")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 12
Brain-derived neurotrophic factor modulates immune reaction in mice with peripheral nerve xenotransplantation
Authors Yu X, Lu L, Liu Z, Yang T, Gong X, Ning Y, Jiang Y
Received 15 October 2015
Accepted for publication 18 November 2015
Published 29 March 2016 Volume 2016:12 Pages 685—694
DOI https://doi.org/10.2147/NDT.S98387
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Wai Kwong Tang
Xin Yu,1 Laijin Lu,1 Zhigang Liu,1 Teng Yang,2 Xu Gong,1 Yubo Ning,3 Yanfang Jiang4
1Department of Hand Surgery, 2Department of Orthopedics, The First Hospital of Jilin University, Changchun, 3Department of Orthopedics, Ningshi Orthopedics Hospital of Tonghua, Tonghua, 4Department of Central Laboratory, The First Hospital of Jilin University, Changchun, People’s Republic of China
Background: Brain-derived neurotrophic factor (BDNF) has been demonstrated to play an important role in survival, differentiation, and neurite outgrowth for many types of neurons. This study was designed to identify the role of BDNF during peripheral nerve xenotransplantation.
Materials and methods: A peripheral nerve xenotransplantation from rats to mice was performed. Intracellular cytokines were stained for natural killer (NK) cells, natural killer T (NKT) cells, T cells, and B cells and analyzed by flow cytometry in the spleen of the recipient mouse. Serum levels of related cytokines were quantified by cytometric bead array.
Results: Splenic NK cells significantly increased in the xenotransplanted mice (8.47±0.88×107 cells/mL) compared to that in the control mice (4.66±0.78×107 cells/mL, P=0.0003), which significantly reduced in the presence of BDNF (4.85±0.87×107 cells/mL, P=0.0004). In contrast, splenic NKT cell number was significantly increased in the mice with xenotransplantation plus BDNF (XT + BDNF) compared to that of control group or of mice receiving xenotransplantation only (XT only). Furthermore, the number of CD3+ T cells, CD3+CD4+ T cells, CD3+CD4- T cells, interferon-γ-producing CD3+CD4+ T cells, and interleukin (IL)-17-producing CD3+CD4+ T cells, as well as CD3-CD19+ B cells, was significantly higher in the spleen of XT only mice compared to the control mice (P<0.05), which was significantly reduced by BDNF (P<0.05). The number of IL-4-producing CD3+CD4+ T cells and CD3+CD4+CD25+Foxp3+ T cells was significantly higher in the spleen of XT + BDNF mice than that in the spleen of XT only mice (P<0.05). Serum levels of IL-6, TNF-α, interferon-γ, and IL-17 were decreased, while IL-4 and IL-10 were stimulated by BDNF following xenotransplantation.
Conclusion: BDNF reduced NK cells but increased NKT cell accumulation in the spleen of xenotransplanted mice. BDNF modulated the number of splenic T cells and its subtype cells in the mice following xenotransplantation. These findings suggest that BDNF inhibits rejection of peripheral nerve following xenotransplantation by regulating innate as well as adaptive immune reaction.
Keywords: peripheral nerve, xenotransplantation, brain-derived neurotrophic factor
Introduction
Neurotrophins are a family of secreted proteins, including brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), neurotrophin-3, and neurotrophin-4, which have been recognized as important regulators of synaptic plasticity.1–4 These molecules have been shown to control the generation, survival, differentiation, and regeneration of neurons in the peripheral and central nervous systems.5 Induction of BDNF in the peripheral nerve and muscle would be important to stimulate Schwann cell differentiation and proliferation, a prerequisite for axonal regeneration. However, neurotrophins are now considered to be growth factors with a wide spectrum of functions outside the nervous system, including modulation and regulation of the immune function.6 NGF has been demonstrated to act as a cytokine-like factor in the development and function of the immune system, and the function of NGF on B cells, T cells, and macrophages has been well described.7 However, limited studies on the role of BDNF in the immune system, especially in transplant immunity, are available.
Two distinct classes of cell surface receptors mediate the biological effects of BDNF, the high affinity receptor tyrosine kinase B (TrkB) and the low affinity p75 neurotrophin receptor. In adult central nervous system, the most prominent expression of BDNF and its receptor, TrkB, is found in structures associated with epilepsy and synaptic plasticity, such as the hippocampus, cerebellum, and neocortex, suggesting that BDNF may modify neuronal plasticity in adult animals. Indeed, accumulating evidence indicates that BDNF is involved in synaptic transmission, neuronal excitability, and the development and maintenance of neuronal plasticity in adult central nervous system. It has been shown that both T and B lymphocytes express TrkB receptors on their surface.8–10 T cells, B cells, and other cells, including innate immune cells, have been demonstrated to play an essential role in the pathogenesis of rejection following organ transplantation. However, the role of BDNF in modulating the function of the immune cells following nerve transplantation has not been investigated.
Therefore, in order to identify whether BDNF could prevent immune rejection after peripheral nerve xenotransplantation and facilitate the nerve regeneration, we performed nerve xenotransplantation from rats to mice and found a dramatic reduction of natural killer (NK) cells, B cells, type 1 T helper (Th1) cells, and type 17 T helper (Th17) cells, while a remarkable increase of natural killer T (NKT) lymphocytes, type 2 T helper (Th2) cells, and regulatory T cells (Tregs) was observed in the spleen of xenotransplanted mice treated with BDNF compared to that in mice receiving xenotransplantation only. In addition, serum concentrations of interleukin (IL)-6, IL-17, interferon γ (IFN-γ), and TNF-α were significantly decreased, while IL-4 and IL-10 were significantly increased in the presence of BDNF following xenotransplantation.
Materials and methods
Animals
Adult male Sprague Dawley rats (6.0±0.1 weeks old and 190±5 g weight in average) and adult male BALB/c mice (70.1±0.2 weeks old and 20.2±0.5 g weight in average) were used as donors and recipients, respectively (at least five mice in each group). All animals were purchased from Jilin University (Changchun, People’s Republic of China). All surgical procedures and postoperative care of the animals were approved by the Institutional Animal Ethics Committee of Jilin University.
