")
Back to Journals » The Application of Clinical Genetics » Volume 13
Association of Cholinergic Muscarinic M4 Receptor Gene Polymorphism with Schizophrenia
Authors Pozhidaev IV , Boiko AS , Loonen AJM , Paderina DZ , Fedorenko OY , Tenin G , Kornetova EG , Semke AV , Bokhan NA , Wilffert B , Ivanova SA
Received 25 January 2020
Accepted for publication 2 April 2020
Published 22 April 2020 Volume 2020:13 Pages 97—105
DOI https://doi.org/10.2147/TACG.S247174
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Martin Maurer
Ivan V Pozhidaev,1,2 Anastasiia S Boiko,1 Anton JM Loonen,3 Diana Z Paderina,1,2 Olga Yu Fedorenko,1,4 Gennadiy Tenin,5 Elena G Kornetova,1,6 Arkadiy V Semke,1 Nikolay A Bokhan,1,2,6 Bob Wilffert,3,7 Svetlana A Ivanova1,4,6
1Mental Health Research Institute, Tomsk National Research Medical Center of the Russian Academy of Sciences, Tomsk, Russian Federation; 2National Research Tomsk State University, Tomsk, Russian Federation; 3Unit of PharmacoTherapy, Epidemiology & Economics, Groningen Research Institute of Pharmacy, University of Groningen, Groningen, the Netherlands; 4National Research Tomsk Polytechnic University, Tomsk, Russian Federation; 5Institute of Cardiovascular Sciences, University of Manchester, Manchester, UK; 6Siberian State Medical University, Tomsk, Russian Federation; 7Department of Clinical Pharmacy and Pharmacology, University Medical Center Groningen, University of Groningen, Groningen, the Netherlands
Correspondence: Anton JM Loonen
University of Groningen, Groningen Research Institute of Pharmacy, PharmacoTherapy, Epidemiology & Economics, Antonius Deusinglaan 1, Groningen 9713AV, the Netherlands
Tel +31 50 363 7576
Fax +31 50 363 2772
Email [email protected]
Background: Previous studies have linked muscarinic M4 receptors (CHRM4) to schizophrenia. Specifically, the rs2067482 polymorphism was found to be highly associated with this disease.
Purpose: To test whether rs2067482 and rs72910092 are potential risk factors for schizophrenia and/or pharmacogenetic markers for antipsychotic-induced tardive dyskinesia.
Patients and Methods: We genotyped DNA of 449 patients with schizophrenia and 134 healthy controls for rs2067482 and rs72910092 polymorphisms of the CHRM4 gene with the use of the MassARRAY® System by Agena Bioscience. Mann–Whitney test was used to compare qualitative traits and χ2 test was used for categorical traits.
Results: The frequency of genotypes and alleles of rs72910092 did not differ between patients with schizophrenia and control subjects. We did not reveal any statistical differences for both rs2067482 and rs72910092 between schizophrenia patients with and without tardive dyskinesia. The frequency of the C allele of the polymorphic variant rs2067482 was significantly higher in healthy persons compared to patients with schizophrenia (OR=0.51, 95% CI [0.33– 0.80]; p=0.003). Accordingly, the CC genotype was found significantly more often in healthy persons compared to patients with schizophrenia (OR=0.49, 95% CI [0.31– 0.80]; p=0.010).
Conclusion: Our study found the presence of the minor allele (T) of rs2067482 variant being associated with schizophrenia. We argue that the association of rs2067482 with schizophrenia may be via its regulatory effect on some other gene with protein kinase C and casein Kknase substrate in neurons 3 (PACSIN3) as a possible candidate. Neither rs2067482 nor rs72910092 is associated with tardive dyskinesia.
Keywords: schizophrenia, suicide, tardive dyskinesia, PACSIN3, CHRM4, gene polymorphism
Introduction
Schizophrenia is a severe mental illness which often leads to significant long-term social or occupational disability of affected patients. The disease is characterized by positive symptoms such as delusions, hallucinations, and psychomotor agitation with hostility, but also by blunted and inadequate affect, concretisms, planning difficulties and many other negative and cognitive symptoms.1,2 The aetiology of schizophrenia has an important genetic component; its heritability is estimated to be between 64% and 81%.3 However, its mode of inheritance is complex and involves the combination of a variety of genes. A large, new body of evidence on genetics of schizophrenia has accumulated over recent years. Many new robustly associated genetic loci have been detected.3 An interesting development is the identification of a genetic variation in a region on chromosome 11, which also turns out to be associated with schizophrenia.4 This region on chromosome 11 contains the candidate genes activating molecule in beclin-1-regulated gene (AMBRA1), diacylglycerol kinase zeta gene (DGKZ), muscarinic acetylcholine receptor M4 gene (CHRM4) and Midkine (MDK). Genomic location of CHRM4 gene is illustrated in Figure 1. Previous studies have already linked muscarinic acetylcholine receptors M1 (CHRM1) and CHRM4 to schizophrenia.5,6 Further findings by Scarr et al showed that CHRM4 but not CHRM1 expression was decreased in the caudate-putamen and hippocampus from patients with this disorder.7
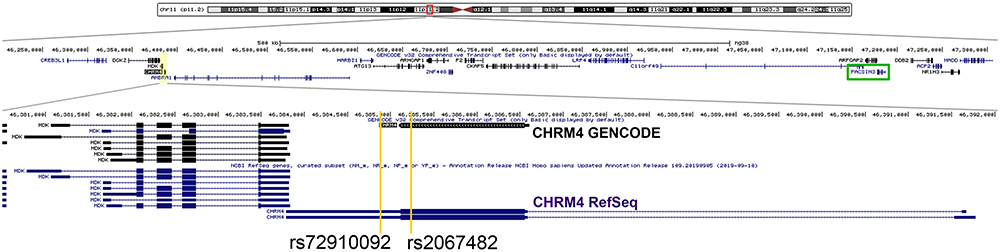 |
Figure 1 Genomic location of CHRM4 gene. Genomic location of the CHRM4 gene (chr15:46,200,000–47,300,000). Yellow lines highlight the location of two investigated variants in CHRM4 gene, rs2067482 and rs72910092. PACSIN3 gene is marked by green rectangle. |
Initially, two muscarinic receptors were distinguished; termed M1 and M2 (based on selective affinity for pirenzepine).8 Screening for cDNA or genes homologous with M1 and M2 revealed the existence of M3, M4, and M5 receptors.9 The localization and characteristics of human muscarinic receptor subtype genes are described by Michel and Teitsma.10 The human M4 receptor gene (CHRM4) consists of a single exon located on chromosome 11p12-p11.2. Two annotations available from GENCODE and RefSeq, differ in the size of 5ʹ- and 3ʹ-UTRs but have the same exon structure (Figure 1).
