")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 17
Association of Two Variable Number of Tandem Repeats in the Monoamine Oxidase A Gene Promoter with Schizophrenia
Authors Tanifuji T, Okazaki S, Otsuka I, Horai T, Shinko Y, Kim S, Sora I, Hishimoto A
Received 12 September 2021
Accepted for publication 28 October 2021
Published 11 November 2021 Volume 2021:17 Pages 3315—3323
DOI https://doi.org/10.2147/NDT.S338854
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Taro Kishi
Takaki Tanifuji,1 Satoshi Okazaki,1 Ikuo Otsuka,1 Tadasu Horai,1 Yutaka Shinko,1 Saehyeon Kim,1 Ichiro Sora,1 Akitoyo Hishimoto1,2
1Department of Psychiatry, Kobe University Graduate School of Medicine, Kobe, Japan; 2Department of Psychiatry, Yokohama City University Graduate School of Medicine, Yokohama, Japan
Correspondence: Akitoyo Hishimoto
Department of Psychiatry, Yokohama City University Graduate School of Medicine, 3-9 Fukuura, Kanazawa, Yokohama, 236-0004, Japan
Tel +81-45-787-2667
Fax +81-45-783-2540
Email [email protected]
Background: Monoamine oxidase-A (MAO-A) decomposes dopamine and serotonin, and decreased MAO-A expression increases monoamine levels and is related to the pathophysiology of schizophrenia. Previous studies have reported that variable number of tandem repeats (VNTR), namely, upstream (u)VNTR, and some single nucleotide polymorphisms (SNPs) in the MAOA gene are associated with schizophrenia.
Methods: We investigated the two VNTRs and their related SNPs (rs6323 and rs1137070) in the MAOA gene promoter in 859 patients with schizophrenia and 826 healthy controls. Distal (d)VNTR and uVNTR were genotyped with fluorescence-based fragment polymerase chain reaction assays, and rs6323 and rs1137070 with TaqMan SNP genotyping assays.
Results: Neither the genotype nor allelic frequency of the VNTRs or SNPs showed significant differences between the schizophrenia and control groups. On the other hand, analysis of the dVNTR-uVNTR-rs6323-rs1137070 haplotype showed significant association for nine repeats (9R)-3R-T-C in female patients (corrected p = 0.0006, odds ratio [confidence interval] = 2.17 [1.446– 3.257]).
Conclusion: Our findings provide novel evidence that MAOA gene polymorphisms are associated with an increased risk of developing schizophrenia in females.
Keywords: haplotype, monoamine oxidase A, polymorphism, schizophrenia, variable number of tandem repeats
Introduction
Schizophrenia is a severe psychiatric disorder that affects approximately 1% of the global population.1 Schizophrenia has high heritability and is associated with complex poly genic factors.2 Pharmacological studies have indicated that a dysfunction of dopaminergic neurons could contribute to the development of schizophrenia.1 Dopamine degradation is catalyzed by monoamine oxidase (MAO) and catechol-o-methyltransferase (COMT) in the brain.3,4 Many studies have investigated the association of MAO and COMT with schizophrenia.4–7
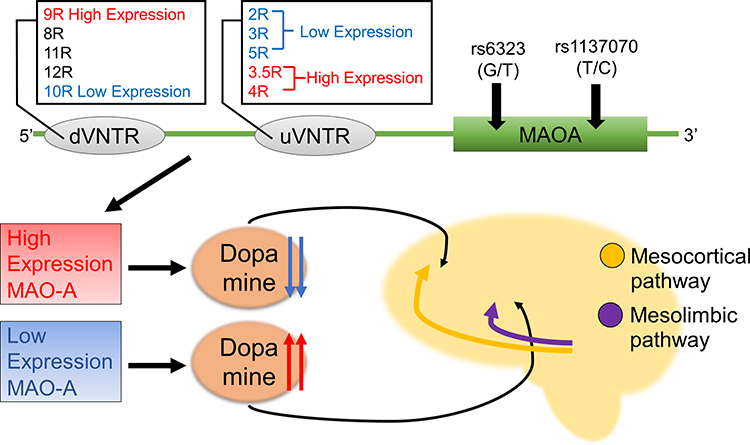 |
Figure 1 Two MAOA gene promoter VNTRs, and their related SNPs have combined effects on the pathogenesis of schizophrenia. For uVNTR, 3.5R and 4R are high-expression alleles, and the 2R, 3R, and 5R are low-expression alleles. For dVNTR, 9R and 10R are high-expression alleles, 8R and 11R are moderate expression alleles, and 12R is unknown. Low levels of MAO-A contribute to increased dopamine levels in the mesolimbic pathway resulting in positive symptoms, and high levels of MAO-A contribute to decreased dopamine levels in the mesocortical pathway cortex leading to negative symptoms with the combined effects of two VNTRs and two SNPs. |