Peripheral nerve xenotransplantation
Rats were euthanized followed by exposing the sciatic nerve of the donor rat through an incision splitting the dorsal gluteal muscle. A segment (0.5 cm) of sciatic nerve was then harvested and used immediately. The recipient mouse was given 3.5% chloral hydrate (0.1 mL/kg) intraperitoneally. The skin over the mouse’s right hind limb was incised, and the underneath muscle was bluntly dissected to expose the sciatic nerve. A 0.5 cm gap on the sciatic nerve was created. The rat sciatic nerve graft (0.5 cm) was interposed to the transected nerve and repaired with 11-0 nylon epineurial sutures. Mice in which the sciatic nerve was exposed without xenotransplantation were used as the sham control group. All mice were sacrificed on the third day after transplantation.
BDNF application
BDNF (eBioscience, San Diego, CA, USA) was diluted with physiological saline (500 μg/50 μL), and 10 μL (total 100 μg) was topically applied at the place of the transplanted nerve graft only once during the operation.
Flow cytometric analysis of intracellular cytokine staining
Whole spleen was homogenized in PBS, and the cell suspension was filtered to remove the debris. Cells were collected by centrifugation at 1,500× g for 10 minutes at 4°C. The cells were then washed and resuspended with FACS buffer. After processing, 1–2×106 cells/mL were stimulated with phorbol 12-myristate 13-acetate (PMA; Sigma Chemicals, Perth, Australia) (50 ng/mL) plus 2 mg/mL of ionomycin for 6 hours. Brefeldin A (10 mg/mL) was then added 2 hours before the cells were collected and stained with antibodies. The cells were stained with antibodies against surface markers (antimouse CD3, CD4, CD25, and NK; eBioscience). Cells were then permeabilized with membrane-permeabilizing reagents (Caltag, Burlingame, CA, USA) and fixed using a fixing solution (Caltag), following the manufacturer’s instructions, followed by incubation with anti-IL-4, anti-IFN-γ, anti-IL-17, and anti-Foxp3 (eBioscience), or isotope-matched control antibodies for 30 minutes. Flow cytometry (FACSCalibur™; Beckton Dickinson, Franklin Lakes, NJ, USA) was performed and data were analyzed using FlowJo software.
Cytometric bead array of serum cytokines
Blood was collected from the orbital sinus of the recipient mice. The concentration of serum cytokines (IFN-γ, TNF-α, IL-4, IL-6, IL-10, and IL-17) was determined by cytometric bead array (CBA), following the manufacturer’s protocol (BD Biosciences, San Jose, CA, USA) with minor modifications. Briefly, 25 mL of individual serum was used in duplicate for analysis. The concentration of serum cytokines was quantified using the CellQuestPro and CBA software (Becton Dickinson) on a FACSCalibur cytometry (BD Biosciences).
Statistical analysis
Difference of paired or unpaired groups was determined by the Student’s t-test for parametric data sets. All statistical analyses were performed using Prism 4 (GraphPad Software, Inc., La Jolla, CA, USA). P≤0.05 was considered statistically significant for all analyses in this study.
Results
Effect of BDNF on the number of splenic NK cells in the xenotransplanted mice
All mice survived post-transplantation regardless of BDNF treatment. The total cell number of splenic mononuclear cells was highest in the mice of xenotransplantation without BDNF treatment (XT only) group (2.68±0.32×109 cells/mL), mildly high in the group of transplantation plus BDNF treatment (XT + BDNF, 2.52±0.41×109 cells/mL), and least in the sham control group (2.41±0.38×109 cells/mL). However, there was no statistical difference among the three groups. The number of splenic NK cells was further counted and characterized, and it was found to be significantly increased in the mice of XT only group (8.47±0.88×107 cells/mL) than that in the sham control group (4.68±0.78×107 cells/mL, P=0.0003, Figure 1A). In contrast, the number of splenic NK cells in the mice of XT + BDNF treatment group (4.85±0.87×107 cells/mL) was remarkably reduced, and it was significantly less than that in the group of XT only group (8.47±0.88×107 cells/mL, P=0.0004). In addition, there was no significant difference between the XT + BDNF group and the sham control group (Figure 1B, P=0.9728).
