")
Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 18
Bufei Yishen Formula Inhibits the Cell Senescence in COPD by Up-Regulating the ZNF263 and Klotho Expression
Authors Wang W , Zhang S, Cui L, Chen Y, Xu X, Wu L
Received 30 July 2022
Accepted for publication 24 February 2023
Published 8 April 2023 Volume 2023:18 Pages 533—539
DOI https://doi.org/10.2147/COPD.S383295
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Richard Russell
Weimin Wang,1,* Shaohong Zhang,1,* Lei Cui,2 Yu Chen,2 Xingxing Xu,2 Longchuan Wu2
1Department of Gerontology, The Affiliated Huaian No.1 People’s Hospital of Nanjing Medical University, Huaian, People’s Republic of China; 2Department of Respiratory Medicine, Huaian Hospital of Traditional Chinese Medicine, Affiliated to Nanjing University of Traditional Chinese Medicine, Huaian, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Longchuan Wu; Xingxing Xu, Email [email protected]; [email protected]
Background: Bufei Yishen formula (BYF) is an effective prescription for the clinical treatment of chronic obstructive pulmonary disease (COPD). However, the molecular mechanism by which it exerts its pharmacological effects remains to be explored.
Methods: The human bronchial cell line BEAS-2B was treated with cigarette smoke extract (CSE). Cellular senescence markers were detected by Western blot and ELISA. Potential transcription factor of klotho was predicted using JASPAR and USCS databases.
Results: CSE induced cellular senescence with intracellular accumulation of cellular senescence biomarkers (p16, p21 and p27) and increased secretion of senescence-related secretory phenotypic (SASP) factors (IL-6, IL-8, and CCL3). In contrast, BYF treatment inhibited CSE-induced cellular senescence. CSE suppressed the transcription, expression and secretion of klotho, whereas BYF treatment rescued its transcription, expression and secretion. CSE downregulated the protein level of ZNF263, whereas BYF treatment rescued the expression of ZNF263. Furthermore, ZNF263-overexpressing BEAS-2B cells could inhibit CSE-induced cellular senescence and SASP factor secretion by upregulating the expression of klotho.
Conclusion: This study revealed a novel pharmacological mechanism by which BYF alleviates clinical symptoms of COPD patients, and regulating ZNF263 and klotho expression may be beneficial to the treatment and prevention of COPD.
Keywords: Bufei Yishen formula, cell senescence, ZNF263, klotho, senescence-related secretory phenotypic
Introduction
Chronic obstructive pulmonary disease (COPD) is a respiratory disease characterized by airflow limitation that is not fully reversible. COPD affects the health and lives of more than 300 million people worldwide and is the third leading cause of death worldwide after ischemic heart disease and stroke.1 COPD imposes a huge economic and social burden and has become a major public health problem. In addition, its morbidity and mortality continue to rise. Currently, no pharmacological treatment is available to reduce the disease progression and mortality. Treatment strategies mainly focus on symptomatic treatment to improve the life quality of patient.2
Bufei Yishen Formula (BYF) is an effective formula for the clinical treatment of COPD, which can effectively alleviate the clinical symptoms of patients, and improve their exercise tolerance and life quality.3 In addition, it can reduce the severity, frequency, and volume of cough and improve wheezing symptoms.3 However, the molecular mechanism by which BYF exerts its pharmacological effects remains to be explored.
Cellular senescence is a state of irreversible permanent cell cycle arrest. Cyclin-dependent kinases inhibitory factors (p16, p21 and p27) accumulate in senescent cells. In addition, senescent cells secrete a variety of senescence-associated secretory phenotypes (SASP) of inflammatory proteins, leading to low-grade chronic inflammation. COPD is described as “accelerated lung aging disease”. Accumulated senescent cells, including alveolar epithelial cells and endothelial cells, are found in the lungs of COPD patients.4 The accumulation of senescent cells may lead to small airway fibrosis and alveolar cell loss in COPD patients.5 Cigarette smoke exposure induced oxidative stress may be an important trigger of COPD aging.6 To date, no studies have reported the effect of BYF on COPD-related cellular senescence.
