")
Back to Journals » Open Access Rheumatology: Research and Reviews » Volume 6
Animal models of systemic sclerosis: their utility and limitations
Authors Artlett C
Received 25 March 2014
Accepted for publication 29 April 2014
Published 1 July 2014 Volume 2014:6 Pages 65—81
DOI https://doi.org/10.2147/OARRR.S50009
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Carol M Artlett
Department of Microbiology and Immunology, Drexel University College of Medicine, Philadelphia, PA, USA
Abstract: Without doubt, animal models have provided significant insights into our understanding of the rheumatological diseases; however, no model has accurately replicated all aspects of any autoimmune disease. Recent years have seen a plethora of knockouts and transgenics that have contributed to our knowledge of the initiating events of systemic sclerosis, an autoimmune disease. In this review, the focus is on models of systemic sclerosis and how they have progressed our understanding of fibrosis and vasculopathy, and whether they are relevant to the pathogenesis of systemic sclerosis.
Keywords: systemic sclerosis, animal models, vasculopathy, fibrosis, chicken, mouse
Introduction
Systemic sclerosis is an autoimmune disease characterized by fibrosis of the skin and internal organs. It is a rare disease with orphan status in the USA. It has a prevalence of 240 cases per million adults, with an annual incidence of 20 cases per million adults.1 The initiating events leading to systemic sclerosis are currently unknown, so understanding the factors driving this disease is difficult and animal models have therefore provided significant insights into its pathology. The hallmarks of systemic sclerosis are inflammation and autoimmunity, endothelial cell dysfunction leading to widespread vasculopathy, and progressive fibrosis.2 Some of the earliest events in the pathology of systemic sclerosis result in microvascular abnormalities, such as endothelial injury leading to a perivascular infiltrate and neointimal thickening of the vessels. In addition, autoantibodies develop. Taken together, these factors culminate in progressive fibrosis that can affect the skin and internal organs, including the kidney, heart, lung, and gastrointestinal tract.
Systemic sclerosis is further divided into two subsets of disease that describe the pattern of skin fibrosis. Limited cutaneous systemic sclerosis includes a subset of patients with fibrosis limited to the distal extremities and with facial involvement, whereas diffuse cutaneous systemic sclerosis is characterized by fibrosis that is proximal to the knees and elbows. Internal organ involvement can occur in both subsets, and patients with diffuse cutaneous disease more frequently have organ involvement early on in their disease course and these patients have sooner morbidity and mortality than those with limited disease.
The autoantibody profile found in patients with systemic sclerosis defines the subset of disease (diffuse cutaneous or limited cutaneous) and can often predict internal organ involvement. Three dominant mutually exclusive autoantibody profiles are found to be associated with systemic sclerosis.3 These are the topoisomerase-1/Scl70 autoantibody associated with diffuse disease, the centromeric autoantibody associated with limited disease, and the autoantibodies against RNA polymerase I and III that are strongly associated with patients who have the diffuse subset and develop hypertensive renal disease.4
Animal models are important and have been used to study numerous aspects of disease that otherwise cannot be studied at length within the human population. They provide the opportunity to investigate genes or therapeutics without initial risk to humans. Rodents have a physiology and anatomy similar to that of man, and most models employ mice, given that over 95% of the mouse genome is similar to that in humans. Rodents have an accelerated lifespan, are small, require less space to house, and are cost-effective; as a result, a multitude of reagents have been developed for use. Rodent genes can be easily manipulated and genetically engineered for selective tissue expression to replicate human disease more accurately. However, animal models do not always replicate disease faithfully, and in many instances they are predictive such that they exhibit one or two features of a disease, rather than being homologous to the human illness by demonstrating all features of the disease.
Investigators researching systemic sclerosis have often borrowed models from other pathological diseases that exhibit features similar to those of fibrosis; for example, the models that were developed for the study of idiopathic pulmonary fibrosis. However, while idiopathic pulmonary fibrosis may be more faithfully replicated in those models, it often falls short as a model for systemic sclerosis due to the systemic nature of the disease. The systemic nature of systemic sclerosis is probably the most profound difference between this and other fibrosing disorders where usually a specific organ is affected.
Not encompassing of all the models used to study the pathogenesis of systemic sclerosis, this review critically focuses on the benefits and disadvantages of some of the more commonly used models and a number of the recently developed animal models (Table 1). We also discuss the advantages and disadvantages of the avian model of systemic sclerosis.