There are two types of MAO, MAO-A and MAO-B, both of which contribute to the degradation of dopamine.3 MAO-A has primary and minor isoforms; however, the functional differences between these isoforms remain unknown.8 The MAOA and MAOB genes are located adjacently to each other on the X chromosome, in the opposite direction.3 MAO-A has been reported to play an important role in mental illnesses such as schizophrenia.9,10 There is an upstream (u) variable number of tandem repeats (VNTR) in the MAOA gene promoter. uVNTR is located 1.2 kb upstream of the MAOA gene, and is present in 3 repeats (R), 3.5R, 4R, or 5R. The 3.5R and 4R are high-expression alleles, and the 2R, 3R, and 5R are low-expression alleles.11 Several studies have reported that the uVNTR and its related single nucleotide polymorphisms (SNPs) of the MAOA gene are associated with schizophrenia;12–15 however, these results are inconsistent with each other.16–18
Recently, a novel VNTR, namely distal (d)VNTR has been identified in the MAOA gene promoter region. dVNTR is located approximately 500 bp upstream of uVNTR and present in 8R, 9R, 10R, 11R or 12R. dVNTR from 8R to 11R were found to be associated with uVNTR, and the corresponding transcripts were evaluated. 9R and 10R are associated with the highest and lowest levels of transcription, respectively, whereas 8R and 11 R show a moderate level of transcription.19 In addition, it was demonstrated that dVNTR and uVNTR are involved in the expression of the two MAO-A isoforms, wherein dVNTR increases the expression of the primary isoform that had little connection to uVNTR, and both VNTRs reduce the expression of a minor isoform that comprised a fraction of the total.8
In neuropsychiatric disorders, the combination of dVNTR and uVNTR was reported to be associated with nicotine dependence.20 However, there is no study that explored the association between the two VNTRs and other mental illnesses such as schizophrenia. In this study, we investigated the association of the two MAOA gene promoter VNTRs, and their related SNPs, with schizophrenia.
Materials and Methods
Participants
We recruited 859 patients with schizophrenia and 826 healthy controls of Japanese descent from the city of Kobe in Japan. The demographic and clinical characteristics of the participants are given in Table 1. At least two psychiatrists diagnosed every patient based on the criteria listed in the Diagnostic and Statistical Manual of Mental Disorders, 4th Edition (DSM-IV) or DSM-5, and performed unstructured interviews and reviews of the patient’s medical records at each hospital. The control participants were interviewed by a psychiatrist and screened for psychiatric disorders on the basis of unstructured interviews; the inclusion criteria were not having a present, past, and family history (first degree relatives) of psychiatric disorders or substance abuse diagnosis (excluding nicotine dependence).
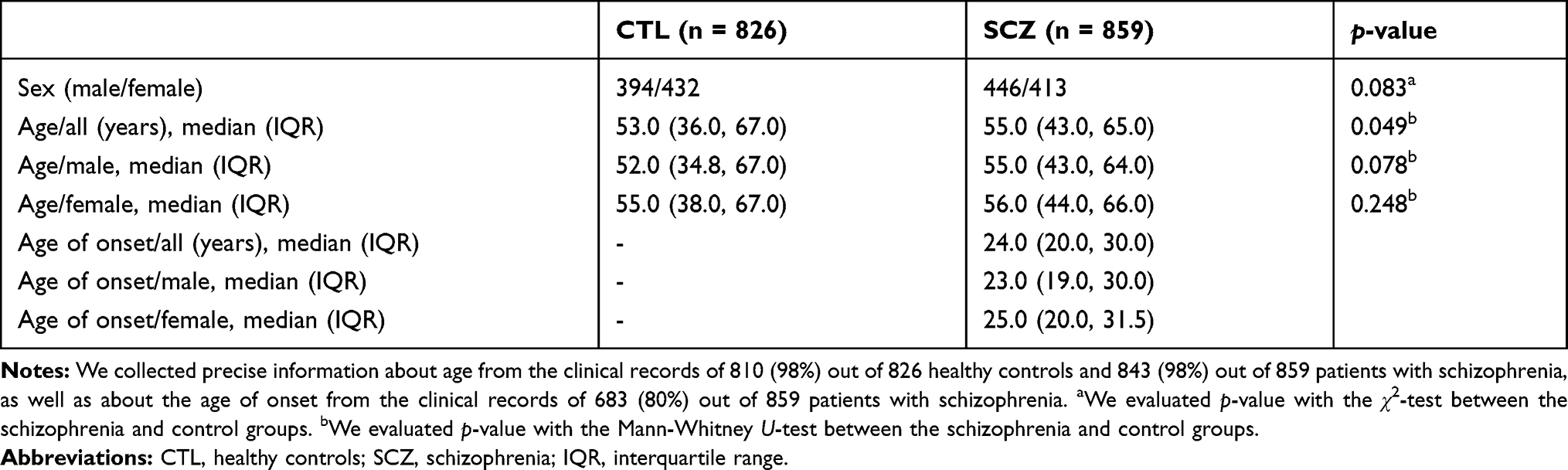 |
Table 1 Demographic and Clinical Characteristics of Participants |
We implemented this study design and all related procedures in accordance with the Declaration of Helsinki. This study was approved by the Ethical Committee for Genetic Studies of Kobe University Graduate School of Medicine. Written informed consent was obtained from all the participants prior to the commencement of the experiments.