The klotho gene is a senescence suppressor gene that encodes a unidirectional transmembrane protein that has been shown to be a key regulator of oxidative stress and cellular senescence. Furthermore, the extracellular domain of the klotho protein can be cleaved and released as a soluble protein, which can be detected in cerebrospinal fluid and blood. Soluble klotho protein can act as an independent hormonal factor that regulates the activity of various growth factor signaling pathways (insulin/IGF-1 and Wnt) and various ion channels. It has been reported that the expression of klotho is significantly decreased in alveolar macrophages and airway epithelial cells of COPD patients.7–9 Our previous research showed that cigarette smoke extract (CSE) can significantly inhibit the expression of klotho in bronchial cells.10 Increasing klotho levels in lung epithelial cells reduces CSE-induced inflammation, oxidative stress and cellular senescence.10,11 However, the regulatory mechanisms underlying the downregulation of klotho expression in COPD remain unknown.
In this study, we will use CSE-induced human bronchial cell lines in vitro to explore the effect of BYF on COPD- associated cellular senescence and the regulatory mechanism of the klotho expression down-regulation in COPD.
Materials and Methods
Preparation of CSE
The smoke from the combustion of one Marlboro cigarette (0.8 mg nicotine and 10 mg tar) was dissolved in 10 mL medium within 2 min using negative pressure. The pH value of the solution was adjusted to 7.4 with hydrochloric acid. The absorbance of the solution at 320 nm was measured by spectrophotometer, and adjusted to 1.8–2.0 with the medium. Subsequently, the solution was filtered through a 0.22 μm membrane. The solution was considered to be 100% CSE. The CSE used for the experiments in this study was 10% CSE diluted with culture medium, and the cells were induced for 24 h for subsequent detection.12,13 CSE was prepared fresh for each experiment, and cells were processed within 15 min.
Preparation of BYF
72 g crude drugs (15 g Astragali Radix, 15 g Ginseng Radix et Rhizoma, 12 g Atractylodes macrocephala, 10 g Radix Rehmanniae Preparata, 10 g Fructus Corni and 10 g Herba Epimedii) were boiled in 1 L distilled water for 2 h, and filtered to remove drug residues. All crude drugs were provided by the Chinese Pharmacy of Huai’an Hospital of Traditional Chinese Medicine. The liquid concentration was adjusted to 72 g/144 mL (0.5 g/mL) and filtered through a 0.22 μm membrane. The BYF concentration used in this study was 100 μg/mL, and the cells were supplemented with BYF in parallel with CSE.
Cell Culture and Transfection
The human bronchial cell line BEAS-2B (ATCC) was cultured in DMEM containing 10% FBS at 5% CO2 and 37 °C. Transfection of pcDNA3.1 ZNF263 overexpression plasmid was performed using Lipofectamine 2000 (Invitrogen Biotechnology Co., Ltd., USA) according to the manufacturer’s instructions.
Western Blot
Cells were lysed on ice using RIPA Lysis Buffer. Total protein was collected, and impurities were removed by centrifugation at 12,000 g for 10 min at 4°C. Equal amounts of protein were separated by SDS-PAGE and electroporated onto PVDF membranes. The membranes were blocked in 5% nonfat milk for 1 h and incubated with primary antibody overnight at 4°C. Subsequently, the membrane was incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. Finally, the signals were visualized by Enhanced Chemiluminescence, and the band density was quantified using Image J software.
Enzyme-Linked Immunosorbent Assay (ELISA)
Concentrations of IL-6 (ab178013, Abcam), IL-8 (b214030, Abcam) and CCL3 (KE00092, Proteintech) in the culture medium were measured using Human ELISA Kits according to the manufacturer’s recommendations.
Quantitative Real-Time PCR (RT-qPCR)
Total RNA was isolated from cells using Trizol reagent and reverse transcribed into cDNA. RT-qPCR was performed on a 7500 Real-Time PCR System (Applied Biosystems) using SYBR Green Master Mix. The relative expression of target gene was analyzed using the 2−ΔΔCT method, and normalized to GAPDH.
Statistical Analysis
All experiments were independently repeated three times. Data were analyzed using GraphPad Prism 8.0, and values are expressed as mean ± SD. Comparisons between two groups were made using Two-tailed t-test. P < 0.05 was considered statistically significant.
