")
Back to Journals » OncoTargets and Therapy » Volume 12
CD48 is a key molecule of immunomodulation affecting prognosis in glioma
Authors Zou C, Zhu C, Guan G, Guo Q, Liu T, Shen S, Yan Z, Xu X, Lin Z, Chen L, Wu A, Cheng W
Received 18 December 2018
Accepted for publication 3 April 2019
Published 28 May 2019 Volume 2019:12 Pages 4181—4193
DOI https://doi.org/10.2147/OTT.S198762
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr XuYu Yang
Cunyi Zou,1,* Chen Zhu,1,* Gefei Guan,1 Qing Guo,1 Tianqi Liu,1 Shuai Shen,1 Zihao Yan,1 Xiaoyan Xu,2 Zhiguo Lin,3 Ling Chen,4 Anhua Wu,1 Wen Cheng1
1Department of Neurosurgery, The First Hospital of China Medical University, Shenyang, Liaoning 110001, People’s Republic of China; 2Department of Pathophysiology, College of Basic Medicine Science, China Medical University, Shenyang, Liaoning 110122, People’s Republic of China; 3Department of Neurosurgery, First Affiliated Hospital of Harbin Medical University, Harbin, Heilongjiang 150001, People’s Republic of China; 4Department of Neurosurgery, General Hospital of People‘s Liberation Army of China (PLA), Medical College of PLA, Institute of Neurosurgery of PLA, Beijing 100853, People’s Republic of China
*These authors contributed equally to this work
Purpose: Glioma is a refractory disease associated with immune cell infiltration, and the effectiveness of checkpoint blockade remains suboptimal. As an adhesion and costimulatory molecule, CD48 plays a significant role in immunomodulation. As such, studying CD48 may provide additional understanding of the immune and inflammation response of glioma.
Methods: Using R language and GraphPad Prism 7, RNA sequencing data of 946 patients from Chinese Glioma Genome Atlas and The Cancer Genome Atlas cohorts were analyzed.
Results: CD48 was highly expressed in the malignant progression of glioma. As an independent risk factor, high-CD48 patients were associated with poor prognosis. CD48 influenced glioma purity and the local immune cell subpopulation. CD48 was closely related to immune function in glioma. Patients with an enhanced immune phenotype, high CD48, were associated with immune suppressive molecules and checkpoints. In addition, CD48 correlated with the immune and inflammatory response. A checkpoint risk score including CD48, SLAMF8 and PD-L1 was used to assess the role of checkpoints. Risk score was particularly high in a malignant subtype of glioma and was an independent predictive indicator of unfavorable outcome. Additionally, age, IDH subtype and MGMT promoter status influenced the predictive significance of checkpoint risk score.
Conclusion: CD48 exhibits a crucial role in reduced survival and immunomodulation in glioma. In addition, we found that checkpoints play a greater role in patients older than 40 years old with IDH wild-type and MGMT methylated status. These findings suggest that combining CD48 blockade with PD-L1 may be a promising approach to glioma immunotherapy for specific subpopulations of patients.
Keywords: CD48, glioma, prognosis, immunomodulation, immunotherapy
Introduction
Diffuse gliomas are common and progressive primary brain tumors that are highly lethal for patients, even for those receiving maximal therapy.1 The median overall survival of glioblastoma (GBM), the most aggressive and invasive type, remains at 14.6 months despite radical radiotherapy and chemotherapy treatment regimes.2 Although novel strategies of glioma immunotherapy are emerging sequentially, such as dendritic cell (DC) therapy and checkpoint blockade, the effectiveness of these novel treatments remains suboptimal at present.3 Therefore, in-depth studies on the immunity status of glioma are urgently needed, in order to identify more effective therapies against this refractory disease.
In glioma, various immune cells and stroma compose the non-tumor component of the tumor parenchyma, such as T cells, tumor-associated macrophages (TAMs) and natural killer (NK) cells. By cross-talking via secretion of cytokines or ligand–receptor interaction, they form a hotbed for the malignant progression of gliomas.4 Moreover, they generate a strong immunosuppressive response by developing multipronged approaches.5 It has been reported that TAMs interact with glioma stem cells to promote tumor progression and immunosuppression.6,7 In addition, immune checkpoints play a significant role in suppressing the antitumor immune response. PD-1/PD-L1 are key regulatory coinhibitory molecules associated with the escape of T and B cell-mediated antitumor immunity in cancer.8 However, the efficacy of glioma immunotherapy against PD-1 has been shown in only a subset of glioma patients and is often accompanied with inflammation and immune-related side effects.3 This indicates that aside from PD-1/PD-L1, there remain other checkpoints that may play an important role in tumor immunity of glioma that are yet to be identified.
Signaling lymphocytic activation molecule family 2 (SLAMF2, CD48) is an adhesion and costimulatory molecule expressed constitutively on most hematopoietic cells, particularly in antigen presenting cells (APC). CD48 binds to CD2 and is involved in a wide variety of innate and adaptive immune responses, ranging from granulocyte activity and allergy to T cell activation and autoimmunity, and CTL or NK function and antimicrobial immunity.9 CD48 expression is increased in autoimmunity and allergy diseases, and anti-CD48 monoclonal antibodies have been shown to attenuate experimental autoimmune encephalomyelitis.10 Recently, it has been demonstrated that CD48 interaction with its high-affinity receptor 2B4 (CD244) leads to monocyte/macrophage-elicited NK cell dysfunction in hepatocellular carcinoma.11 These results suggest that CD48 may play an important role in mediating the immune response in both immune activation or suppression. Nevertheless, there has been little investigation into the immunomodulatory role of CD48 in glioma. Here, we explore the clinical and functional role of CD48 in glioma using RNA sequencing (RNA-seq) data of 946 patients in Chinese Glioma Genome Atlas (CGGA) and The Cancer Genome Atlas (TCGA), which may provide deeper insight into immune checkpoints in glioma treatment.