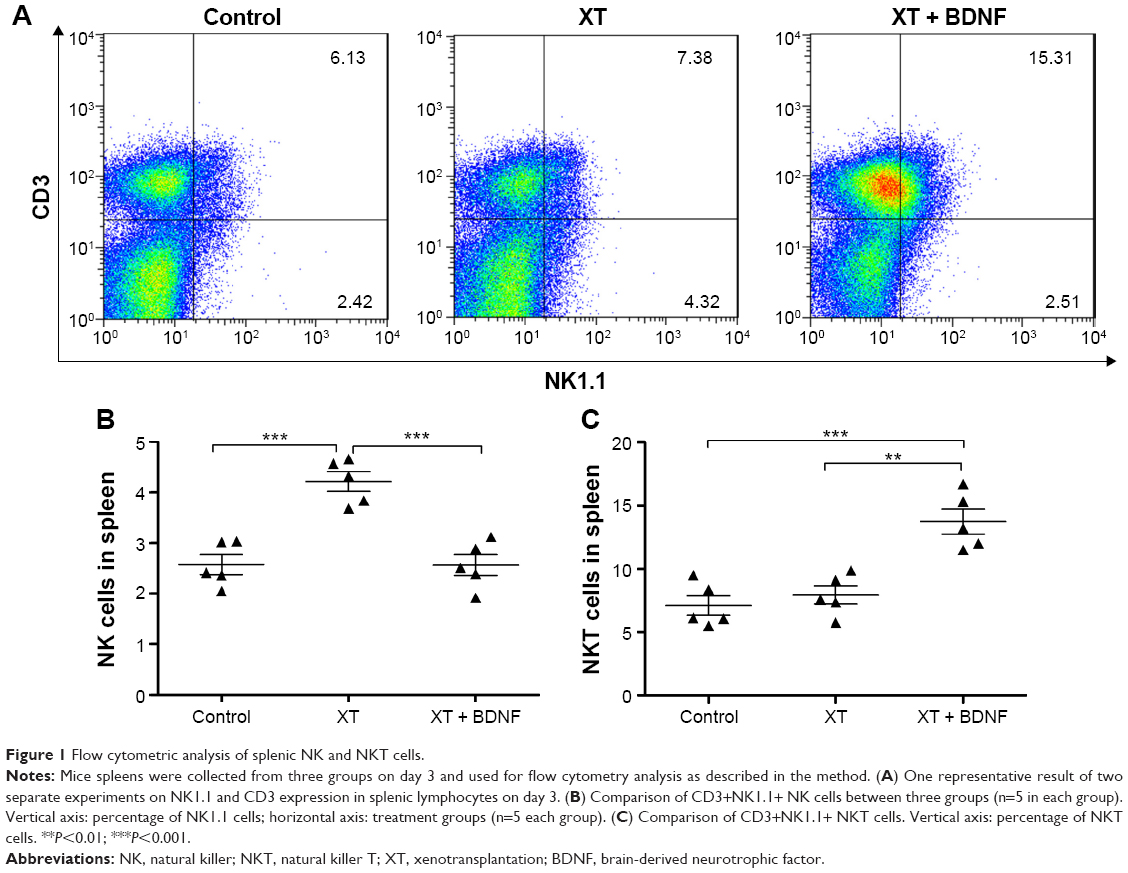 | Figure 1 Flow cytometric analysis of splenic NK and NKT cells. |
Effect of BDNF on the number of splenic NKT cells in the xenotransplanted mice
The number of NKT cells was significantly increased in the spleen of mice receiving XT + BDNF (2.60±0.42×108 cells/mL) compared to that in the sham control group (1.29±0.31×108 cells/mL, P=0.0007) or the XT only mice (1.60±0.32×108 cells/mL, P=0.0015, Figure 1C). However, there was no significant difference in the number of NKT cells between the XT only group and the sham control mice (P=0.4551).
Effect of BDNF on the number of splenic T cells in the xenotransplanted mice
As shown in Figures 2 and 3, the percentage of T cell and its subgroup was analyzed in the splenic mononuclear cells. The number of CD3+ T cells (12.56±0.94×108 cells/mL), CD3+CD4+ T cells (6.56±0.74×108 cells/mL), CD3+CD4− T cells (3.15±0.39×108 cells/mL), IFN-γ-producing CD3+CD4+ T cells (7.51±0.96×107 cells/mL), and IL-17-producing CD3+CD4+ T cells (3.18±0.20×107 cells/mL) was significantly increased in the spleen of XT only mice compared to that in the sham control group (CD3+ T cells: 6.0±0.54×108 cells/mL, CD3+CD4+ T cells: 3.46±0.42×108 cells/mL, CD3+CD4− T cells: 1.55±0.16×108 cells/mL, IFN-γ-producing CD3+CD4+ T cells: 1.76±0.09×107 cells/mL, and IL-17-producing CD3+CD4+ T cells: 0.51±0.09×107 cells/mL, respectively, P<0.0001 in all paired comparison; Figure 3A–D, and F, respectively). However, in the presence of BDNF, the number of the aforementioned sub-group T cells was significantly reduced in the mice of XT + BDNF group (CD3+ T cells: 8.55±0.43×108 cells/mL, CD3+CD4+ T cells: 4.36±0.33×108 cells/mL, CD3+CD4− T cells: 2.16±0.18×108 cells/mL, IFN-γ-producing CD3+CD4+ T cells: 3.33±0.30×107 cells/mL, and IL-17-producing CD3+CD4+ T cells: 1.21±0.15×107 cells/mL, respectively) compared to that in the group of XT only (P<0.01 in all paired comparison, Figure 3A–D, and F). In addition, the IL-4-producing CD3+CD4+ T cells and CD3+CD4+CD25+Foxp3+ T cells were significantly increased in the mice of XT + BDNF group (3.36±0.54×107 cells/mL and 6.85±0.34×106 cells/mL, respectively) compared to those in the mice of XT only group (1.84±0.30×107 cells/mL and 3.58±0.36×106 cells/mL, respectively, P<0.05, Figure 3E and G).
