")
Back to Journals » OncoTargets and Therapy » Volume 13
Circ-PRKDC Contributes to 5-Fluorouracil Resistance of Colorectal Cancer Cells by Regulating miR-375/FOXM1 Axis and Wnt/β-Catenin Pathway
Authors Chen H, Pei L, Xie P, Guo G
Received 11 March 2020
Accepted for publication 31 May 2020
Published 23 June 2020 Volume 2020:13 Pages 5939—5953
DOI https://doi.org/10.2147/OTT.S253468
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Geoffrey Pietersz
Hao Chen,1 Lingyu Pei,2 Peng Xie,1 Guancheng Guo1
1Department of Emergency Surgery, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, People’s Republic of China; 2Department of Digestive System, Yongcheng People’s Hospital, Shangqiu, Henan, People’s Republic of China
Correspondence: Guancheng Guo
Department of Emergency Surgery, The First Affiliated Hospital of Zhengzhou University, No. 1, East Jianshe Road, Zhengzhou 450052, Henan, People’s Republic of China
Tel +86 371 66271227
Email [email protected]
Purpose: Diverse circular RNAs (circRNAs) participate in the regulation of drug resistance in human cancers. However, the role of circRNAs in drug resistance in colorectal cancer (CRC) is dismal. In this study, we aimed to explore the effect of circ-PRKDC on 5-fluorouracil (5-FU) resistance in CRC.
Materials and Methods: The levels of circ-PRKDC, microRNA-375 (miR-375) and forkhead box protein M1 (FOXM1) mRNA were determined by quantitative real-time polymerase chain reaction (qRT-PCR). IC50 of 5-FU, cell colony formation ability and invasion were assessed by Cell Counting Kit-8 (CCK-8) assay, colony formation assay and transwell assay, respectively. The protein levels of P-glycoprotein (P-gp), multidrug resistance protein 1 (MRP1), FOXM1, β-catenin and c-Myc were measured via Western blot assay. The targeting relationship between miR-375 and circ-PRKDC or FOXM1 was investigated by dual-luciferase reporter assay and RNA immunoprecipitation (RIP) assay. The effect of circ-PRKDC in vivo was explored by murine xenograft model assay.
Results: Circ-PRKDC was upregulated in 5-FU-resistant CRC tissues and cells. Circ-PRKDC silencing repressed 5-FU resistance, cell colony formation and invasion in 5-FU-resistant CRC cells in vitro and inhibited 5-FU resistance in vivo. MiR-375 was a target of circ-PRKDC and miR-375 inhibition reversed the effects of circ-PRKDC silencing on 5-FU resistance, cell colony formation and invasion. FOXM1 was a direct target gene of miR-375. MiR-375 suppressed 5-FU resistance by targeting FOXM1. Moreover, circ-PRKDC knockdown decreased FOXM1 expression by targeting miR-375. Additionally, circ-PRKDC knockdown impeded wnt/β-catenin pathway by regulating miR-375 and FOXM1.
Conclusion: Circ-PRKDC enhanced 5-FU resistance in CRC by regulating FOXM1/miR-375 axis and wnt/β-catenin pathway.
Keywords: CRC, 5-FU, circ-PRKDC, miR-375, FOXM1, wnt/β-catenin
Introduction
Colorectal cancer (CRC) is a malignant tumor with high morbidity and mortality, seriously threatening human health.1,2 Conventional treatment strategy for CRC patients is surgery combined with chemotherapy.3 5-fluorouracil (5-FU) is the basis of standard chemotherapy for CRC.4 However, chemoresistance seriously affects the effectiveness of CRC treatment.5 Thus, it is of great significance to clarify the mechanisms of drug resistance and find novel targets to counter chemoresistance in CRC.
Circular RNAs (circRNAs) are defined as non-coding RNAs (ncRNAs) with covalently closed-loop structures but without 5ʹ cap and 3ʹ poly (A) tail.6 Increasing evidence has shown that the dysregulation of circRNAs is closely related to tumor progression and chemoresistance in many types of cancers. For example, Huang et al demonstrated that the elevated expression of circAKT3 improved cisplatin resistance in gastric cancer (GC).7 Zhou et al claimed that deficiency of circ_0004015 repressed cell viability and metastasis and gefitinib resistance in non-small cell lung cancer (NSCLC).8 Multiple circRNAs are differentially expressed in drug-resistant CRC cells,9 but the exact roles of circRNAs in drug resistance in CRC are not clear. In this research, we explored the role of circ-PRKDC (also termed as circ_0136666) in 5-FU resistance of CRC.
MicroRNAs (miRNAs) are a series of ncRNAs (~22 nucleotides) which exert their functions mainly by recognizing the 3ʹ untranslated region (3ʹ UTR) of target genes.10 To date, diverse miRNAs have been reported to be involved in chemoresistance of CRC. For example, Sun et al declared that miR-302a could hinder cell metastasis and improve cetuximab sensitivity in CRC by targeting NFIB and CD44.11 Liang et al disclosed that the downregulation of miR-483-3p improved oxaliplatin resistance via suppressing cell apoptosis and promoting migration in CRC.12 MiR-375 has been confirmed to participate in the process of drug sensitivity in cancers, such as prostate cancer,13 breast cancer (BC),14 and cervical cancer.15 Moreover, miR-375 was reduced in CRC and played vital roles in CRC development.16 Nonetheless, the studies of miR-375 in drug resistance of CRC are very insufficient.
Transcription factor Forkhead box protein M1 (FOXM1) belongs to Forkhead Box family and plays crucial roles in various cancers.17,18 More importantly, FOXM1 has been proved to take part in drug resistance in some cancers, such as prostate cancer,19 pancreatic cancer,20 nasopharyngeal carcinoma21 and CRC.22 However, whether miR-375 can target FOXM1 to alter drug resistance in CRC has not been determined.
In this paper, we explored the expression of circ-PRKDC in 5-FU-resistant CRC and further investigated the biological roles and potential mechanisms of circ-PRKDC in 5-FU resistance in CRC.
Materials and Methods
Tissues Acquisition
Thirty pairs of tissue samples were collected from 5-FU-resistant and sensitive CRC patients who underwent surgery at the First Affiliated Hospital of Zhengzhou University. Patients with progressive disease during primary chemotherapy or patients with recurrent disease within 6 months after completing primary chemotherapy were termed 5-FU resistance. Patients with recurrence more than 6 months or without recurrence were called 5-FU sensitive. The samples were collected after chemotherapy and maintained at −80°C before use. Approval was obtained from the Ethics Committee of the First Affiliated Hospital of Zhengzhou University. Written informed consents were signed by the patients. The clinicopathological characteristics of CRC patients were shown in Table 1.
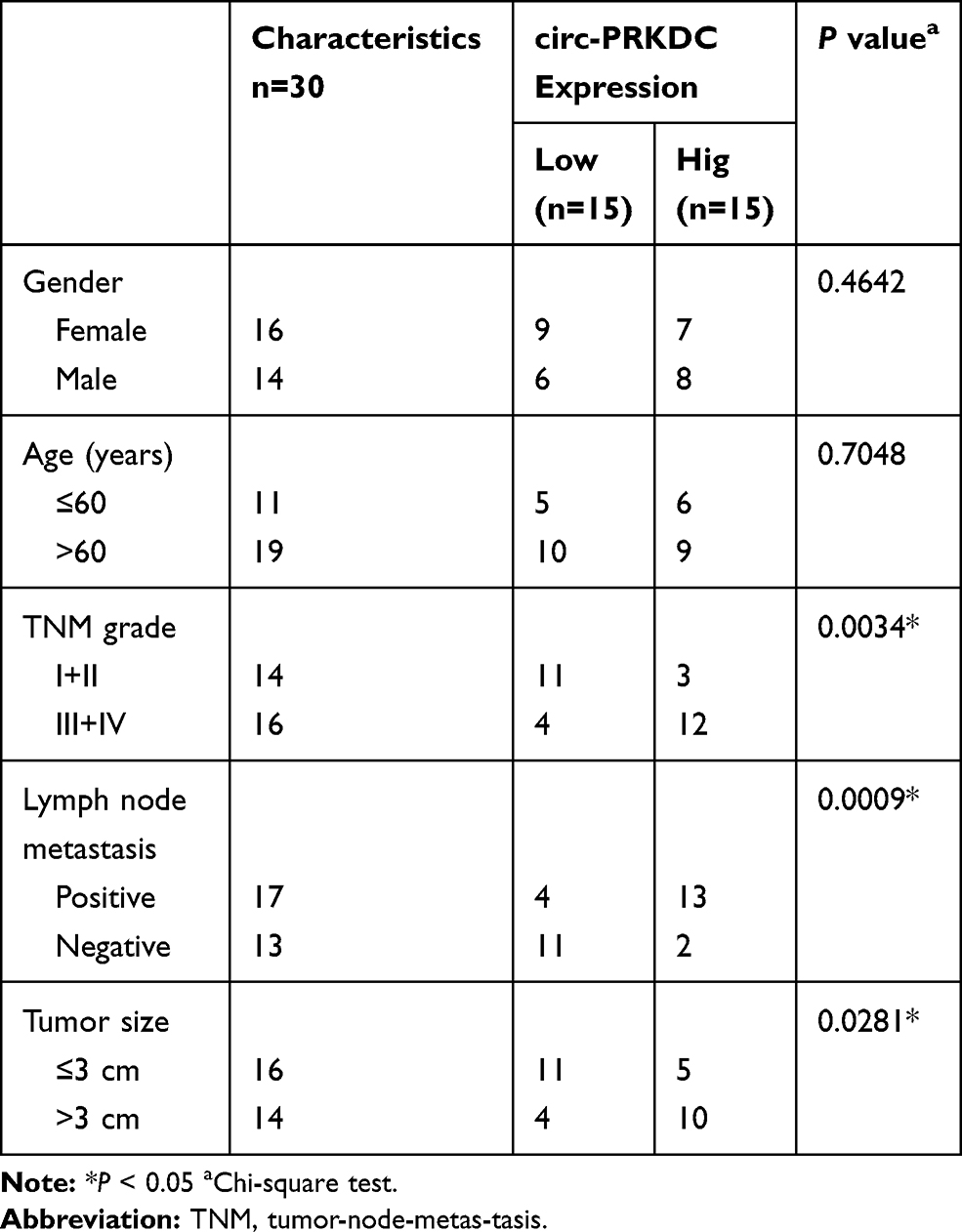 |
Table 1 Relationship Between Circ-PRKDC Expression and Clinicopathologic Features of Colorectal Cancer Patients |
Cell Culture
Normal human fetal colonic mucosa cells (FHC) and CRC cells (SW620 and SW480) were bought from the American Type Culture Collection (ATCC, Manassas, VA, USA). 5-FU-resistant CRC cells (SW620/5-FU and SW480/5-FU) were generated by treating SW620 and SW480 cells with increasing doses of 5-FU (Solarbio, Beijing, China), as previously described.23 All these cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; HyClone, South Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; HyClone) and 1% penicillin-streptomycin (HyClone) at an atmosphere of 5% CO2 and 37°C. To maintain the resistance of SW620/5-FU and SW480/5-FU cells, 15.0 μg/mL 5-FU (Solarbio) was additionally added into the medium.