Material and methods
Patient samples
Our study included 976 glioma samples with detailed clinical information, 310 from the CGGA database and 636 from TCGA database. TCGA cohort was downloaded from public databases (
Bioinformatics analysis
Using methods described previously we calculated stromal and immune scores, glioma purity and microenvironment cell populations.14,15 After Pearson correlation analysis, genes (r >0.4) were chosen for gene ontology (GO) analysis via DAVID (
Statistical analysis
Statistical analysis was carried out using SPSS, GraphPad Prism 7 and R 3.3.3 (
Results
CD48 is enriched in glioblastoma and the mesenchymal subtype
To explore the expression pattern of CD48 in glioma, we examined the RNA-sequencing data in CGGA cohorts. The expression level of CD48 increased along with glioma grade and was most enriched in grade IV glioma, GBM. An investigation of the different molecular subtypes of glioma revealed higher CD48 expression in wild-type IDH, compared to lower grade glioma (LGG) or GBM (Figure 1A). Moreover, we obtained a consistent result in the TCGA database, suggesting that CD48 expression was associated with malignant progression (
 | Figure 1 Expression profile and prognostic value of CD48 in glioma. CD48 was differentially expressed in different grades, histopathologic classifications, molecular and TCGA subtypes (A). CD48 was a good predictor of the mesenchymal subtype (B). Expression of CD48 determined prognosis of glioma in CGGA (C and D) and TCGA (E and F) cohortsAbbreviations: Exp, expression; LGG, lower grade glioma; GBM, glioblastoma multiforme; IDHwt, IDH wild-type; IDHmut, IDH mutant; 1p19qCod, 1p19q codeleted; 1p19qNod, 1p19q non-codeleted; AUC, area under curve; OS, overall survival; HGG, high grade glioma. |
Mesenchymal glioma is the most malignant TCGA subtype. Consistent with this, CD48 was the most enriched in the mesenchymal subtype (CGGA, Figure 1A; TCGA,
CD48 is an independent risk factor for poor outcome
To uncover whether CD48 expression determined prognosis of glioma patients, we compared OS between the low- and high-CD48 groups. In the CGGA cohort, high-CD48 patients had a shorter OS compared to low-CD48 patients in glioma, including high-grade glioma (HGG) (Figure 1C and D). The same result was validated in TCGA cohort (Figure 1E and F). Considering the prognostic importance of CD48, we then assessed the predictive value of CD48 in the CGGA dataset. Results of this analysis indicated that CD48 was an accurate predictor of 1–3-year survival (AUC =77.7%, 79.2% and 81.3%, respectively) (
After univariate and multivariate cox analysis of key clinical and molecular factors, we found that CD48 expression was an independent prognostic factor of glioma in the CGGA dataset (HR =1.202, P=0.006) (Table 1). Consistent with this, CD48 also independently indicated unfavorable prognosis in TCGA dataset (HR =1.144, P=0.006) (
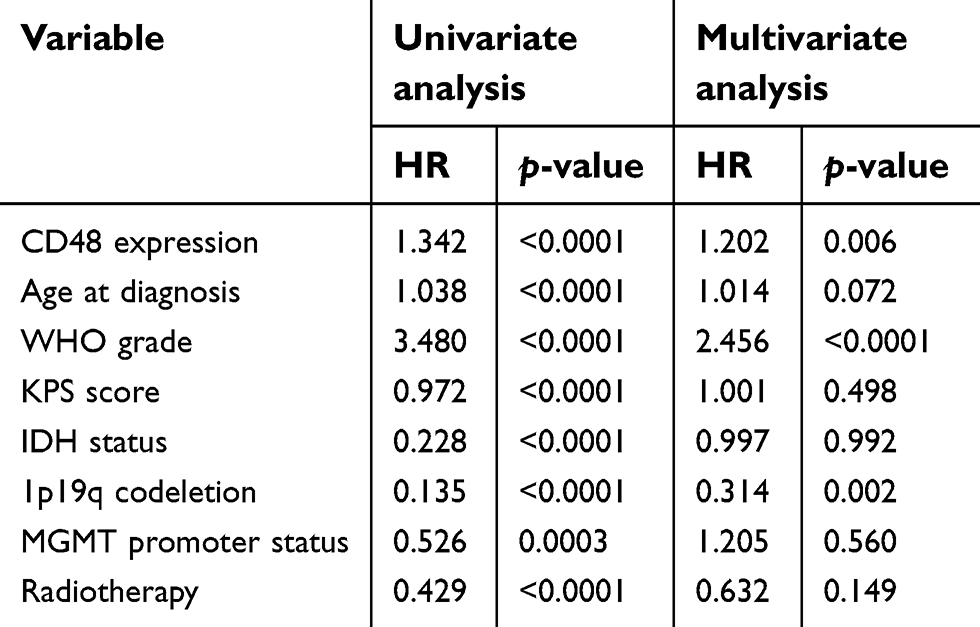 | Table 1 Univariate and multivariate cox analysis of CD48 in CGGA |
CD48 influences glioma purity and local microenvironment cell populations