Genotyping of uVNTR and dVNTR in the MAOA Gene Promoter
Peripheral blood samples were drawn from the participants, and DNA was extracted using QIAamp DNA Blood Midi Kit (Qiagen Inc., Valencia, CA, USA). The quantity and purity of the DNA were assessed via NanoDrop (Thermo Fisher Scientific, Waltham, MA, USA) and the DNA samples were stored at −80 °C until analysis. For the MAOA uVNTR and dVNTR genotyping, we performed the fluorescence-based fragment polymerase chain reaction (PCR) assay according to previous studies.8,20 The MAOA uVNTR PCR assay volume (10 µL) contained 1 ng of genomic DNA, 5 µL of AmpliTaq Gold Master Mix (Applied Biosystems, Foster City, CA, USA), and 15 pmol each of the following primers: 5ʹ-GAA CGG ACG CTC CAT TCG GA-3ʹ as a forward primer labeled with 6- Fluorescence (FAM) and 5ʹ-ACA GCC TGA CCG TGG AGA AG-3ʹ as a reverse primer (Invitrogen, Carlsbad, CA, USA). Thermal cycling comprised 10 min of initial denaturing at 95 °C followed by 40 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, and a final extension step of 7 min at 72 °C. The MAOA dVNTR PCR assay volume (20 µL) contained 10 ng of genomic DNA, 10 µL of AmpliTaq Gold Master Mix with 10% GC enhancer (Applied Biosystems), 0.1 µM of 7 deaza-dGTP (Bio Labs, NewYork, NY, USA) and 25 pmol of each the primers: 5ʹ-GGG TTA AGC GCC TCA GCT TC-3ʹ as a forward primer labeled with 6-FAM and 5ʹ-CAA GAG TGG ACT TAA GGA AGC AG-3ʹ as a reverse primer (Invitrogen). Thermal cycling comprised 10 min of initial denaturing at 95 °C followed by 10 cycles of 95 °C for 20 s, the touchdown annealing steps from 65–56 °C for 20 s, and 72 °C for 30 s, 35 cycles of 95 °C for 20 s, 55 °C for 20 s, and 72 °C for 30 s, followed by a final extension step of 7 min at 72 °C. Both PCR the products were analyzed using SeqStudio Genetic Analyzer (Applied Biosystems) and GeneMapper Software version 6 (Applied Biosystems).
Genotyping of rs6323 and rs1137070
We used TaqMan SNP genotyping assays, rs6323 (Assy ID: ANEP6VZ) and rs1137070 (Assy ID:C___8878813_20), obtained from Thermo Fisher Scientific database (http://www.thermofisher.com) as described previously.21 Genotyping was performed on a 7500 Real-Time PCR System (Applied Biosystems) according to the manufacturer’s protocol.
Statistical Analysis
The data were analyzed using R version 4.0.0 (R development core team, Vienna, Austria) and EZR version 1.42 (Saitama Medical Center, Jichi Medical University, Saitama, Japan). We used Haploview version 4.2 (Dlay Lab, Broad Institute Cambridge, MA, USA)22 to analyze allele/haplotype frequencies and genetic association in females. Differences between the groups were analyzed using χ2 and Mann-Whitney U-tests. We examined genotype–based associations, and alleles and haplotypes using the Cochran–Armitage trend test and χ2_test, respectively, and permutation tests based on 10,000 replications were performed for the correction, as necessary. The threshold for statistical significance was defined as a two tailed p < 0.05.
Results
Since the X chromosome contains the MAOA gene, we analyzed each sex separately. We tested VNTR polymorphisms via Hardy–Weinberg equilibrium (HWE) in females with Fisher’s exact test. (dVNTR, control p = 0.783 and schizophrenia p = 0.984; uVNTR, control p = 0.780 and schizophrenia p = 0.108; rs6323, control p = 0.397 and schizophrenia p = 0.606; rs1137070, control p = 0.233 and schizophrenia p = 0.584). The genotype and allelic frequency for dVNTR, uVNTR, rs6323, and rs1137070 are shown in Tables 2 and 3. Neither the genotype nor allelic frequency of the VNTRs or SNPs was significantly different between the schizophrenia and control groups.