Results
BYF Inhibits CSE-Induced Cellular Senescence
BEAS-2B cells were exposed to CSE or CSE+BYF. p16, p21 and p27 proteins accumulated intracellularly in the CSE group compared to the negative control (NC) group (Figure 1A and B), whereas the protein levels of p16, p21 and p27 were reduced in the CSE+BYF group compared to the CSE group (Figure 1A and B). In addition, the secretion of SASP factors into the medium was measured by ELISA. The levels of IL-6, IL-8 and CCL3 secreted by cells in the CSE group were significantly higher than those in the NC group (Figure 1C–E), whereas the levels of IL-6, IL-8 and CCL3 in the CSE+BYF group were significantly lower than those in the CSE group (Figure 1C–E). These results suggest that BYF treatment inhibits CSE-induced senescence and secretion of SASP factors in BEAS-2B cells.
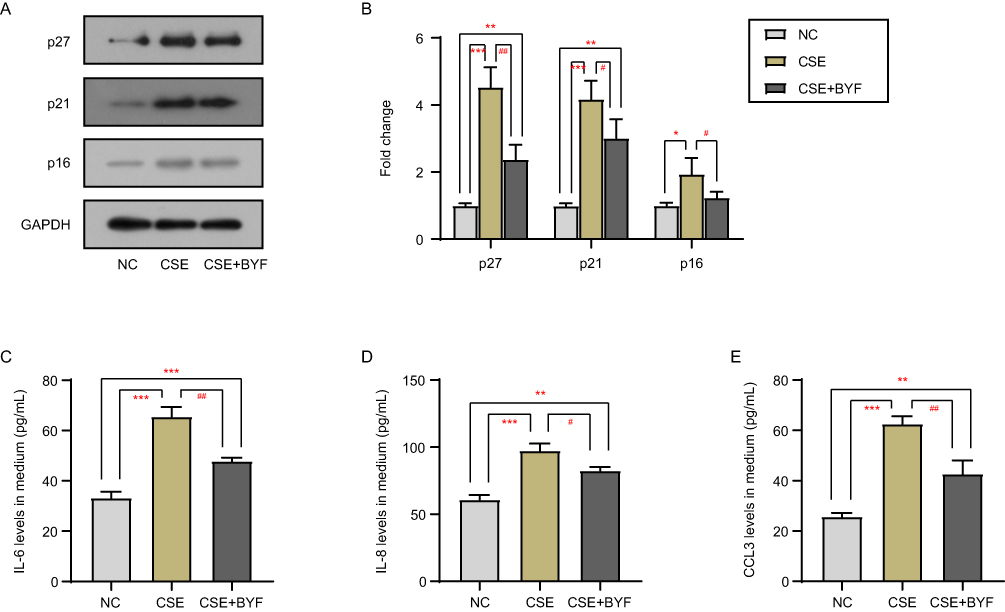 |
Figure 1 BYF inhibits CSE-induced cellular senescence. The protein levels of senescence markers p16, p21 and p27 were detected by Western blot (A and B). The secretion of IL-6 (C), IL-8 (D), and CCL3 (E) into the medium was measured by ELISA. *Compared into NC group, P<0.05; **Compared into NC group, P<0.01; ***Compared into NC group, P<0.001; #Compared into CSE group, P<0.05; ##Compared into CSE group, P<0.01. |
BYF Up-Regulates the Transcription, Expression and Secretion of Klotho
The effects of CSE+BYF treatment on the transcription, expression and secretion of klotho in BEAS-2B cells were examined by RT-qPCR, Western blot and ELISA, respectively. As shown in Figure 2, CSE decreased the mRNA and protein levels of klotho in BEAS-2B cells, as well as the levels of soluble klotho in the medium. Compared with the CSE group, the CSE+BYF group showed significantly increased levels of klotho mRNA and protein, as well as soluble klotho levels (Figure 2). These results indicate that CSE inhibited the transcription, expression and secretion of klotho in BEAS-2B cells, while BYF treatment activated the transcription, expression and secretion of klotho.
