")
Back to Journals » Infection and Drug Resistance » Volume 15
Association of CRISPR-Cas System with the Antibiotic Resistance and Virulence Genes in Nosocomial Isolates of Enterococcus
Authors Tao S , Chen H, Li N, Fang Y , Xu Y, Liang W
Received 8 September 2022
Accepted for publication 26 October 2022
Published 30 November 2022 Volume 2022:15 Pages 6939—6949
DOI https://doi.org/10.2147/IDR.S388354
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Shuan Tao,1,2 Huimin Chen,1 Na Li,3 Yewei Fang,2 Yao Xu,4 Wei Liang2
1School of Medicine, Jiangsu University, Zhenjiang, People’s Republic of China; 2Department of Clinical Laboratory, Ningbo First Hospital, Ningbo, People’s Republic of China; 3Bengbu Medical College, Bengbu, People’s Republic of China; 4School of Medicine, Ningbo University, Ningbo, People’s Republic of China
Correspondence: Wei Liang, Department of Clinical Laboratory, Ningbo First Hospital, No. 59. Liuting Road, Haishu District, Ningbo, ZheJiang, 315010, People’s Republic of China, Tel +86 15351883016, Fax +86-051885213100, Email [email protected]
Purpose: This study aimed to investigate the prevalence of the CRISPR-Cas system in nosocomial isolates of Enterococcus and their possible association with antibiotic resistance and virulence genes.
Materials and Methods: Identification and antimicrobial susceptibility of the microorganism were performed by the automatized VITEK 2 Compact system (bioMerieux, France). A total of 100 Enterococcus isolates were collected and identified by VITEK 2 Compact automatic microbial identification drug susceptibility analyzer. The prevalence of various CRISPR-Cas systems, antibiotic resistance genes and virulence genes were investigated by polymerase chain reaction (PCR). The prevalence of CRISPR-Cas systems associated with antibiotic resistance and virulence genes was performed by appropriate statistical tests.
Results: A total of 100 isolates of Enterococcus were identified and there were 62/100(62.0%) Enterococcus faecalis isolates and 38/100(38.0%) Enterococcus faecalis isolates. In total, 46 (46.0%) of 100 isolates had at least one CRISPR-Cas locus. CRISPR elements were more prevalent in Enterococcus faecalis isolates. The results of PCR demonstrated that CRISPR1-Cas, orphan CRISPR2, and CRISPR3-Cas were present in 23 (23.0%), 42 (42.0%) and 5 (5.0%) Enterococcus isolates, respectively. Compared with CRISPR-Casnegative isolates, the CRISPR-Cas positive isolates showed significant lower resistance rates against ampicillin, erythromycin, levofloxacin, tetracycline, vancomycin, gentamicin, streptomycin, and rifampicin. Presumably consistent with drug susceptibility, fewer CRISPR loci were identified in vanA, tetM, ermB, aac6’-aph(2”), aadE, and ant(6) positive isolates. There was a significant negative correlation between the CRISPR-Cas locus and the enterococcal virulence factors enterococcal surface protein (esp) gene.
Conclusion: In conclusion, the results indicated that the absence of the CRISPR-Cas system was negatively associated with some antibiotic resistance in clinical isolates of Enterococcus faecalis and Enterococcus faecium. Also, there was a negative correlation with the carriage of antibiotic resistance genes. Furthermore, CRISPR-Cas may prevent some isolates from acquiring certain virulence factors.
Keywords: Enterococcus, CRISPR-Cas system, antimicrobial-antibiotic resistance gene, virulence genes
Introduction
Enterococci are a group of facultative anaerobic Gram-positive opportunistic pathogens.1 Enterococcus faecium and Enterococcus faecalis are the dominant species of enterococci.2 One of the most prominent features of enterococci is their high level of intrinsic and acquired resistance to many antibiotics.3 Enterococci are inherently resistant to many first-line antimicrobial agents, such as clindamycin, cephalosporins, compound sulfonamide, and low-concentration aminoglycosides.4,5 In addition, Enterococci can acquire antimicrobial resistance through horizontal genes transfer (HGT) mediated by mobile gene elements (MGEs) such as plasmids, phages, and transposons, which is one of the main mechanisms that contribute to the spread of resistance genes between bacteria.6,7 Antibiotic resistance of enterococci and the ability to acquire and spread antibiotic resistance is a challenge in the clinical setting and increases the difficulty of treating enterococcal infectious diseases.8
The Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-associated (Cas) system is an acquired immune defense system with immune memory in prokaryotic genomes, which can effectively resist the acquisition of foreign mobile gene elements such as plasmids or phages to maintain the stability of bacterial genomes.9,10 CRISPR-Cas system is one of the factors limiting the development and evolution of bacterial antibiotic resistance.11 CRISPR-Cas is considered as a natural barrier to horizontal gene transfer and transmission of antibiotic resistance.12 The CRISPR-Cas system is widely present in the genomes of bacteria and archaea and is an integral part of their immune systems.13 The CRISPR-Cas system consists of a CRISPR array and the Cas gene family, in which the CRISPR array is composed of highly conserved repeat sequences and spacer sequences, with upstream leader sequences responsible for their transcription.14 The Cas gene family, the CRISPR-related protein genes, is highly conserved, which encodes the Cas protein with a domain capable of cleaving DNA double-strands.15 In the CRISPR array, the palindromic structure contained in the repeat sequence can form a conserved RNA secondary structure, which can bind to the Cas protein to form a complex and work together.16
The mechanism of CRISPR-Cas mainly includes adaptation, expression, and interference.17 When exogenous plasmids or bacteriophages invade bacteria/archaea, the internal CRISPR-Cas system of these prokaryotes is activated, and some gene segments of exogenous DNA (called pre-spacer sequences or protospacer sequences, proto-spacer) integration into the original CRISPR array, the process called adaptation.18 During the expression stage, when the homologous foreign gene invades the bacterium again, the CRISPR array is transcribed into the precursor CRISPR RNA (pre-crRNA) long chain, and then processed by Cas proteins into mature RNA units (crRNA), which are combined to form the Cas-crRNA complex.19,20 In the interference stage, crRNA recognizes, binds, and cleaves the homologous exogenous nucleic acid sequence, directing the Cas-crRNA complex to degrade the target molecule.21,22 Antibiotic resistance is mediated by the HGT between bacteria.23 HGT is mainly achieved through transformation, conjugation, and phage-mediated transduction, which is also one of the main ways for bacteria to obtain drug resistance genes beneficial to their survival.24 CRISPR-Cas system can defend against mobile genetic elements such as phages or plasmids.25 Recent studies have revealed that the presence of CRISPR-Cas system is associated with antibiotic sensitivity and lack of virulence traits. Studies have demonstrated a highly significant negative correlation between the presence of the CRISPR-Cas system in Enterococcus faecalis and acquired antibiotic resistance.26 Several studies have proposed the potential of the CRISPR-Cas system to regulate bacterial virulence. However, some researchers have found no evidence by a statistical model that the CRISPR-Cas system can prevent the occurrence of HGT on the time scale of bacterial evolution.27 Gholizadeh et al11 demonstrated that there was no significant relationship between the CRISPR-Cas system and antibiotic resistance in Escherichia coli. The study of the function of the CRISPR-Cas system and its effect on HGT should be more comprehensive and in-depth.