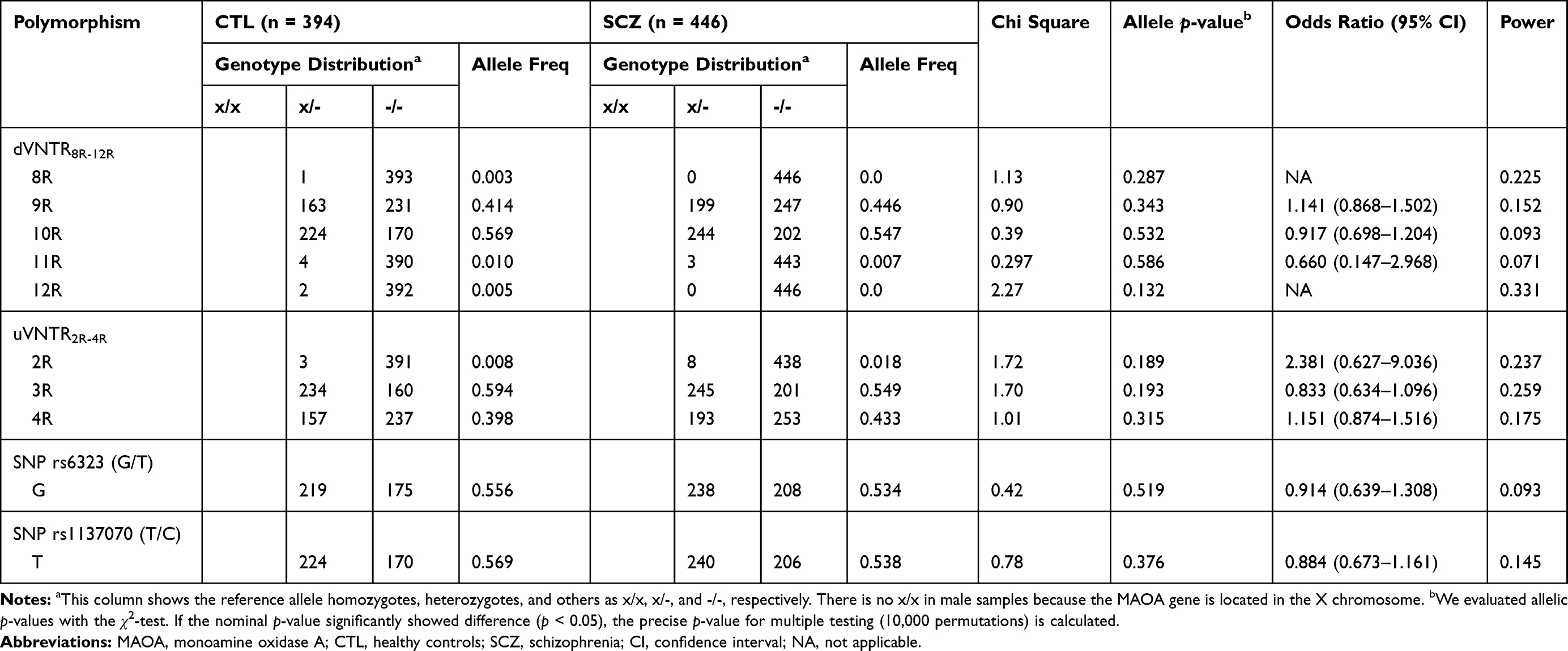 |
Table 2 Allelic and Genotypic Distribution of the Polymorphisms in the MAOA Promoter in Healthy Controls and Male Patients with Schizophrenia |
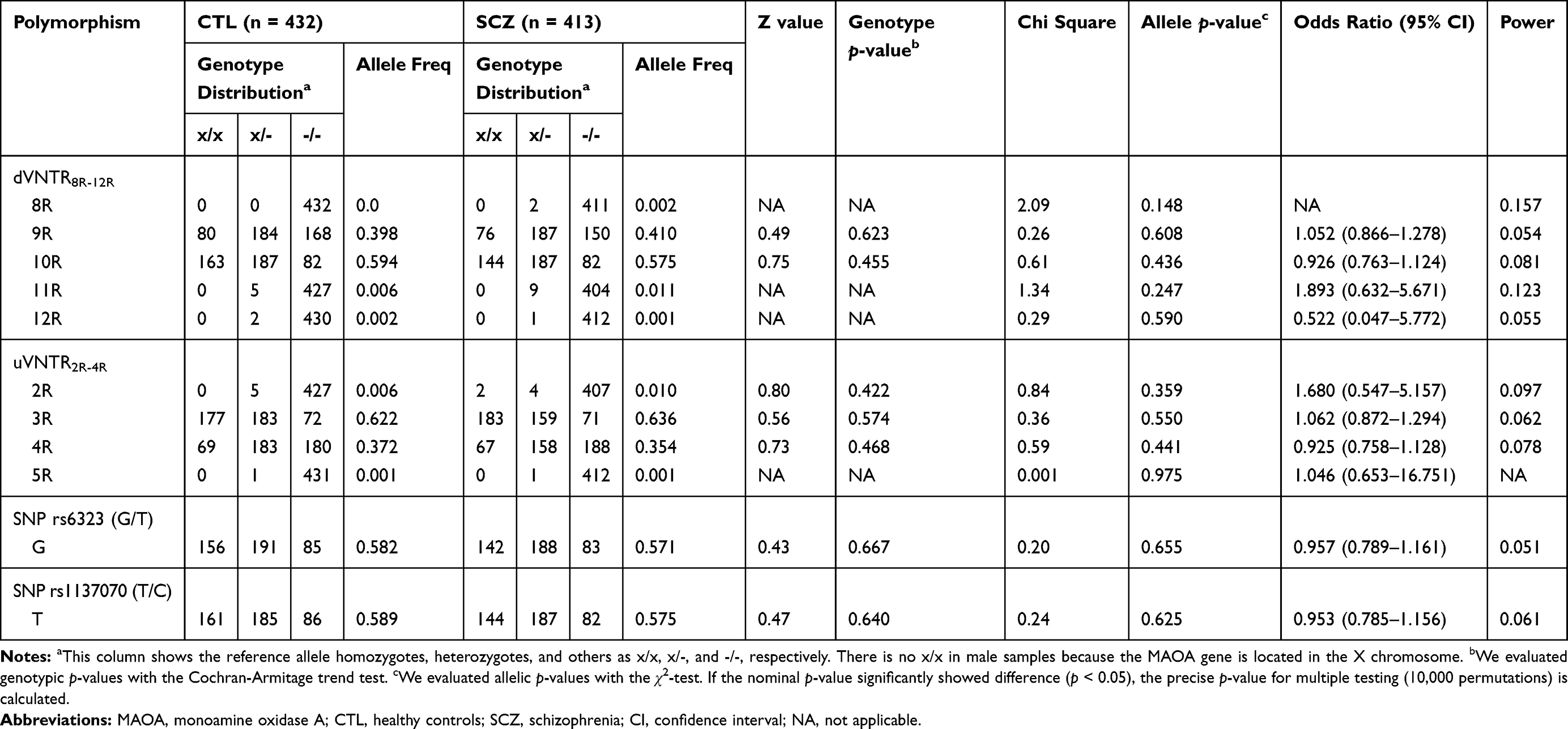 |
Table 3 Allelic and Genotypic Distribution of the Polymorphisms in the MAOA Promoter in Healthy Controls and Female Patients with Schizophrenia |
The association analysis of dVNTR-uVNTR-rs6323-rs1137070 haplotype is given in Table 4. The distribution of the haplotype comprising two VNTRs and the two SNPs showed a significant association for 9R-3R-T-C (p = 0.0001) and 10R-4R-G-T (p = 0.0139) in the female patients. Permutation tests based on 10,000 replications were performed, and