RNase R Digestion Assay
To assess the stability of circ-PRKDC, total RNA (20 μg) in SW620/5-FU and SW480/5-FU cells was maintained with or without 40 U RNase R (Epicentre, Madison, Wisconsin, USA) for 30 min at 37° followed by qRT-PCR analysis for circ-PRKDC and linear PRKDC expression.
Cell Transfection
Small interfering RNA targeting circ-PRKDC (si-circ-PRKDC) and its control (si-NC), circ-PRKDC overexpression plasmid (circ-PRKDC), FOXM1 overexpression plasmid (FOXM1) and their control (pcDNA), mimics of miR-375 (miR-375) and its control (miR-NC), inhibitors of miR-375 (anti-miR-375) and its control (anti-miR-NC), short hairpin RNA targeting circ-PRKDC (sh-circ-PRKDC) and its control (sh-NC) were synthesized by GeneCopoeia (Guangzhou, China). Cell transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA).
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Assay
Total RNA was extracted from tissues and cells with RNAiso Plus (Takara, Dalian, China) and examined on a NanoDrop 2000c spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Then reverse transcription experiment was conducted using Primer Script™ RT reagent kit (Takara) or miRNA first Strand cDNA Synthesis Kit (Vazyme, Nanjing, China). Next, AceQ Universal SYBR qPCR Master Mix (Vazyme) was adopted to examine the expression of circ-PRKDC, miR-375 and FOXM1. The expression was calculated using the 2−ΔΔCt method. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) or U6 was used as the internal control. The primers were: (circ-PRKDC: 5ʹ-CAGAGACGATTGGCTGGTGAG-3ʹ and R: 5ʹ-TGATAAATTGCCCAACAAAGAGACT-3ʹ); miR-375: (F: 5ʹ-TTTGTTCGTTCGGCTCGC-3ʹ and R: 5ʹ-GCTGTCAACGATACGCTACGT-3ʹ); FOXM1: (F: 5ʹ-CGTCGGCCACTGATTCTCAAA-3ʹ and R: 5ʹ-GGCAGGGGATCTCTTAGGTTC-3ʹ); GAPDH: (F: 5ʹ-AATGGATTTGGACGCATTGGT-3ʹ and R: 5ʹ-TTTGCACTGGTACGTGTTGAT-3ʹ); U6: (F: 5ʹ-CTCGCTTCGGCAGCACATATACTA-3ʹ and R: 5ʹ-ACGAATTTGCGTGTCATCCTTGCG-3ʹ). The nanogram amount of circ-PRKDC in resistant CRC tissues was 7.68 ng, and the nanogram amount of circ-PRKDC in sensitive CRC tissues was 0.56 ng. The nanogram amount of circ-PRKDC in FHC, SW620, SW480, SW620/5-FU and SW480/5-FU cells were 0.42 ng, 1.85 ng, 2.48 ng, 4.25 ng and 6.26 ng, respectively.
Cell Counting Kit-8 (CCK-8) Assay
IC50 of 5-FU was evaluated by CCK-8 assay kit (Beyotime, Shanghai, China). In brief, SW620, SW480, SW620/5-FU or SW480/5-FU cells were plated into 96-well plates (5×103 cells/well) and maintained overnight. Then various doses of 5-FU (Solarbio) were added into the well and incubated for further 48 h. Next, 10 µL CCK-8 (Beyotime) was added into each well and kept for 4 h. The absorbance at 450 nm was measured with a microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Colony Formation Assay
CRC cells were put into 6-well plates (600 cells/well) and kept in DMEM (HyClone) at 37°C. The medium was replaced every 3 days. Two weeks later, the cells were washed with phosphate-buffered saline (PBS; Solarbio) and fixed with 4% paraformaldehyde (Sangon, Shanghai, China). Next, cell colonies were stained with crystal violet (Sangon) and observed under a microscope (Olympus, Tokyo, Japan).
Transwell Assay
The invasion of CRC cells was estimated through transwell insert chambers (Corning Incorporated, Corning, NY, USA) pre-coated with Matrigel (Solarbio). In brief, CRC cells (5×104 cells) were resuspended in serum-free DMEM (HyClone) and then put into the top chamber. The bottom chamber was added with DMEM (HyClone) including 10% FBS (HyClone). After 48 h, cells those still on the top membrane were removed and invaded cells were fixed with methanol, stained with crystal violet (Sangon) and counted under a microscope (Olympus).
Western Blot Assay
Total protein was isolated using RIPA buffer (Beyotime) and determined with a BCA Protein Quantification Kit (Vazyme). The proteins were separated by 10% sodium dodecyl sulfonate-polyacrylamide gel (SDS-PAGE; Solarbio) and transferred onto polyvinylidene difluoride membranes (Millipore, Billerica, MA, USA). Then the membranes were blocked in skim milk for 1 h and probed with primary antibodies: GAPDH (ab181602; Abcam, Cambridge, MA, USA), P-glycoprotein (P-gp; ab170904; Abcam), multidrug resistance protein 1 (MRP1; ab32574; Abcam), FOXM1 (ab180710; Abcam), β-catenin (ab16051; Abcam) or c-Myc (ab39688; Abcam) overnight at 4°C followed by incubation with relevant secondary antibody (ab6728; Abcam) at room temperature for 2 h. The proteins were observed by an enhanced chemiluminescence kit (Vazyme).
Dual-Luciferase Reporter Assay
The fragments of circ-PRKDC or 3ʹUTR of FOXM1 containing the predicted wild-type or mutant complementary sequences of miR-375 were cloned into the pmirGLO vector (Promega, Madison, WI, USA) to establish luciferase reporter plasmids: WT-circ-PRKDC, MUT-circ-PRKDC, FOXM1 3ʹ UTR-WT and FOXM1 3ʹUTR-MUT. Next, SW620/5-FU and SW480/5-FU cells were seeded into 24-well-plates. MiR-375 or miR-NC together with indicated vector were transfected into cells. The luciferase activity was estimated via a Dual-Luciferase Reporter Assay Kit (Promega) after 48 h.
RNA Immunoprecipitation (RIP) Assay
RIP assay was conducted using the Magna RNA-binding protein immunoprecipitation kit (Millipore). In brief, SW620/5-FU and SW480/5-FU cells were disrupted in RIP buffer and then cell extracts were interacted with immunoglobulin G (Anti-IgG; Abcam) or Argonaute-2 (Ago2; Abcam) which was coated with magnetic beads for 4 h at 4°C. Finally, the levels of circ-PRKDC, miR-375 and FOXM1 on the beads were quantified using qRT-PCR analysis.