The purpose of this study was to investigate the prevalence of CRISPR-Cas in clinical Enterococcus isolates and to analyze their association with antibiotic resistance genes and virulence factors. Understanding the relationship between CRISPR loci and antibiotic resistance phenotypes and genotypes as well as virulence genes may provide new insights into the fight against infections caused by enterococcal drug-resistant pathogens.
Methods and Materials
Bacterial Isolates
A total of 100 clinical enterococcal isolates, including 62 Enterococcus faecalis and 38 Enterococcus faecalis, were randomly collected in the Ningbo First Hospital from January to August 2022. All Enterococcus isolates were inoculated on Colombian blood agar plates and incubated at 37°C for 24 hours. And identified by VITEK 2 Compact automatic microbial identification drug susceptibility analyzer. Enterococcal drug susceptibility testing was performed with the automated broth microdilution methods (VITEK 2 Compact, BioMérieux, France) and matching cards for GP67. Operation and results interpretation was performed strictly according to the American Society for Clinical and Laboratory Standardization (CLSI) 2019 standard. All experiments were approved by the Medical Ethics Committee of Ningbo First Hospital.
Bacterial Nucleic Acid Extraction
The isolates were taken out from the −80 °C refrigerator and resuscitated, inoculated on blood plates, and cultured in 37 °C incubators for 18 h. One colony of each isolate was taken and inoculated in 1mL of Luria Bertani (LB) medium liquid medium at 37°C and 200 r/min for 24 hours. The DNA was extracted using the bacterial genomic DNA extraction kit (TIANGEN) according to the instructions.
Genotypic Detection of antibiotic Resistance Genes and Virulence Genes
Genotype analysis of antibiotic resistance genes and virulence genes for each Enterococcus isolate was based on the multiplex polymerase chain reaction (PCR). The antibiotic resistance genes included those encoding resistance to macrolide (ermA and ermB), tetracycline (tetM and tetO), penicillin (blaZ), aminoglycosides (aadE, aac(6’)-aph(2”), ant(6)), and glycopeptides (vancomycin/teicoplanin) (vanA and vanB) (Table 1). The investigated virulence genes were: the cylA (cytolysin activator), esp (enterococcal surface protein), asal (the sugar aggregation material), hyl (hyaluronidase), gelE (gelatinase), agg (aggregation substance) (Table 1). PCRs experiments were done in a final volume of 50 µL containing 25µL TaqPCR MasterMix (2X), 0.5 µL of each primer (10 pmol), 2 µL of template genomic DNA, and 22 µL of sterile water. The PCR cycle conditions consisted of 94°C pre-denaturation for 5 min; 30 cycles of 94°C for 30s, 60 °C for 30s, 72°C for 45s, followed by a final extension at 72°C for 5 min.
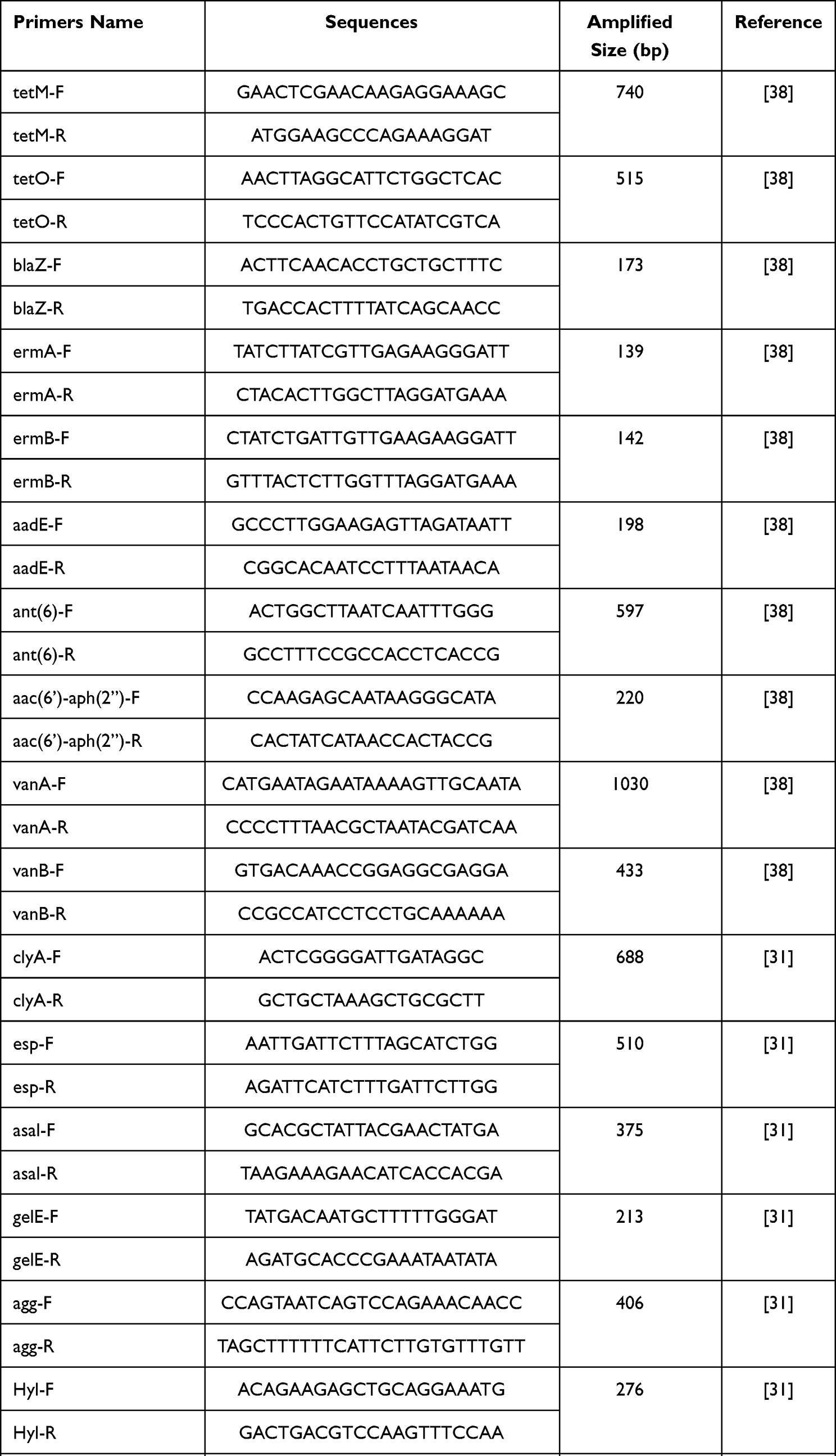 |
Table 1 Primers Used for the Detection of Antibiotic Resistance Genes, Virulence Genes and CRISPR-Cas Associated Genes Among Enterococcus Isolates |
Detection of CRISPR-Cas Systems
The presence of the CRISPR locus was identified by PCR, including the three CRISPR sites and the Cas genes of CRISPR1 and CRISPR3 (Table 1). The 50 µL PCR reaction mixture for each locus contained 25 µL PCR master mix (2X), 1 µL of each primer (10pMol), 2 µL of template DNA, and 21 µL of sterile water. The amplification condition consisted of 94°C for 5 min, 30 cycles of 94°C for 30s, 60°C for 30s, and 72°C for 45s, followed by final elongation at 72°C for 5 min. All PCR products were separated by electrophoresis on 1% agarose gel in 1xTBE buffer at 100V for 30 minutes. Then the analysis was performed using the gel imaging system to identify the amplification of the target gene.