Muscarinic receptors belong to a family of over 800 G protein-coupled receptors (GPCRs).11,12 Based on the present knowledge in the brain M2 and M4 receptors interact with Gi-, Go-, and Gn-type G proteins (all inhibitory vs adenylate cyclase), while M1, M3 and M5 are coupled to Gq-type G proteins (activating phospholipase C which catalyzes the formation of diacylglycerol and inositol triphosphate (IP3) from phosphatidyl-inositol-4,5-biphosphate (PIP2)).13 Within the brain CHRM4 levels are particularly high in the dorsal striatum and relatively low in comparison to cholinergic muscarinic M1 receptors (CHRM1) in neocortex, amygdala and hippocampus.14 From anatomical studies, it can be concluded that a prominent role of M4 muscarinic receptors is mainly restricted to the (dorsal) striatum. In addition, within the striatum the inhibitory CHRM4 are primarily located on projection neurons constituting the excitatory direct extrapyramidal pathway, while stimulatory CHRM1 are present on projection neurons constituting the excitatory direct as well as the inhibitory indirect pathways.15 Hence, activation of excitatory CHRM1 as well as inhibitory CHRM4 results in a decrease of behavioral and cognitive output. This data suggests that changes in CHRM4 may be involved in the pathophysiology of positive and negative schizophrenia symptoms. According to online resources for research GeneMANIA database CHRM4 is involved in complicated genetic networks and pathways with a number of protein kinases, receptors and translation factors (Figure S1).
In a previous study, Scarr et al made an attempt to determine whether variations in the CHRM4 gene sequence were associated with an altered risk of schizophrenia.16 The CHRM4 gene was studied in 76 schizophrenia patients and 74 persons with no history of psychiatric disorders, and two genomic variants were identified, rs2067482 and rs72910092. Of these, only rs2067482, was found to be highly associated with schizophrenia, further linking CHRM4 to the disorder. Authors considered cholinergic muscarinic M4 receptor gene polymorphism not only as a potential risk factor, but also as a pharmacogenomic marker for schizophrenia. Motivated by their findings, we aimed to test two hypotheses in larger cohort of 472 patients with schizophrenia in comparison to 134 healthy controls, ie, whether these 2 SNPs (rs2067482 and rs72910092) are: 1) potential risk factors for schizophrenia, and 2) pharmacogenetic markers for antipsychotic-induced tardive dyskinesia, and discuss their possible significance.
Patients and Methods
Patients
This study was carried out in accordance with the Code of Ethics of the World Medical Association (Declaration of Helsinki 1975, revised in Fortaleza, Brazil, 2013), established for experiments involving humans. Each patient provided written informed consent, after the study was approved (protocol N63/7.2014) by the Local Bioethics Committee of the Mental Health Research Institute. Patient selection has been previously described.17 We recruited patients from three psychiatric hospitals located in the Tomsk, Kemerovo, and Chita oblasts (regions) of Siberia, Russia. The inclusion criteria were a clinical diagnosis of schizophrenia, according to the International Statistical Classification of Diseases and Related Health Problems, 10th Revision (ICD-10: F20), and age 18–75 years old. Detailed descriptive statistics are presented below (Table 1). Exclusion criteria were non-Caucasian physical appearance or family members (eg, Mongoloid, Buryats, or Khakassians); pregnancy, or any relevant physical disorder including organic brain disorders (eg, epilepsy, Parkinson’s disease); or relevant pharmacological withdrawal symptoms. Clinical examination and diagnostic assessment were carried out using the Positive and Negative Syndrome Scale (PANSS). The course of schizophrenia (continuous or episodic) was determined by ICD-10 – in the classification of ICD-10, the fifth character is used for this. In order to assess the presence or absence of tardive dyskinesia (TD), patients were assessed according to the abnormal involuntary movement scale (AIMS).18–20 The AIMS scores were transformed into a binary form (presence or absence of dyskinesia) with Schooler and Kane’s criteria.21 Schooler–Kane criteria require: (i) at least 3 months of cumulative exposure to neuroleptics; (ii) the absence of other conditions that might cause involuntary movements and (iii) at least moderate dyskinetic movements in one body area (≥3 on AIMS) or mild dyskinetic movements in two body areas (≥2 on AIMS). The presence of orofacial dyskinesia and limb-truncal dyskinesia was established by a cut-off score of 2 (mild but definite) on any of the items from 1 to 4 and from 5 to 7 of AIMS, respectively. To compare dosage of antipsychotic medications, all dosages were converted into chlorpromazine equivalents (CPZeq).22
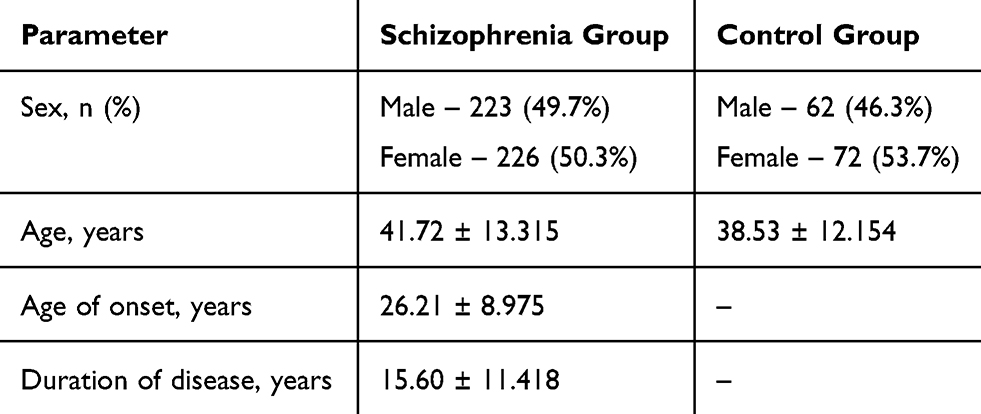 |
Table 1 Characteristics of Schizophrenia and Control Groups |
Healthy Probands
The control group (134 subjects) consisted of mentally and somatically healthy individuals with Caucasian background (based on both physical appearance and familial history) of both genders and aged 19–60 years old, who provided written informed consent. Exclusion criteria were chronically physical pathology in exacerbation, relevant pharmacological withdrawal symptoms, any organic brain disorders (eg, epilepsy, Parkinson’s disease) or mental disorders.