As an immune costimulatory molecule, we investigated whether CD48 influenced the composition of the tumor environment using glioma purity. We found CD48 had strongly positive correlation with glioma immune score (r=0.717) and stromal score (r=0.571), but negative correlation with glioma purity (r=−0.685) (Figure 2A). Additionally, local microenvironment cell population analysis showed that CD48 was positively associated with an enrichment of myeloid dendritic cells (r=0.470), B lineage (r=0.397), CD8 T cells (r=0.343) and monocytic lineage (r=0.236) (Figure 2A). Taken together, these results indicated that CD48 exerted an important function in influencing the local immune microenvironment of glioma.
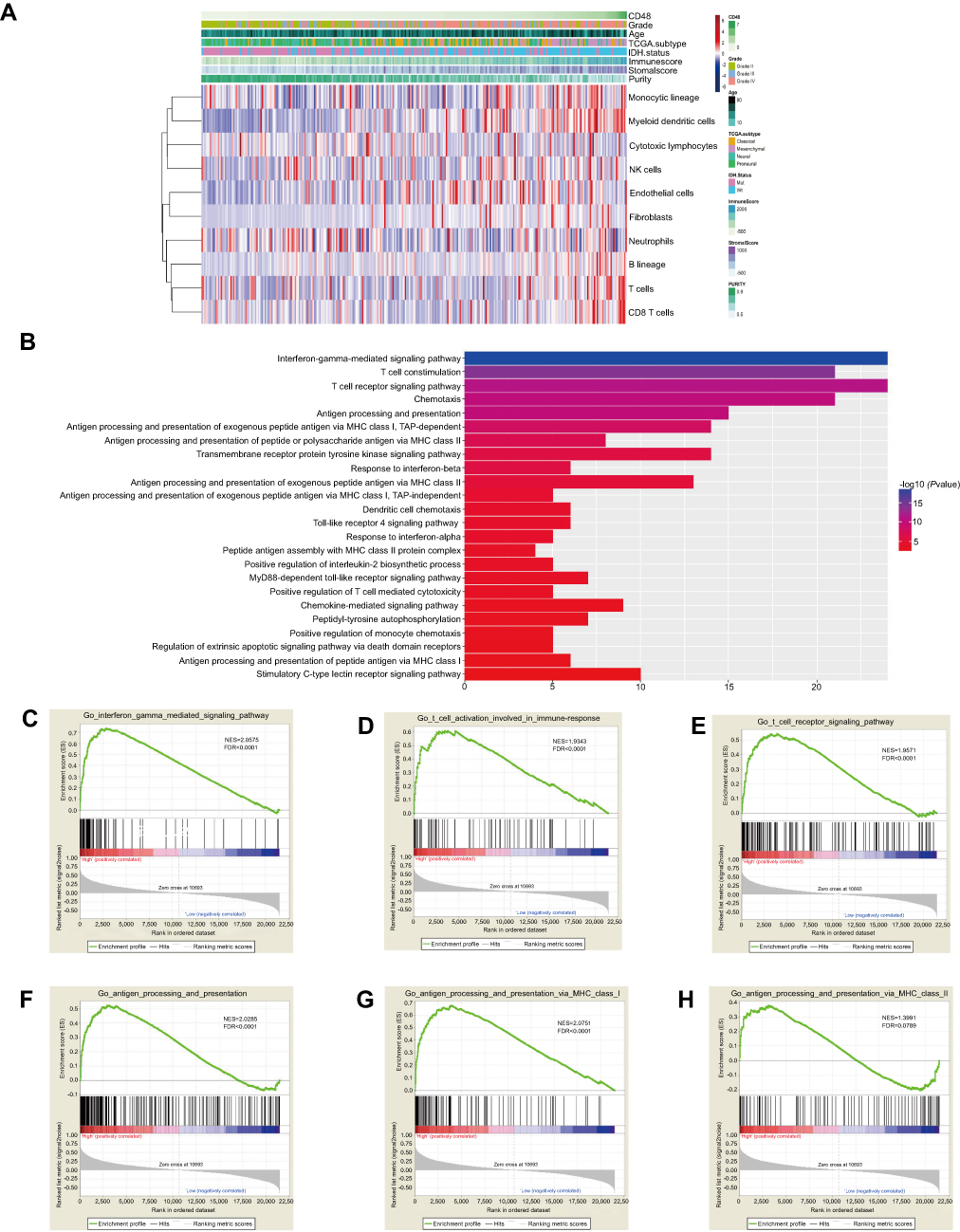 | Figure 2 Microenvironment and biological function of CD48. High CD48 was associated with low purity, enrichment of myeloid dendritic cells, B lineage, CD8 T cells and monocytic lineage (A). Biological function was analyzed by DAVID among correlative genes (B). Biological processes relative to CD48 were validated using GSEA (C–H).Abbreviations: NK, natural killer; GO, gene ontology. |
CD48 is related to immune function
To explore the biological process of CD48, we first made PCA to study transcriptomic features. Based on CD48 expression level, whole transcriptome expression profiles were found to be different between high and low groups (
After correlation analysis, genes strongly correlated with CD48 (r>0.4) in both CGGA and TCGA datasets were selected for functional annotation. These genes were predominately enriched in the interferon-gamma-mediated signaling pathway, T cell costimulation, T cell receptor signaling pathway, chemotaxis and antigen processing and presentation (Figure 2B). GSEA further confirmed that in the high-CD48 group an activated phenotype of antigen processing and presenting, the IFN-γ-mediated signaling pathway, T cell activation and T cell receptor signaling pathway were present (CGGA, Figure 2C–H; TCGA,
CD48 correlates with immune suppressive molecules
Using PCA, we found high-CD48 patients had distinct phenotypes of innate immunity, T cell-mediated immune response and immune response to tumor cells from low-CD48 patients (CGGA, Figure 3A–C; TCGA,
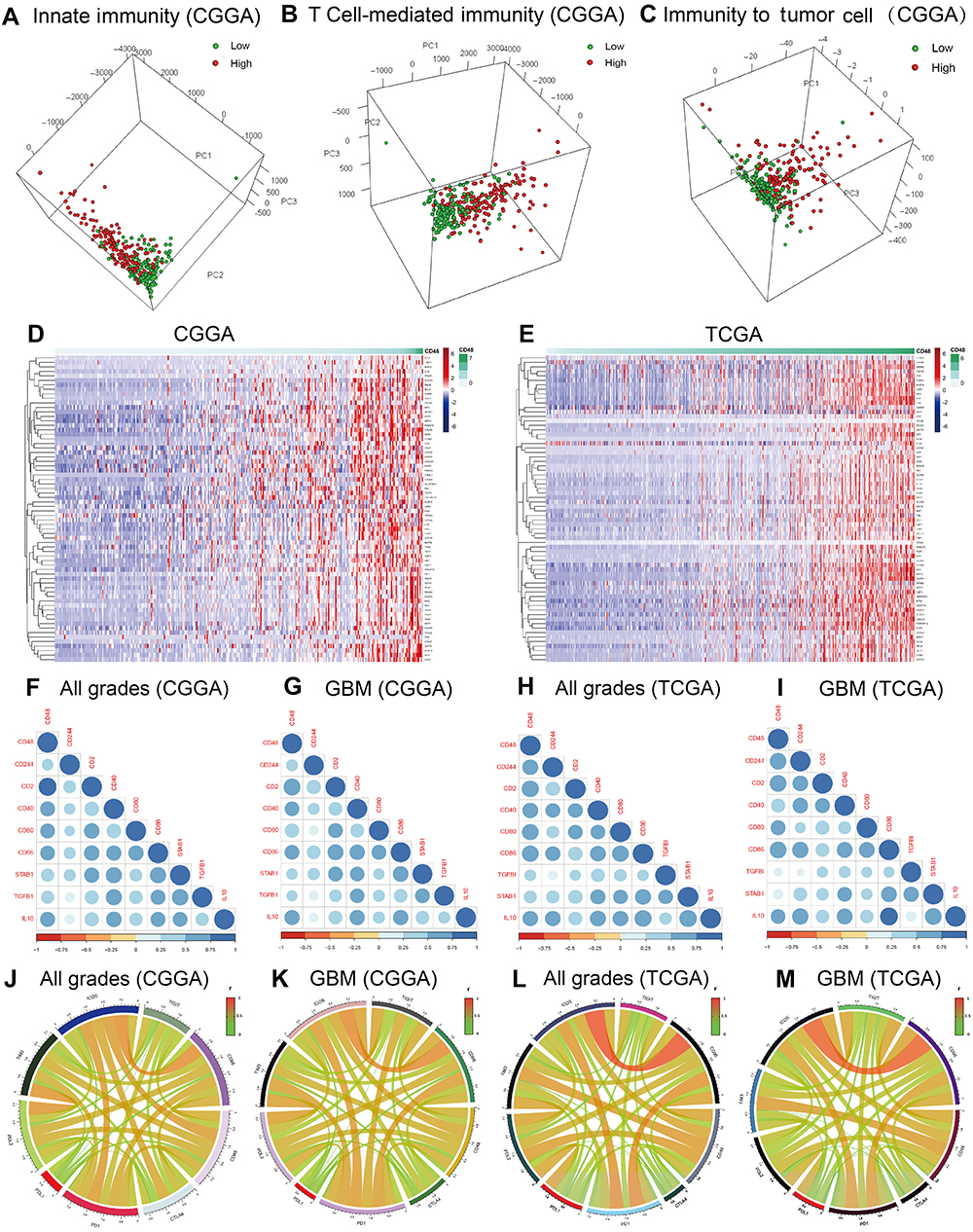 | Figure 3 Immune response is influenced by CD48. Innate immunity, T cell-mediated immunity and immune response to tumor cell were dissimilar between low- and high-CD48 groups in CGGA (A–C). CD48 was closely associated with most immune suppressive factors (D and E). Immune suppressive molecules were positively associated with CD48 (F–I). CD48 was strongly correlated with immune checkpoints (J–M).Abbreviations: CGGA,Chinese Glioma Genome Atlas; TCGA, The Cancer Genome Atlas; GBM, glioblastoma multiforme. |
To confirm our speculation, we further performed analysis of correlation. CD48 was significantly correlated with immune suppressive molecules in all grades of glioma, as well in GBM of CGGA and TCGA cohorts (Figure 3F–I). Moreover, a strong association between CD48 and immune checkpoints, which suppressed antitumor immunity, was seen in glioma and GBM (Figure 3J–M). However, CD48 did not correlate with CTLA-4 and PD-L1. These results implied that CD48 exerted an immunosuppressive role in glioma immunity.