Statistical Analysis
SPSS version 17.0 was used for all descriptive statistical analyses. Chi-squared test and Fisher’s exact test were used to compare the association of the presence of CRISPR-Cas with antimicrobial susceptibility, antibiotic resistance genes, and virulence genes in both Enterococcus faecalis and Enterococcus faecium, and P < 0.05 was considered as statistical significance.
Results
Source of Isolates
A total of 100 Enterococcus isolates were collected (62 Enterococcus faecalis and 38 Enterococcus faecium). The main specimens of clinical Enterococcus faecalis isolates were urine (38,61.29%), pus (20, 28.99%), puncture fluid (3, 4.84%), and secretions (1, 1.61%). The main specimen sources of Enterococcus faecium isolates were urine (28, 73.68%), blood (4, 10.53%), secretion (3, 7.89%), and puncture fluid (2, 5.26%). The distribution of these isolates is shown in Table 2.
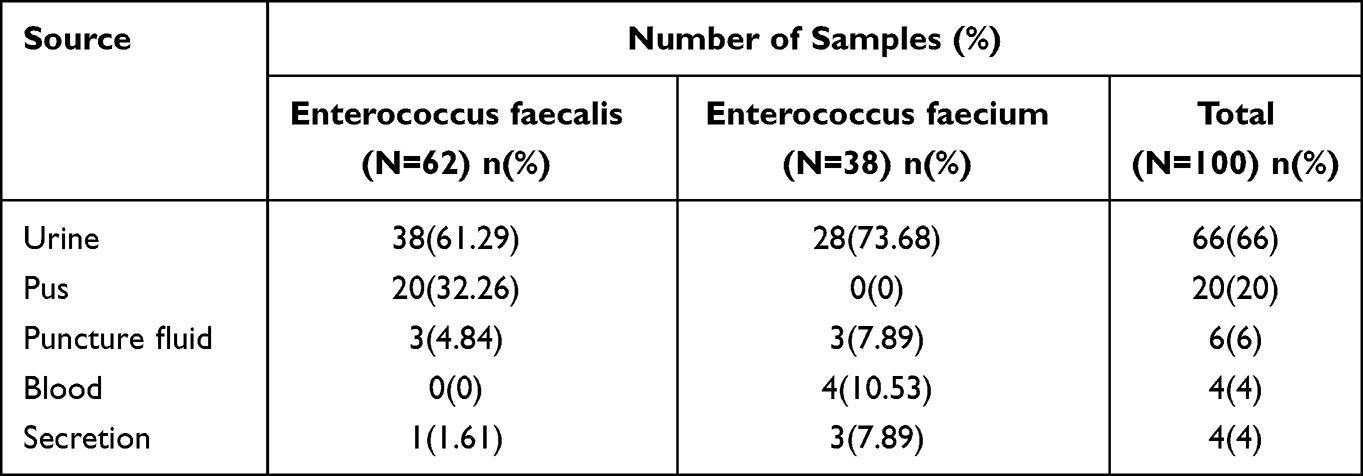 |
Table 2 The Distribution of Enterococcus Isolates |
Drug Susceptibility Test of Enterococcus Isolates
The phenotypic drug susceptibility testing among Enterococcus faecalis and Enterococcus faecium are shown in Table 3. Enterococci showed the highest rates of resistance to tetracycline, followed by erythromycin, ciprofloxacin, and levofloxacin, and showed the lowest rates of resistance to linezolid, teicoplanin, and vancomycin. Overall, the resistance rate of Enterococcus faecium was higher than that of Enterococcus faecalis, and the prevalence rate of ampicillin, ciprofloxacin, nitrofurantoin, levofloxacin, vancomycin, streptomycin, teicoplanin, and rifampicin resistance of Enterococcus faecium was significantly higher than that of Enterococcus faecalis (P <0.05).
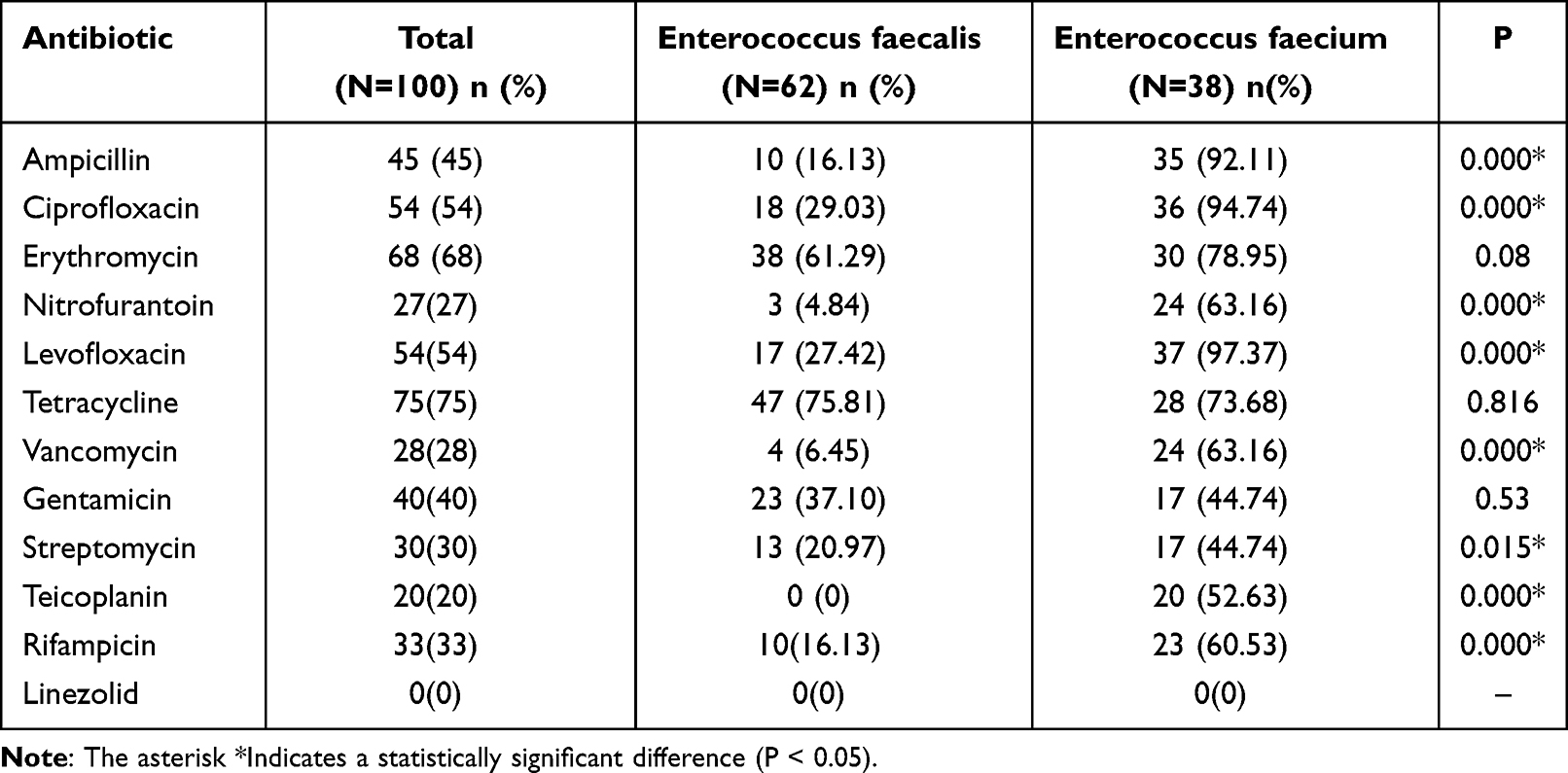 |
Table 3 Phenotypic Characteristics and Resistance Rates of Different Enterococcus Isolates to the Antibiotics |
Distribution of Antibiotic Resistance Genes and Virulence Genes in Enterococcus Isolates
The distribution of resistance and virulence genes in Enterococcus faecalis and Enterococcus faecium isolates are shown in Table 4. Overall, the ermB, aac (6 “) -aph (2“), aadE, and tetM genes were present in most isolates, while the blaZ gene was present in only 5% of isolates. The tetO, ermA, and vanB genes were not detected in all isolates. The prevalence rate of tetM, aac (6”)-aph (2”), and vanA genes were significantly different between E. faecalis and E. faecium isolates (p <0.05).