DNA Analysis
DNA was isolated from antecubital venous blood samples by means of a standard phenol-chloroform method. DNA concentrations were measured with a Thermo Scientific NanoDrop 8000 UV-Vis Spectrophotometer. All samples were genotyped without having knowledge of the patient’s clinical conditions. Genotyping for rs2067482 and rs72910092 polymorphisms of CHRM4 gene was carried out with the use of the MassARRAY® System by Agena Bioscience (San Diego, USA), kit iPLEX Gold 384, in the Laboratory of Genetics of the University of Groningen. Processing of the DNA samples included several steps: a standard PCR reaction to obtain the amplificate, a shrimp alkaline phosphatase (SAP) reaction to neutralize the unincorporated dNTPs in the amplificate, the PCR iPLEX Gold extension reaction, followed by placing the samples on a special chip (SpectroCHIP Array) using NanoDispenser RS1000 and loading them into the SEQUENOM MassARRAY® Analyzer 4.
Real-time mass spectrum analysis was conducted with special program software MassARRAY TYPER 4.0.
Statistical Analysis
Mann–Whitney test was used to compare qualitative traits and χ2 test was used for categorical traits. Genotype prevalence was checked for meeting Hardy–Weinberg equilibrium expectations by means of χ2 test. Allele and genotype frequencies were compared using χ2 test and Fisher’s exact test, if necessary. Statistically significant differences were considered at p<0.05. Odds ratios (ORs) and 95% confidence intervals (CIs) were calculated when estimating the effect size of genetic polymorphisms on the risk of schizophrenia and TD. All statistical analyses were performed using SPSS for Windows, v. 17. Bonferroni correction for multiple testing was applied to the resulting p values.
Results
After obtaining informed consent, 472 patients with schizophrenia were recruited in total, but only 449 of them were fulfilling the criterion for tardive dyskinesia (TD) of receiving the antipsychotic treatment for more than three months and therefore eligible for the subsequent study (Table 1).17 [40]. According to the other predefined criteria, 121 out of these 449 patients suffered from TD.
Table 2 presents the frequency of genotypes and alleles for rs2067482 and rs72910092 polymorphisms of CHRM4 gene in patients with schizophrenia and healthy persons.
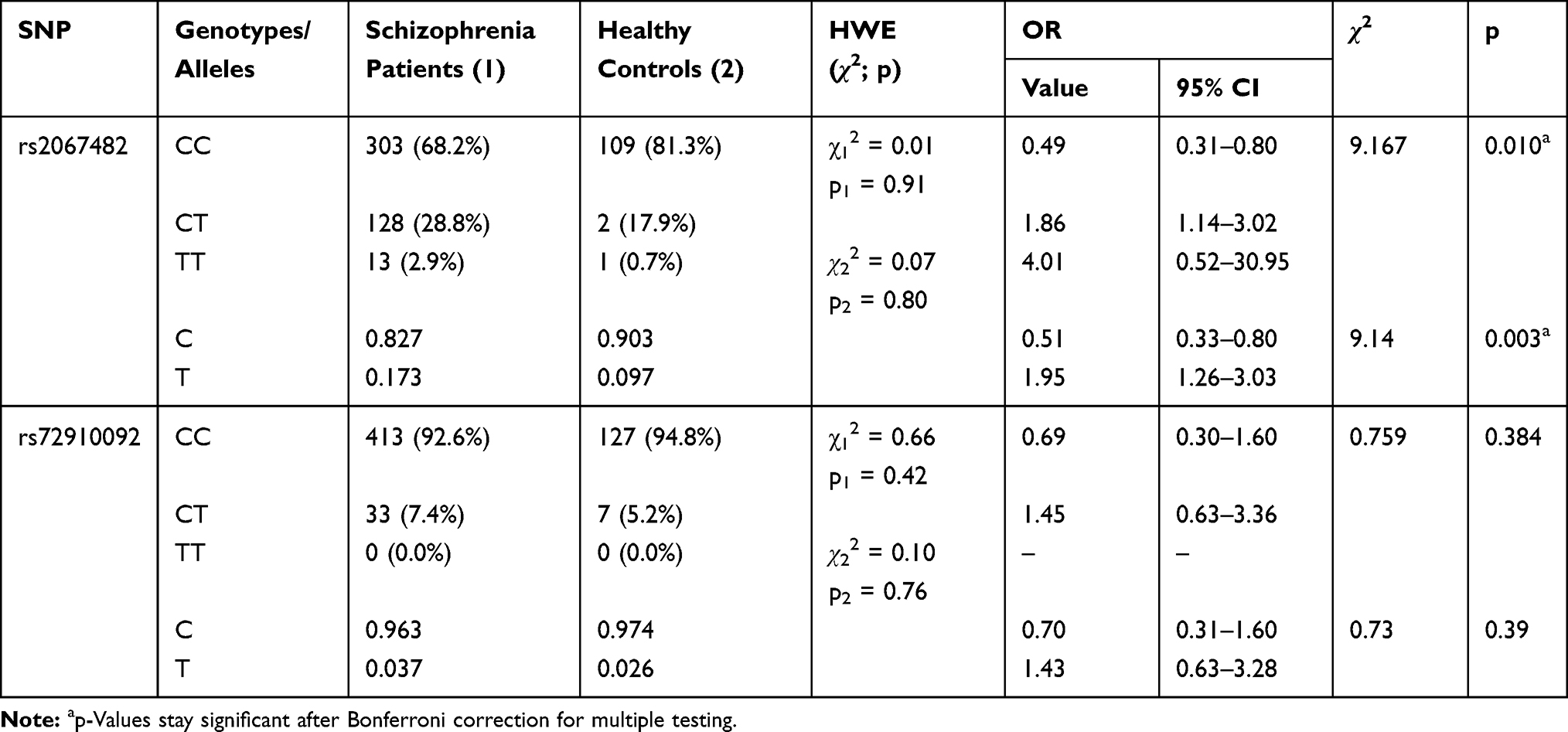 |
Table 2 Frequency of CHRM4 Genotypes and Alleles in Patients with Schizophrenia and Healthy Controls |
The prevalence of genotypes in both the schizophrenia and healthy groups was consistent with Hardy-Weinberg equilibrium. The frequency of the С allele of the polymorphic variant rs2067482 was significantly higher in healthy persons compared to patients with schizophrenia (Table 2; χ2=9.14, p=0.003; OR=0.51, 95% CI [0.33–0.80]). Accordingly, the CC genotype was found significantly more often in healthy persons compared to patients with schizophrenia (χ2=9.167, p=0.010; OR=0.49, 95% CI [0.31–0.80]).