High-CD48 glioma is associated with an enhanced inflammatory response
An inflammatory response is always accompanied by an immune response and plays an important role in the malignant progression of glioma. We chose 7 metagenes of 104 genes representing different types of inflammatory and immune responses;18 CD48 was positively associated with six of these clusters except IgG (Figure 4A and B). Using GO terms involved in the inflammatory response, we performed GSVA to further explore the relation between CD48 and inflammation. We found that CD48 was correlated with inflammation, with no differences observed between chronic or acute inflammatory signatures in glioma (Figure 4C and D). These suggested that CD48 was crucial in promoting inflammation in glioma.
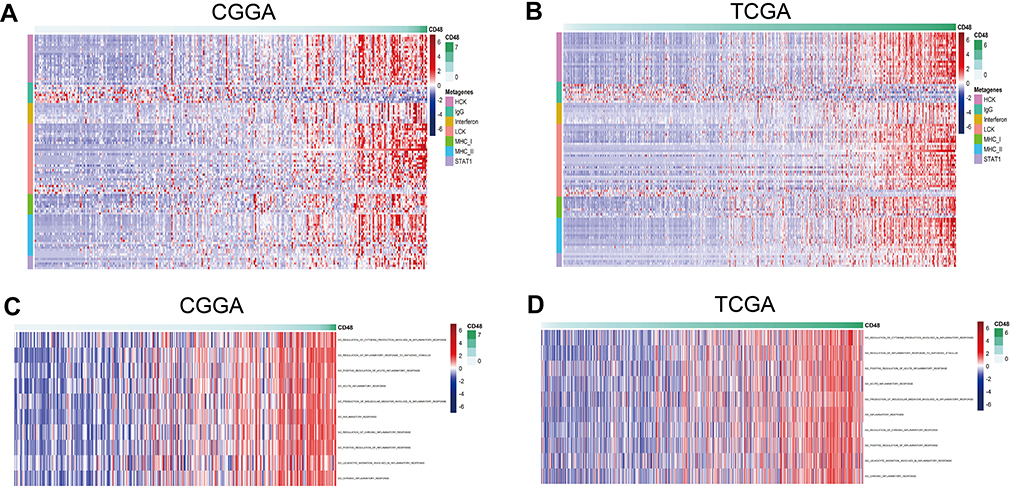 | Figure 4 Association between CD48 and the inflammatory response. Genes involved in the inflammatory response correlated with CD48 in CGGA and TCGA cohorts (A and B). GSVA confirmed correlation between CD48 and the inflammation process, including acute and chronic inflammation (C and D).Abbreviations: CGGA,Chinese Glioma Genome Atlas; TCGA, The Cancer Genome Atlas; GSVA, gene set variation analysis. |
Checkpoint risk signature of CD48, SLAMF8 and PD-L1 reflects glioma prognosis
SLAMF8, a family member molecule of CD48, plays a significant role in both the immune and inflammatory response.19 CD48 and SLAMF8 both were strongly correlated with classical checkpoints, with little association with PD-L1. Therefore, we assembled a risk signature including CD48, SLAMF8 and PD-L1 to assess the role of checkpoints in glioma (Risk score = 0.2029 * PD-L 1+0.2942 * CD48+0.4562 * SLAMF8). The risk score was much higher in malignant progression, such as GBM, IDH-wild type of LGG and GBM, and the mesenchymal subtype (CGGA, Figure 5A; TCGA,
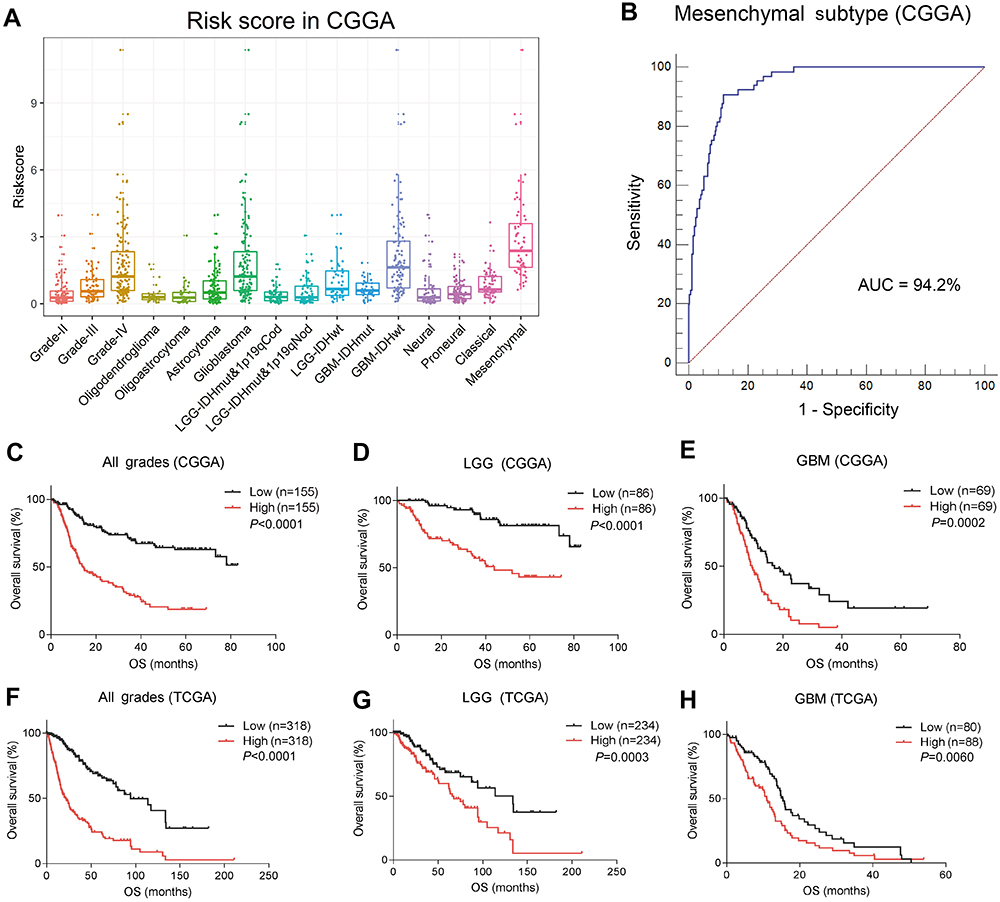 | Figure 5 Distinct distribution and prognosis of checkpoint risk score. Checkpoint risk score exhibited various distributions among grades, histopathologic classifications, molecular and TCGA subtypes (A). Checkpoint risk score was especially high in the mesenchymal subtype (B). High checkpoint risk score was associated with unfavorable outcomes in glioma in CGGA (C–E) and TCGA (F–H) cohorts.Abbreviations: CGGA, China Glioma Genome Atlas; AUC, area under curve; TCGA,The Cancer Genome Atlas; OS, overall survival; LGG,lower grade glioma; GBM, glioblastoma multiforme. |