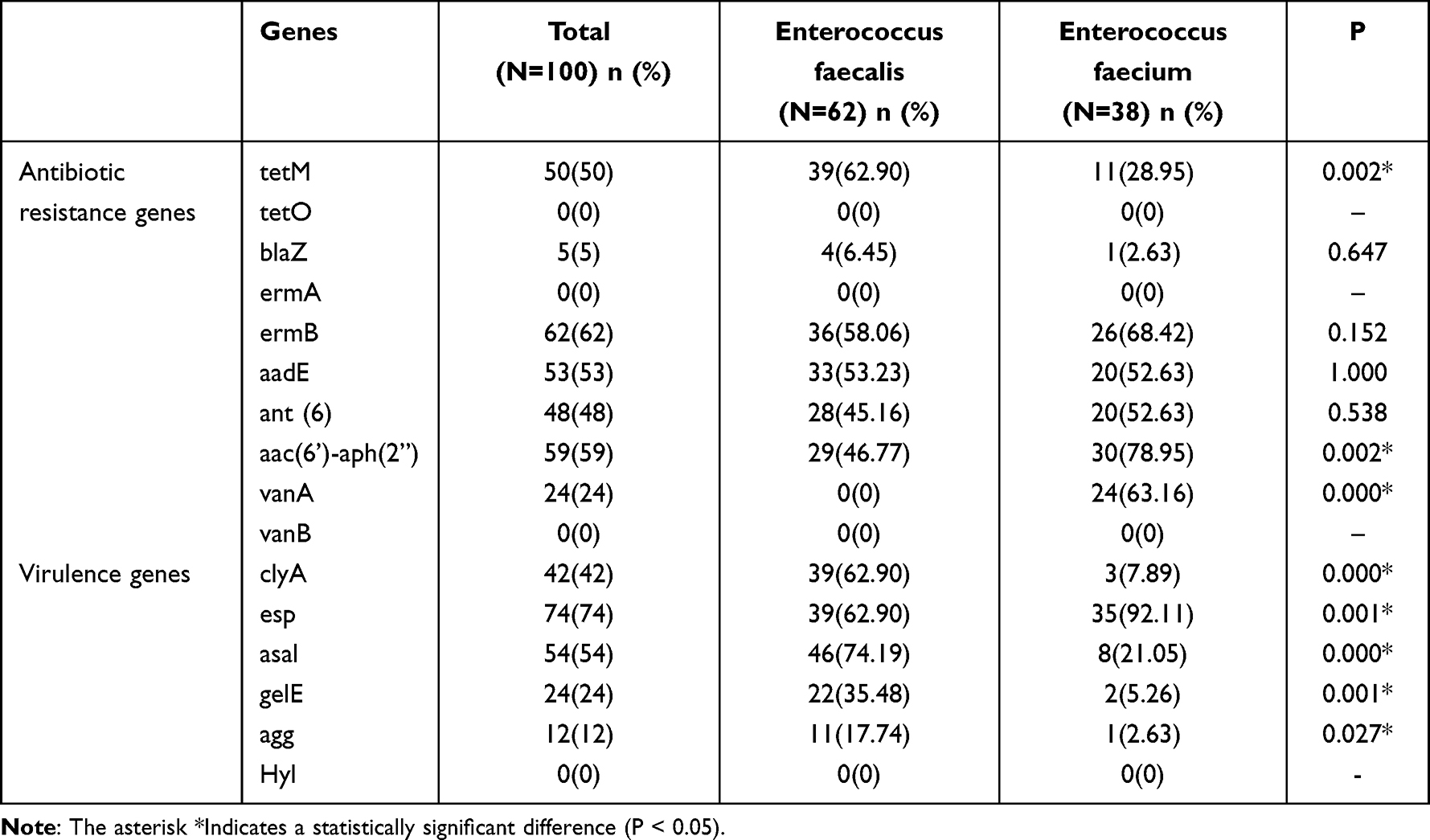 |
Table 4 The Distribution of Antibiotic Resistance-Related Genes and Virulence Genes Between Enterococcus faecalis and Enterococcus faecium Isolates |
The distribution of virulence genes of Enterococcus isolates is shown in Table 4. The most presence of the virulence genes among Enterococcus isolates was the esp gene, followed by asal and clyA. The prevalence of clyA, asal, gelE, and agg genes in Enterococcus faecalis were significantly higher than that in Enterococcus faecium, while the prevalence of esp genes was lower than that in Enterococcus faecium (p <0.05).
Prevalence of CRISPR-Cas Systems in Enterococcus
The prevalence of CRISPR-Cas in Enterococcus faecalis isolates and Enterococcus faecium isolates are shown in Table 5. Overall, CRISPR-Cas were detected in 46/100 isolates, with 23(23%) isolates positive for CRISPR1-Cas, 42(42%) isolates for CRISPR2-positive and 5 (5%) isolates positive for CRISPR3-Cas. The occurrence of CRISPR-Cas was more common in Enterococcus faecalis (n = 32, 51.63%) than in Enterococcus faecium (n=14, 36.84%) isolates. Specifically, Among the Enterococcus faecalis isolates, 18(29.03%), 30(48.39%), and 4(6.45%) were identified as CRISPR1-Cas, orphan CRISPR2, and CRISPR3-Cas, respectively. CRISPR1-Cas, orphan CRISPR2, and CRISPR3-Cas were observed in 5 (13.16%), 8(31.58%), and 1 (2.63%) Enterococcus faecium isolates, respectively.