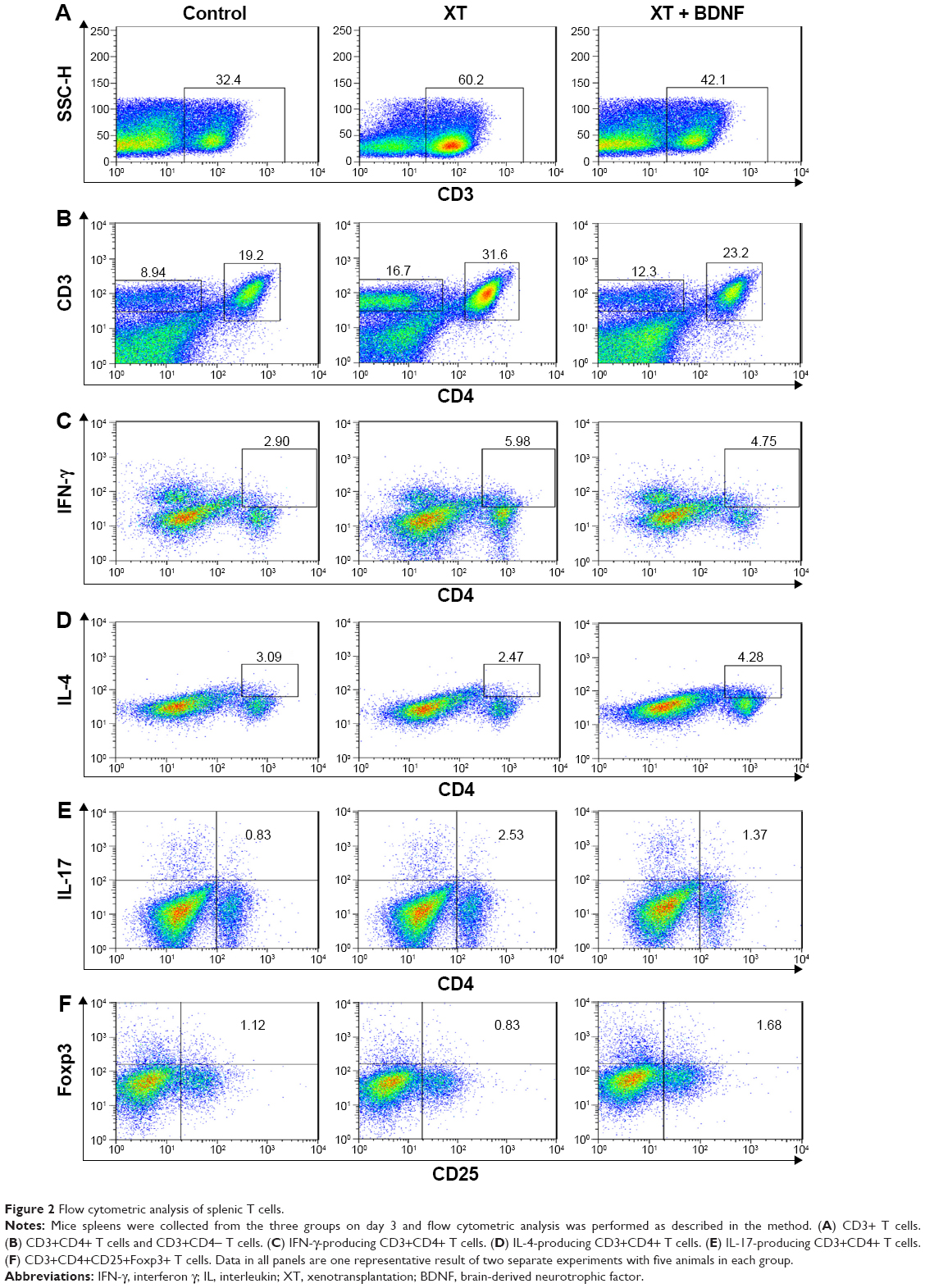 | Figure 2 Flow cytometric analysis of splenic T cells. |
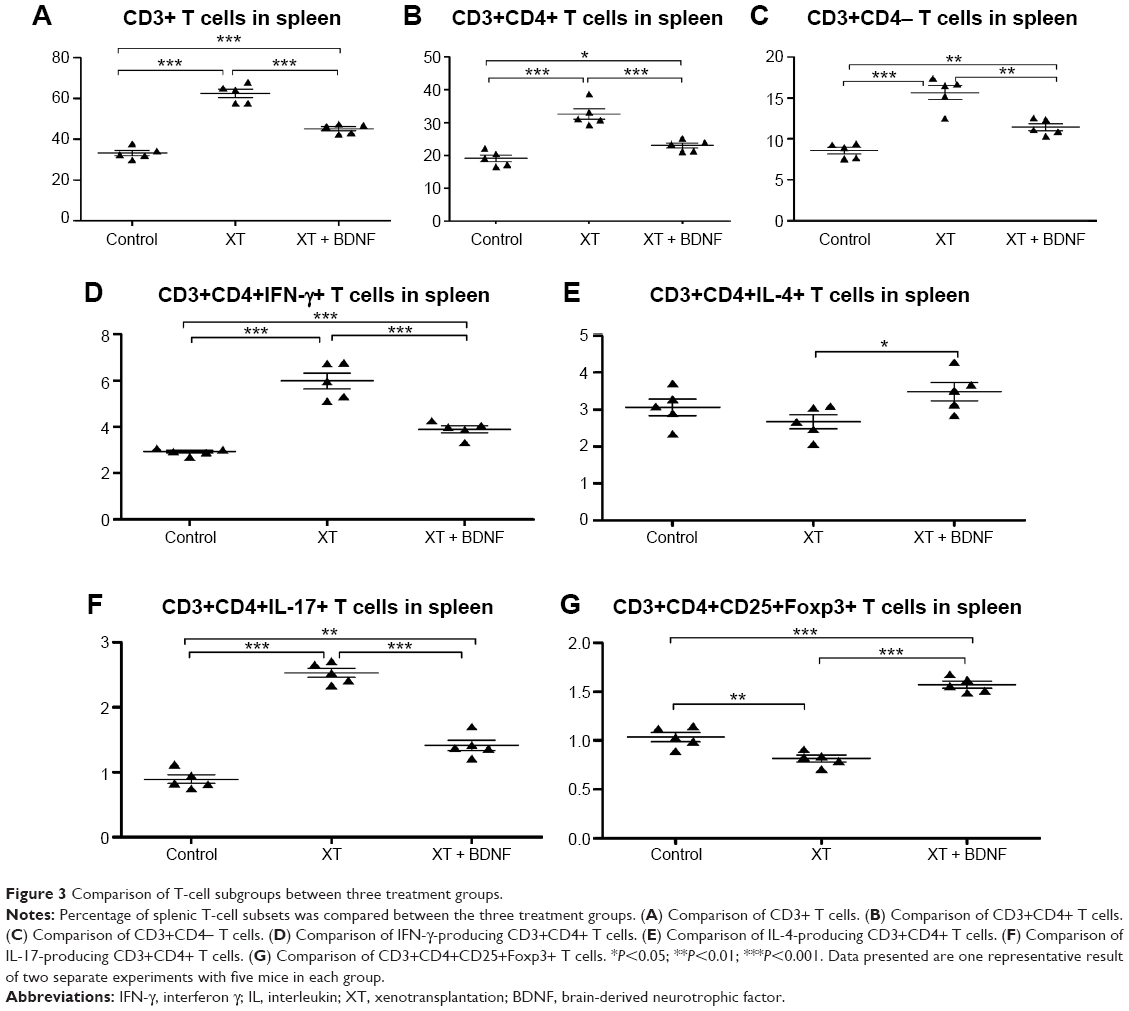 | Figure 3 Comparison of T-cell subgroups between three treatment groups. |
Effect of BDNF on the number of splenic B cells in the xenotransplanted mice
The number of CD3+CD19+ B cells was significantly decreased in the spleen of XT + BDNF group (6.01±0.52×108 cells/mL) compared to that in the sham control group (7.47±0.31×108 cells/mL, P<0.0001) or the XT only group (10.13±0.34×108 cells/mL, P<0.0001, Figure 4A and B). There was no significant difference in the number of CD3+CD19+ B cells between the XT only group and the sham control mice (P=0.1128).