In all grades of glioma, higher risk score was associated with shorter OS. As such, a higher risk score was correlated with an unfavorable prognosis in LGG and GBM of CGGA datasets (Figure 5C–E). Additionally, these results were validated in the TCGA dataset (Figure 5F–H). After univariate and multivariate cox analyses, we demonstrated that the checkpoint risk score was an independent prognostic factor in CGGA (Table 2) and TCGA (
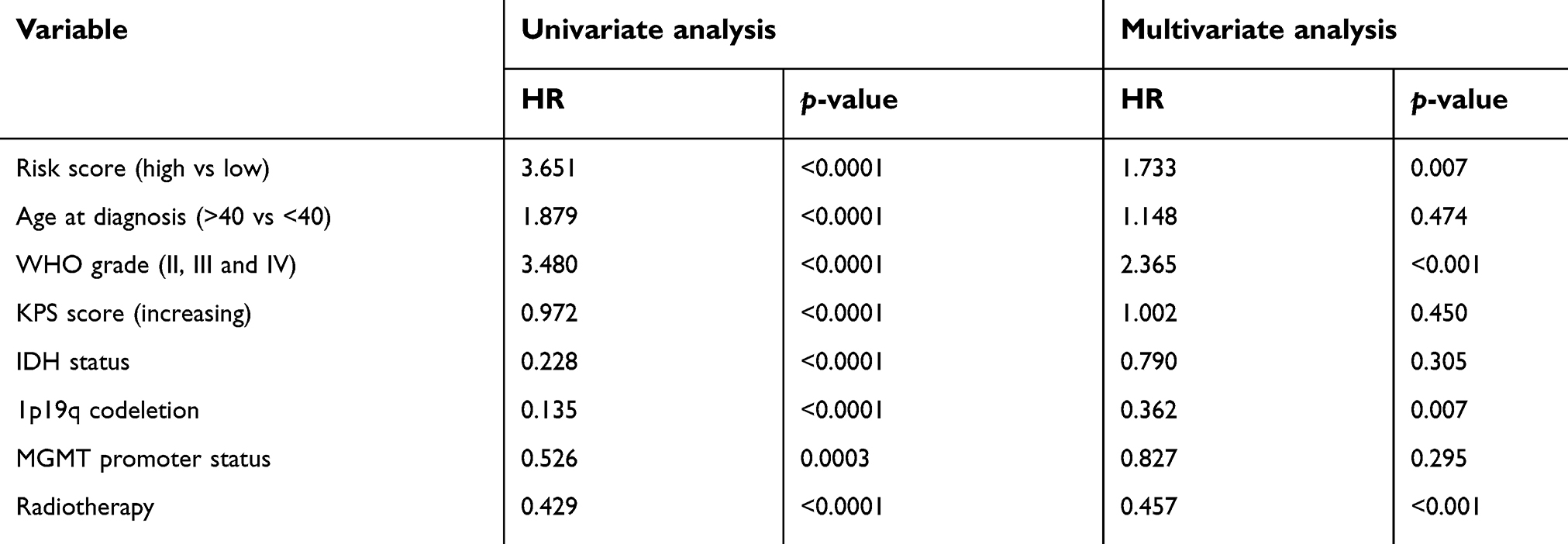 | Table 2 Univariate and multivariate cox analysis of checkpoint risk score in CGGA |
Checkpoint risk signatures have different determined values in different subtypes in GBM
GBM is the most malignant and refractory type of glioma. Recent research has focused on immune checkpoint studies and immunotherapy, which may provide new avenues for treatment of glioma. We identified that checkpoint risk scores had predictive value in both female and male groups (Figure 6A–D). Additionally, similar outcomes were observed in patients younger than 40 between low- and high-risk scores. However, survival differed among patients older than 40 (Figure 6E–H), suggesting that checkpoint factors might influence prognosis in older patients. Considering ethnicity, we found increased risk score indicated shorted survival time in Asian of CGGA (Figure 5E) and in white of TCGA (
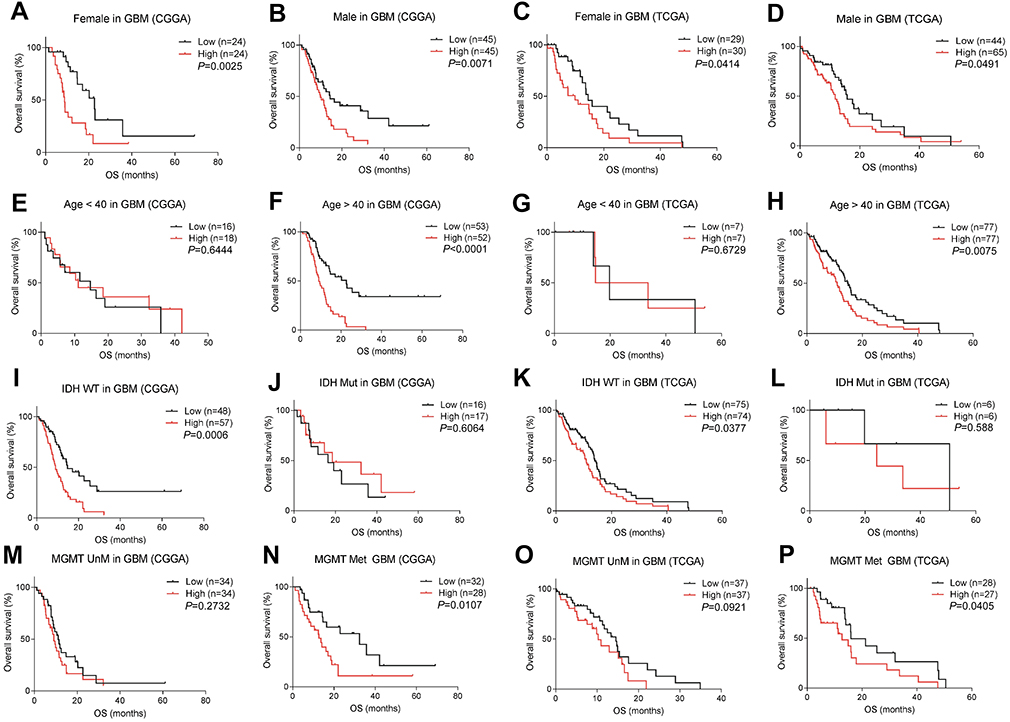 | Figure 6 Prognostic value of checkpoint risk score in different subtypes of GBM. The predictive value of risk score was equal in male and female groups (A–D). Risk score correlated with prognosis among patients older than 40, but not in the group of younger patients (E–H). High-risk score indicated poor prognosis with IDH wild type and MGMT methylation status, but not IDH mutation and MGMT methylation status (I–P).Abbreviations: GBM, glioblastoma multiforme; OS,overall survival; CGGA, Chinese Glioma Genome Atlas; TCGA, The Cancer Genome Atlas; WT, wild-type; Mut, mutant; Met, methylated; unM, unmethylated. |
Next, we investigated the prognostic role of risk score in the different molecular subtypes of GBM. In IDH wild-type GBM, prognosis was significantly different compared to other subtypes, which was attributed to variable risk scores. However, no difference in survival was observed in IDH mutant GBM (Figure 6I–L). Moreover, we observed obvious differences in survival time between low and high-risk score groups in MGMT-methylated patients, but not in GBM with unmethylated MGMT promoter (Figure 6M–P).