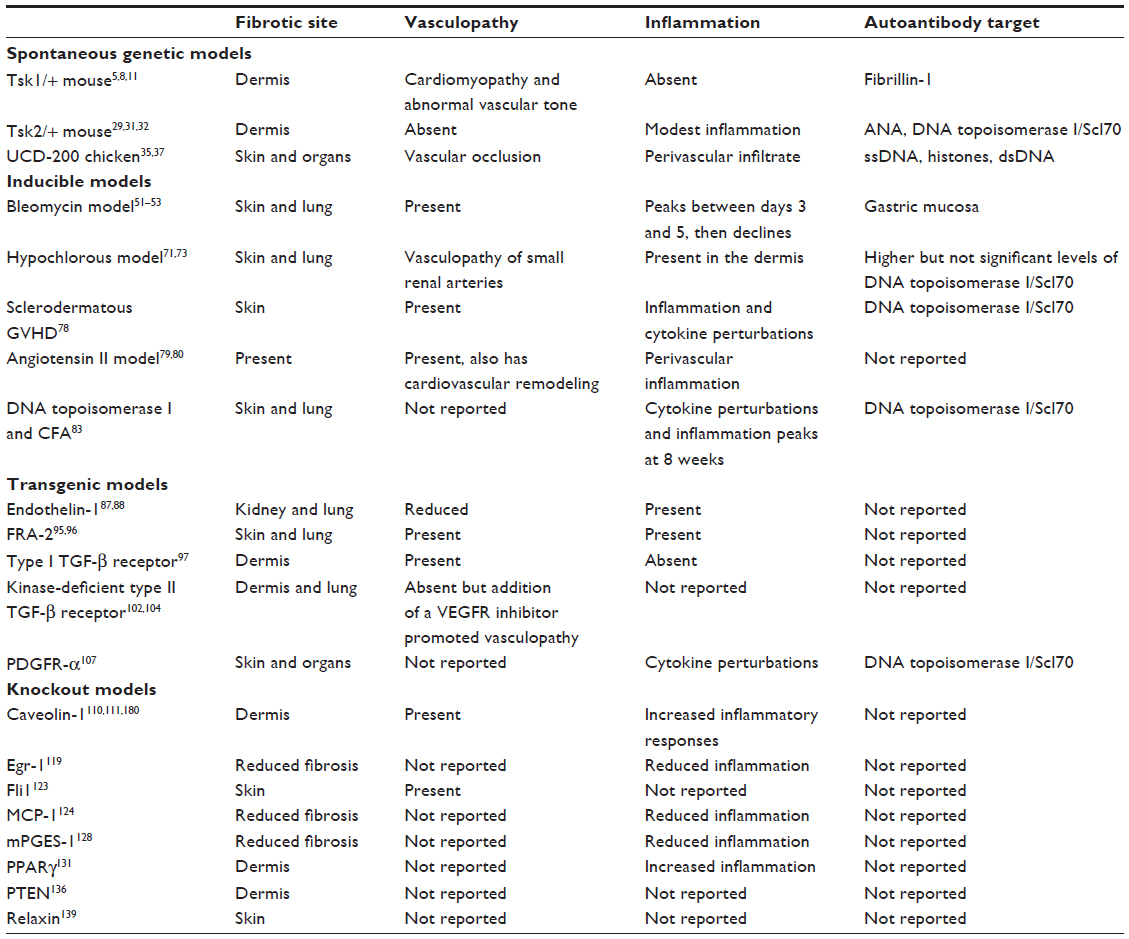 | Table 1 Prominent features of animal models of systemic sclerosis |
Genetic models of systemic sclerosis
There are three models of systemic sclerosis that have arisen by spontaneous mutations to the genome, resulting in increased extracellular matrix deposition. Two spontaneous mutations arose in mice and one in the chicken. All these models have proven to be useful in studying fibrosis and vasculopathy; however, the mutations present in these animals do not appear to be relevant to human disease from a genetic standpoint given that systemic sclerosis is not hereditary.