We did not reveal any significant contribution of CHRM4 rs2067482 and rs72910092 to the course of schizophrenia (continuous or episodic) (Table 3).
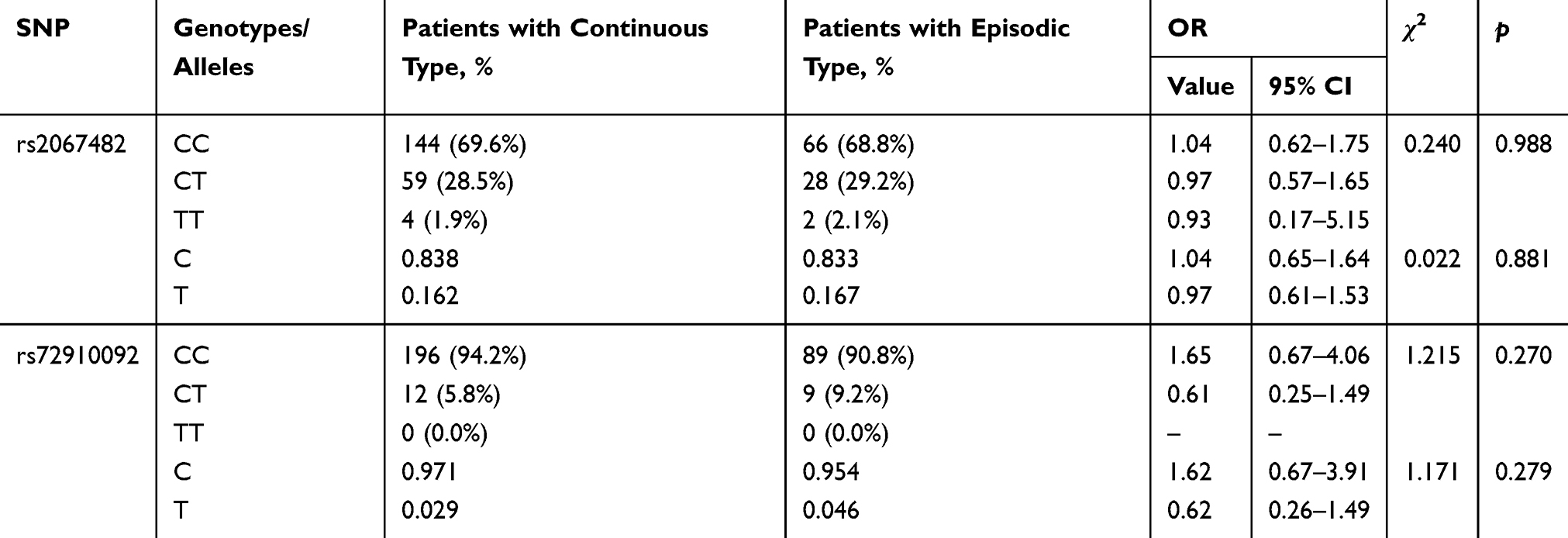 |
Table 3 Frequency of CHRM4 Genotypes and Alleles in Schizophrenia Patients with Different Types of Course of the Disease (Continuous vs Episodic) |
From the two SNPs studied for CHRM4, only rs2067482 showed significant association with leading symptoms of schizophrenia (positive vs negative, χ2=7.267, p=0.026). The rs2067482*CT (OR=1.63, 95% CI: 1.04–2.59) genotype was associated with positive leading symptoms (Table 4).
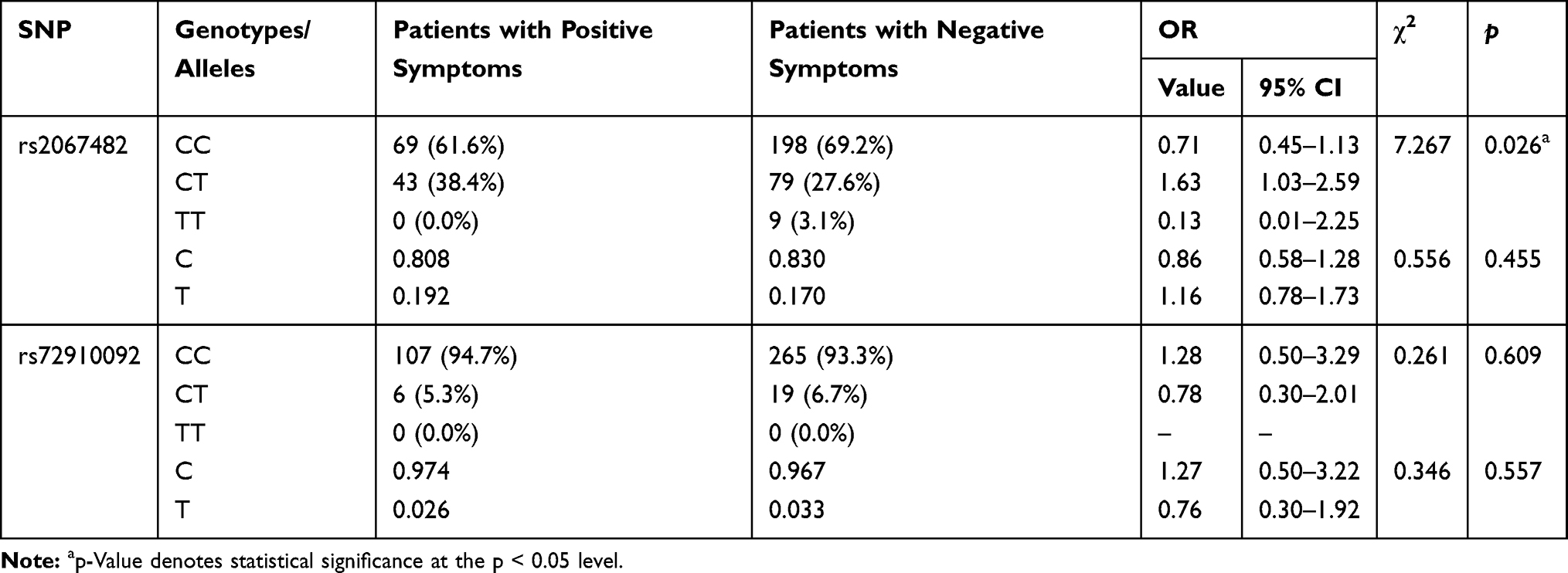 |
Table 4 Frequency of CHRM4 Genotypes and Alleles in Schizophrenia Patients with Different Leading Symptoms (Positive vs Negative) |
We did not reveal any statistical differences in the distribution of genotypes and alleles for rs2067482 and rs72910092 of CHRM4 between patients with and without TD (Table 5). In our statistical analysis both rs2067482 and rs72910092 were not linked to neither orofacial nor limb-truncal TD (Tables S1 and S2).