Discussion
Malignant glioma is a heterogeneous disease accompanied by the infiltration of multiple immune cells which mount a complex immune response. Immunomodulation among immune cells or between tumor cells and immune cells plays a significant role in promoting progression of neoplasm.20 Checkpoints are key molecules in immunomodulation, and checkpoint blockade shows significant potential for immune therapy of glioma.21 Here, we demonstrate that the expression of the adhesion and costimulatory molecule of immune cells, CD48, correlated with malignant phenotype and unfavorable prognosis. Identification and characterization of such molecules may aid in our understanding of immune mechanisms in glioma, paving the way for novel treatments.
After analysis of glioma RNA-seq data of 946 patients, we identified that CD48 expression was increased along with glioma grade and was particularly enriched in GBM. CD48 has been reported to be highly expressed in breast cancer, with high tumor-infiltrating lymphocytes (TIL), as one molecule of eight-gene TIL signature (including CD19, CD3D, CD48, GZMB, LCK, MS4A1, PRF1 and SELL),22 which suggested that CD48 is commonly enriched in more malignant tumors associated with TIL. We found CD48 was enriched in IDH-wild glioma and mesenchymal subtypes, which indicates involvement in an active immune response.23,24 In addition, it has been reported that CD48 had a positive correlation with the disease status of primary Sjögren’s syndrome, in both serum and salivary glands.25,26 Taken together, these findings indicate that CD48 is highly expressed in the immune and inflammatory response, as well as in glioma. As such, the detection of CD48 in serum may be an indicator to reflect the immune and inflammation status of glioma patients; however, further study is needed.
Previously, we found that immune-related factors, STAB1, TGF-β and IL-10 play crucial roles in controlling the development and prognosis of glioma cells.27,28 Jiang’s team has reported that coinhibitory checkpoints, PD-1,29 PD-L130 and TIM-316 are independent prognostic risk factors with immune-related function in glioma. Here, we confirmed that the costimulatory molecule CD48 also predicted prognosis of glioma independently, with high-CD48 expressing glioma associated with shorter OS. These results suggest that immune-related molecules play a crucial role in promoting the malignant progress of gliomas and despite the presence of the blood–brain barrier, glioma still could promote tumor development by immunomodulation.
Glioma is a heterogeneous tumor with a complex microenvironment. Infiltrating stromal and immune cells form the major fraction of non-tumor cells, such as endothelial cells, T cells and TAMs and so on, diluting the purity of tumor cells. Higher levels of immature blood vessels and TAMs serve as robust indicators of poor prognosis.31,32 Glioma purity is an intrinsic characteristic of this tumor in the development of a suitable microenvironment. Macrophages, microglia and neutrophils are enriched in low purity glioma. Low glioma purity means malignant entities, reduced survival time and intensive immune phenotypes.14 A positive correlation exists between CD48 and immune score and stromal score, and a negative correlation exists between CD48 and glioma purity. This suggests that CD48 expression in glioma may influence the composition of the tumor microenvironment to boost the advancement of glioma.
Myeloid dendritic cells, B lineage, CD8 T cells and monocytic lineage, which are the main functional immune cells during the anti-tumor immune response, were enriched in high-CD48 glioma. T cells are functional immune cells of adaptive immunity that can kill tumor cells with the costimulatory signal from APC through MHC molecules. Dendritic cells and monocyte-macrophage lineages are the main APC. As a costimulatory signaling molecule, CD48 may interact with its ligand to regulate immunological activity of these immune cells. GO analysis confirmed that CD48 was involved in antigen processing and presentation, T cell costimulation and the T cell receptor signaling pathway. High-CD48 glioma correlated with antigen processing and presenting via MHC I, which interacts with CD8 T cells. Considering the local microenvironment cell population, these results verified that CD48 plays a significant role in immunomodulation of CD8 T cell-mediated immunity and may be involved in anti-tumor immunity.
Although CD48 has little correlation with the enrichment of NK cells in glioma, CD48 is reported to regulate activity and NK cell function.33 NK cells can control innate immunity and also adaptive immunity through receptor/ligand-specific cross talk, mainly CD48/2B4.34 Additionally, it has been reported that CD48 prolonged the time frame of effective T cell activation by supporting mature DC survival.35 In glioma, high-CD48 glioma followed a more activated innate immunity and T cell-mediated adaptive immunity, consistent with our previous finding that malignant gliomas are associated with a more enhanced immune phenotype.36 However, other research has demonstrated that CD48 also inhibits the activity of NK cells.11 Therefore, we suggest that high-CD48 glioma is associated with stronger immunosuppression at the same time.
Studies on immunosuppression have revealed that ligation of CD80 and CD86 on T cells by the cell surface molecule CTLA-4 on suppressor cells results in suppression of T cell activation.37 Suppression of primed immune responses by DCs correlates with ReIB nuclear binding activity and CD40 expression, with immature DCs also inducing CD8+ T regulatory cells to produce high levels of IL-10.38 Regulatory CD4+ T cells induced by the DCs transfer tolerance to primed recipients in an IL-10-dependent fashion.39 TGF-β could suppress T-cell proliferation and suppression of CD8+ T cells that mediate tumor rejection by CD4+CD25+. T cells require TGF-β to interact with CD8+ T cells.40 These suppressive molecules are positively associated with CD48 in our study, suggesting that CD48 works with them to promote immunosuppression.
Immune checkpoints have been shown to play immunosuppressive roles in cancer, such as CTLA-4, PD-1, PD-L1/2, Tim-3, ICOS, TIGIT and CD96. These checkpoint blockades are under investigation, and new inhibitory pathways are being investigated.41,42 CD48 has a strong association with most checkpoints, but little with CTLA-4 and PD-L1, which demonstrates that CD48 may have a different immunomodulating role from CTLA-4 and PD-L1. While therapies targeting CTLA-4 and PD-1 have shown remarkable success in glioma mouse models, clinical patients benefit little from these treatments,43 and some patients experience serious immune-related side effects or even fatal cerebral edema and myocarditis.44,45 High-CD48 gliomas revealed close association with the inflammatory response. Therefore, we suggest that multiple checkpoint blockades may promote antitumor immunity without excessive inflammation. As such, CD48 may be an appropriate target, potentially by use of an inhibitor against PD-L1 or CTLA-4.