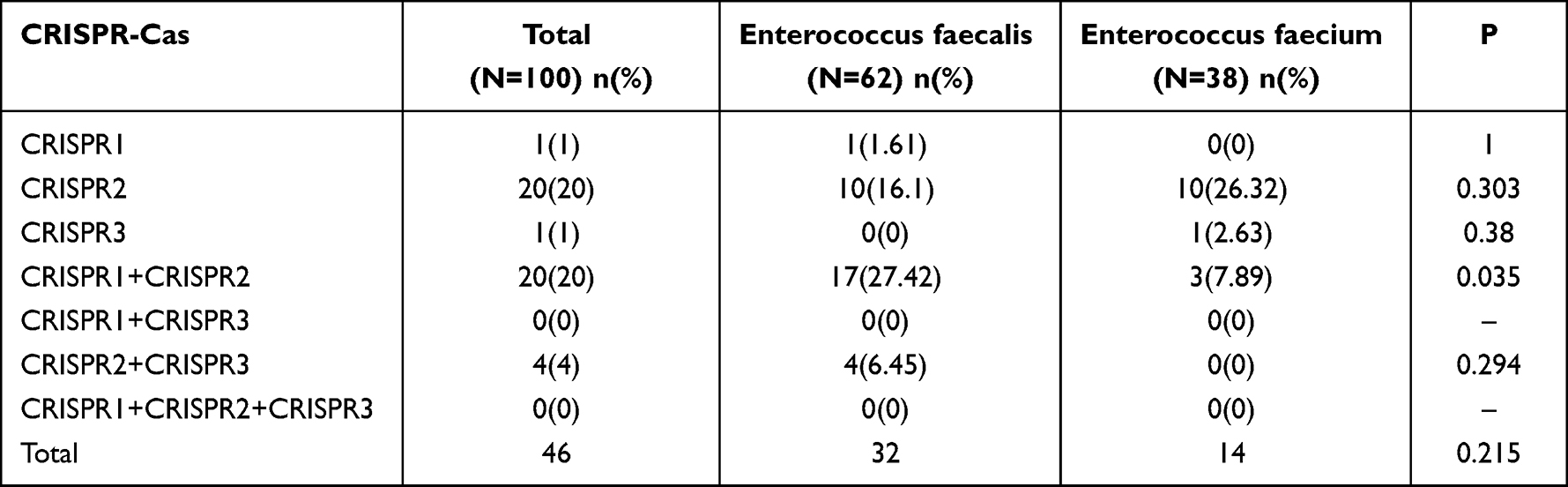 |
Table 5 The Prevalence of CRISPR-Cas in Enterococcus faecalis Isolates and Enterococcus faecium Isolates |
Association Between Antibiotic Resistance Genes, Virulence Genes, and the CRISPR-Cas System
The occurrence of antibiotic resistance genes and virulence genes between CRISPR-Cas-positive and -negative isolates of Enterococcus faecalis and Enterococcus faecium are shown in Table 6. The occurrence of CRISPR-Cas elements in Enterococcus isolates negatively correlated with antibiotic resistance rates. The prevalence of these antibiotic-resistance-associated genes was lower in the CRISPR- Cas-positive isolates compared to the CRISPR-Cas-negative isolates. The distribution of tetM, ermB, aadE, ant (6), and aac (6’) -aph (2”) between the two groups was statistically significant (p <0.05) (Table 6). In addition, compared with CRISPR-Cas-negative Enterococcus faecium, CRISPR-Cas-positive Enterococcus faecium isolates were significantly less resistant to vancomycin (Table 6).
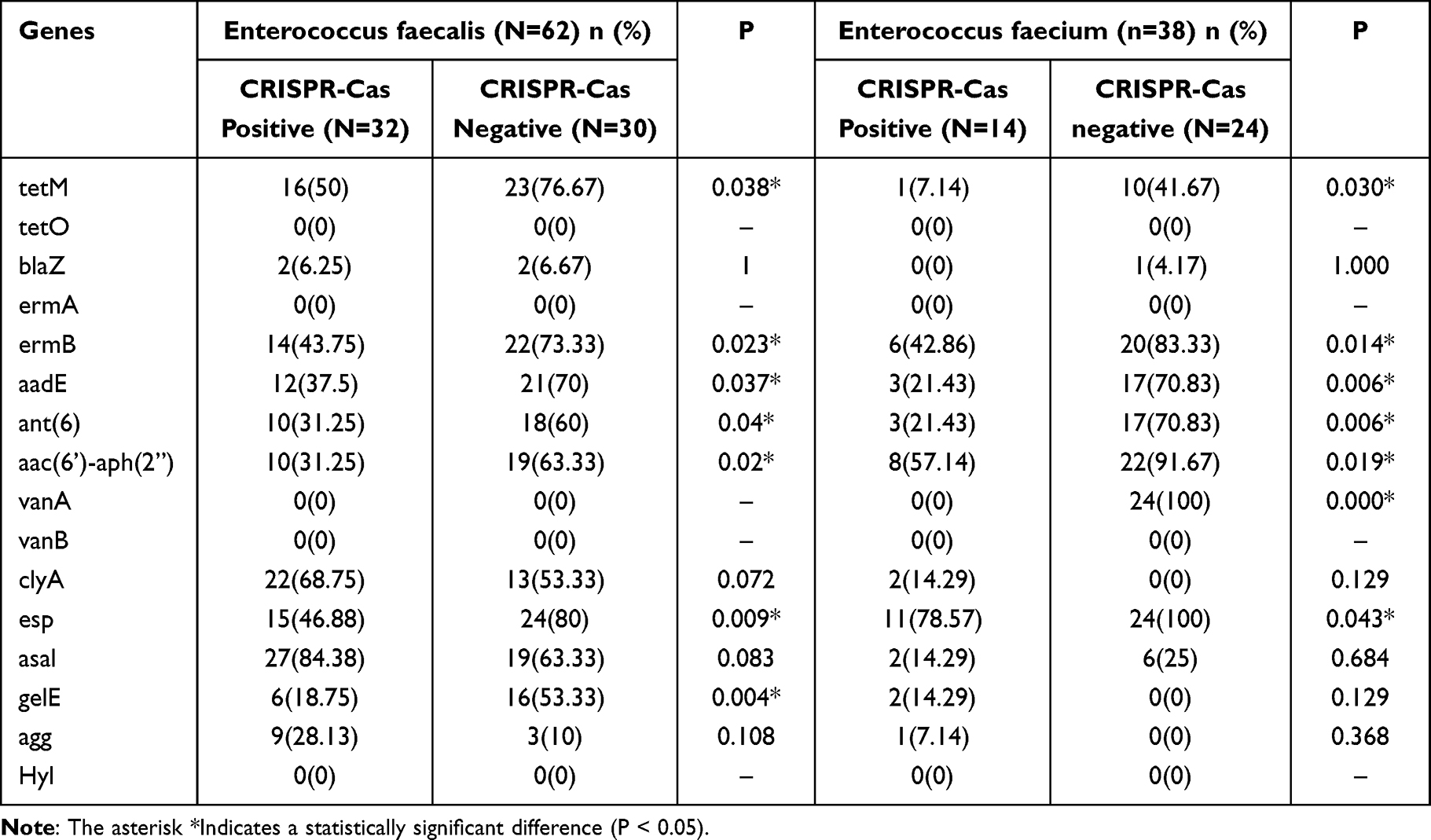 |
Table 6 The Correlation Between Drug-Resistance Genes and CRISPR-Cas of Enterococcus faecalis and Enterococcus faecium |
In Enterococcus faecalis isolates, the presence of asal virulence genes was the highest (74.19%), followed by esp (62.90%). The number of virulence genes esp (92.11%) in Enterococcus faecium isolates was higher. Both Enterococcus faecalis and Enterococcus faecium isolates have lower virulence genes agg (19.35%, 2.63%, respectively). None of the virulence gene hyl was found in all Enterococcus faecalis and Enterococcus faecium isolates. Compared with CRISPR-negative isolates, the Enterococcus faecalis virulence gene esp had a lower presence in CRISPR-positive isolates. In addition, the absence of Enterococcus faecalis virulence gene gelE was significantly associated with the presence of CRISPR-Cas (p<0.05).