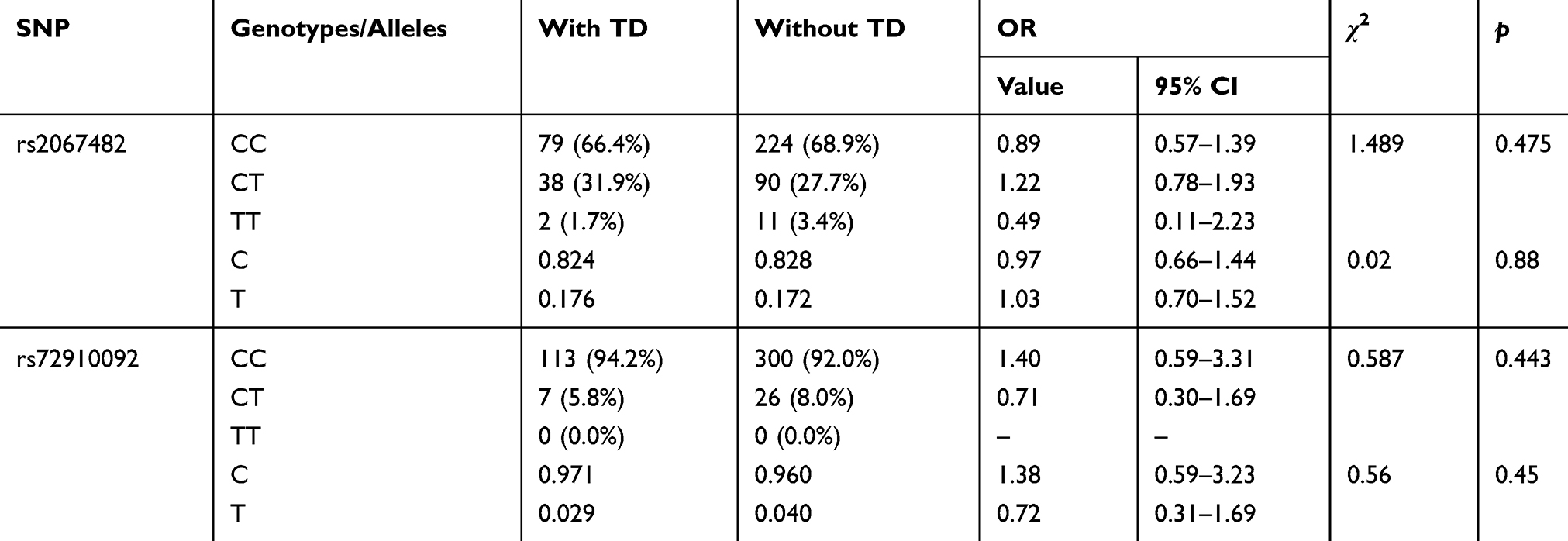 |
Table 5 CHRM4 Genotype Distribution in Schizophrenia Patients with and without Tardive Dyskinesia |
We did, however, obtain evidence of an association of rs2067482 with TD in our patient population in comparison with healthy controls (Table 6; χ2=7.42, p=0.02).
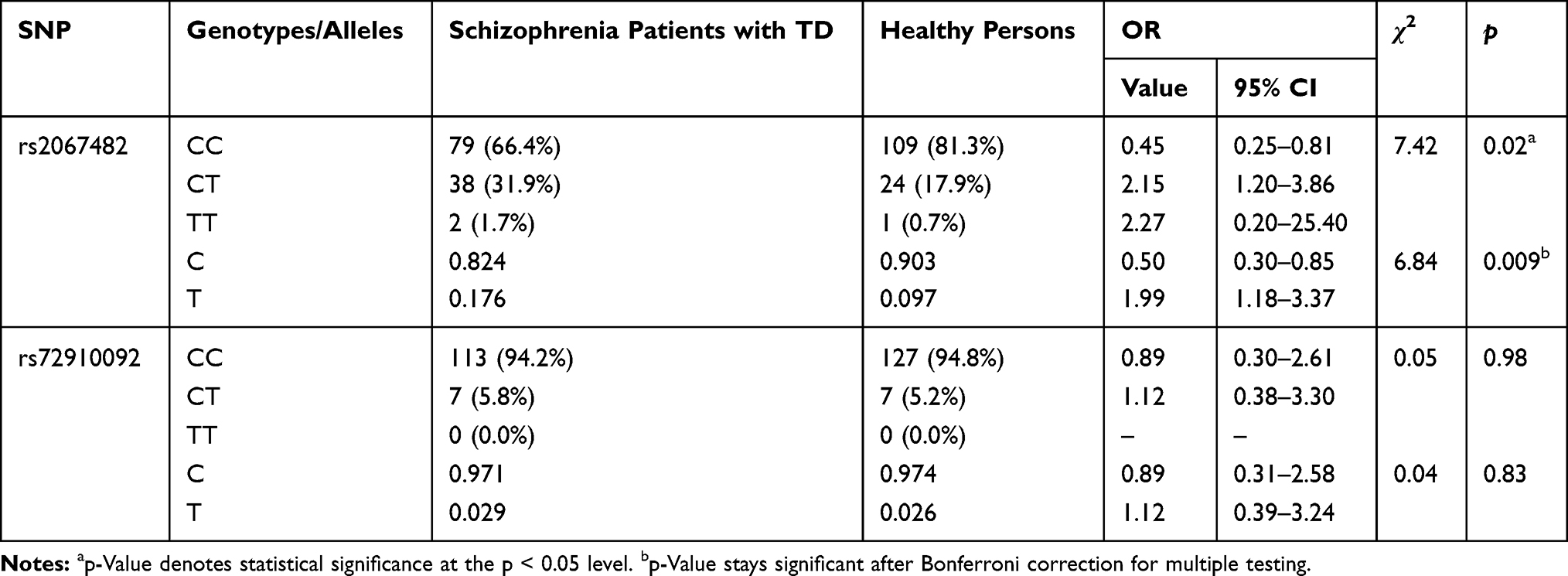 |
Table 6 CHRM4 Genotype Distribution in Schizophrenia Patients with Tardive Dyskinesia and Healthy Persons |
Our results could be interpreted in favor of reducing risk of schizophrenia for carriers of CC genotype and increasing risk of disease for carriers of CT genotype respectively (OR=0.45, 95% CI [0.25–0.81]; OR=2.15, 95% CI [1.20–3.86]). Certainly, we have an association in allelotype (χ2=6.84, p=0.009) and can conclude that allele C has a possible effect against schizophrenia (OR=0.50, 95% CI [0.30–0.85] whereas allele T has a predisposing effect (OR=1.99, 95% CI [1.18–3.37]) towards the disease.