Murine Xenograft Model
Nude mice (5-week-old) were bought from Shanghai SLAC Laboratory Animal Co., Ltd (Shanghai, China) and divided into 4 groups (n=8). Lentivirus-mediated sh-circ-PRKDC or sh-NC was stably transfected into SW480/5-FU cells and then 3×106 transfected cells were subcutaneously injected into the mice. Eight days after inoculation, the mice were administrated with 20.0 μg/mL 5-FU (Solarbio) or the same volume of PBS (Solarbio) by intraperitoneal injection every 3 days. Tumor length (L) and width (W) were monitored every 3 days after 8 days. Tumor volume was calculated by (L×W2)/2. After 29 days, the mice were euthanized and tumor samples were collected for weight, qRT-PCR assay and Western blot assay. The study was performed in accordance with the Guidelines for Care and Use of Laboratory Animals of “National Institutes of Health” and approved by the Ethics Committee of Animal Research of the First Affiliated Hospital of Zhengzhou University.
Statistical Analysis
The experiments were repeated three times. Statistical analysis was conducted using software GraphPad Prism 7 (GraphPad Inc., La Jolla, CA, USA) and data were exhibited as mean ± standard deviation (SD). The correlation between the expression of miR-375 and circ-PRKDC or FOXM1 was estimated by Spearman correlation coefficient analysis. Student’s t-test or one-way analysis of variance (ANOVA) was utilized for difference analysis. P<0.05 was defined as significant.
Results
Circ-PRKDC Expression Was Elevated in 5-FU-Resistant CRC Tissues and Cell Lines
To begin with, the expression of circ-PRKDC in 5-FU-resistant CRC tissues (n=30) and 5-FU-sensitive CRC tissues was measured. The data of qRT-PCR showed that compared to 5-FU-sensitive tissues, circ-PRKDC level was notably increased in 5-FU-resistant CRC tissues (Figure 1A). Moreover, we found that circ-PRKDC level was related to TNM grades, lymph node metastasis and tumor size and was irrelevant to the gender and age of patients (Table 1). Next, we determined circ-PRKDC expression in normal human fetal colonic mucosa cells (FHC), CRC cells (SW620 and SW480) and 5-FU-resistant CRC cells (SW620/5-FU and SW480/5-FU) via qRT-PCR. Our results displayed that circ-PRKDC expression was markedly raised in CRC cells compared to FHC cells; moreover, circ-PRKDC was higher expressed in 5-FU-resistant CRC cells in reference to parental sensitive CRC cells (Figure 1B). Then the stability of circ-PRKDC was analyzed by RNase R digestion assay, showing that circ-PRKDC was resistant to RNase R, while linear PRKDC was digested by RNase R (Figure 1C and D). Thus, these data demonstrated that the dysregulation of circ-PRKDC might participate in 5-FU resistance in CRC.
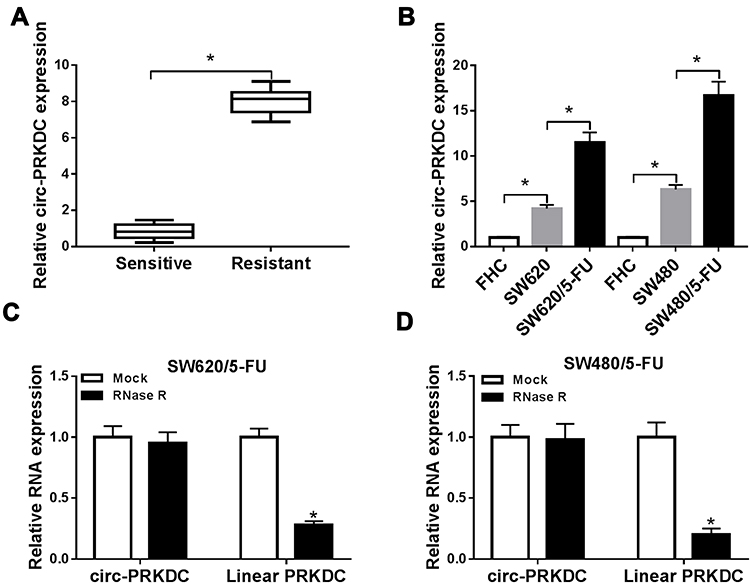 |
Figure 1 High expression of circ-PRKDC was observed in 5-FU-resistant CRC tissues and cells. (A) Circ-PRKDC expression in 5-FU-resistant and 5-FU-sensitive CRC tissues was determined by qRT-PCR. (B) Circ-PRKDC expression in FHC, SW620, SW620/5-FU, SW480 and SW480/5-FU cells was measured by qRT-PCR. (C and D) The expression of circ-PRKDC and linear PRKDC in SW480 and SW480/5-FU cells treated with or without RNase R were measured by qRT-PCR assay. *P<0.05. |
Circ-PRKDC Knockdown Suppressed 5-FU Resistance in 5-FU-Resistant CRC Cells
To explore the exact role of circ-PRKDC in the resistance of 5-FU-resistant CRC cells to 5-FU, we firstly determined 5-FU resistance in 5-FU-resistant CRC cells by treating cells with different doses of 5-FU. As shown in Figure 2A, IC50 of 5-FU was increased in SW620/5-FU and SW480/5-FU cells in comparison with SW620 and SW480 cells, indicating 5-FU resistance was produced in SW620/5-FU and SW480/5-FU cells. Next, loss-of-function experiments were conducted through transfecting si-circ-PRKDC into SW620/5-FU and SW480/5-FU cells. The data of qRT-PCR exhibited that si-circ-PRKDC transfection led to a marked reduction of circ-PRKDC expression in SW620/5-FU and SW480/5-FU cells compared to si-NC groups (Figure 2B). CCK-8 assay displayed that circ-PRKDC knockdown remarkedly enhanced 5-FU sensitivity in SW620/5-FU and SW480/5-FU cells, as indicated by decreased IC50 value (Figure 2C). Colony formation assay exhibited that the colony numbers of SW620/5-FU and SW480/5-FU cells were evidently decreased after circ-PRKDC silencing compared to control groups (Figure 2D). Transwell assay indicated that circ-PRKDC deficiency remarkably impeded SW620/5-FU and SW480/5-FU cell invasion when compared to si-NC group (Figure 2E). Moreover, the effects of circ-PRKDC silencing on drug resistance-associated proteins (P-gp and MRP1) were investigated via Western blot assay. The data showed that P-gp and MRP1 levels were all decreased in SW620/5-FU and SW480/5-FU cells transfected with si-circ-PRKDC compared to those in cells transfected with si-NC (Figure 2F and G). Collectively, silencing of circ-PRKDC inhibited the resistance of 5-FU-resistant CRC cells to 5-FU.
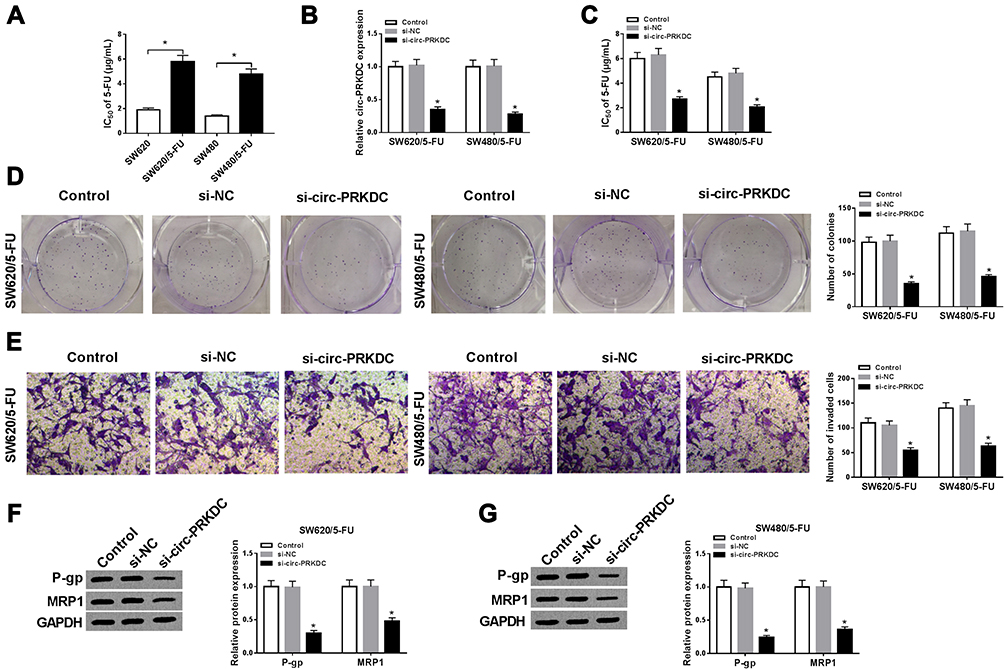 |
Figure 2 Downregulation of circ-PRKDC improved 5-FU sensitivity in 5-FU-resistant CRC cells. (A) IC50 of 5-FU in SW620, SW620/5-FU, SW480 and SW480/5-FU cells was examined by CCK-8 assay. (B–G) SW620/5-FU and SW480/5-FU cells were transfected with si-PRKDC or si-NC. (B) The expression of circ-PRKDC in SW620/5-FU and SW480/5-FU cells was determined by qRT-PCR. (C) IC50 of 5-FU in SW620/5-FU and SW480/5-FU cells was evaluated by CCK-8 assay. (D) The colony formation ability of SW620/5-FU and SW480/5-FU cells was analyzed by colony formation assay. (E) The invasion of SW620/5-FU and SW480/5-FU cells was assessed by transwell assay. (F and G) The protein levels of P-gp and MRP1 in SW620/5-FU and SW480/5-FU cells were measured using Western blot assay. *P<0.05. |
Circ-PRKDC Silencing Repressed 5-FU Resistance in 5-FU-Resistant CRC Cells by Targeting miR-375