Discussion
In this study, we assessed 62 clinical isolates of Enterococcus faecalis and 38 clinical isolates of Enterococcus faecium to determine the association between antibiotic resistance, virulence genes, and the CRISPR system. We found that the prevalence of antibiotic resistance of Enterococcus faecium is significantly higher than that of Enterococcus faecalis. In addition, the results demonstrated that the presence of the CRISPR-Cas system is associated with the absence of antibiotic resistance genes. The results also showed a significant correlation between the presence of the CRISPR-Cas system and fewer virulence factors.
In the current study, we found that the presence of orphan CRISPR2(42%) was more in Enterococcus than CRISPR1-Cas (23%) and CRISPR3-Cas (5%), which was similar to Alduhaidhawi.28 In non-clinical Enterococcus faecium isolates, Huescas et al29 demonstrated a higher prevalence rate of CRISPR2 and lower frequency of CRISPR1-Cas (5.6%) and CRISPR3-Cas (1.8%) system. In contrast to our study, Palmer et al30 found that CRISPR2 was present in all Enterococcus faecalis isolates. In addition. Dos Santos et al31 reported that CRISPR3 (67.4%) was the most prevalent type among Enterococcus isolates. Our results indicated that co-occurrence of CRISPR1 and CRISPR3, as well as CRISPR1, CRISPR2, and CRISPR3 were not found in the same isolates, which is similar to Linderstrauss et al.32 Similarly, in a study by Gholizadeh et al33 found that none of the isolates had the CRISPR1 and CRISPR3, as well as CRISPR1, CRISPR2, and CRISPR3 at the same time in the dental-root canal and hospital-acquired isolates of Enterococcus faecalis. However, Burley et al34 found three simultaneous sites of CRISPR1 and CRISPR3 (0.03%) in 88 root canals, oral cavity, and hospital-acquired isolates. Orphan CRISPR2 is thought to be inactive due to the lack of associated Cas proteins. A study revealed most multidrug-resistant Enterococcus faecalis isolates lack functional CRISPR-Cas and possess only the orphan CRISPR2.35 In Enterococcus isolates with only orphan CRISPR2, it was found to exist with some antibiotic resistance genes, such as aadE and ant(6), aac6’-aph(2”), indicating that CRISPR2 alone is not immune to Enterococcus. Studies have shown that the consensus repeats of the CRISPR1 and CRISPR2 loci are identical, which suggests that the two sites are functionally linked.36 In a study by Price et al demonstrated that the orphan CRISPR2 locus requires the presence of CRISPR1-Cas from Enterococcus faecalis for genomic defense against MGE.35
Antibiotic resistance genes are mainly transmitted through HGT.6 The acquired immune system CRISPR-Cas is considered as a barrier to HGT.37 Based on the presence of CRISPR-Cas, we compared the resistance rates of enterococci to different antibacterial drugs. In our study, there was a significant correlation between phenotypic antibiotic resistance, antibiotic resistance-related genes, and the absence of the CRISPR locus. Similarly, several studies have reported that multi-drug resistance was associated with the absence of the CRISPR locus.38–40 It has also been found that the loss of antibiotic resistance is associated with the presence of CRISPR3 in Enterococcus faecalis.34 Studies have shown that the presence of CRISPR1 in Enterococcus faecalis is associated with the lower pro-phage content of isolates such as Enterococcus faecalis OG1RF.41–43Studies show that CRISPR3-Cas is active in genomic defense with sequence specificity, which can be observed when the CRISPR3-mutant of T11 acquired cas9 (Δcas9 + CRISPR3) and interfered with pAD1 acquisition. They also observed that the deletion of only two loci could cause a significant reduction in genomic defense against the mobile genomic element(MGE).35
The presence of CRISPR-Cas was found to be associated not only with antibiotic sensitivity but also with the lack of virulence genes.33,44 The present results showed that the presence of the CRISPR locus was significantly associated with fewer virulence factors. Enterococcal infections and their pathogenicity are related to their carriage of virulence factors. Enterococcus virulence factors such as esp, asal, hyl, gelE, clyA, agg, etc. can play a role in anti-phagocytosis, adhesion, biofilm formation, exoenzyme, toxin, and so on.45 Literature reported that the CRISPR-Cas system is inversely correlated with some virulence factors.46 In this study, we found a correlation between the presence of CRISPR-Cas and the absence of the esp and gelE gene in Enterococcus faecalis (P = 0.009, P=0.004, respectively) and a correlation between the presence of CRISPR-Cas and the absence of esp (P = 0.0043) in Enterococcus faecium. In addition, there was a correlation between the absence of CRISPR loci and the absence of some virulence factors (such as clyA, agg). In a study by Gholizadeh et al indicated that CRISPR-Cas may prevent the acquisition of some pathogenic factors in dental root canals and hospital-acquired isolates of Enterococcus faecalis.33
Several studies have shown that the presence of the CRISPR-Cas system is inversely associated with the incidence of bacterial resistance.47 The negative impact of the CRISPR-Cas system on antibiotic resistance has also been demonstrated in K. pneumoniae.48 In addition, Ren et al49 revealed that point mutations in the cas1 and cas2 genes may be associated with multiple drug resistance in Shigella.
However, studies have also reported that there was no significant correlation between CRISPR-Cas and the acquisition of integron, plasmids, antibiotic resistance, and virulence genes in Escherichia coli.50,51 Shabbir et al52 claimed that the CRISPR-Cas system promotes antimicrobial resistance in Campylobacter jejuni. Therefore, further studies of CRISPR-Cas with mobile genetic elements associated with drug resistance are required.
Although the limited number of isolates included in the analysis in this study, reference can be provided for the prevention and control of enterococcal nosocomial infection. The relationship between CRISPR loci and antibiotic resistance phenotypes and genotypes may provide new insights into combating infections caused by resistant pathogens.