To account for multiple testing, Bonferroni correction was applied to the resulting p values. The findings in Table 2 stay significant, while in Table 6, only the p value allelic frequency remains significant.
Discussion
Previously, Scarr et al have found a reduced level of expression of CHRM4 but not of CHRM1 in the hippocampus of schizophrenia patients.7 Thereafter, they reported the synonymous polymorphism rs2067482 in CHRM4 to be associated with an increased risk of schizophrenia.16 In the current study, we have confirmed an association of the rs2067482 variant with schizophrenia in 449 patients versus 134 healthy subjects. No difference in genotype was detected between patients with and without TD. Surprisingly, the results of the allelic association of the rs2067482 polymorphism with the disease were opposite to the ones reported earlier: Scarr et al found allele C being associated with increased risk of schizophrenia.16 In contrast, our study identified allele T having higher prevalence in schizophrenia patients (Table 2). It is hard to explain such inconsistency but several reasons can be discussed. First, our patient population is markedly different from the one Scarr et al studied. Both populations belong to different ethnical groups (White Russian vs Australian). Secondly, no data is available concerning diagnostic subtyping and mental co-morbidity of their patients which might affect the selection of the particular individual to the patient group. Moreover, many patients (exactly half of them in their 2013 study)16 committed suicide, while none of the individuals in our study had suicidal tendencies, pointing out to different subtypes/severity of the disease. Thus, CHRM4 may be involved in anti-suicidal effects and not in schizophrenia in general. The atypical antipsychotic drug clozapine has specific activity in preventing suicide attempts in high-risk patients with schizophrenia or schizoaffective disorder.23 Clozapine is well known to act as a partial agonist at the CHRM4.24 Moreover, many findings in human and animal studies indicate an association between cholinergic hyperactivity and bipolar depression.25,26 The same mechanism, which in theory might involve insufficient activity of inhibitory CHRM4, might also induce suicidality in patients with schizophrenia. Therefore, we believe that the findings of Scarr et al might be related to the specific deceased patient sample they studied. Furthermore, considering that schizophrenia affects less than 1% of the population, it seems quite unlikely that the common allele with the frequency of 85–95% in the population can be associated with the disease, as found by Scarr et al.16
We also investigated a possible relationship of rs2067482 with TD and did not identify any significant connection. Dyskinesia could be induced by malfunctioning of CHRM4 in the dorsal striatum (particularly putamen).27 However, as adaptation of the activity of direct and indirect extrapyramidal pathways is one of the major functions to regulate motor activity, a genetically induced CHRM4 functionality could be compensated during development. Nevertheless, the relatively high level of expression of CHRM4 in ventral and dorsal striatum in comparison to amygdala and hippocampus,14 makes it difficult to attribute much importance to dysfunction of CHRM4 within the latter region as suggested by Scarr et al.7 A well-known theory holds that in schizophrenia delusions are caused by aberrant salience (function of the amygdala) resulting from dopaminergic hyperactivity (function of the midbrain).28,29 We have argued that the primary mechanism could also be the other way around meaning that dysfunction of amygdala and hippocampus results in activation of monoaminergic centers of the midbrain which activate circuits regulating pleasure and happiness.30 This would place the suggestion by Scarr et al7 about altered CHRM4 expression in hippocampus of patients with schizophrenia in another perspective. Further study of the role of CHRM4 for the activity of fibres originating within amygdala and hippocampus and projecting to ventral striatum31 and/or via the habenula to monoaminergic midbrain centers32 is of potential interest to explain delusional symptoms of schizophrenia.30
Scarr et al did not report finding any de novo mutations in the CHRM4 gene neither in patients nor in controls.16 Out of eight common variants reported for the gene, they only found two (rs2067482 and rs72910092), and only the first one was shown to be associated with schizophrenia (albeit at the border of significance). However, although there is enough evidence that the CHRM4 gene itself might be involved in schizophrenia, no direct link exists between rs2067482 and the gene. Polymorphism rs2067482 does not cause amino acid change in the protein sequence but the option still remains that it has a regulatory effect on gene expression. GTEx portal search (gtexportal.org) reveals 300eQTL SNPs associated with CHRM4 expression. Variant rs72910092 does not appear in that list as well as it is not a significant eQTL for any other gene. In contrast, rs2067482 is found to be a significant eQTL for CHRM4 (P value 1.4 x 10−4) in skin tissue. In addition, rs2067482 is also found to be eQTL for 10 more protein coding genes, namely AMBRA1, ARFGAP2, ARHGAP1, ATG13, C11orf49, DDB2, LRP4, MADD, MDK and PACSIN3 (PACSIN3 is the only gene with association found in brain cerebellum). Therefore, it is possible that the association of rs2067482 with schizophrenia is via its regulatory effect on CHRM4 and/or another gene. In the original GWA study, which identified chr11p11.2 locus association with schizophrenia,4 four highly correlated SNPs (rs7112229, rs11819869, rs7130141 and rs12575668) were found. In GTEx, these SNPs are significant eQTLs for the genes ACP2, AMBRA1, ARFGAP2, ARHGAP1, ATG13, C11orf49, CHRM4, DDB2, LRP4, MADD, MDK and PACSIN3. Of those, only one association is found in brain, namely PACSIN3 (in cerebellum). Interestingly, linkage disequilibrium analysis with LDlink (ldlink.nci.nih.gov) shows that rs2067482 and four schizophrenia variants from GWAS are not highly linked (r2 0.35 to 0.57), suggesting independent regulation of the target genes. Moreover, this analysis identifies PACSIN3 as the common gene regulated by all (described above) genomic variants in the brain. PACSIN3 is a member of pacsins family of cytoplasmic adapter proteins. Three members of the family are found to date, with high sequence conservation, and involved in endocytosis33 and actin dynamics.34 PACSIN3 is not a well-studied gene; it plays a key role in the embryonic morphogenesis that has been demonstrated in the PACSIN3 knockout zebrafish model.35 Pacsin1 knock-out in mice results, among other defects, in tonic-clonic seizures, suggesting pacsins involvement in brain functioning.36