Immunotherapy, in particular checkpoint blockade, has become the current focus of cancer research. Checkpoint risk signature including PD-L1, CD48 and SLAMF8 was confirmed as an independent risk factor of glioma, correlating with malignant progression. These results emphasize that risk factor immune checkpoints exert significant influence in determining development and outcome of glioma. Moreover, checkpoint risk score was an accurate predictor of the mesenchymal subtype, further indicating that the mesenchymal subtype is accompanied by a more complex immune microenvironment and immune response. This suggests that the mesenchymal subtype of GBM may be more suitable for immunotherapy.
Considering the factors of gender and age, we found checkpoint risk score had a crucial influence on prognosis in groups of patients older than 40, but not in groups of young patients. This indicates that age is an important factor in influencing tumor immunity, and older patients might have a better response to checkpoint blockade immunotherapy than young patients, which may be due to the increased expression of regulatory immune checkpoints on T cells with age in human and mice.46,47 However, patients older than 75 are associated with a decline in the immune system, which theoretically can reduce the efficacy of immunotherapy.48 In addition, our result confirmed that checkpoint risk score plays an important role in IDH wild-type GBM rather than mutant IDH. IDH wild-type GBM presents with a more complex microenvironment and stronger immune response,24 which results in an unfavorable outcome.49 Therefore, checkpoint blockade may be the optimal approach to block immunosuppression and activate effective antitumor immunity.
Conclusion
Through multiple analysis of large glioma cohorts, we found the costimulatory and adhesion molecule CD48 correlated with malignant progression and poor prognosis. By influencing local immune microenvironments, CD48 plays an important role in antigen presenting and T cell activation. With an active immune phenotype, CD48 was strongly associated with immunosuppression and inflammatory responses. Further studies will need to further investigate the immunomodulatory role of CD48 in antitumor immunity of glioma. Due to its role in immunomodulation, CD48 may become an ideal target by checkpoint blockade, which is most suitable for IDH wild-type GBM patients above 40 years old.
Ethical approval
This article does not contain any studies with human participants or animals.
Acknowledgments
We thank all the members of Dr Anhua Wu’s laboratory for help with our study. This work was supported by grants from the National Natural Science Foundation of China (grant numbers: 81172409, 81472360, 81402045, and 81872054) and the Science and Technology Department of Liaoning Province (grant number: 2011225034).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rich JN, Bigner DD. Development of novel targeted therapies in the treatment of malignant glioma. Nat Rev Drug Discovery. 2004;3(5):430–446. doi:10.1038/nrd1380
2. Ladomersky E, Genet M, Zhai L, et al. Improving vaccine efficacy against malignant glioma. Oncoimmunology. 2016;5:e1196311. doi:10.1080/2162402X.2016.1196311
3. Lucca LE, Hafler DA. Co-inhibitory blockade while preserving tolerance: checkpoint inhibitors for glioblastoma. Immunol Rev. 2017;276:9–25. doi:10.1111/imr.12529
4. Yan D, Kowal J, Akkari L, et al. Inhibition of colony stimulating factor-1 receptor abrogates microenvironment-mediated therapeutic resistance in gliomas. Oncogene. 2017;36:6049–6058. doi:10.1038/onc.2017.261
5. Kamran N, Kadiyala P, Saxena M, et al. Immunosuppressive myeloid cells‘ blockade in the glioma microenvironment enhances the efficacy of immune-stimulatory gene therapy. Mol Ther. 2017;25:232–248. doi:10.1016/j.ymthe.2016.10.003
6. Zhou W, Ke SQ, Huang Z, et al. Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat Cell Biol. 2015;17:170–182. doi:10.1038/ncb3090
7. Jia XH, Feng GW, Wang ZL, et al. Activation of mesenchymal stem cells by macrophages promotes tumor progression through immune suppressive effects. Oncotarget. 2016;7:20934–20944. doi:10.18632/oncotarget.8064
8. Zuazo M, Gato-Canas M, Llorente N, et al. Molecular mechanisms of programmed cell death-1 dependent T cell suppression: relevance for immunotherapy. Ann Transl Med. 2017;5:385. doi:10.21037/atm.2017.06.11
9. McArdel SL, Terhorst C, Sharpe AH. Roles of CD48 in regulating immunity and tolerance. Clin Immunol. 2016;164:10–20. doi:10.1016/j.clim.2016.01.008
10. McArdel SL, Brown DR, Sobel RA, Sharpe AH. Anti-CD48 monoclonal antibody attenuates experimental autoimmune encephalomyelitis by limiting the number of pathogenic CD4+ T cells. J Immunol. 2016;197:3038–3048. doi:10.4049/jimmunol.1600706
11. Wu Y, Kuang DM, Pan WD, et al. Monocyte/macrophage-elicited natural killer cell dysfunction in hepatocellular carcinoma is mediated by CD48/2B4 interactions. Hepatology. 2013;57:1107–1116. doi:10.1002/hep.26192
12. Cheng W, Li M, Jiang Y, et al. Association between small heat shock protein B11 and the prognostic value of MGMT promoter methylation in patients with high-grade glioma. J Neurosurg. 2016;125:7–16. doi:10.3171/2015.5.JNS142437
13. Cheng W, Li M, Cai J, et al. HDAC4, a prognostic and chromosomal instability marker, refines the predictive value of MGMT promoter methylation. J Neurooncol. 2015;122:303–312. doi:10.1007/s11060-014-1709-6
14. Zhang C, Cheng W, Ren X, et al. Tumor purity as an underlying key factor in glioma. Clin Cancer Res. 2017;23:6279–6291. doi:10.1158/1078-0432.CCR-16-2598
15. Becht E, Giraldo NA, Lacroix L, et al. Estimating the population abundance of tissue-infiltrating immune and stromal cell populations using gene expression. Genome Biol. 2016;17:218. doi:10.1186/s13059-016-1070-5
16. Li G, Wang Z, Zhang C, et al. Molecular and clinical characterization of TIM-3 in glioma through 1,024 samples. Oncoimmunology. 2017;6:e1328339. doi:10.1080/2162402X.2017.1328339
17. Cai J, Chen Q, Cui Y, et al. Immune heterogeneity and clinicopathologic characterization of IGFBP2 in 2447 glioma samples. Oncoimmunology. 2018;7:e1426516. doi:10.1080/2162402X.2018.1490854