Conclusion
The study revealed that the CRISPR-Cas system of Enterococcus faecalis has a higher carriage rate compared to Enterococcus faecium and concluded that CRISPR-Cas system may hinder the transmission of antibiotic resistance genes and virulence genes, which provides a reference for the prevention and control of enterococcal nosocomial infection.
Acknowledgments
Thanks to Jiangsu University and the Central Laboratory of Ningbo First Hospital for supporting the work. Special thanks to my tutor Dr. Liang for my guidance.
Funding
This study was supported by grants from the Natural Science Foundation of Jiangsu Province (BK20191210), the fifth phase of the “333 Project” scientific research project in Jiangsu Province (BRA2019248), the Jiangsu Commission of Health (H2018073), and the Subject of Lianyungang Science and Technology Bureau (SF2015).
Disclosure
The authors have no conflict of interest to disclose.
References
1. Said MS, Tirthani E, Lesho E. Enterococcus infections. In: StatPearls. Treasure Island (FL): StatPearls Publishing Copyright © 2022, StatPearls Publishing LLC.; 2022.
2. Bhardwaj SB, Mehta M, Sood S, Sharma J. Isolation of a novel phage and targeting biofilms of drug-resistant oral enterococci. J Glob Infect Dis. 2020;12(1):11–15. doi:10.4103/jgid.jgid_110_19
3. Mull RW, Harrington A, Sanchez LA, Tal-Gan Y. Cyclic peptides that govern signal transduction pathways: from prokaryotes to multi-cellular organisms. Curr Top Med Chem. 2018;18(7):625–644. doi:10.2174/1568026618666180518090705
4. Gołaś-Prądzyńska M, Rola JG. Occurrence and antimicrobial resistance of enterococci isolated from goat’s milk. J Vet Res. 2021;65(4):449–455. doi:10.2478/jvetres-2021-0071
5. Shahini Shams Abadi M, Taji A, Salehi F, Kazemian H, Heidari H. High-level gentamicin resistance among clinical isolates of enterococci in Iran: a systematic review and meta-analysis. Folia Med (Plovdiv). 2021;63(1):15–23. doi:10.3897/folmed.63.e53506
6. McInnes RS, McCallum GE, Lamberte LE, van Schaik W. Horizontal transfer of antibiotic resistance genes in the human gut microbiome. Curr Opin Microbiol. 2020;53:35–43. doi:10.1016/j.mib.2020.02.002
7. Boccella M, Santella B, Pagliano P, et al. Prevalence and antimicrobial resistance of enterococcus species: a retrospective cohort study in Italy. Antibiotics. 2021;10(12):1552. doi:10.3390/antibiotics10121552
8. Chanda W, Manyepa M, Chikwanda E, et al. Evaluation of antibiotic susceptibility patterns of pathogens isolated from routine laboratory specimens at Ndola Teaching Hospital: a retrospective study. PLoS One. 2019;14(12):e0226676. doi:10.1371/journal.pone.0226676
9. Haider MZ, Shabbir MAB, Yaqub T, et al. CRISPR-Cas system: an adaptive immune system’s association with antibiotic resistance in Salmonella enterica serovar enteritidis. Biomed Res Int. 2022;2022:9080396. doi:10.1155/2022/9080396
10. Xu Y, Li Z. CRISPR-Cas systems: overview, innovations and applications in human disease research and gene therapy. Comput Struct Biotechnol J. 2020;18:2401–2415. doi:10.1016/j.csbj.2020.08.031
11. Gholizadeh P, Köse Ş, Dao S, et al. How CRISPR-Cas system could be used to combat antimicrobial resistance. Infect Drug Resist. 2020;13:1111–1121. doi:10.2147/IDR.S247271
12. Wheatley RM, MacLean RC. CRISPR-Cas systems restrict horizontal gene transfer in Pseudomonas aeruginosa. ISME J. 2021;15(5):1420–1433. doi:10.1038/s41396-020-00860-3
13. Cho S, Shin J, Cho BK. Applications of CRISPR/Cas system to bacterial metabolic engineering. Int J Mol Sci. 2018;19(4):1089. doi:10.3390/ijms19041089
14. Guo M, Chen H, Dong S, Zhang Z, Luo H. CRISPR-Cas gene editing technology and its application prospect in medicinal plants. Chin Med. 2022;17(1):33. doi:10.1186/s13020-022-00584-w
15. Koujah L, Shukla D, Naqvi AR. CRISPR-Cas based targeting of host and viral genes as an antiviral strategy. Semin Cell Dev Biol. 2019;96:53–64. doi:10.1016/j.semcdb.2019.04.004
16. Wright AV, Liu JJ, Knott GJ, Doxzen KW, Nogales E, Doudna JA. Structures of the CRISPR genome integration complex. Science. 2017;357(6356):1113–1118. doi:10.1126/science.aao0679
17. Nidhi S, Anand U, Oleksak P, et al. Novel CRISPR-Cas systems: an updated review of the current achievements, applications, and future research perspectives. Int J Mol Sci. 2021;22(7):3327. doi:10.3390/ijms22073327
18. Amitai G, Sorek R. CRISPR-Cas adaptation: insights into the mechanism of action. Nat Rev Microbiol. 2016;14(2):67–76. doi:10.1038/nrmicro.2015.14
19. Makarova KS, Wolf YI, Alkhnbashi OS, et al. An updated evolutionary classification of CRISPR-Cas systems. Nat Rev Microbiol. 2015;13(11):722–736. doi:10.1038/nrmicro3569
20. Charpentier E, Richter H, van der Oost J, White MF. Biogenesis pathways of RNA guides in archaeal and bacterial CRISPR-Cas adaptive immunity. FEMS Microbiol Rev. 2015;39(3):428–441. doi:10.1093/femsre/fuv023
21. Hille F, Charpentier E. CRISPR-Cas: biology, mechanisms and relevance. Philos Trans R Soc Lond B Biol Sci. 2016;371:1707.