In order to explore the potential mechanism of circ-PRKDC in regulating 5-FU resistance in 5-FU-resistant CRC cells, online website starBase v2.0 was utilized to search the potential target of circ-PRKDC. MiR-375 was predicted to be a target of circ-PRKDC and their binding sites were presented in Figure 3A. To verify this prediction, dual-luciferase reporter assay was carried out. The data indicated that miR-375 together with WT-circ-PRKDC transfection led to an obvious suppression in the luciferase activity in SW620/5-FU and SW480/5-FU cells compared to miR-NC and WT-circ-PRKDC co-transfected groups; however, no change was observed in MUT-circ-PRKDC groups (Figure 3B and C). Then we further conducted RIP assay to demonstrate the interaction between circ-PRKDC and miR-375. The results showed that the expression levels of circ-PRKDC and miR-375 were markedly elevated in Ago2 immunoprecipitates in SW620/5-FU and SW480/5-FU cells compared to IgG control groups (Figure 3D and E). Afterward, the expression of miR-375 in 5-FU-resistant CRC tissues was measured. The data showed that miR-375 was weakly expressed in 5-FU-resistant CRC tissues relative to 5-FU-sensitive CRC tissues (Figure 3F). Likewise, miR-375 level was decreased in CRC cells compared to FHC cells; moreover, there was a lower expression of miR-375 in 5-FU-resistant CRC cells compared to CRC cells (Figure 3G). As analyzed by Spearman correlation coefficient analysis, miR-375 level was inversely correlated with circ-PRKDC level in CRC tissues (Figure 3H). Besides, we observed that si-circ-PRKDC transfection distinctly decreased circ-PRKDC level and increased miR-375 level in SW620/5-FU and SW480/5-FU cells, whereas circ-PRKDC transfection showed the opposite results (Figure 3I and J). Subsequently, the association between circ-PRKDC and miR-375 in regulating 5-FU resistance was explored. SW620/5-FU and SW480/5-FU cells were transfected with si-NC, si-circ-PRKDC, si-circ-PRKDC+anti-miR-NC or si-circ-PRKDC+anti-miR-375. QRT-PCR assay indicated that the upregulation of miR-375 in SW620/5-FU and SW480/5-FU cells caused by circ-PRKDC knockdown was effectively reversed by miR-375 inhibition (Figure 3K). CCK-8 assay showed that IC50 of 5-FU in SW620/5-FU and SW480/5-FU cells were reduced by circ-PRKDC deficiency, while miR-375 deletion ameliorated the reduction (Figure 3L). The suppressive roles of circ-PRKDC knockdown in cell colony formation and cell invasion in SW620/5-FU and SW480/5-FU cells were all abrogated following the inhibition of miR-375, as illustrated by colony formation assay and transwell assay (Figure 3M and N). Moreover, miR-375 inhibition reversed the downregulation of P-gp and MRP1 in SW620/5-FU and SW480/5-FU cells mediated by circ-PRKDC knockdown (Figure 3O and P). To sum up, miR-375 inhibition overturned the inhibitory effect of circ-PRKDC silencing on 5-FU resistance in 5-FU-resistant CRC cells.
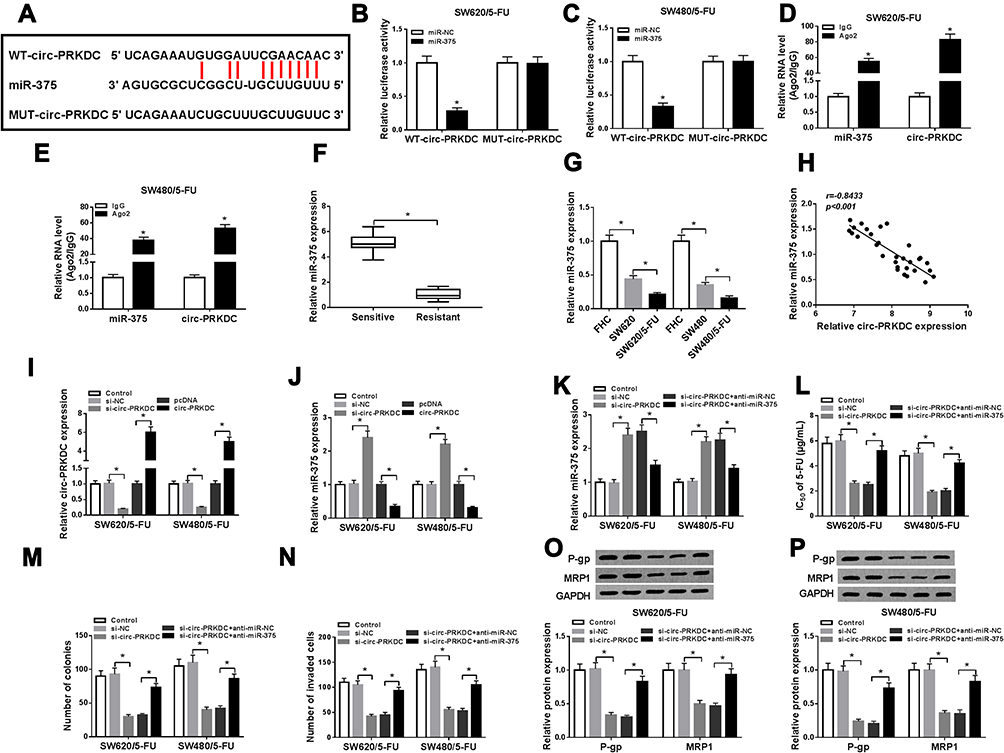 |
Figure 3 Circ-PRKDC targeted miR-375 to regulate 5-FU resistance in 5-FU-resistant CRC cells. (A) The binding sites between circ-PRKDC and miR-375 were predicted by starBase v2.0. (B and C) The combination between circ-PRKDC and miR-375 was investigated by dual-luciferase reporter assay. (D and E) After RIP assay, the expression levels of circ-PRKDC and miR-375 in IgG or Ago2 immunoprecipitates were measured by qRT-PCR analysis. (F) MiR-375 expression in 5-FU-resistant and 5-FU-sensitive CRC tissues was measured using qRT-PCR assay. (G) MiR-375 expression in FHC, SW620, SW480, SW620/5-FU and SW480/5-FU cells was determined by qRT-PCR. (H) The correlation between circ-PRKDC and miR-375 was analyzed by Spearman correlation coefficient analysis. (I and J) The expression levels of circ-PRKDC and miR-375 in SW620/5-FU and SW480/5-FU cells transfected with si-NC, si-circ-PRKDC, pcDNA or circ-PRKDC were examined by qRT-PCR. (K–P) SW620/5-FU and SW480/5-FU cells were assigned to control, si-NC, si-circ-PRKDC, si-circ-PRKDC+anti-miR-NC and si-circ-PRKDC+anti-miR-375 groups. (K) The expression of miR-375 in SW620/5-FU and SW480/5-FU cells was examined through qRT-PCR. (L) IC50 of 5-FU in SW620/5-FU and SW480/5-FU cells was evaluated by CCK-8 assay. (M) The colony numbers of SW620/5-FU and SW480/5-FU cells were tested by colony formation assay. (N) The invasion of SW620/5-FU and SW480/5-FU cells was detected via transwell assay. (O and P) The protein levels of P-gp and MRP1 in SW620/5-FU and SW480/5-FU cells were measured via Western blot assay. *P<0.05. |
FOXM1 Was a Target Gene of miR-375
Through further searching bioinformatics software starBase v2.0, FOXM1 was predicted to be a target gene of miR-375 and their potential binding sites are presented in Figure 4A. Dual-luciferase reporter assay displayed that FOXM1 3ʹUTR-WT and miR-375 co-transfection caused a remarkable reduction of the luciferase activity in SW620/5-FU and SW480/5-FU cells compared to FOXM1 3ʹUTR-WT and miR-NC co-transfected groups, but the luciferase activity in FOXM1 3ʹUTR-MUT groups was not changed, indicating the combination between FOXM1 and miR-375 (Figure 4B and C). RIP results showed that miR-375 and FOXM1 levels were enhanced in Ago2 immunoprecipitates in SW620/5-FU and SW480/5-FU cells compared to IgG control groups, further confirming the combination between miR-375 and FOXM1 (Figure 4D and E). Thereafter, the mRNA and protein levels of FOXM1 in 5-FU-sensitive and 5-FU-resistant CRC tissues were measured. Our data showed that FOXM1 mRNA and protein levels were all distinctly raised in 5-FU-resistant tissues compared to 5-FU-sensitive tissues (Figure 4F and G). Spearman correlation coefficient analysis showed that there was an inverse correlation between the levels of FOXM1 and miR-375 in CRC tissues (Figure 4H). Likewise, we also observed that the mRNA and protein levels of FOXM1 were increased in SW620 and SW480 cells compared to FHC cells; moreover, FOXM1 mRNA and protein were higher expressed in SW620/5-FU and SW480/5-FU cells than in SW620 and SW480 cells (Figure 4I and J). Collectively, miR-375 negatively regulated FOXM1 expression by directly targeting.
