circ_0006282 might be a potential therapeutic target for GC treatment.
Recent studies have shown that circRNAs can exert as a sponge on miRNAs to regulate the expression of downstream target genes of miRNAs and affect tumor occurrence and development.18–20 MiR-155 is an important member of the microRNAs family, and due to its versatile nature, miR-155 has attracted extensive attention and extensive research. A large number of studies have shown that miR-155 has a wide range of functions, involving many biological processes such as developmental differentiation of hematopoietic cells, developmental differentiation of immune cells, inflammatory response, immune response, muscle development, and fat differentiation.21–24 At the same time, miR-155 is highly expressed in various cancer tissues or cell lines such as liver cancer, lymphoma, breast cancer, pancreatic cancer and lung cancer, and is closely related to tumor occurrence, invasion and metastasis.25–28 Moreover, with the deepening of research, miR-155 is very likely to become a new tumor marker and a new target for tumor gene therapy. As a member of the F-box protein, FBXO22 can cause instability of the ubiquitinated complex, thereby promoting or inhibiting tumorigenesis and metastasis. The expression of FBXO22 is up-regulated in hepatocellular carcinoma and can promote the development of hepatocellular carcinoma by regulating the ubiquitination and degradation of p21.29 Similarly, in another study, FBXO22 was also found to promote the progression of hepatocellular carcinoma by regulating the ubiquitination and degradation of KLF4.30 At present, there is no research report on the relationship between circ_0006282, miR-155 and FBXO22. In this study, we demonstrated that circ_0006282 promotes FBXO22 expression and gastric cancer progression through sponging miR-155. Moreover, we observed that the expression of miR-155 was negatively correlated with that of circ_0006282 and FBXO22 in GC tissues.
Conclusion
In conclusion, we verified that circ_0006282 is highly expressed in GC tissues and GC cell lines. Meanwhile, circ_0006282 promotes the proliferation, migration and invasion of GC through regulating miR-155/FBXO22 signal pathway. Our findings demonstrated that circ_0006282 might be an effective therapeutic target for the treatment of GC.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
2. Farran B, Muller S, Montenegro RC. Gastric cancer management: kinases as a target therapy. Clin Exp Pharmacol Physiol. 2017;44(6):613–622. doi:10.1111/cep.2017.44.issue-6
3. Kim HS, Kim JH, Kim JW, Kim BC. Chemotherapy in elderly patients with gastric cancer. J Cancer. 2016;7(1):88–94. doi:10.7150/jca.13248
4. Kristensen LS, Hansen TB, Veno MT, Kjems J. Circular RNAs in cancer: opportunities and challenges in the field. Oncogene. 2018;37(5):555–565. doi:10.1038/onc.2017.361
5. Zhu X, Yang G, Xu J, Zhang C. Silencing of SNHG6 induced cell autophagy by targeting miR-26a-5p/ULK1 signaling pathway in human osteosarcoma. Cancer Cell Int. 2019;19:82. doi:10.1186/s12935-019-0794-1
6. Yao T, Chen Q, Fu L, Guo J. Circular RNAs: biogenesis, properties, roles, and their relationships with liver diseases. Hepatol Res. 2017;47(6):497–504. doi:10.1111/hepr.v47.6
7. Han D, Li J, Wang H, et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology. 2017;66(4):1151–1164. doi:10.1002/hep.29270
8. Du WW, Yang W, Liu E, Yang Z, Dhaliwal P, Yang BB. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016;44(6):2846–2858. doi:10.1093/nar/gkw027
9. Zhang J, Liu H, Hou L, et al. Circular RNA_LARP4 inhibits cell proliferation and invasion of gastric cancer by sponging miR-424-5p and regulating LATS1 expression. Mol Cancer. 2017;16(1):151. doi:10.1186/s12943-017-0719-3