Tight skin 1 mouse model
This mouse was first reported in 1976 by Green et al,5 occurred as a result of spontaneous mutation in the B10.-D2(58N)/Sn strain, and was detected by a feeling of tethering in the skin in the interscapular region. The mutation is homozygous lethal and mice are bred as heterozygotes. These mice have thickened skin that is firmly bound to the subcutaneous tissue. There is loss of elasticity in the skin, although the mice have increased deposition of elastin.6 Due to the increased elastin, the mouse also displays an emphysema-like pathology in the lung tissue.7 The tight skin 1 (Tsk1/+) mouse has an enlarged heart and has been studied in the context of cardiomyopathy.8 It has also been reported to have an enlarged skeleton, with increased bone and cartilage growth.5 The mutation was found to be a partial duplication of the fibrillin-1 gene on chromosome 2.9 This mutation is thought to sequester increased levels of transforming growth factor (TGF)-β in the extracellular matrix, driving collagen synthesis.10 With identification of this mutation in the Tsk1/+ mouse and the presence of autoantibodies to fibrillin-1,11 clinicians and researchers have tried to correlate this mouse better with systemic sclerosis. Autoantibodies found in patient serum were reported to be directed against fibrillin-1 in a study cohort of Choctaw Native American Indians.12 A follow-up study failed to demonstrate fibrillin-1 autoantibodies in patients of other ethnicities with systemic sclerosis.13 Another study reported an association with a microsatellite marker near the fibrillin-1 gene in Choctaw Indians and Japanese patients with systemic sclerosis,14,15 but this was not reproduced in other cohorts.16,17 Further studies in systemic sclerosis have shown that the fibroblasts are able to assemble fibrillin-1 microfibrils but these fibrils are unstable, suggesting that there is an inherent defect in fibrillin-1 in systemic sclerosis.18
More recent analyses of fibrillin-1 have suggested that the mutations associated with this protein are correlated with stiff skin syndrome, which is an autosomal dominant congenital form of systemic sclerosis. Stiff skin syndrome is characterized by increased synthesis of fibrillin, elastin, and collagen. Microfibrillar assembly is increased but abnormal, and the microfibrils appear to be shorter. There is also increased activation of the TGF-β pathway, as indicated by the nuclear localization of Smad2 and expression of connective tissue growth factor.19 A mouse model employing the fibrillin-1 missense mutation identified in stiff skin syndrome was found to recapitulate fibrosis in the dermis, and this signaling was via α5β1 and αvβ3 integrins, which was driven by TGF-β.20 Intriguingly, the stiff skin model had autoantibodies toward topoisomerase I.20 Inflammation in the skin was also prominent and there was enrichment of plasmacytoid dendritic cells and Th2, Th17, and Th9 CD4 subpopulations.20 The authors argued that the abnormal fibrillin-1 results in microfibrillar aggregates that fail to make adequate contact with cells but are able to retain the ability to sequester latent TGF-β, which becomes activated due to stiffness in the extracellular matrix resulting in a feed-forward signaling mechanism to drive fibrosis.20 Integrins are known to sense the extracellular environment, and by that sensing are able to dictate the response of the fibroblast.21–24 The stiff skin syndrome model hints at dysregulated signaling mechanisms driven by integrins that may be relevant for the pathology of systemic sclerosis; however, it does not address the etiology of systemic sclerosis because these fibrillin-1 mutations are not identified in the disease.
The Tsk1/+ mouse has been used to study the efficacy of various drugs for the treatment of fibrosis, and the model has proved valuable in this regard. Imatinib is a small-molecule tyrosine kinase inhibitor that is able to inhibit both TGF-β and platelet-derived growth factor (PDGF) pathways. Imatinib prevents the development of inflammation-driven experimental fibrosis mediated by the drug bleomycin. Furthermore, imatinib reduced dermal thickening in the Tsk1/+ model,25 suggesting the involvement of TGF-β and PDGF in the development of fibrosis in this model. Interferon-gamma therapy administered by nanoparticles also successfully reduced skin thickness in the Tsk1/+ mouse,26 and edaravone, a free radical scavenger, decreased collagen synthesis,27 as did rapamycin.28
Tight skin 2 mouse model
The tight skin 2 (Tsk2/+) mouse was first described in 1986, and arose in the offspring of a 101/H mouse exposed to the mutagenic agent ethylnitrosourea. As with Tsk1/+, tight skin is found in the interscapular region.29 The gene mutation is located on chromosome 1 between 42.5 and 52.5 Mb,30 and to date the genetic defect has not been published. Like Tsk1/+, Tsk2/+ traits are highly penetrant and are homozygous embryonic lethal. Tsk2/+ mice replicate many features of systemic sclerosis, including tight skin, increased deposition of dermal extracellular matrix, and autoantibodies.31,32 Tsk2/+ shows increased transcription rates of type I and III collagen in dermal fibroblasts,33 suggesting that the increased collagen synthesis is cell autonomous rather than dependent on the integrity of the skin as a whole. Tsk2/+ mice were also reported to have increased autoimmunity, and a majority of the mice developed multiple autoantibodies specific for systemic sclerosis, including Scl70, centromere, and DNA.32 A restricted T-cell receptor bias was noted in skin infiltrates that encompasses specific T-cell receptor subtypes.34 Until the genetic mutation has been identified, the value of this model cannot be fully realized. Another caveat to this model is that few laboratories choose to study it, possibly because the gene mutation is yet to be reported.