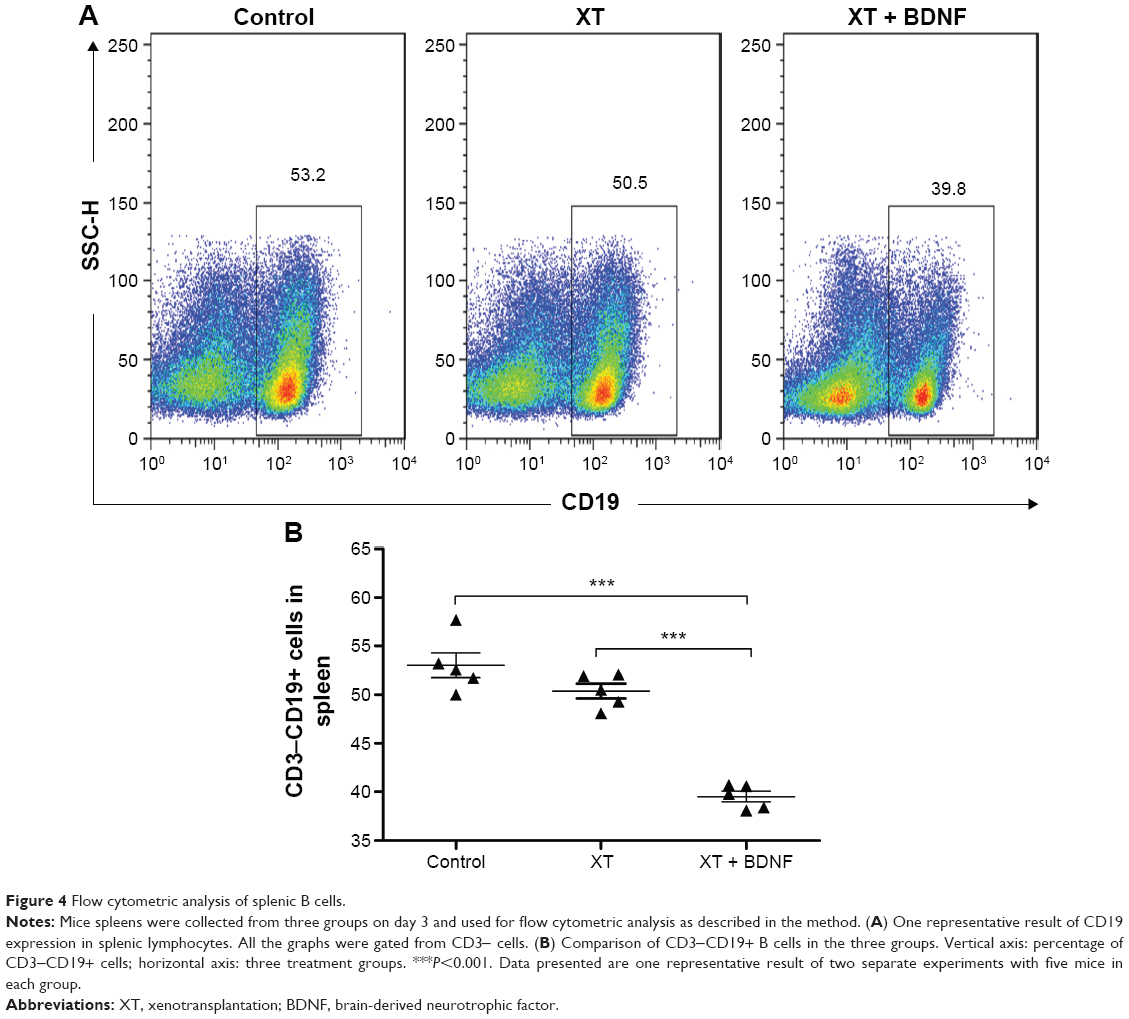 | Figure 4 Flow cytometric analysis of splenic B cells. |
Effect of BDNF on the levels of cytokines in the serum of xenotransplanted mice
Compared to the sham control group, the following cytokines were significantly increased in the serum of XT only mice: IL-6 (51.49±17.19 pg/mL of control vs 152.71±14.18 pg/mL of XT only, P<0.0001), IL-17 (5.76 ±1.52 pg/mL of control vs 38.93±5.04 pg/mL of XT only, P<0.0001), IFN-γ (4.12±0.57 pg/mL of control vs 23.49±5.18 pg/mL of XT only, P<0.0001), and TNF-α (7.15±1.85 pg/mL of control vs 34.74±3.05 pg/mL of XT only, P<0.0001). In contrast, IL-4 and IL-10 were significantly decreased in the serum of XT only mice (1.89±0.66 pg/mL and 45.66±18.59 pg/mL, respectively) compared to those in the sham control mice (3.39±0.68 pg/mL, P=0.0076, and 43.27±28.37 pg/mL, P=0.0002, respectively).
Compared to XT only mice and XT + BDNF mice, the levels of IL-6 (152.71±14.18 pg/mL of XT only vs 84.78±7.84 pg/mL of XT + BDNF, P<0.0001), IL-17 (38.93±5.04 pg/mL of XT only vs 24.16±1.44 pg/mL of XT + BDNF, P=0.0002), IFN-γ (23.49±5.18 pg/mL of XT only vs 15.34±3.11 pg/mL of XT + BDNF, P=0.0167), and TNF-α (34.74±3.50 pg/mL of XT only vs 18.55±6.66 pg/mL of XT + BDNF, P=0.0013) were significantly decreased, while IL-4 (1.89±0.66 pg/mL of XT only vs 24.73±2.13 pg/mL of XT + BDNF, P<0.0001) and IL-10 (45.66±18.59 pg/mL of XT only vs 375.06±57.11 pg/mL of XT + BDNF, P<0.0001) were significantly increased (Figure 5).