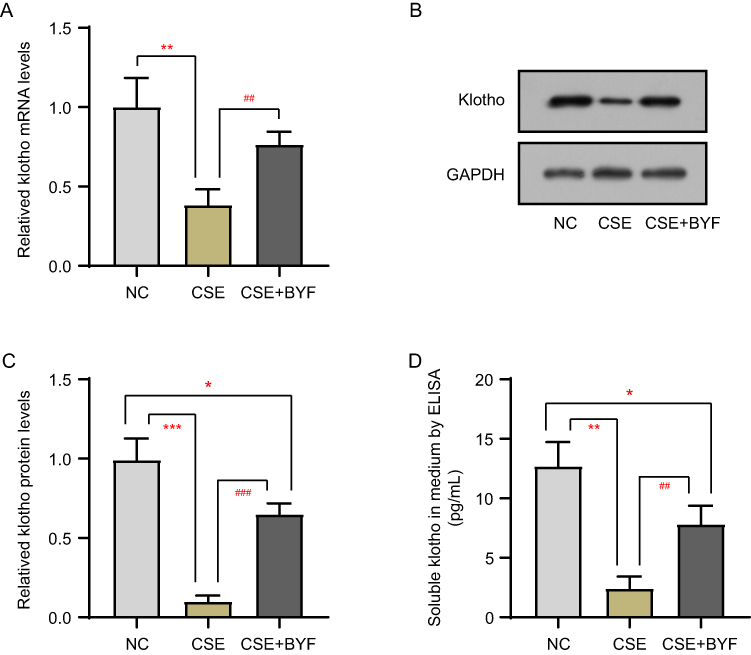 |
Figure 2 BYF up-regulates the transcription, expression and secretion of klotho. The transcription (A), expression (B and C) and secretion (D) of klotho in BEAS-2B cells were detected by RT-qPCR, Western blot and ELISA, respectively. *Compared into NC group, P<0.05; **Compared into NC group, P<0.01; ***Compared into NC group, P<0.001; ##Compared into CSE group, P<0.01; ###Compared into CSE group, P<0.001. |
BYF Activates Klotho Transcription by Upregulating the Expression of Transcription Factor ZNF263
Given the effects of CSE and BYF treatment on klotho transcript levels, we predicted potential transcription factors for klotho using the JASPAR and USCS databases (Figure 3A). We found that CSE downregulated the protein level of ZNF263 in BEAS-2B cells, whereas BYF treatment rescued the ZNF263 expression (Figure 3B and C). Furthermore, ZNF263-overexpressing BEAS-2B cells could inhibit CSE-induced cellular senescence (Figure 3D–F) and SASP factor secretion (Figure 3G–I) by upregulating the klotho expression (Figure 3D and E).
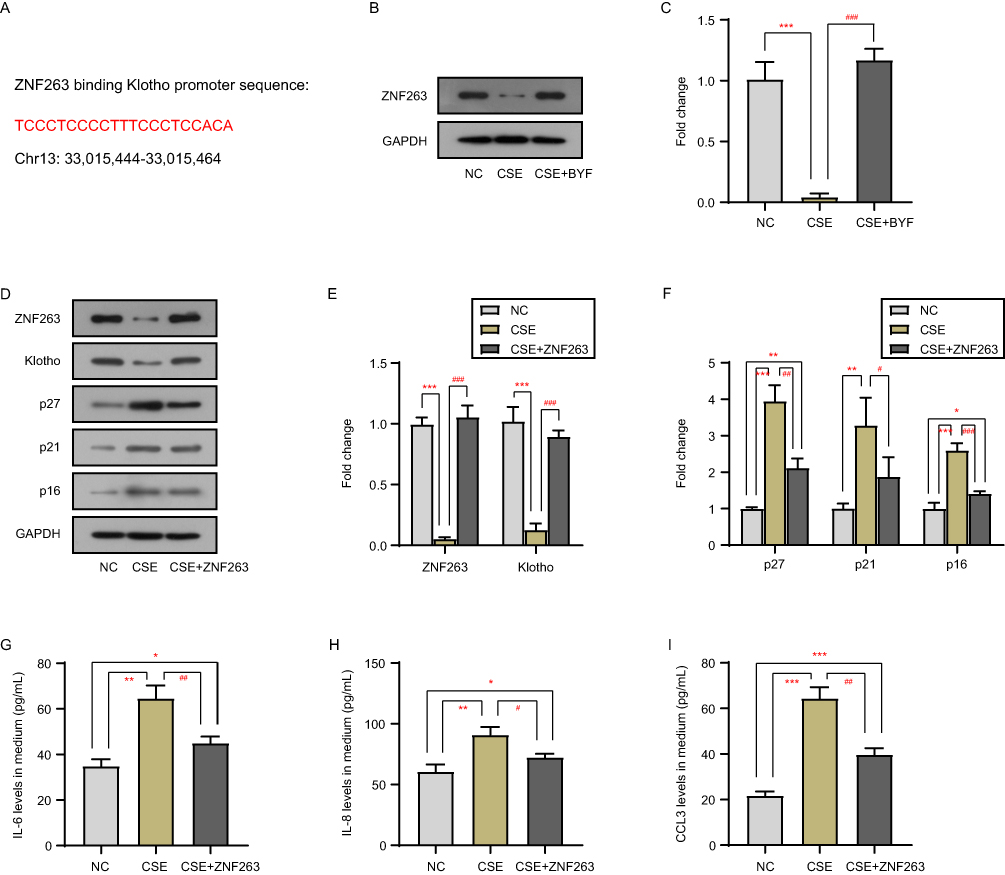 |
Figure 3 BYF activates klotho transcription by upregulating the expression of transcription factor ZNF263. ZNF263 binding klotho promoter sequence with JASPAR and USCS databases (A). The protein level of ZNF263 in BEAS-2B cells with CSE exposure and BYF treatment was detected by Western blot (B and C). The protein levels of ZNF263, klotho and senescence markers in BEAS-2B cells with CSE exposure and ZNF263-expression plasmid transfection were detected by Western blot (D–F). The secretion of IL-6 (G), IL-8 (H), and CCL3 (I) into the medium was measured by ELISA. *Compared into NC group, P<0.05; **Compared into NC group, P<0.01; ***Compared into NC group, P<0.001; #Compared into CSE group, P<0.05; ##Compared into CSE group, P<0.01; ###Compared into CSE group, P<0.001. |
Discussion
In a previous study, we found that CSE suppresses the expression of klotho in BEAS-2B cells, whereas BYF can inhibit CSE-induced inflammation by upregulating the expression of klotho.10 In this study, we found that CSE induced senescence in BEAS-2B cells, whereas BYF could inhibit CSE-induced cellular senescence by upregulating the transcription, expression and secretion of the anti-aging gene klotho. Currently, studies on the mechanism of BYF to alleviate clinical symptoms of COPD patients have focused on the regulation of oxidative stress and immune responses. In this study, we report that BYF can inhibit the cellular senescence of COPD, revealing a new molecular mechanism of by which BYF exerts its function.