significant differences were seen in the 9R-3R-T-C (p = 0.0006) haplotype, but not in the 10R-4R-G-T (p = 0.0799). Participants with the 9R-3R-T-C haplotype had 2.17 times increased odds of developing schizophrenia (odds ratio [confidence interval] = 2.17 [1.446–3.257], p = 0.0006). The haplotype containing dVNTR(9R) showed clearly significantly differences, when analyzed using the sliding window method (Table 5).
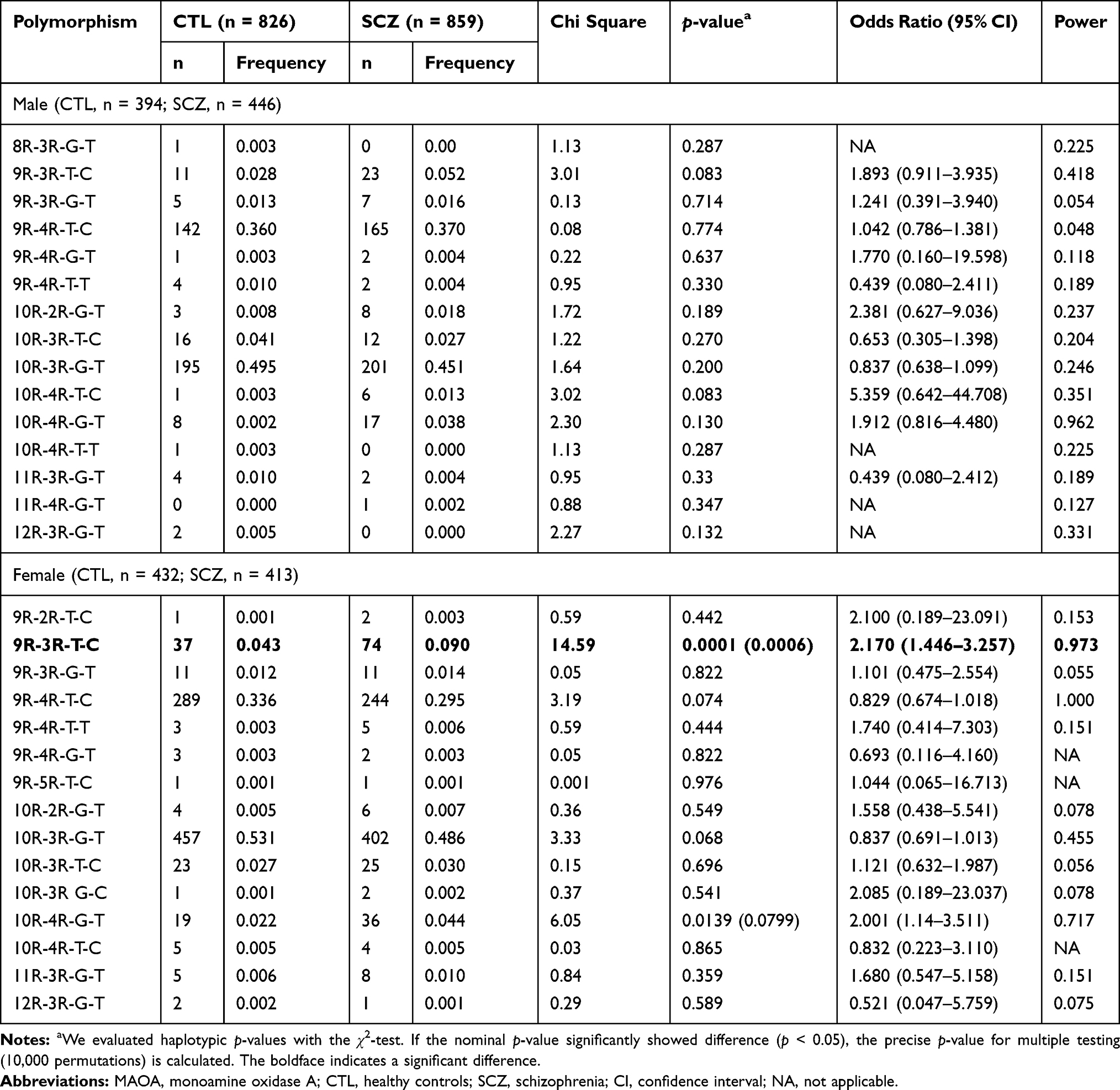 |
Table 4 Haplotypic Distribution of Polymorphisms in the MAOA Promoter in Controls and Patients with Schizophrenia |
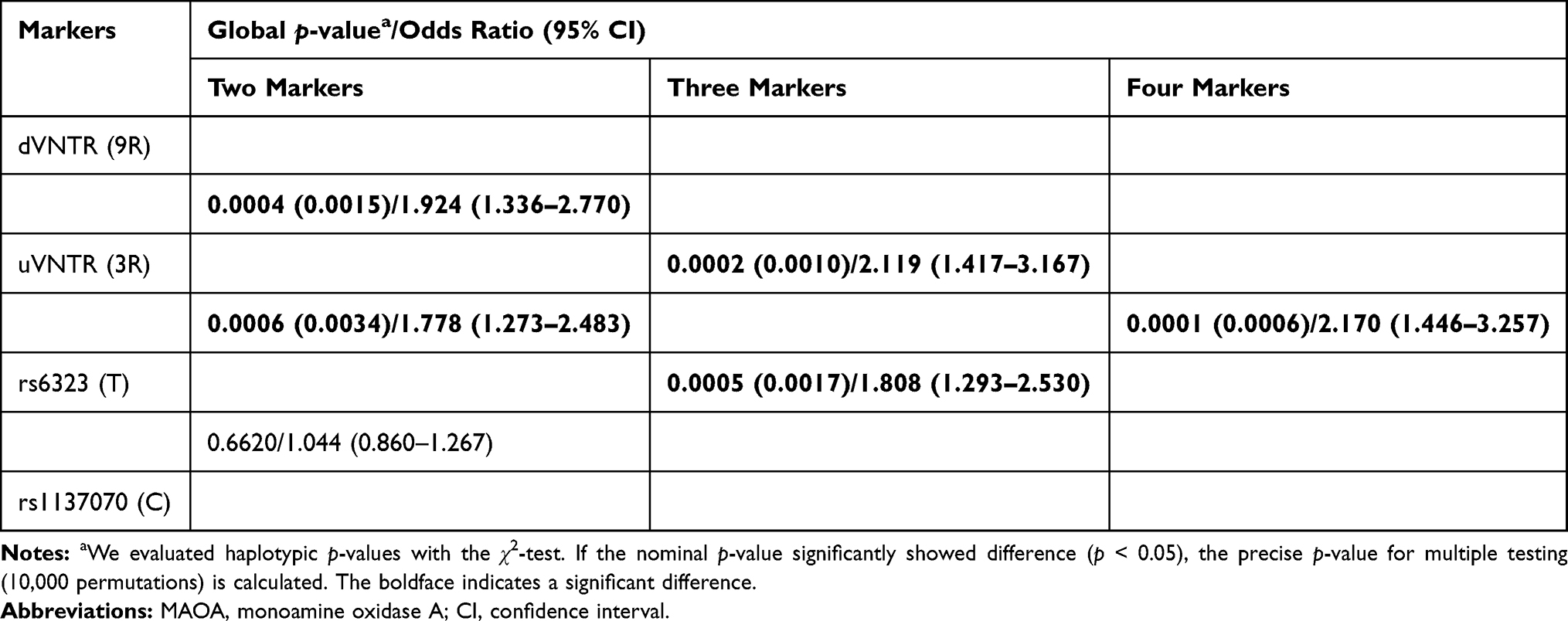 |
Table 5 Haplotype Analysis of the MAOA Promoter in Controls and Female Patients with Schizophrenia |
Discussion
In this study, we investigated whether the two VNTRs (dVNTR and uVNTR) and two SNPs (rs6323 and rs1137070) in the MAOA gene promoter are associated with schizophrenia. We found that the distribution of a haplotype consisting of the two VNTRSs and two SNPs was significantly associated with schizophrenia in females.
Although multiple studies have investigated the association of uVNTR and SNPs in the MAOA gene with schizophrenia, their results are not consistent.12–18 Furthermore, no studies have investigated the association between dVNTR and schizophrenia. To the best of our knowledge, this is the first study to investigate the association of the two MAOA VNTRs with schizophrenia.