UCD-200 chicken
University of California at Davis line 200 (UCD-200) chickens develop a hereditary connective tissue disease that is similar in many ways to systemic sclerosis. It is characterized by a perivascular lymphocytic infiltration,35 endothelial injury resulting in vascular occlusion,36 speckled antinuclear antibodies,37,38 and fibrosis of the skin and visceral organs.37 During experiments to map the genetic mutation in UCD-200 chickens crossed with red jungle fowl chickens, it was noted that the F1 generation were disease-free, whereas 36% of the back-crossed population (F2 generation) had early onset avian systemic sclerosis and 61% had late onset avian systemic sclerosis. The incidence of avian systemic sclerosis was significantly higher in male chickens than in female chickens.39 This suggests that the genetic defect in the UCD-200 line has an autosomal recessive mode of inheritance, or is comprised of multiple genetic loci or multiple loci interactions that are required to drive disease.39 Five genes have been identified that confer a high association with the disease, and include TGFBR1, IGFBP3, EXOC2/IRF4, CCR8, and SOCS1. All of the genes are located on chromosome 2, with the exception of SOCS1, which is located on chromosome 14. All of these genes have been reported to have significance in the pathology of systemic sclerosis or to play a role in autoimmunity.40–44 In addition, COL1A2 was found to be located in close proximity to the quantitative trait loci on chromosome 2, suggesting that this gene may play a role in avian systemic sclerosis. A shorter mRNA transcript has been identified in the early inflammatory stage in the comb and esophagus of diseased chickens.39
Of the three known spontaneous genetic animal models, this model probably represents systemic sclerosis the closest in terms of its pathological features. Especially important to systemic sclerosis and the UCD-200 model is the perivascular infiltrate that precedes vasculopathy. Inflammation of the skin occurs early on in the disease, and this is predominantly found in the comb, which becomes swollen and erythematous. The vasculopathy appears to be driven by endothelial apoptosis.36,38 Histology of the comb demonstrates that there is vessel occlusion with neointimal formation and a perivascular inflammatory infiltrate.36 The model also has internal organ involvement. Autoantigens present in the sera include single-stranded DNA, double-stranded DNA, histones, poly(G), and cardiolipin; however, they did not reflect the antinuclear antibody spectrum that is observed in diffuse systemic sclerosis.45 The caveats to this model are that few animal facilities are equipped to house these chickens, which require significantly more room than mice and have to be housed separately. Few laboratories have studied this model because most researchers prefer smaller animals with greater reagent resources.
Inducible models of fibrosis
Animal models predicated on inducing fibrosis in the mouse are of great value because they allow for study of the initiating events in fibrosis. These models have proven to be very useful. However, induction of fibrosis in animals is limited to a small number of chemicals, and sclerodermatous graft versus host disease (GVHD) is difficult to work with.