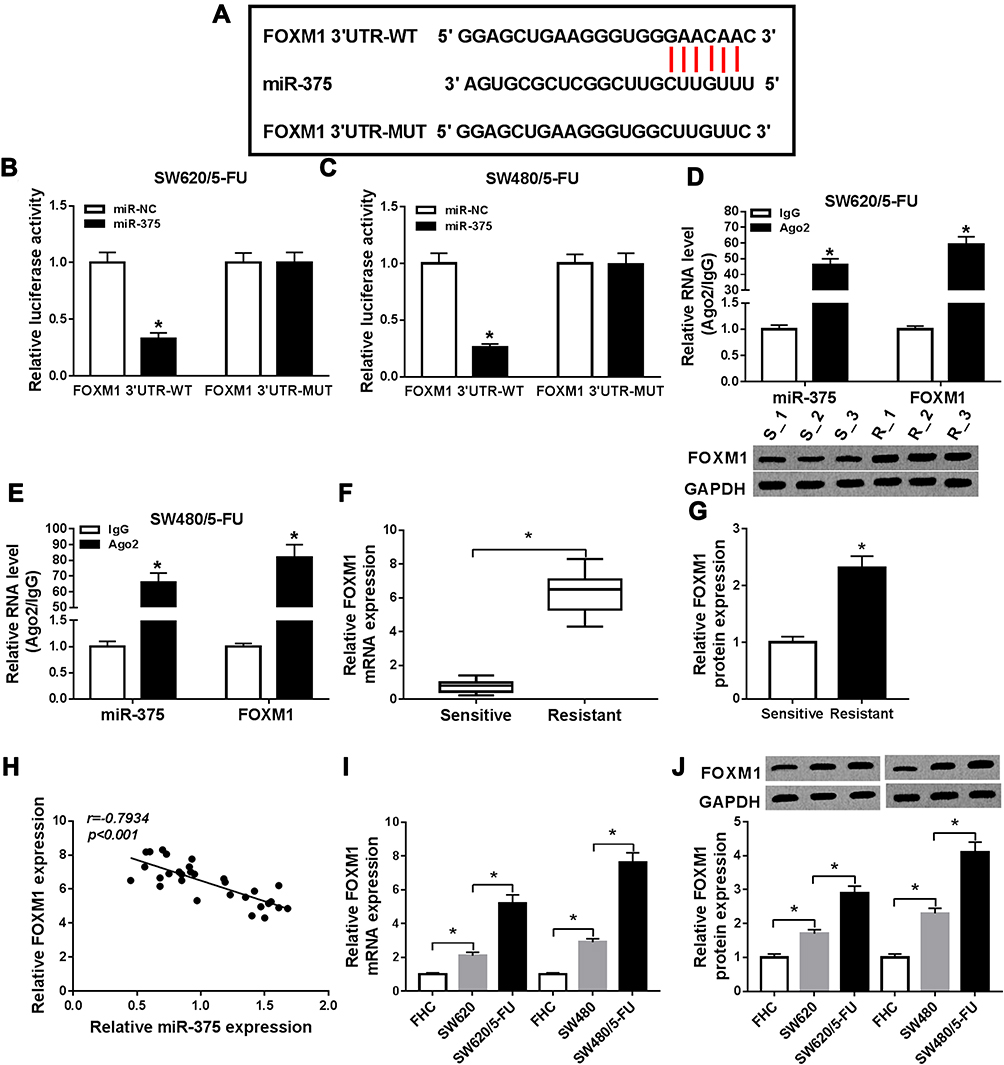 |
Figure 4 FOXM1 was a direct target gene of miR-375. (A) The potential binding sites between miR-375 and FOXM1 were presented. (B and C) The relationship between miR-375 and FOXM1 was verified by dual-luciferase reporter assay. (D and E) The interaction between FOXM1 and miR-375 were analyzed by RIP assay and qRT-PCR analysis. (F and G) The mRNA and protein levels of FOXM1 in 5-FU-resistant and 5-FU-sensitive CRC tissues were measured by qRT-PCR assay and Western blot assay, respectively. (H) The correlation between miR-375 and FOXM1 in CRC tissues was analyzed by Spearman correlation coefficient analysis. (I and J) The mRNA and protein levels of FOXM1 in FHC, SW620, SW480, SW620/5-FU and SW480/5-FU cells were measured by qRT-PCR assay and Western blot assay, respectively. *P<0.05. |
FOXM1 Overexpression Reversed the Inhibitory Effect of miR-375 on 5-FU Resistance in 5-FU Resistant CRC Cells
In order to investigate whether miR-375 could regulate 5-FU resistance in 5-FU-resistant CRC cells by targeting FOXM1, SW620/5-FU and SW480/5-FU cells were transfected with miR-NC, miR-375, miR-375+pcDNA or miR-375+FOXM1. As we observed in Figure 5A and B, miR-375 transfection obviously decreased the protein level of FOXM1 in SW620/5-FU and SW480/5-FU cells, while FOXM1 transfection partially ameliorated the decrease. As demonstrated by CCK-8 assay, colony formation assay and transwell assay, IC50 of 5-FU, cell colony formation ability and cell invasion in SW620/5-FU and SW480/5-FU cells were all apparently suppressed by miR-375 overexpression, while the elevation of FOXM1 effectively rescued the impacts (Figure 5C–E). In addition, we examined the protein levels of P-gp and MRP1. The data of Western blot assay displayed that miR-375 reduced the levels of P-gp and MRP1 in SW620/5-FU and SW480/5-FU cells, while these effects were abolished by FOXM1 overexpression (Figure 5F and G). All these data illustrated that miR-375 could repress 5-FU resistance by targeting FOXM1 in 5-FU-resistant CRC cells.
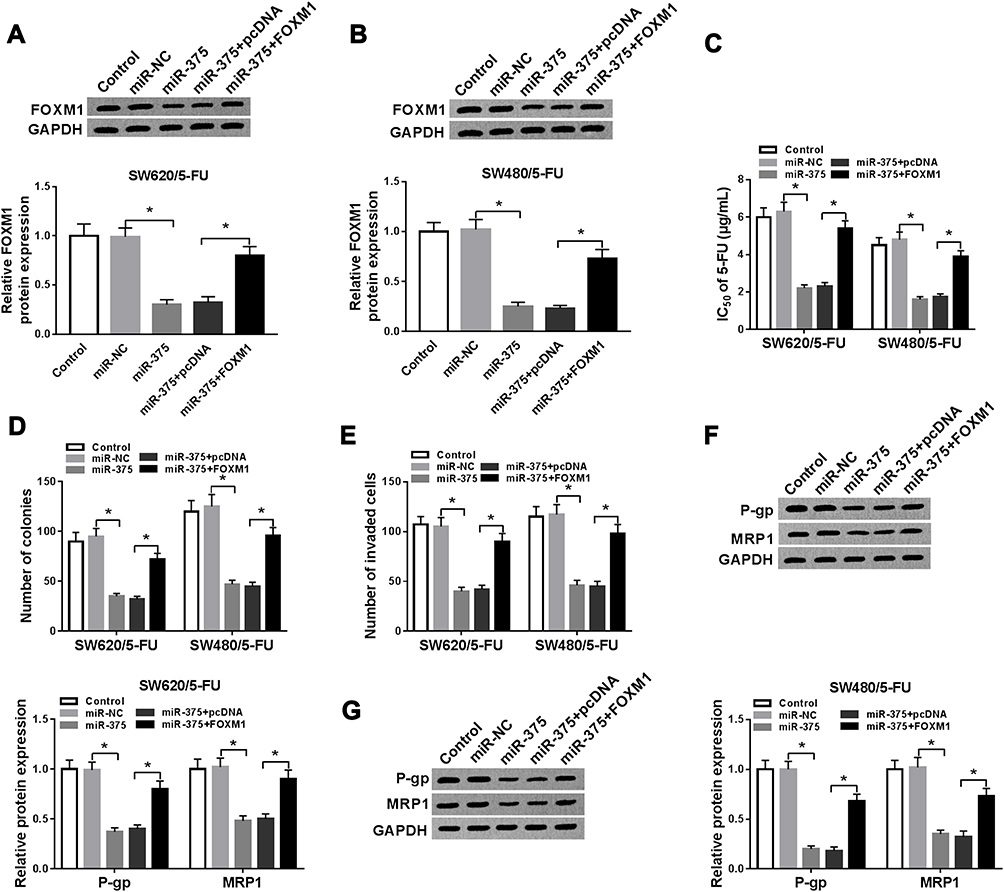 |
Figure 5 MiR-375 inhibited 5-FU resistance by binding to FOXM1 in 5-FU-resistant CRC cells. SW620/5-FU and SW480/5-FU cells were divided into 5 groups: control, miR-NC, miR-375, miR-375+pcDNA and miR-375+FOXM1. (A and B) The protein level of FOXM1 in SW620/5-FU and SW480/5-FU cells was measured by Western blot assay. (C) IC50 of 5-FU in SW620/5-FU and SW480/5-FU cells was evaluated by CCK-8 assay. (D) The colony formation ability of SW620/5-FU and SW480/5-FU cells was analyzed by colony formation assay. (E) SW620/5-FU and SW480/5-FU cell invasion was explored by transwell assay. (F and G) The protein levels of P-gp and MRP1 in SW620/5-FU and SW480/5-FU cells were detected by Western blot assay. *P<0.05. |
Circ-PRKDC Knockdown Decreased FOXM1 Expression via Sponging miR-375
In order to further investigate the relationship among circ-PRKDC, miR-375 and FOXM1, the protein level of FOXM1 in SW620/5-FU and SW480/5-FU cells transfected with si-NC, si-circ-PRKDC, si-circ-PRKDC+anti-miR-NC or si-circ-PRKDC+anti-miR-375 was measured by Western blot assay. The results displayed that circ-PRKDC knockdown drastically downregulated the protein level of FOXM1 in SW620/5-FU and SW480/5-FU cells, while miR-375 inhibition overturned the effect (Figure 6A and B). These data indicated that circ-PRKDC could positively modulate FOXM1 expression by targeting miR-375 in 5-FU-resistant CRC cells.