We found that the dVNTR(9R)-uVNTR(3R)-rs6323(T)-rs1137070(C) haplotype was associated with schizophrenia in females. Previous studies have reported dVNTR(9R) as a high-expression allele,19 uVNTR(3R) as a low-expression allele,11 rs1137070(C) as a relatively high-expression allele,23 and that rs6323(T)-rs1137070(C) haplotype can lead to decreased MAO-A expression, leading to an increased risk of developing schizophrenia.14,15,24 These results including those of the current study also appear to be inconsistent. Only considering the function of each allele, and that low-expression alleles increase dopamine, which increases the risk of schizophrenia, is insufficient to clarify the molecular mechanism of schizophrenia.7,9,11,13,19 Schizophrenia is a multifactorial genetic disorder, and thus we need to investigate haplotypes with multiple variants and have a different viewpoint.15 The current study shows that several variants in the MAOA locus had a combined effects on the risk of developing schizophrenia. In the similar catabolic enzyme of dopamine, the variants allele affects dopamine levels in a specific region of the brain, which is related to the risk of developing schizophrenia.4 There is a possibility that low levels of MAO-A contribute to hyperfunction in the mesolimbic pathway resulting in positive symptoms of schizophrenia, whereas high levels of MAO-A contribute to hypofunction in the mesocortical pathway cortex leading to negative symptoms of schizophrenia with the combined effects of variant alleles.5,7 Therefore, the 9R-3R-T-C haplotype may have different functions for MAO-A expression dependent on brain regions (Figure 1). Further studies focusing on differences in the brain regions are required to determine the effects on MAO-A expression. Indeed, previous studies have reported that uVNTR is associated with the activity of different brain regions, contributing to the developments of psychiatric disorders.24,25
It was previously reported that uVNTR (low-expression alleles)-rs6323(T)-rs1137070(C) haplotype is associated with schizophrenia,14,15 in line with our findings. We found that the odds ratio of the 9R-3R-T-C haplotype (corrected p = 0.0006, odds ratio = 2.170) was higher than that of the 3R-T-C- haplotype (corrected p = 0.0017, odds ratio = 1.808) (Table 5), indicating that the haplotypes containing dVNTR(9R) may increase the role of MAO-A in the pathophysiology of schizophrenia. It is important to emphasize the combined effects that several variants had on the risk of schizophrenia and that finding new variants may help increase our current knowledge of the molecular mechanism underlying this disease.
Although recent large genome-wide association studies (GWASs) have shown that various SNPs are associated with schizophrenia,26–28 they did not include the MAOA gene polymorphisms that we considered in this study. Therefore, our findings warrant reconsideration of the previous studies while also bearing in mind the effects of VNTRs and haplotypes as well as sex differences.
Our study has several limitations. First, our sample size was relatively small, and our cohort comprised participants of only Japanese descent. Further studies with large samples sizes and different populations are required to validate our findings. Second, we did not consider longitudinal effects or detailed history of the symptoms, such as whether negative or positive symptoms were dominant.
Conclusion
To the best of our knowledge, this is the first study reporting the association of the two MAOA gene promoter VNTRs, and their related SNPs with schizophrenia. Our findings show that the dVNTR(9R)-uVNTR(3R)-rs6323(T)-rs1137070(C) haplotype was associated with schizophrenia in females, which may increase the risk of developing and help reveal the molecular mechanism of this disease.
Acknowledgments
We thank Makiko Ohnishi and Yasuko Nagashima for their technical assistance. This research was partly supported by JSPS KAKENHI grant numbers 15K19727, 18K15483, and 21K07520 (SO), as well as 26461718, 17H04249, and 21H02852 (AH).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Stepnicki P, Kondej M, Kaczor AA. Current concepts and treatments of schizophrenia. Molecules. 2018;23(8):2087. doi:10.1038/nature13595
2. Kendler KS, Diehl SR. The genetics of schizophrenia: a current, genetic-epidemiologic perspective. Schizophr Bull. 1993;19(2):261–285. doi:10.1093/schbul/19.2.261
3. Bortolato M, Chen K, Shih JC. Monoamine oxidase inactivation: from pathophysiology to therapeutics. Adv Drug Deliv Rev. 2008;60(13–14):1527–1533. doi:10.1016/j.addr.2008.06.002
4. Tunbridge EM, Harrison PJ, Weinberger DR. Catechol-o-methyltransferase, cognition, and psychosis: Val158Met and beyond. Biol Psychiatry. 2006;60(2):141–151. doi:10.1016/j.biopsych.2005.10.024