Klotho is an important anti-aging gene. Klotho knockout mice exhibit syndromes similar to human aging, including shortened lifespan and early-onset emphysema.14 In contrast, mice with klotho overexpression have significantly longer lifespan. However, there are few studies on the expression regulation mechanisms of klotho. This study identified a novel transcription factor of klotho, ZNF263. ZNF263 activated the transcription of klotho and upregulated its expression. Importantly, we also found that BYF activated the transcription of klotho by upregulating the expression of the transcription factor ZNF263.
ZNF263 is a transcriptional repressor and activator that contains an N-terminal scanning domain and nine C2H2 zinc finger domains at the C-terminus, as well as a kruppel association box (KRAB) domain.15,16 Frietze et al find that the gene with the greatest decrease in expression upon loss of ZNF263 is “cellular component organization and biogenesis”.17 ZNF263 has been reported to be involved in early human embryonic development,18 cardiac regeneration19 and adipogenesis.20 However, these studies are only based on software analysis and lack experimental validation. Our study demonstrates the role of ZNF263 in regulating of cellular senescence. Other target genes regulated by ZNF263 remain to be further investigated.
To our knowledge, several different BYF components exist in some hospitals. In our hospital, the formula includes 6 Chinese herbs including Astragali Radix, Ginseng Radix et Rhizoma, Atractylodes macrocephala, Radix Rehmanniae Preparata, Fructus Corni and Herba Epimedii in a ratio of 1.5:1.5:1.2:1.0:1.0:1.0. The formulation of Chen et al’s team (self-defined as BYF-2 in this study) contains 12 Chinese herbs, minus the use of Atractylodes macrocephala and Radix Rehmanniae Preparata, and adds Lycii Fructus, Schisandrae Chinensis Fructus, Fritillariae Thunbergii Bulbus, Paeoniae Rubra Radix, Pheretima, Perillae Fructus, Ardisiae Japonicae Herba and Citri Reticulatae Pericarpium.21–23 Their findings suggest that BYF-2 exerts beneficial effects on COPD by modulating pathways such as lipid metabolism, inflammatory responses, oxidative stress, and cell connectivity.21–23 The formulation of Li et al’s team (self-defined as BYF-3 in this study) includes only 4 Chinese herbs: Asragali Radix, Ginseng Radix et Rhizoma, Fructus Corni and Schisandrae Chinensis Fructus in a ratio of 1:1:1:0.6.24,25 Their findings suggest that BYF-3 has a therapeutic effect on COPD in rats through its effects on interleukin expression and/or secretion and lipid metabolism.24,25 Other groups have used systems pharmacology to identify the main active components of BYF, including icariin, astragaloside IV, nobiletin, ginsenoside Rh1, paeonol, etc.3,25,26 These studies all show the promise of the formulation in COPD treatment. However, the standardization and unification of formulations, the identification of all effective components, the molecular mechanism and pharmacological effects of the formulations (or effective components), basic and preclinical research, and cohort analysis of clinical applications will all be important contents of future research.