According to the Gene network constructed through the online resource for research GeneMANIA (Figure S1) PACSIN3 is interconnected with CHRM4 via EEF1A2. Eukaryotic elongation factor 1, alpha-2 (eEF1A2) protein is involved in protein synthesis, suppression of apoptosis, and regulation of actin function and cytoskeletal structure. EEF1A2 gene is highly expressed in the central nervous system and EEF1A2 knockout mice show neuronal degeneration.37 Mutations in EEF1A2 have been recently shown to cause epileptic encephalopathy associated with neurodevelopmental disorders such as intellectual disability, autistic spectrum disorder, hypotonia and dysmorphic facial features.38
Our study has three limitations. 1) The sample size of the control group is not large enough although the minor allele frequency (MAF) for rs2067482 in our control group is 9.7%, which is close to MAF 11% according to the 1000 Genomes Project; 2) Despite the fact that we tried to select individuals of only Caucasian background, in the population of the given region of Siberia a national admixture driving these results is possible; 3) It is possible that rs2067482 is in high linkage disequilibrium with another SNP that does increase the risk of schizophrenia. This is quite common especially when several studies give conflicting results, as it is appears to be the case in current study. In prospect, it would have been helpful to evaluate the entire LD region surrounding CHRM4 and test the tagging SNPs in the LD region.
Conclusion
To sum up, our findings confirm association of rs2067482 polymorphism with schizophrenia. However, current data does not support the hypothesis that this link is via the regulation of the CHRM4 gene. Rather than CHRM4, combined evidence from our and previous studies suggests PACSIN3 might be regulated by rs2067482 and identifies PACSIN3 as a novel candidate gene for schizophrenia. There are no de novo mutations in CHRM4 in schizophrenia patients and all regulatory variants, which were described earlier, point to different target genes. In addition, we can speculate that decreased level of CHRM4 in schizophrenic patients was a consequence, but not a cause of illness. There is enough evidence that the reduction in CHRM4 gene expression contributes to the course of the disease, it is however unlikely that CHRM4 is a causative gene. Nevertheless, our data demonstrate the link between the CHRM4 locus and schizophrenia, and indicate a need for further research on the role of this gene in the pathogenesis of the disease.
Acknowledgments
This work resulted from a collaboration between the Mental Health Research Institute in Tomsk and the Groningen Research Institute of Pharmacy (GRIP) of the University of Groningen. The Russian part is carried out within the framework of the Tomsk Polytechnic University Competitiveness Enhancement Program, which did not provide financial assistance for research.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kay SR. Significance of the positive-negative distinction in schizophrenia. Schizophr Bull. 1990;16:635–652.
2. Millan MJ, Fone K, Steckler T, Horan WP. Negative symptoms of schizophrenia: clinical characteristics, pathophysiological substrates, experimental models and prospects for improved treatment. Eur Neuropsychopharmacol. 2014;24:645–692. doi:10.1016/j.euroneuro.2014.03.008
3. Giegling I, Hosak L, Mössner R, et al. Genetics of schizophrenia: a consensus paper of the WFSBP task force on genetics. World J Biol Psychiatry. 2017;18:492–505. doi:10.1080/15622975.2016.1268715
4. Rietschel M, Mattheisen M, Degenhardt F, et al. Association between genetic variation in a region on chromosome 11 and schizophrenia in large samples from Europe. Mol Psychiatry. 2012;17:906–917. doi:10.1038/mp.2011.80
5. Felder CC, Porter AC, Skillman TL, et al. Elucidating the role of muscarinic receptors in psychosis. Life Sci. 2001;68:2605–2613. doi:10.1016/S0024-3205(01)01059-1
6. Crook JM, Tomaskovic-Crook E, Copolov DL, Dean B. Decreased muscarinic receptor binding in subjects with schizophrenia: a study of the human hippocampal formation. Biol Psychiatry. 2000;48:381–388. doi:10.1016/S0006-3223(00)00918-5
7. Scarr E, Sundram S, Keriakous D, Dean B. Altered hippocampal muscarinic M4, but not M1, receptor expression from subjects with schizophrenia. Biol Psychiatry. 2007;61:1161–1170. doi:10.1016/j.biopsych.2006.08.050
8. Haga T. Molecular properties of muscarinic acetylcholine receptors. Proc Jpn Acad Ser B Phys Biol Sci. 2013;2013(89):226–256. doi:10.2183/pjab.89.226
9. Bonner TI. The molecular basis of muscarinic receptor diversity. Trends Neurosci. 1989;12:148–151. doi:10.1016/0166-2236(89)90054-4
10. Michel MC, Teitsma CA. Polymorphisms in human muscarinic receptor subtype genes. Handb Exp Pharmacol. 2012;208:49–59.
11. Lin HH. G-protein-coupled receptors and their (Bio) chemical significance win 2012 nobel prize in chemistry. Biomed J. 2013;36:118–124. doi:10.4103/2319-4170.113233
12. Stevens RC, Cherezov V, Katritch V, et al. The GPCR network: a large-scale collaboration to determine human GPCR structure and function. Nat Rev Drug Discov. 2013;12:25–34. doi:10.1038/nrd3859
13. Goldberg JA, Ding JB, Surmeier DJ. Muscarinic modulation of striatal function and circuitry. Handb Exp Pharmacol. 2012;208:223–241.