22. Wan F, Draz MS, Gu M, Yu W, Ruan Z, Luo Q. Novel strategy to combat antibiotic resistance: a sight into the combination of CRISPR/Cas9 and nanoparticles. Pharmaceutics. 2021;13(3):352. doi:10.3390/pharmaceutics13030352
23. Lerminiaux NA, Cameron ADS. Horizontal transfer of antibiotic resistance genes in clinical environments. Can J Microbiol. 2019;65(1):34–44. doi:10.1139/cjm-2018-0275
24. Arnold BJ, Huang IT, Hanage WP. Horizontal gene transfer and adaptive evolution in bacteria. Nat Rev Microbiol. 2022;20(4):206–218. doi:10.1038/s41579-021-00650-4
25. Pinilla-Redondo R, Russel J, Mayo-Muñoz D, et al. CRISPR-Cas systems are widespread accessory elements across bacterial and archaeal plasmids. Nucleic Acids Res. 2022;50(8):4315–4328. doi:10.1093/nar/gkab859
26. Mulkerrins KB, Lyons C, Shiaris MP. Draft genome sequence of Enterococcus faecalis AS003, a strain possessing all three type II-a CRISPR loci. Microbiol Resource Announcements. 2021;10(11). doi:10.1128/MRA.01449-20
27. Gophna U, Kristensen DM, Wolf YI, Popa O, Drevet C, Koonin EV. No evidence of inhibition of horizontal gene transfer by CRISPR-Cas on evolutionary timescales. ISME J. 2015;9(9):2021–2027. doi:10.1038/ismej.2015.20
28. Alduhaidhawi AHM, AlHuchaimi SN, Al-Mayah TA, et al. Prevalence of CRISPR-Cas systems and their possible association with antibiotic resistance in Enterococcus faecalis and Enterococcus faecium collected from hospital wastewater. Infect Drug Resist. 2022;15:1143–1154. doi:10.2147/IDR.S358248
29. Huescas CGY, Pereira RI, Prichula J, Azevedo PA, Frazzon J, Frazzon APG. Frequency of clustered regularly interspaced short palindromic repeats (CRISPRs) in non-clinical Enterococcus faecalis and Enterococcus faecium strains. Braz J Biol. 2019;79(3):460–465. doi:10.1590/1519-6984.183375
30. Palmer KL, Gilmore MS. Multidrug-resistant enterococci lack CRISPR-cas. mBio. 2010;1(4). doi:10.1128/mBio.00227-10
31. Dos Santos BA, de Oliveira JDS, Parmanhani-da-Silva BM, Ribeiro RL, Teixeira LM, Neves FPG. CRISPR elements and their association with antimicrobial resistance and virulence genes among vancomycin-resistant and vancomycin-susceptible enterococci recovered from human and food sources. Infect Genet Evol. 2020;80:104183. doi:10.1016/j.meegid.2020.104183
32. Lindenstrauss AG, Pavlovic M, Bringmann A, Behr J, Ehrmann MA, Vogel RF. Comparison of genotypic and phenotypic cluster analyses of virulence determinants and possible role of CRISPR elements towards their incidence in Enterococcus faecalis and Enterococcus faecium. Syst Appl Microbiol. 2011;34(8):553–560. doi:10.1016/j.syapm.2011.05.002
33. Gholizadeh P, Aghazadeh M, Ghotaslou R, et al. CRISPR-cas system in the acquisition of virulence genes in dental-root canal and hospital-acquired isolates of Enterococcus faecalis. Virulence. 2020;11(1):1257–1267. doi:10.1080/21505594.2020.1809329
34. Burley KM, Sedgley CM. CRISPR-Cas, a prokaryotic adaptive immune system, in endodontic, oral, and multidrug-resistant hospital-acquired Enterococcus faecalis. J Endod. 2012;38(11):1511–1515. doi:10.1016/j.joen.2012.07.004
35. Price VJ, Huo W, Sharifi A, Palmer KL. CRISPR-Cas and restriction-modification act additively against conjugative antibiotic resistance plasmid transfer in Enterococcus faecalis. mSphere. 2016;1(3). doi:10.1128/mSphere.00064-16
36. Tomida J, Morita Y, Shibayama K, et al. Diversity and microevolution of CRISPR loci in Helicobacter cinaedi. PLoS One. 2017;12(10):e0186241. doi:10.1371/journal.pone.0186241
37. Zheng Z, Zhang Y, Liu Z, et al. The CRISPR-Cas systems were selectively inactivated during evolution of Bacillus cereus group for adaptation to diverse environments. ISME J. 2020;14(6):1479–1493. doi:10.1038/s41396-020-0623-5
38. Wu Q, Cui L, Liu Y, et al. CRISPR-Cas systems target endogenous genes to impact bacterial physiology and alter mammalian immune responses. Mol Biomed. 2022;3(1):22. doi:10.1186/s43556-022-00084-1
39. Hullahalli K, Rodrigues M, Palmer KL. Exploiting CRISPR-Cas to manipulate Enterococcus faecalis populations. eLife. 2017;6. doi:10.7554/eLife.26664
40. Gholizadeh P, Aghazadeh M, Ghotaslou R, et al. Role of CRISPR-Cas system on antibiotic resistance patterns of Enterococcus faecalis. Ann Clin Microbiol Antimicrob. 2021;20(1):49. doi:10.1186/s12941-021-00455-6
41. Palmer KL, Godfrey P, Griggs A, et al. Comparative genomics of enterococci: variation in Enterococcus faecalis, clade structure in E. faecium, and defining characteristics of E. gallinarum and E. casseliflavus. mBio. 2012;3(1):e00318–11. doi:10.1128/mBio.00318-11
42. McBride SM, Fischetti VA, Leblanc DJ, Moellering RC, Gilmore MS. Genetic diversity among Enterococcus faecalis. PLoS One. 2007;2(7):e582. doi:10.1371/journal.pone.0000582
43. Bourgogne A, Garsin DA, Qin X, et al. Large scale variation in Enterococcus faecalis illustrated by the genome analysis of strain OG1RF. Genome Biol. 2008;9(7):R110. doi:10.1186/gb-2008-9-7-r110
44. Hullahalli K, Rodrigues M, Nguyen UT, Palmer K. An attenuated CRISPR-Cas system in Enterococcus faecalis permits DNA acquisition. mBio. 2018;9(3). doi:10.1128/mBio.00414-18
45. Pillay S, Zishiri OT, Adeleke MA. Prevalence of virulence genes in Enterococcus species isolated from companion animals and livestock. Onderstepoort J Vet Res. 2018;85(1):e1–e8. doi:10.4102/ojvr.v85i1.1583
46. Roy S, Naha S, Rao A, Basu S. CRISPR-Cas system, antibiotic resistance and virulence in bacteria: through a common lens. Prog Mol Biol Transl Sci. 2021;178:123–174.
47. Pursey E, Dimitriu T, Paganelli FL, Westra ER, van Houte S. CRISPR-Cas is associated with fewer antibiotic resistance genes in bacterial pathogens. Philos Trans R Soc Lond B Biol Sci. 2022;377(1842):20200464. doi:10.1098/rstb.2020.0464
48. Wang G, Song G, Xu Y. Association of CRISPR/Cas system with the drug resistance in Klebsiella pneumoniae. Infect Drug Resist. 2020;13:1929–1935. doi:10.2147/IDR.S253380
49. Ren L, Deng LH, Zhang RP, et al. Relationship between drug resistance and the clustered, regularly interspaced, short, palindromic repeat-associated protein genes cas1 and cas2 in Shigella from giant panda dung. Medicine. 2017;96(7):e5922. doi:10.1097/MD.0000000000005922
50. Toro M, Cao G, Ju W, et al. Association of clustered regularly interspaced short palindromic repeat (CRISPR) elements with specific serotypes and virulence potential of Shiga toxin-producing Escherichia coli. Appl Environ Microbiol. 2014;80(4):1411–1420. doi:10.1128/AEM.03018-13
51. Touchon M, Charpentier S, Pognard D, et al. Antibiotic resistance plasmids spread among natural isolates of Escherichia coli in spite of CRISPR elements. Microbiology. 2012;158(Pt 12):2997–3004. doi:10.1099/mic.0.060814-0
52. Shabbir MA, Wu Q, Shabbir MZ, et al. The CRISPR-cas system promotes antimicrobial resistance in Campylobacter jejuni. Future Microbiol. 2018;13:1757–1774. doi:10.2217/fmb-2018-0234
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.