10. Chen S, Li T, Zhao Q, Xiao B, Guo J. Using circular RNA hsa_circ_0000190 as a new biomarker in the diagnosis of gastric cancer. Clin Chim Acta. 2017;466:167–171. doi:10.1016/j.cca.2017.01.025
11. Chen J, Li Y, Zheng Q, et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. 2017;388:208–219. doi:10.1016/j.canlet.2016.12.006
12. Shao Y, Li J, Lu R, et al. Global circular RNA expression profile of human gastric cancer and its clinical significance. Cancer Med. 2017;6(6):1173–1180. doi:10.1002/cam4.2017.6.issue-6
13. Hu L, Wu H, Wan X, et al. MicroRNA-585 suppresses tumor proliferation and migration in gastric cancer by directly targeting MAPK1. Biochem Biophys Res Commun. 2018;499(1):52–58. doi:10.1016/j.bbrc.2018.03.116
14. Zou H, Xu X, Luo L, et al. Hsa_circ_0101432 promotes the development of hepatocellular carcinoma (HCC) by adsorbing miR-1258 and miR-622. Cell Cycle. 2019;18(19):2398–2413. doi:10.1080/15384101.2019.1618120
15. Correa P. Gastric cancer: overview. Gastroenterol Clin North Am. 2013;42(2):211–217. doi:10.1016/j.gtc.2013.01.002
16. Amieva M, Peek RM
17. Zhang HD, Jiang LH, Sun DW, Hou JC, Ji ZL. CircRNA: a novel type of biomarker for cancer. Breast Cancer. 2018;25(1):1–7. doi:10.1007/s12282-017-0793-9
18. Bach DH, Lee SK, Sood AK. Circular RNAs in cancer. Mol Ther Nucleic Acids. 2019;16:118–129. doi:10.1016/j.omtn.2019.02.005
19. Yi Y, Liu Y, Wu W, Wu K, Zhang W. Reconstruction and analysis of circRNAmiRNAmRNA network in the pathology of cervical cancer. Oncol Rep. 2019. doi:10.3892/or.2019.7028
20. Rong D, Sun H, Li Z, et al. An emerging function of circRNA-miRNAs-mRNA axis in human diseases. Oncotarget. 2017;8(42):73271–73281. doi:10.18632/oncotarget.v8i42
21. Georgantas RW
22. Ruggiero T, Trabucchi M, De Santa F, et al. LPS induces KH-type splicing regulatory protein-dependent processing of microRNA-155 precursors in macrophages. FASEB J. 2009;23(9):2898–2908. doi:10.1096/fj.09-131342
23. Rodriguez A, Vigorito E, Clare S, et al. Requirement of bic/microRNA-155 for normal immune function. Science. 2007;316(5824):608–611. doi:10.1126/science.1139253
24. Seok HY, Tatsuguchi M, Callis TE, He A, Pu WT, Wang DZ. miR-155 inhibits expression of the MEF2A protein to repress skeletal muscle differentiation. J Biol Chem. 2011;286(41):35339–35346. doi:10.1074/jbc.M111.273276
25. Kluiver J, Poppema S, de Jong D, et al. BIC and miR-155 are highly expressed in Hodgkin, primary mediastinal and diffuse large B cell lymphomas. J Pathol. 2005;207(2):243–249. doi:10.1002/path.1825
26. Lee EJ, Gusev Y, Jiang J, et al. Expression profiling identifies microRNA signature in pancreatic cancer. Int J Cancer. 2007;120(5):1046–1054. doi:10.1002/ijc.22394
27. Iorio MV, Ferracin M, Liu CG, et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005;65(16):7065–7070. doi:10.1158/0008-5472.CAN-05-1783
28. Van Roosbroeck K, Fanini F, Setoyama T, et al. Combining anti-miR-155 with chemotherapy for the treatment of lung cancers. Clin Cancer Res. 2017;23(11):2891–2904. doi:10.1158/1078-0432.CCR-16-1025
29. Zhang L, Chen J, Ning D, et al. FBXO22 promotes the development of hepatocellular carcinoma by regulating the ubiquitination and degradation of p21. J Exp Clin Cancer Res. 2019;38(1):101. doi:10.1186/s13046-019-1058-6
30. Tian X, Dai S, Sun J, et al. F-box protein FBXO22 mediates polyubiquitination and degradation of KLF4 to promote hepatocellular carcinoma progression. Oncotarget. 2015;6(26):22767–22775. doi:10.18632/oncotarget.v6i26
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.