In conclusion, BYF inhibits the cell senescence of COPD by up-regulating the expression of transcription factor ZNF263 and activating the transcription, expression and secretion of the senescence suppressor gene klotho. This study reveals a novel possible molecular mechanism by which BYF alleviates clinical symptoms of COPD patients. In addition, the expression of transcription factor ZNF263 and senescence suppressor gene klotho are suppressed in COPD, and regulating their expression may be beneficial to COPD treatment and prevention.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
This work was supported by Huai’an Science and Technology Bureau (No. HAB202037), the Innovation Service Capacity Building Program of Huai’an (No. HAP202003), Special fund of Huai’an Science and Technology Bureau: Huai’an Key Laboratory of Geriatric Diseases and Geriatric Syndrome (No. HAP202105) and The research project of elderly health in Jiangsu Province (No. LR2021050). Weimin Wang and Shaohong Zhang are co-first authors for this study. Longchuan Wu and Xingxing Xu are co-correspondence authors for this study.
Funding
This work was supported by Huai’an Science and Technology Bureau (No. HAB202037), the Innovation Service Capacity Building Program of Huai’an (No. HAP202003), Special fund of Huai’an Science and Technology Bureau: Huai’an Key Laboratory of Geriatric Diseases and Geriatric Syndrome (No. HAP202105) and The research project of elderly health in Jiangsu Province (No. LR2021050).
Disclosure
The authors declare no competing interests in this work.
References
1. Choi JY, Rhee CK. Diagnosis and treatment of early chronic obstructive lung disease (COPD). J Clin Med. 2020;9:11. doi:10.3390/jcm9113426
2. Li LC, Han YY, Zhang ZH, et al. Chronic obstructive pulmonary disease treatment and pharmacist-led medication management. Drug Des Devel Ther. 2021;15:111–124. doi:10.2147/DDDT.S286315
3. Wu L, Chen Y, Yi J, et al. Mechanism of action of bu-fei-yi-shen formula in treating chronic obstructive pulmonary disease based on network pharmacology analysis and molecular docking validation. Biomed Res Int. 2020;2020:9105972. doi:10.1155/2020/9105972
4. Rutten EP, Gopal P, Wouters EF, et al. Various mechanistic pathways representing the aging process are altered in COPD. Chest. 2016;149(1):53–61. doi:10.1378/chest.15-0645
5. Barnes PJ, Baker J, Donnelly LE. Cellular senescence as a mechanism and target in chronic lung diseases. Am J Respir Crit Care Med. 2019;200(5):556–564. doi:10.1164/rccm.201810-1975TR
6. Birch J, Anderson RK, Correia-Melo C, et al. DNA damage response at telomeres contributes to lung aging and chronic obstructive pulmonary disease. Am J Physiol Lung Cell Mol Physiol. 2015;309(10):L1124–37. doi:10.1152/ajplung.00293.2015
7. Li L, Wang Y, Gao W, et al. Klotho reduction in alveolar macrophages contributes to cigarette smoke extract-induced inflammation in chronic obstructive pulmonary disease. J Biol Chem. 2015;290(46):27890–27900. doi:10.1074/jbc.M115.655431
8. Kureya Y, Kanazawa H, Ijiri N, et al. Down-regulation of soluble alpha-klotho is associated with reduction in serum irisin levels in chronic obstructive pulmonary disease. Lung. 2016;194(3):345–351. doi:10.1007/s00408-016-9870-7
9. Gao W, Yuan C, Zhang J, et al. Klotho expression is reduced in COPD airway epithelial cells: effects on inflammation and oxidant injury. Clin Sci. 2015;129(12):1011–1023. doi:10.1042/CS20150273
10. Wang WM, Wu LC, Chen B, et al. Effects of Bufei Yishen formula on klotho gene expression and inflammation-related RIG-I/NF-κB pathway in cigarette smoke extract-treated BEAS-2B cells. Trop J Pharm Res. 2018;17(10):1925–1931. doi:10.4314/tjpr.v17i10.5