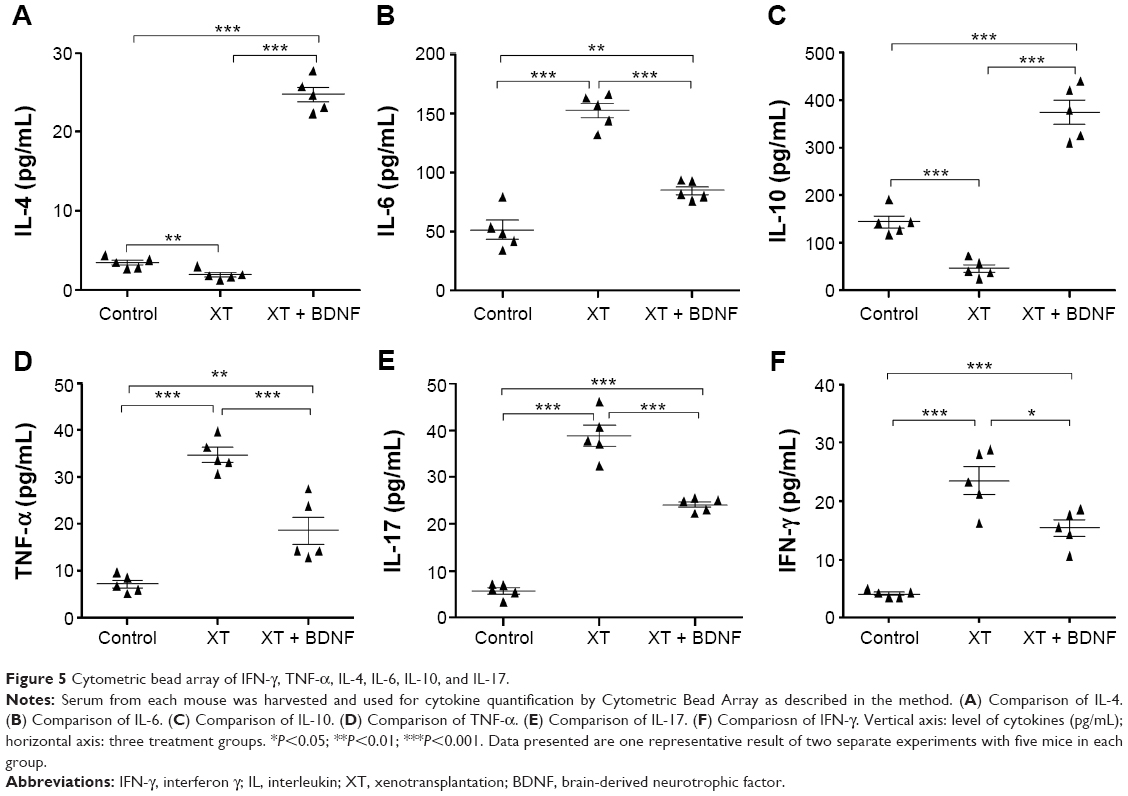 | Figure 5 Cytometric bead array of IFN-γ, TNF-α, IL-4, IL-6, IL-10, and IL-17. |
Discussion
This study investigated the effect of BDNF on the immune reaction following peripheral nerve xenotransplantation from rats to mice. It was found that the number of NK cells, T cells, CD3+CD4+ T cells, CD3+CD4− T cells, Th1 cells, Th17 cells, and B cells was significantly increased in the mice receiving xenotransplantation and that BDNF could significantly suppress upregulation of the aforementioned T cells and B cells in the xenotransplanted mice. Paradoxically, the number of NKT cells, Th2 cells, and Tregs was significantly decreased at the acute phase of peripheral nerve xenotransplantation in the mice, and BDNF could significantly upregulate NKT cells, Th2 cells, and Tregs in the mice receiving nerve xenotransplantation. Furthermore, serum levels of IL-6, IL-17, IFN-γ, and TNF-α were significantly inhibited, whereas serum levels of IL-4 and IL-10 were significantly stimulated by BDNF in the xenotransplanted mice.
As a member of the neurotrophin family, BDNF was initially identified as a target-derived factor that promotes the survival of several populations of central neurons.11–14 Both in vitro15 and in vivo,16–18 it has been reported that application of BDNF was effective for motoneuron survival. Induction of BDNF in the peripheral nerve and muscle would be important to stimulate Schwann cell differentiation and proliferation, a prerequisite for axonal regeneration. It has been reported that BDNF mRNA and its TrkB receptor mRNA coexist in the same neuron,19,20 and antisense BDNF oligonucleotides21,22 or anti-BDNF antiserum23,24 could reduce the survival of cultured neurons, even as isolated single cells. Thus, BDNF may act as an autocrine factor for maintaining neuronal survival during target-independent stages of development.25 It has been reported that BDNF served as a self-amplifying autocrine factor in promoting axon formation in embryonic hippocampal neurons by triggering two-nested positive feedback mechanisms. First, BDNF elevates cytoplasmic cyclic adenosine monophosphate and protein kinase A activity, which triggers further secretion of BDNF and membrane insertion of its receptor TrkB. Second, BDNF/TrkB signaling activates PI3-kinase that promotes anterograde transport of TrkB in the putative axon, further enhancing local BDNF/TrkB signaling. Together, these self-amplifying BDNF actions ensure stable elevation of local cyclic adenosine monophosphate/protein kinase A activity that is critical for axon differentiation and growth.