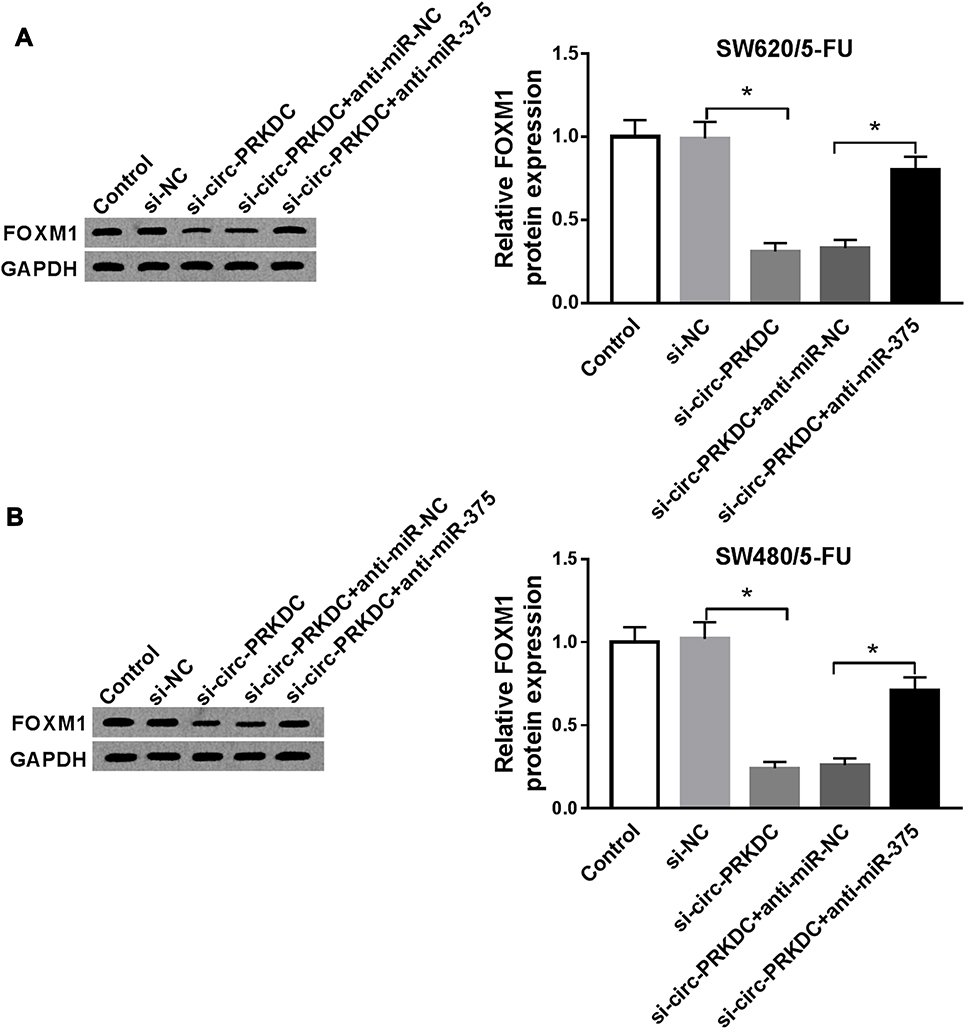 |
Figure 6 Silencing of circ-PRKDC downregulated FOXM1 expression via binding to miR-375 in 5-FU-resistant CRC cells. (A and B) SW620/5-FU and SW480/5-FU cells were transfected with si-NC, si-circ-PRKDC, si-circ-PRKDC+anti-miR-NC or si-circ-PRKDC+anti-miR-375 and then the protein level of FOXM1 was determined by Western blot assay. *P<0.05. |
MiR-375 Inhibition or FOXM1 Overexpression Abrogated the Suppressive Role of Circ-PRKDC Silencing in Wnt/β-Catenin Pathway
To further explore the molecular mechanism of circ-PRKDC in the regulation of 5-FU resistance in 5-FU-resistant CRC cells, the levels of wnt/β-catenin-associated proteins (β-catenin and c-Myc) in SW620/5-FU and SW480/5-FU cells transfected with si-NC, si-circ-PRKDC, si-circ-PRKDC+anti-miR-NC, si-PRKDC+anti-miR-375, si-circ-PRKDC+pcDNA or si-circ-PRKDC+FOXM1 were measured. The data showed that circ-PRKDC deficiency led to noteworthy reduction in β-catenin and c-Myc levels in SW620/5-FU and SW480/5-FU cells, while miR-375 deletion or FOXM1 overexpression rescued the effects (Figure 7A and B). To sum up, circ-PRKDC silencing blocked the activation of wnt/β-catenin pathway by regulating miR-375 and FOXM1 in 5-FU-resistant CRC cells.
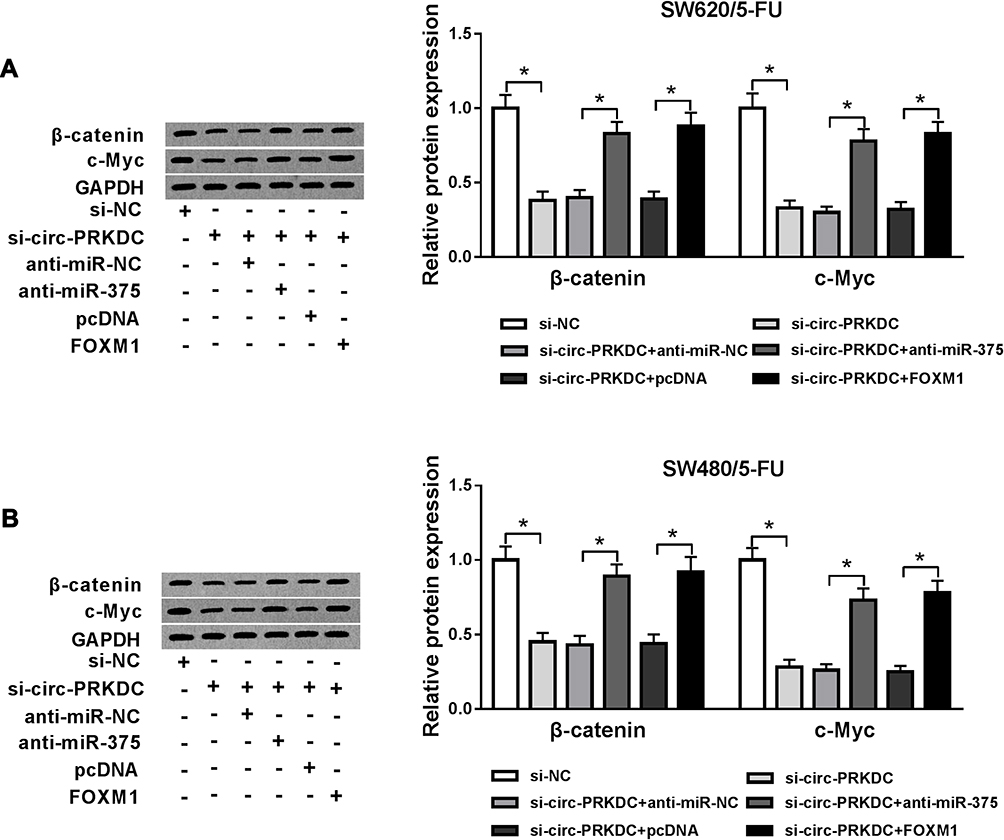 |
Figure 7 Circ-PRKDC knockdown suppressed wnt/β-catenin pathway in 5-FU-resistant CRC cells by regulating miR-375 and FOXM1. (A and B) SW620/5-FU and SW480/5-FU cells were assigned to control, si-NC, si-circ-PRKDC, si-circ-PRKDC+anti-miR-NC, si-PRKDC+anti-miR-375, si-circ-PRKDC+pcDNA and si-circ-PRKDC+FOXM1 groups and then the protein levels of β-catenin and c-Myc were examined by Western blot assay. *P<0.05. |
Circ-PRKDC Deficiency Repressed 5-FU Resistance of CRC in vivo
In order to reveal the role of circ-PRKDC in 5-FU resistance in vivo, sh-NC or sh-circ-PRKDC transfected SW480/5-FU cells were injected into the mice. After 8 days, the mice were treated with 5-FU or PBS every 3 days. We found that tumor volume and weight were all blocked by circ-PRKDC knockdown or 5-FU treatment (Figure 8A and B). Moreover, the expression levels of circ-PRKDC, FOXM1, β-catenin and c-Myc were reduced in sh-circ-PRKDC+PBS and sh-circ-PRKDC+5-FU treated mice compared to corresponding sh-NC+PBS and sh-NC+5-FU groups (Figure 8C and D). Thus, we drew a conclusion that circ-PRKDC knockdown enhanced 5-FU sensitivity of CRC in vivo. Next, we presented the schematic representation of circ-PRKDC on 5-FU resistance, showing that circ-PRKDC overexpression contributed to 5-FU resistance of CRC by repressing miR-375, promoting FOXM1 and activating wnt/β-catenin pathway (Figure 8E).