11. Blake DJ, Reese CM, Garcia M, et al. Soluble extracellular Klotho decreases sensitivity to cigarette smoke induced cell death in human lung epithelial cells. Toxicol In Vitro. 2015;29(7):1647–1652. doi:10.1016/j.tiv.2015.06.019
12. Dang X, He B, Ning Q, et al. Alantolactone suppresses inflammation, apoptosis and oxidative stress in cigarette smoke-induced human bronchial epithelial cells through activation of Nrf2/HO-1 and inhibition of the NF-kappaB pathways. Respir Res. 2020;21(1):95. doi:10.1186/s12931-020-01358-4
13. Chen CH, Li YR, Lin SH, et al. Tiotropium/Olodaterol treatment reduces cigarette smoke extract-induced cell death in BEAS-2B bronchial epithelial cells. BMC Pharmacol Toxicol. 2020;21(1):74. doi:10.1186/s40360-020-00451-0
14. Kuro-o M, Matsumura Y, Aizawa H, et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature. 1997;390(6655):45–51. doi:10.1038/36285
15. Yokoyama M, Nakamura M, Okubo K, et al. Isolation of a cDNA encoding a widely expressed novel zinc finger protein with the LeR and KRAB-A domains. Biochim Biophys Acta. 1997;1353(1):13–17. doi:10.1016/S0167-4781(97)00074-2
16. Williams AJ, Blacklow SC, Collins T. The zinc finger-associated SCAN box is a conserved oligomerization domain. Mol Cell Biol. 1999;19(12):8526–8535. doi:10.1128/MCB.19.12.8526
17. Frietze S, Lan X, Jin VX, et al. Genomic targets of the KRAB and SCAN domain-containing zinc finger protein 263. J Biol Chem. 2010;285(2):1393–1403. doi:10.1074/jbc.M109.063032
18. Leng L, Sun J, Huang J, et al. Single-cell transcriptome analysis of uniparental embryos reveals parent-of-origin effects on human preimplantation development. Cell Stem Cell. 2019;25(5):697–712 e6. doi:10.1016/j.stem.2019.09.004
19. Afify AY. A miRNA’s insight into the regenerating heart: a concise descriptive analysis. Heart Fail Rev. 2020;25(6):1047–1061. doi:10.1007/s10741-019-09896-w
20. Khan R, Raza SHA, Junjvlieke Z, et al. Function and transcriptional regulation of bovine TORC2 gene in adipocytes: roles of C/EBP, XBP1, INSM1 and ZNF263. Int J Mol Sci. 2019;20:18. doi:10.3390/ijms20184338
21. Zhao P, Li J, Yang L, et al. Integration of transcriptomics, proteomics, metabolomics and systems pharmacology data to reveal the therapeutic mechanism underlying Chinese herbal Bufei Yishen formula for the treatment of chronic obstructive pulmonary disease. Mol Med Rep. 2018;17(4):5247–5257. doi:10.3892/mmr.2018.8480
22. Zhao P, Liu X, Dong H, et al. Bufei yishen formula restores Th17/treg balance and attenuates chronic obstructive pulmonary disease via activation of the adenosine 2a receptor. Front Pharmacol. 2020;11:1212. doi:10.3389/fphar.2020.01212
23. Guan Q, Zhao P, Tian Y, et al. Identification of cancer risk assessment signature in patients with chronic obstructive pulmonary disease and exploration of the potential key genes. Ann Med. 2022;54(1):2309–2320. doi:10.1080/07853890.2022.2112070
24. Li J, Yang L, Yao Q, et al. Effects and mechanism of bufei yishen formula in a rat chronic obstructive pulmonary disease model. Evid Based Complement Alternat Med. 2014;2014:381976. doi:10.1155/2014/381976
25. Jin F, Zhang L, Chen K, et al. Effective-component compatibility of bufei yishen formula III combined with electroacupuncture suppresses inflammatory response in rats with chronic obstructive pulmonary disease via regulating SIRT1/NF-kappaB signaling. Biomed Res Int. 2022;2022:3360771. doi:10.1155/2022/3360771
26. Li J, Zhao P, Yang L, et al. System biology analysis of long-term effect and mechanism of Bufei Yishen on COPD revealed by system pharmacology and 3-omics profiling. Sci Rep. 2016;6:25492. doi:10.1038/srep25492
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.