5. Schildkraut JJ, Orsulak PJ, Schatzberg AF, Herzog JM. Platelet monoamine oxidase activity in subgroups of schizophrenic disorders. Schizophr Bull. 1980;6(2):220–225. doi:10.1093/schbul/6.2.220
6. Meyer-Lindenberg A, Nichols T, Callicott JH, et al. Impact of complex genetic variation in COMT on human brain function. Mol Psychiatry. 2006;11(9):867–877. doi:10.1038/sj.mp.4001860
7. Berger PA, Ginsburg RA, Barchas JD, Murphy DL, Wyatt RJ. Platelet monoamine oxidase in chronic schizophrenic patients. Am J Psychiatry. 1978;135(1):95–99. doi:10.1176/ajp.135.1.95
8. Manca M, Pessoa V, Lopez AI, et al. The regulation of monoamine oxidase a gene expression by distinct variable number tandem repeats. J Mol Neurosci. 2018;64(3):459–470. doi:10.1007/s12031-018-1044-z
9. Meyer JH, Ginovart N, Boovariwala A, et al. Elevated monoamine oxidase a levels in the brain: an explanation for the monoamine imbalance of major depression. Arch Gen Psychiatry. 2006;63(11):1209–1216. doi:10.1001/archpsyc.63.11.1209
10. Ziegler C, Domschke K. Epigenetic signature of MAOA and MAOB genes in mental disorders. J Neural Transm (Vienna). 2018;125(11):1581–1588. doi:10.1007/s00702-018-1929-6
11. Sabol SZ, Hu S, Hamer D. A functional polymorphism in the monoamine oxidase A gene promoter. Hum Genet. 1998;103(3):273–279. doi:10.1007/s004390050816
12. Culej J, Nikolac Gabaj N, Stefanovic M, Karlovic D. Prediction of schizophrenia using MAOA-uVNTR polymorphism: a case-control study. Indian J Psychiatry. 2020;62(1):80–86. doi:10.4103/psychiatry.IndianJPsychiatry_54_19
13. Jönsson EG, Norton N, Forslund K, et al. Association between a promoter variant in the monoamine oxidase A gene and schizophrenia. Schizophr Res. 2003;61(1):31–37. doi:10.1016/s0920-9964(02)00224-4
14. Qiu HT, Meng HQ, Song C, et al. Association between monoamine oxidase (MAO)-A gene variants and schizophrenia in a Chinese population. Brain Res. 2009;1287:67–73. doi:10.1016/j.brainres.2009.06.072
15. Sun Y, Zhang J, Yuan Y, Yu X, Shen Y, Xu Q. Study of a possible role of the monoamine oxidase A (MAOA) gene in paranoid schizophrenia among a Chinese population. Am J Med Genet B Neuropsychiatr Genet. 2012;159B(1):104–111. doi:10.1002/ajmg.b.32009
16. Li D, He L. Meta-study on association between the monoamine oxidase A gene (MAOA) and schizophrenia. Am J Med Genet B Neuropsychiatr Genet. 2008;147B(2):174–178. doi:10.1002/ajmg.b.30570
17. Sasaki T, Hattori M, Sakai T, et al. The monoamine oxidase-a gene and major psychosis in japanese subjects. Biol Psychiatry. 1998;44(9):922–924. doi:10.1016/S0006-3223(97)00522-2
18. Syagailo YV, Stöber G, Grässle M, et al. Association analysis of the functional monoamine oxidase A gene promoter polymorphism in psychiatric disorders. Am J Med Genet B Neuropsychiatr Genet. 2001;105(2):168–171. doi:10.1002/ajmg.1193
19. Philibert RA, Wernett P, Plume J, Packer H, Brody GH, Beach SR. Gene environment interactions with a novel variable Monoamine Oxidase A transcriptional enhancer are associated with antisocial personality disorder. Biol Psychol. 2011;87(3):366–371. doi:10.1016/j.biopsycho.2011.04.007
20. Koks G, Prans E, Ho XD, et al. Genetic interaction between two VNTRs in the MAOA gene is associated with the nicotine dependence. Exp Biol Med (Maywood). 2020;245(8):733–739. doi:10.1177/1535370220916888
21. Okazaki S, Hishimoto A, Otsuka I, et al. Increased serum levels and promoter polymorphisms of macrophage migration inhibitory factor in schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2018;83:33–41. doi:10.1016/j.pnpbp.2018.01.001
22. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005;21(2):263–265. doi:10.1093/bioinformatics/bth457
23. Hotamisligil GS, Breakefield XO. Human monoamine oxidase A gene determines levels of enzyme activity. Am J Hum Genet. 1991;49:383–392.
24. Zhang J, Chen Y, Zhang K, et al. A cis-phase interaction study of genetic variants within the MAOA gene in major depressive disorder. Biol Psychiatry. 2010;68(9):795–800. doi:10.1016/j.biopsych.2010.06.004
25. Lee B-T, Ham B-J. Monoamine oxidase A–uVNTR genotype affects limbic brain activity in response to affective facial stimuli. Neuroreport. 2008;19(5):515–519. doi:10.1097/WNR.0b013e3282f94294
26. Ikeda M, Takahashi A, Kamatani Y, et al. Genome-wide association study detected novel susceptibility genes for schizophrenia and shared trans-populations/diseases genetic effect. Schizophr Bull. 2019;45(4):824–834. doi:10.1093/schbul/sby140
27. Lam M, Chen CY, Li Z, et al. Comparative genetic architectures of schizophrenia in East Asian and European populations. Nat Genet. 2019;51(12):1670–1678. doi:10.1038/s41588-019-0512-x
28. Schizophrenia Working Group of the Psychiatric Genomics C. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511(7510):421–427. doi:10.1038/nature13595.
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.