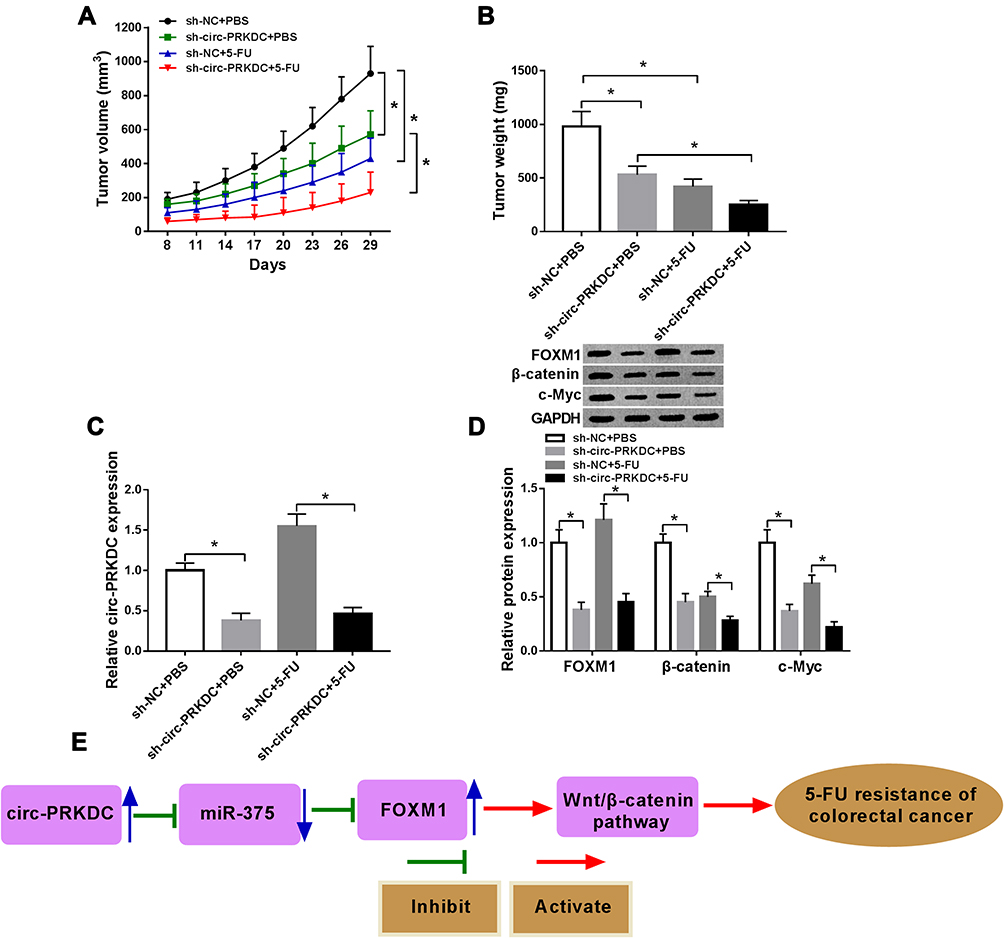 |
Figure 8 Knockdown of circ-PRKDC improved 5-FU sensitivity of CRC in vivo. Sh-NC or sh-circ-PRKDC transfected SW480/5-FU cells were injected into the mice and treated with PBS or 5-FU every 3 days after 8 days. (A) Tumor volume was examined every 3 days after 8 days. (B) Tumor weight was measured after 29 days. (C) The expression of circ-PRKDC in the harvested tissues was determined by qRT-PCR. (D) The protein levels of FOXM1, β-catenin and c-Myc in the harvested tissues were determined by Western blot assay. (E) The schematic representation of circ-PRKDC/miR-375/FOXM1/wnt/β-catenin pathway in the modulation of 5-FU resistance in CRC. *P<0.05. |
Discussion
5-FU is a first-line drug for the therapy of various types of tumors in the clinic.4 However, the acquisition of drug resistance is one of the major obstacles for tumor treatment.24 Accumulating evidence has elucidated that circRNAs act as essential regulators in drug resistance in cancers.25 In this paper, we investigated the role and mechanism of circ-PRKDC in 5-FU resistance in CRC. We observed that circ-PRKDC level was raised in 5-FU-resistant CRC and circ-PRKDC knockdown improved the sensitivity of CRC cells to 5-FU by modulating miR-375/FOXM1 axis and wnt/β-catenin pathway.
Liu et al unraveled that circ-PVT1 was conspicuously elevated in paclitaxel (PTX)-resistant GC and its silencing improved PTX sensitivity via promoting cell apoptosis and repressing invasion in PTX-resistant GC cells.26 Yan et al claimed that circ_0035483 contributed to the resistance of renal cancer cells to gemcitabine.27 These findings suggested the important role of circRNAs in chemoresistance in cancers. However, the effect of circRNAs on drug resistance in CRC is barely studied. Circ_0136666 was increased in CRC and circ_0136666 deficiency hampered CRC cell viability, colony formation, cell cycle process and metastasis,28 indicating the tumorigenic role of circ_0136666 in CRC. Nonetheless, the contribution of circ_0136666 to drug resistance in CRC remains unknown. Herein, we investigated the effect of circ-PRKDC in 5-FU sensitivity in CRC for the first time. We found that circ-PRKDC was drastically elevated in 5-FU-resistant CRC. Moreover, deficiency of circ-PRKDC improved 5-FU sensitivity and repressed cell colony formation and invasion in vitro. Circ-PRKDC knockdown also increased 5-FU sensitivity in vivo. In a word, circ-PRKDC played a positive role in 5-FU resistance of CRC.
Convincing evidence has shown that circRNAs can exert their functions via acting as miRNA sponges.29 For example, circ_0002483 could regulate taxol sensitivity of NSCLC via interacting with miR-182-5p.30 Circ_0003998 facilitated the resistance of lung adenocarcinoma to docetaxel (DTX) by binding to miR-326.31 We identified that miR-375 was a target of circ-PRKDC. Inhibition of miR-375 weakened the suppressive roles of circ-PRKDC interference in drug resistance, colony formation and cell invasion in 5-FU-resistant cells, suggesting that circ-PRKDC facilitated 5-FU resistance in CRC by modulating miR-375. Wang et al reported that miR-375 could promote DTX resistance in prostate cancer by modulating SEC23A and YAP1.13 Ward et al manifested that miR-375 was reduced in tamoxifen-resistant cells, and miR-375 overexpression improved tamoxifen sensitivity in BC by targeting MTDH.32 The different role of miR-375 in drug resistance of cancers might be associated with different tumor microenvironments and drug types. Xu et al suggested that miR-375 was weakly expressed in CRC and its elevation restrained the resistance of CRC cells to 5-FU by interacting with YAP1 and SP1.33 Another study in CRC showed that miR-375 was reduced in CRC cells and tissues, and enhanced 5-FU sensitivity in CRC cells through promoting cell apoptosis and cycle arrest and repressing growth and motility, partially by targeting TYMS.34 In the research, miR-375 was decreased and miR-375 overexpression facilitated 5-FU sensitivity and repressed cell colony formation and invasion in 5-FU-resistant CRC cells. Additionally, FOXM1 was a target gene of miR-375. In CRC, Varghese et al demonstrated that FOXM1 level was elevated and repressed 5-FU sensitivity in 5-FU-resistant CRC.22 Liu et al proved that FOXM1 was increased and downregulation of FOXM1 improved 5-FU sensitivity in drug-resistant CRC cells.35 In the study, a high expression of FOXM1 was observed in 5-FU-resistant CRC. Moreover, FOXM1 overexpression restored the impacts of miR-375 on 5-FU resistance, colony formation and motility in 5-FU-resistant CRC cells, indicating that miR-375 could regulate 5-FU resistance through targeting FOXM1.