14. Lebois EP, Thorn C, Edgerton JR, Popiolek M, Xi S. Muscarinic receptor subtype distribution in the central nervous system and relevance to aging and Alzheimer’s disease. Neuropharmacology. 2018;136(Pt C):362–373. doi:10.1016/j.neuropharm.2017.11.018
15. Santiago MP, Potter LT. Biotinylated m4-toxin demonstrates more M4 muscarinic receptor protein on direct than indirect striatal projection neurons. Brain Res. 2001;894:12–20. doi:10.1016/S0006-8993(00)03170-X
16. Scarr E, Um JY, Cowie TF, Dean B. Cholinergic muscarinic M4 receptor gene polymorphisms: a potential risk factor and pharmacogenomic marker for schizophrenia. Schizophr Res. 2013;146:279–284. doi:10.1016/j.schres.2013.01.023
17. Boiko AS, Ivanova SA, Pozhidaev IV, et al. Pharmacogenetics of tardive dyskinesia in schizophrenia: the role of CHRM1 and CHRM2 muscarinic receptors. World J Biol Psychiatry. 2019;9:1–6. doi:10.1080/15622975.2018.1548780
18. Loonen AJM, van Praag HM. Measuring movement disorders in antipsychotic drug trials: the need to define a new standard. J Clin Psychopharmacol. 2007;27:423–430. doi:10.1097/jcp.0b013e31814f1105
19. Loonen AJM, Doorschot CH, van Hemert DA, Oostelbos MC, Sijben AE. The Schedule for the Assessment of Drug-Induced Movement Disorders (SADIMoD): test-retest reliability and concurrent validity. Int J Neuropsychopharmcol. 2000;3:285–296. doi:10.1017/S1461145700002066
20. Loonen AJM, Doorschot CH, van Hemert DA, Oostelbos MC, Sijben AE. The schedule for the assessment of drug-induced movement disorders (SADIMoD): inter-rater reliability and construct validity. Int J Neuropsychopharmacol. 2001;4:347–360. doi:10.1017/S1461145701002589
21. Schooler NR, Kane JM. Research diagnoses for tardive dyskinesia. Arch Gen Psychiatry. 1982;39:486–487. doi:10.1001/archpsyc.1982.04290040080014
22. Andreasen NC, Pressler M, Nopoulos P, Miller D, Ho BC. Antipsychotic dose equivalents and dose-years: a standardized method for comparing exposure to different drugs. Biol Psychiatry. 2010;67:255–262. doi:10.1016/j.biopsych.2009.08.040
23. Meltzer HY, Alphs L, Green AI, et al. Clozapine treatment for suicidality in schizophrenia: international suicide prevention Trial (InterSePT). Arch Gen Psychiatry. 2003;60:82–91. doi:10.1001/archpsyc.60.1.82
24. Michal P, Lysíková M, El-Fakahany EE, Tucek S. Clozapine interaction with the M2 and M4 subtypes of muscarinic receptors. Eur J Pharmacol. 1999;376:119–125. doi:10.1016/S0014-2999(99)00341-6
25. Van Enkhuizen J, Janowsky DS, Olivier B, et al. The catecholaminergic-cholinergic balance hypothesis of bipolar disorder revisited. Eur J Pharmacol. 2015;753:114–126. doi:10.1016/j.ejphar.2014.05.063
26. Loonen AJM, Kupka RW, Ivanova SA. Circuits regulating pleasure and happiness in bipolar disorder. Front Neural Circuits. 2017;11:35. doi:10.3389/fncir.2017.00035
27. Loonen AJM, Ivanova SA. New insights into the mechanism of drug-induced dyskinesia. CNS Spectr. 2013;18:15–20. doi:10.1017/S1092852912000752
28. Kapur S. Psychosis as a state of aberrant salience: a framework linking biology, phenomenology, and pharmacology in schizophrenia. Am J Psychiatry. 2003;160:13–23. doi:10.1176/appi.ajp.160.1.13
29. Howes OD, Kapur S. The dopamine hypothesis of schizophrenia: version III–the final common pathway. Schizophr Bull. 2009;35:549–562. doi:10.1093/schbul/sbp006
30. Loonen AJM, Ivanova SA. Circuits regulating pleasure and happiness in schizophrenia: the neurobiological mechanism of delusion. In: Shen YC, editor. Schizophrenia Treatment - the New Facets. Rijeka, Croatia: Intech; 2016:109–134.
31. Heilbronner SR, Rodriguez-Romaguera J, Quirk GJ, Groenewegen HJ, Haber SN. Circuit-based corticostriatal homologies between rat and primate. Biol Psychiatry. 2016;80:509–521. doi:10.1016/j.biopsych.2016.05.012
32. Loonen AJM, Ivanova SA. Circuits regulating pleasure and happiness: the evolution of the amygdalar-hippocampal-habenular connectivity in vertebrates. Front Neurosci. 2016;10:539. doi:10.3389/fnins.2016.00539
33. Modregger J, Ritter B, Witter B, Paulsson M, Plomann M. All three PACSIN isoforms bind to endocytic proteins and inhibit endocytosis. J Cell Sci. 2000;113:4511–4521.
34. Sumoy L, Pluvinet R, Andreu N, Estivill X, Escarceller M. PACSIN 3 is a novel SH3 domain cytoplasmic adapter protein of the pacsin-syndapin-FAP52 gene family. Gene. 2001;262:199–205. doi:10.1016/S0378-1119(00)00531-X
35. Edeling MA, Sanker S, Shima T, et al. Structural requirements for PACSIN/Syndapin operation during zebrafish embryonic notochord development. PLoS One. 2009;4(12):e8150. doi:10.1371/journal.pone.0008150
36. Koch D, Spiwoks-Becker I, Sabanov V. Proper synaptic vesicle formation and neuronal network activity critically rely on syndapin I. EMBO J. 2011;30:4955–4969. doi:10.1038/emboj.2011.339
37. Nakajima J, Okamoto N, Tohyama J, et al. De novo EEF1A2 mutations in patients with characteristic facial features, intellectual disability, autistic behaviors and epilepsy. Clin Genet. 2015;87(4):356–361. doi:10.1111/cge.12394
38. De Rinaldis M, Giorda R, Trabacca A. Mild epileptic phenotype associates with de novo eef1a2 mutation: case report and review. Brain Dev. 2020;42(1):77–82. doi:10.1016/j.braindev.2019.08.001
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.