18. Rody A, Holtrich U, Pusztai L, et al. T-cell metagene predicts a favorable prognosis in estrogen receptor-negative and HER2-positive breast cancers. Breast Cancer Res. 2009;11:R15. doi:10.1186/bcr2234
19. Zou CY, Guan GF, Zhu C, et al. Costimulatory checkpoint SLAMF8 is an independent prognosis factor in glioma. CNS Neurosci Ther. 2019 Mar;25(3):333–342. doi:10.1111/cns.13041
20. Kamran N, Alghamri MS, Nunez FJ, et al. Current state and future prospects of immunotherapy for glioma. Immunotherapy. 2018;10:317–339. doi:10.2217/imt-2017-0122
21. Saha D, Martuza RL, Rabkin SD. Macrophage polarization contributes to glioblastoma eradication by combination immunovirotherapy and immune checkpoint blockade. Cancer Cell. 2017;32:253–267.e255. doi:10.1016/j.ccell.2017.07.006
22. West NR, Milne K, Truong PT, Macpherson N, Nelson BH, Watson PH. Tumor-infiltrating lymphocytes predict response to anthracycline-based chemotherapy in estrogen receptor-negative breast cancer. Breast Cancer Res. 2011;13:R126. doi:10.1186/bcr3052
23. Carro MS, Lim WK, Alvarez MJ, et al. The transcriptional network for mesenchymal transformation of brain tumours. Nature. 2010;463:318–325. doi:10.1038/nature08712
24. Amankulor NM, Kim Y, Arora S, et al. Mutant IDH1 regulates the tumor-associated immune system in gliomas. Genes Dev. 2017;31:774–786. doi:10.1101/gad.294991.116
25. Nishikawa A, Suzuki K, Kassai Y, et al. Identification of definitive serum biomarkers associated with disease activity in primary Sjogren‘s syndrome. Arthritis Res Ther. 2016;18:106. doi:10.1186/s13075-016-1006-1
26. Aqrawi LA, Jensen JL, Oijordsbakken G, et al. Signalling pathways identified in salivary glands from primary Sjogren‘s syndrome patients reveal enhanced adipose tissue development. Autoimmunity. 2018;1–12. doi:10.1080/08916934.2018.1446525
27. Han S, Xia J, Qin X, Han S, Wu A. Phosphorylated SATB1 is associated with the progression and prognosis of glioma. Cell Death Dis. 2013;4:e901. doi:10.1038/cddis.2013.433
28. Qiu B, Zhang D, Wang C, et al. IL-10 and TGF-beta2 are overexpressed in tumor spheres cultured from human gliomas. Mol Biol Rep. 2011;38:3585–3591. doi:10.1007/s11033-010-0469-4
29. Liu S, Wang Z, Wang Y, et al. PD-1 related transcriptome profile and clinical outcome in diffuse gliomas. Oncoimmunology. 2018;7:e1382792. doi:10.1080/2162402X.2018.1490854
30. Wang Z, Zhang C, Liu X, et al. Molecular and clinical characterization of PD-L1 expression at transcriptional level via 976 samples of brain glioma. Oncoimmunology. 2016;5:e1196310. doi:10.1080/2162402X.2016.1196310
31. Jia ZZ, Gu HM, Zhou XJ, et al. The assessment of immature microvascular density in brain gliomas with dynamic contrast-enhanced magnetic resonance imaging. Eur J Radiol. 2015;84:1805–1809. doi:10.1016/j.ejrad.2015.05.035
32. Wang H, Zhang L, Zhang IY, et al. S100B promotes glioma growth through chemoattraction of myeloid-derived macrophages. Clin Cancer Res. 2013;19:3764–3775. doi:10.1158/1078-0432.CCR-12-3725
33. Lee KM, Forman JP, McNerney ME, et al. Requirement of homotypic NK-cell interactions through 2B4(CD244)/CD48 in the generation of NK effector functions. Blood. 2006;107:3181–3188. doi:10.1182/blood-2005-01-0185
34. Assarsson E, Kambayashi T, Schatzle JD, et al. NK cells stimulate proliferation of T and NK cells through 2B4/CD48 interactions. J Immunol. 2004;173:174–180. doi:10.4049/jimmunol.173.1.174
35. Kis-Toth K, Tsokos GC. Engagement of SLAMF2/CD48 prolongs the time frame of effective T cell activation by supporting mature dendritic cell survival. J Immunol. 2014;192:4436–4442. doi:10.4049/jimmunol.1302909
36. Cheng W, Ren X, Zhang C, et al. Bioinformatic profiling identifies an immune-related risk signature for glioblastoma. Neurology. 2016;86:2226–2234. doi:10.1212/WNL.0000000000002770
37. Paust S, Lu L, McCarty N, Cantor H. Engagement of B7 on effector T cells by regulatory T cells prevents autoimmune disease. Proc Natl Acad Sci U S A. 2004;101:10398–10403. doi:10.1073/pnas.0403342101
38. Dhodapkar MV, Steinman RM, Krasovsky J, Munz C, Bhardwaj N. Antigen-specific inhibition of effector T cell function in humans after injection of immature dendritic cells. J Exp Med. 2001;193:233–238.
39. Martin E, O‘Sullivan B, Low P, Thomas R. Antigen-specific suppression of a primed immune response by dendritic cells mediated by regulatory T cells secreting interleukin-10. Immunity. 2003;18:155–167.
40. von Boehmer H. Mechanisms of suppression by suppressor T cells. Nat Immunol. 2005;6:338–344. doi:10.1038/ni1180
41. Marin-Acevedo JA, Dholaria B, Soyano AE, Knutson KL, Chumsri S, Lou Y. Next generation of immune checkpoint therapy in cancer: new developments and challenges. J Hematol Oncol. 2018;11:39. doi:10.1186/s13045-018-0582-8
42. Dougall WC, Kurtulus S, Smyth MJ, Anderson AC. TIGIT and CD96: new checkpoint receptor targets for cancer immunotherapy. Immunol Rev. 2017;276:112–120. doi:10.1111/imr.12518
43. Boussiotis VA, Charest A. Immunotherapies for malignant glioma. Oncogene. 2017. doi:10.1038/s41388-41017-40024-z
44. Zhu X, McDowell MM, Newman WC, Mason GE, Greene S, Tamber MS. Severe cerebral edema following nivolumab treatment for pediatric glioblastoma: case report. J Neurosurgery Pediatrics. 2017;19:249–253. doi:10.3171/2016.8.PEDS16326
45. Moslehi JJ, Salem JE, Sosman JA, Lebrun-Vignes B, Johnson DB. Increased reporting of fatal immune checkpoint inhibitor-associated myocarditis. Lancet. 2018;391:933. doi:10.1016/S0140-6736(18)30302-7
46. Canaday DH, Parker KE, Aung H, Chen HE, Nunez-Medina D, Burant CJ. Age-dependent changes in the expression of regulatory cell surface ligands in activated human T-cells. BMC Immunol. 2013;14:45. doi:10.1186/1471-2172-14-45
47. Hurez V, Daniel BJ, Sun L, et al. Mitigating age-related immune dysfunction heightens the efficacy of tumor immunotherapy in aged mice. Cancer Res. 2012;72:2089–2099. doi:10.1158/0008-5472.CAN-11-3019
48. Elias R, Karantanos T, Sira E, Hartshorn KL. Immunotherapy comes of age: immune aging & checkpoint inhibitors. J Geriatr Oncol. 2017;8:229–235. doi:10.1016/j.jgo.2017.02.001
49. Cheng W, Ren X, Zhang C, Cai J, Han S, Wu A. Gene expression profiling stratifies IDH1-mutant glioma with distinct prognoses. Mol Neurobiol. 2017;54:5996–6005. doi:10.1007/s12035-016-0150-6
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.