NK cells are large granular cytotoxic lymphocytes that represent a fundamental component of the innate immune system. In transplantation, NK cells are perhaps best known for their ability to directly reject major histocompatibility complex-mismatched bone marrow allografts, as illustrated by recent studies.26,27 NK cells have the capacity to exacerbate multiple forms of allograft injury; in this regard, NK cells can induce cardiac, skin, and kidney allograft rejection.28–33 In the current study, the number of NK cells after 3 days of peripheral nerve xenotransplantation was significantly higher than that in the sham control mice, indicating that NK cells may induce the acute rejection of peripheral nerve xenotransplantation. However, in the presence of BDNF, the number of NK cells in the xenotransplanted mice was significantly low, suggesting that BDNF inhibits the rejection of peripheral nerve xenotransplantation by decreasing the level of NK cells.
NKT cells are innate-like T cells that recognize lipid antigens presented by the nonpolymorphic major histocompatibility complex class I-like molecule CD1d. It has been demonstrated that NKT cells are required for the induction of tolerance of islet xenografts and cardiac allografts.34,35 In the current study, there was no significant difference in the number of NKT cells between the XT only mice and the sham control group, but it increased significantly in the mice of XT plus BDNF group, suggesting that BDNF enhances the effect of NKT cells in inducing peripheral nerve xenograft tolerance by increasing the number of NKT cells.
Following the activation of innate immune cells, such as NK and NKT cells, adaptive immune cells, including T cells and B cells, are activated during the acute rejection of peripheral nerve xenotransplantation. In this context, it has been demonstrated that cellular immunity, especially T cells, plays a vital role during organ xenotransplant rejection.36–39 In addition to direct killing activity by cytotoxic T lymphocytes, xenograft rejection can occur through T cell-mediated mechanisms, including cytokine production, recruitment and activation of other cytotoxic cells, and by providing help for B cells that produce xenoreactive antibodies.40 We demonstrated that both IFN-γ- and IL-17-producing CD4+ and CD8+ T cells play a crucial role during the acute rejection of peripheral nerve xenotransplantation.41 We further showed that the number of CD3+ T cells, Th1 cells, and Th17 cells was suppressed by BDNF, while the subsets of Th2 cells and Tregs were upregulated by BDNF in the xenotransplanted mice. In addition, serum concentrations of IL-6, IL-17, TNF-α, and IFN-γ were significantly inhibited by BDNF in the xenotransplanted mice. These results indicate that topical application of BDNF after peripheral nerve xenotransplantation results in the inhibition of Th1 and Th17 cells, but activation of Th2 cells and Tregs, and by using this mechanism, BDNF may inhibit the rejection of xenotransplanted peripheral nerve segment.
Besides T cell and its subgroups, B cells also play a crucial role in transplant rejection such as in kidney and liver transplantation by secreting target-specific antibodies. In the present study, B cells were also significantly suppressed by BDNF in the xenotransplanted mice, indicating that BDNF regulates target-specific antibodies by suppressing B cells.
In contrast to organ transplantation such as the kidney, heart, or liver transplantation, in which survival of the transplanted organs is crucial to have long-term organ function, the peripheral nerve xenograft functions for a limited time only. Thus, once the regenerative nerve can pass the nerve xenografts in a short period following the transplantation, the function of peripheral nerve xenografts is completed, regardless of the survival or death of the transplanted nerve. The current study demonstrated that BDNF plays an essential role in inhibiting the acute rejection of peripheral nerve xenotransplantation by modulating the immune reaction in the xenotranplanted nerves.
Conclusion
In summary, the current study demonstrated that the number of NK cells, Th1 cells, Th17 cells, and B cells in the xenotransplated nerves was significantly reduced by BDNF, while the number of NKT, Th2 cells, and Tregs in the xenotransplanted spleen was significantly increased by BDNF. In addition, cytokines released by the aforementioned immune cells were also modulated by BDNF. These findings suggest that BDNF inhibits the peripheral nerve xenograft rejection by regulating the paradigm of NK/NKT and Th1/Th2/Th17/Tregs.
Acknowledgments
This study was supported by the grants from the National Natural Science Foundation of China (Nos 81171145 and 81201382) and Post-doctor Science Foundation of China (No 2013M541315).
Disclosure
The authors report no conflicts of interest in this work.
References
Lu B. BDNF and activity-dependent synaptic modulation. Learn Mem. 2003;10(2):86–98. | ||
Mendell LM, Munson JB, Arvanian VL. Neurotrophins and synaptic plasticity in the mammalian spinal cord. J Physiol. 2001;533(pt 1):91–97. | ||
Schinder AF, Poo M. The neurotrophin hypothesis for synaptic plasticity. Trends Neurosci. 2000;23(12):639–645. | ||
Lewin GR, Barde YA. Physiology of the neurotrophins. Annu Rev Neurosci. 1996;19:289–317. | ||
Barde YA. Trophic factors and neuronal survival. Neuron. 1989;2(6):1525–1534. | ||
Nockher WA, Renz H. Neurotrophins in inflammatory lung diseases: modulators of cell differentiation and neuroimmune interactions. Cytokine Growth Factor Rev. 2003;14(6):559–578. | ||
Nassenstein C, Braun A, Erpenbeck VJ, et al. The neurotrophins nerve growth factor, brain-derived neurotrophic factor, neurotrophin-3, and neurotrophin-4 are survival and activation factors for eosinophils in patients with allergic bronchial asthma. J Exp Med. 2003;198(3):455–467. | ||