The activation of wnt/β-catenin pathway is related to drug resistance in human cancers. For example, circ_001569 facilitated osteosarcoma progression and drug resistance via promoting wnt/β-catenin pathway.36 Circ_0003418 knockdown repressed cisplatin sensitivity through the activation of wnt/β-catenin pathway in hepatocellular carcinoma.37 In this study, circ-PRKDC deficiency decreased β-catenin and c-Myc levels in 5-FU-resistant CRC cells, suggesting the inactivation of wnt/β-catenin pathway, while miR-375 deletion or FOXM1 overexpression abolished the effect.
In conclusion, we verified that circ-PRKDC and FOXM1 were apparently increased and miR-375 was drastically decreased in 5-FU resistant CRC. Further research indicated circ-PRKDC knockdown enhanced the sensitivity of 5-FU-resistant cells to 5-FU by regulating miR-375/FOXM1 axis and wnt/β-catenin pathway, which might provide a promising strategy to counter 5-FU-resistant in CRC.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi:10.3322/caac.21492
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7–30. doi:10.3322/caac.21332
3. Nordlinger B, Sorbye H, Glimelius B, et al. Perioperative FOLFOX4 chemotherapy and surgery versus surgery alone for resectable liver metastases from colorectal cancer (EORTC 40983): long-term results of a randomised, controlled, Phase 3 trial. Lancet Oncol. 2013;14:1208–1215. doi:10.1016/S1470-2045(13)70447-9
4. Longley DB, Harkin DP, Johnston PG. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003;3(5):330–338. doi:10.1038/nrc1074
5. Meyerhardt JA, Mayer RJ. Systemic therapy for colorectal cancer. N Engl J Med. 2005;352(5):476–487. doi:10.1056/NEJMra040958
6. Ashwal-Fluss R, Meyer M, Pamudurti NR, et al. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell. 2014;56(1):55–66. doi:10.1016/j.molcel.2014.08.019
7. Huang X, Li Z, Zhang Q, et al. Circular RNA AKT3 upregulates PIK3R1 to enhance cisplatin resistance in gastric cancer via miR-198 suppression. Mol Cancer. 2019;18(1):71. doi:10.1186/s12943-019-0969-3
8. Zhou Y, Zheng X, Xu B, et al. Circular RNA hsa_circ_0004015 regulates the proliferation, invasion, and TKI drug resistance of non-small cell lung cancer by miR-1183/PDPK1 signaling pathway. Biochem Biophys Res Commun. 2019;508(2):527–535. doi:10.1016/j.bbrc.2018.11.157
9. Abu N, Hon KW, Jeyaraman S, et al. Identification of differentially expressed circular RNAs in chemoresistant colorectal cancer. Epigenomics. 2019;11(8):875–884. doi:10.2217/epi-2019-0042
10. Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014;15:509–524. doi:10.1038/nrm3838
11. Sun L, Fang Y, Wang X, et al. miR-302a inhibits metastasis and cetuximab resistance in colorectal cancer by targeting NFIB and CD44. Theranostics. 2019;9:8409–8425. doi:10.7150/thno.36605
12. Liang H, Xu Y, Zhang Q, et al. MiR-483-3p regulates oxaliplatin resistance by targeting FAM171B in human colorectal cancer cells. Artif Cells Nanomed Biotechnol. 2019;47:725–736. doi:10.1080/21691401.2019.1569530
13. Wang Y, Lieberman R, Pan J, et al. miR-375 induces docetaxel resistance in prostate cancer by targeting SEC23A and YAP1. Mol Cancer. 2016;15:70. doi:10.1186/s12943-016-0556-9
14. Fu H, Fu L, Xie C, et al. miR-375 inhibits cancer stem cell phenotype and tamoxifen resistance by degrading HOXB3 in human ER-positive breast cancer. Oncol Rep. 2017;37:1093–1099. doi:10.3892/or.2017.5360
15. Shen Y, Wang P, Li Y, et al. miR-375 is upregulated in acquired paclitaxel resistance in cervical cancer. Br J Cancer. 2013;109:92–99. doi:10.1038/bjc.2013.308
16. Cui F, Wang S, Lao I, et al. miR-375 inhibits the invasion and metastasis of colorectal cancer via targeting SP1 and regulating EMT-associated genes. Oncol Rep. 2016;36:487–493. doi:10.3892/or.2016.4834
17. Katoh M, Katoh M. Human FOX gene family (Review). Int J Oncol. 2004;25:1495–1500.
18. Halasi M, Gartel AL. Targeting FOXM1 in cancer. Biochem Pharmacol. 2013;85:644–652. doi:10.1016/j.bcp.2012.10.013
19. Lin JZ, Wang WW, Hu TT, et al. FOXM1 contributes to docetaxel resistance in castration-resistant prostate cancer by inducing AMPK/mTOR-mediated autophagy. Cancer Lett. 2020;469:481–489. doi:10.1016/j.canlet.2019.11.014
20. Liu C, Shi J, Li Q, et al. STAT1-mediated inhibition of FOXM1 enhances gemcitabine sensitivity in pancreatic cancer. Clin Sci (Lond). 2019;133:645–663. doi:10.1042/CS20180816
21. Li D, Ye L, Lei Y, et al. Downregulation of FoxM1 sensitizes nasopharyngeal carcinoma cells to cisplatin via inhibition of MRN-ATM-mediated DNA repair. BMB Rep. 2019;52:208–213. doi:10.5483/BMBRep.2019.52.3.249.
22. Varghese V, Magnani L, Harada-Shoji N, et al. FOXM1 modulates 5-FU resistance in colorectal cancer through regulating TYMS expression. Sci Rep. 2019;9:1505. doi:10.1038/s41598-018-38017-0
23. Meng X, Fu R. miR-206 regulates 5-FU resistance by targeting Bcl-2 in colon cancer cells. Onco Targets Ther. 2018;11:1757–1765. doi:10.2147/OTT.S159093
24. Gillet JP, Gottesman MM. Mechanisms of Multidrug Resistance in Cancer. Multi-Drug Resistance in Cancer. Springer; 2010:47–76.
25. Hua X, Sun Y, Chen J, et al. Circular RNAs in drug resistant tumors. Biomed Pharmacother. 2019;118:109233. doi:10.1016/j.biopha.2019.109233
26. Liu YY, Zhang LY, Du WZ. Circular RNA circ-PVT1 contributes to paclitaxel resistance of gastric cancer cells through regulates ZEB1 expression by sponging miR-124-3p. Biosci Rep. 2019;39. doi:10.1042/BSR20193045.
27. Yan L, Liu G, Cao H, et al. Hsa_circ_0035483 sponges hsa-miR-335 to promote the gemcitabine-resistance of human renal cancer cells by autophagy regulation. Biochem Biophys Res Commun. 2019;519:172–178. doi:10.1016/j.bbrc.2019.08.093
28. Jin C, Wang A, Liu L, et al. Hsa_circ_0136666 promotes the proliferation and invasion of colorectal cancer through miR-136/SH2B1 axis. J Cell Physiol. 2019;234:7247–7256. doi:10.1002/jcp.27482
29. Kulcheski FR, Christoff AP, Margis R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J Biotechnol. 2016;238:42–51. doi:10.1016/j.jbiotec.2016.09.011
30. Li X, Yang B, Ren H, et al. Hsa_circ_0002483 inhibited the progression and enhanced the Taxol sensitivity of non-small cell lung cancer by targeting miR-182-5p. Cell Death Dis. 2019;10:953. doi:10.1038/s41419-019-2180-2
31. Yu W, Peng W, Sha H, et al. Hsa_circ_0003998 promotes chemoresistance via modulation of miR-326 in lung adenocarcinoma cells. Oncol Res. 2019;27:623–628. doi:10.3727/096504018X15420734828058
32. Ward A, Balwierz A, Zhang JD, et al. Re-expression of microRNA-375 reverses both tamoxifen resistance and accompanying EMT-like properties in breast cancer. Oncogene. 2013;32:1173–1182. doi:10.1038/onc.2012.128
33. Xu X, Chen X, Xu M, et al. miR-375-3p suppresses tumorigenesis and partially reverses chemoresistance by targeting YAP1 and SP1 in colorectal cancer cells. Aging (Albany NY). 2019;11:7357–7385. doi:10.18632/aging.102214
34. Xu F, Ye ML, Zhang YP, et al. MicroRNA-375-3p enhances chemosensitivity to 5-fluorouracil by targeting thymidylate synthase in colorectal cancer. Cancer Sci. 2020;2:1–14. doi:10.1111/cas.14356
35. Liu X, Xie T, Mao X, et al. MicroRNA-149 increases the sensitivity of colorectal cancer cells to 5-Fluorouracil by targeting forkhead box transcription factor FOXM1. Cell Physiol Biochem. 2016;39:617–629. doi:10.1159/000445653
36. Zhang H, Yan J, Lang X, et al. Expression of circ_001569 is upregulated in osteosarcoma and promotes cell proliferation and cisplatin resistance by activating the Wnt/beta-catenin signaling pathway. Oncol Lett. 2018;16:5856–5862. doi:10.3892/ol.2018.9410
37. Chen H, Liu S, Li M, et al. circ_0003418 inhibits tumorigenesis and cisplatin chemoresistance through Wnt/beta-Catenin pathway in hepatocellular carcinoma. Onco Targets Ther. 2019;12:9539–9549. doi:10.2147/OTT.S229507
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.