Besser M, Wank R. Cutting edge: clonally restricted production of the neurotrophins brain-derived neurotrophic factor and neurotrophin-3 mRNA by human immune cells and Th1/Th2-polarized expression of their receptors. J Immunol. 1999;162(11):6303–6306. | ||
Maroder M, Bellavia D, Vacca A, et al. The thymus at the crossroad of neuroimmune interactions. Ann N Y Acad Sci. 2000;917:741–747. | ||
D’Onofrio M, de Grazia U, Morrone S, et al. Expression of neurotrophin receptors in normal and malignant B lymphocytes. Eur Cytokine Netw. 2000;11(2):283–291. | ||
Barde YA, Edgar D, Thoenen H. Purification of a new neurotrophic factor from mammalian brain. EMBO J. 1982;1(5):549–553. | ||
Huang EJ, Reichardt LF. Neurotrophins: roles in neuronal development and function. Annu Rev Neurosci. 2001;24:677–736. | ||
Huang EJ, Reichardt LF. Trk receptors: roles in neuronal signal transduction. Annu Rev Biochem. 2003;72:609–642. | ||
Thoenen H. Neurotrophins and neuronal plasticity. Science. 1995;270(5236):593–598. | ||
Henderson CE, Camu W, Mettling C, et al. Neurotrophins promote motor neuron survival and are present in embryonic limb bud. Nature. 1993;363(6426):266–270. | ||
Friedman B, Kleinfeld D, Ip NY, et al. BDNF and NT-4/5 exert neurotrophic influences on injured adult spinal motor neurons. J Neurosci. 1995;15(2):1044–1056. | ||
Munson JB, Shelton DL, McMahon SB. Adult mammalian sensory and motor neurons: roles of endogenous neurotrophins and rescue by exogenous neurotrophins after axotomy. J Neurosci. 1997;17(1):470–476. | ||
Yan Q, Matheson C, Lopez OT, et al. The biological responses of axotomized adult motoneurons to brain-derived neurotrophic factor. J Neurosci. 1994;14(9):5281–5291. | ||
Miranda RC, Sohrabji F, Toran-Allerand CD. Neuronal colocalization of mRNAs for neurotrophins and their receptors in the developing central nervous system suggests a potential for autocrine interactions. Proc Natl Acad Sci U S A. 1993;90(14):6439–6443. | ||
Kokaia Z, Bengzon J, Metsis M, et al. Coexpression of neurotrophins and their receptors in neurons of the central nervous system. Proc Natl Acad Sci U S A. 1993;90(14):6711–6715. | ||
Wright EM, Vogel KS, Davies AM. Neurotrophic factors promote the maturation of developing sensory neurons before they become dependent on these factors for survival. Neuron. 1992;9(1):139–150. | ||
Acheson A, Conover JC, Fandl JP, et al. A BDNF autocrine loop in adult sensory neurons prevents cell death. Nature. 1995;374(6521):450–453. | ||
Hyman C, Hofer M, Barde YA, et al. BDNF is a neurotrophic factor for dopaminergic neurons of the substantia nigra. Nature. 1991;350(6315):230–232. | ||
Ghosh A, Carnahan J, Greenberg ME. Requirement for BDNF in activity-dependent survival of cortical neurons. Science. 1994;263(5153):1618–1623. | ||
Davies AM, Wright EM. Neurotrophic factors. Neurotrophin autocrine loops. Curr Biol. 1995;5(7):723–726. | ||
Kean LS, Hamby K, Koehn B, et al. NK cells mediate costimulation blockade-resistant rejection of allogeneic stem cells during nonmyeloablative transplantation. Am J Transplant. 2006;6(2):292–304. | ||
Hamby K, Trexler A, Pearson TC, et al. NK cells rapidly reject allogeneic bone marrow in the spleen through a perforin- and Ly49D-dependent, but NKG2D-independent mechanism. Am J Transplant. 2007;7(8):1884–1896. | ||
Maier S, Tertilt C, Chambron N, et al. Inhibition of natural killer cells results in acceptance of cardiac allografts in CD28-/- mice. Nat Med. 2001;7(5):557–562. | ||
McNerney ME, Lee KM, Zhou P, et al. Role of natural killer cell subsets in cardiac allograft rejection. Am J Transplant. 2006;6(3):505–513. | ||
Kroemer A, Xiao X, Degauque N, et al. The innate NK cells, allograft rejection, and a key role for IL-15. J Immunol. 2008;180(12):7818–7826. | ||
Ito A, Shimura H, Nitahara A, et al. NK cells contribute to the skin graft rejection promoted by CD4+ T cells activated through the indirect allorecognition pathway. Int Immunol. 2008;20(10):1343–1349. | ||
Garrod KR, Liu FC, Forrest LE, et al. NK cell patrolling and elimination of donor-derived dendritic cells favor indirect alloreactivity. J Immunol. 2010;184(5):2329–2336. | ||
Hidalgo LG, Sis B, Sellares J, et al. NK cell transcripts and NK cells in kidney biopsies from patients with donor-specific antibodies: evidence for NK cell involvement in antibody-mediated rejection. Am J Transplant. 2010;10(8):1812–1822. | ||
Ikehara Y, Yasunami Y, Kodama S, et al. CD4(+) Valpha14 natural killer T cells are essential for acceptance of rat islet xenografts in mice. J Clin Invest. 2000;105(12):1761–1767. | ||
Seino KI, Fukao K, Muramoto K, et al. Requirement for natural killer T (NKT) cells in the induction of allograft tolerance. Proc Natl Acad Sci U S A. 2001;98(5):2577–2581. | ||
Dorling A, Lombardi G, Binns R, et al. Detection of primary direct and indirect human anti-porcine T cell responses using a porcine dendritic cell population. Eur J Immunol. 1996;26(6):1378–1387. | ||
Davila E, Byrne GW, LaBreche PT, et al. T-cell responses during pig-to-primate xenotransplantation. Xenotransplantation. 2006;13(1):31–40. | ||
Cardona K, Korbutt GS, Milas Z, et al. Long-term survival of neonatal porcine islets in nonhuman primates by targeting costimulation pathways. Nat Med. 2006;12(3):304–306. | ||
Hering BJ, Wijkstrom M, Graham ML, et al. Prolonged diabetes reversal after intraportal xenotransplantation of wild-type porcine islets in immunosuppressed nonhuman primates. Nat Med. 2006;12(3):301–303. | ||
Yang YG, Sykes M. Xenotransplantation: current status and a perspective on the future. Nat Rev Immunol. 2007;7(7):519–531. | ||
Yu X, Jiang Y, Lu L, et al. A crucial role of IL-17 and IFN-gamma during acute rejection of peripheral nerve xenotransplantation in mice. PLoS One. 2012;7(3):e34419. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.