Bleomycin model of fibrosis
The bleomycin mouse model of fibrosis is commonly used to replicate systemic sclerosis, or dermal or pulmonary fibrosis. Bleomycin is a glycopeptide antibiotic produced by Streptomyces verticillus and has been used to treat Hodgkin’s lymphoma, squamous cell carcinoma, and testicular cancer, and is usually used in combination with other antineoplastic drugs.46 It has also been used to treat plantar warts.47 However, use of bleomycin is not without significant risks, and a systemic sclerosis-like syndrome can ensue in susceptible individuals.48–50 This observation paved the way for development of the bleomycin mouse model of fibrosis by Yamamoto et al.51 Local dermal injections of bleomycin in mice induced collagen synthesis at the injection site over 4 weeks. The overall effects were found to be systemic because the lung also showed increased collagen synthesis.51 The sensitivity to bleomycin was found to be strain-specific, and skin thickening was greater in some strains than in others. Yamamoto et al52 found that fibrosis was induced by approximately two-fold in C3H/He, DBA/2, B10.D2, and B10.A strains, and that dermal thickness was increased by more than 2.5-fold in A/J, C3H/He, B10.A, and B10.D2 strains. One of the initiating events in bleomycin-induced fibrosis is mediated by inflammation. During the first 3–5 days of injections, there is a peak in the skin inflammatory infiltrate which then decreases with extended treatment. Bleomycin has been reported to induce vascular changes in the skin at the injection site.53 Upon withdrawal of bleomycin, the fibrosis resolves over time. Investigators usually terminate the study prior to resolution, but this model could be utilized in study pathways involved in the resolution of fibrosis. In a similar manner, skin involved in systemic sclerosis appears to soften during the course of the disease.54 The model replicates some of the most important biomarkers of systemic sclerosis but is by its very nature a model with limited uses.
There are several modes for administration of bleomycin that have been used to induce fibrosis depending on the organ to be investigated. As described above, bleomycin can be injected subdermally to induce synthesis of collagen; however, this is labor-intensive and has to be performed daily for 3–4 weeks for a robust fibrotic lesion to develop. In addition to the skin becoming fibrotic, this method also causes lung fibrosis.55 A single administration of bleomycin via intratracheal instillation in weight-adjusted dosages is frequently used to study pulmonary fibrosis. The fibrotic stage exists until 3–4 weeks post-administration.56 There can be a poorer survival rate depending on the strain of mouse used.57 Finally, bleomycin can be administered via Alzet® osmotic pumps (DURECT Corporation, Cupertino, CA, USA) that are embedded under the skin and deliver a constant amount of compound over a certain number of days, making this approach to administration convenient on many levels. When used with bleomycin, the mortality observed in mice is significantly less than with intratracheal instillation.58 Further, this method induces a pattern of interstitial lung disease that replicates the lung pathology of systemic sclerosis and produces fibrosis in the skin and internal organs also in a pattern that is similar to systemic sclerosis.58
Bleomycin activates the NLRP3 inflammasome, leading to downstream signaling events in fibroblasts and resulting in synthesis of collagen.55 Many of the signals that result in activation of the NLRP3 inflammasome are still being elucidated. Reactive oxygen species, oxidative stress, and the mitochondria are integrally involved in activation of the NLRP3 inflammasome.59,60 Bleomycin induces oxidative stress61,62 by activating the mitochondria, and it is this induction of oxidative stress that activates the inflammasome. Further evidence in support of this finding is that cells depleted of mitochondria are resistant to bleomycin-induced activation.63 To further underscore the importance of reactive oxygen species and oxidative stress in this model, fibrosis was found to be abrogated when N-acetylcysteine was administered.64 The culmination of inflammasome assembly and activation results in cleavage of caspase-1, which is then able to cleave interleukin (IL)-1β and IL-18 for secretion, leading to fibrosis.55 IL-1β and IL-18 were found to be elevated in systemic sclerosis serum65,66 and it was not until our laboratory specifically studied the involvement of the inflammasome in the dermal fibrosis component of systemic sclerosis that the association between an activated inflammasome, fibrosis, and systemic sclerosis was found.55 Further, we found that activated caspase-1 was driving the increased collagen synthesis and myofibroblast phenotype.55
Although the bleomycin model replicates some of the earliest signaling patterns observed in systemic sclerosis, the model does not present with the profound autoantibody patterns typical of systemic sclerosis, but the mouse can establish an autoantibody that cross-reacts with the gastric mucosa.67
Hypochlorous model of systemic sclerosis
Reactive oxygen species have been implicated in the pathogenesis of systemic sclerosis, and systemic sclerosis monocytes68 and fibroblasts69 release increased amounts of reactive oxygen species, which triggers synthesis of collagen. The endoplasmic reticulum itself can induce reactive oxygen species and can also induce mitochondrial-reactive oxygen species,70 suggesting that increased reactive oxygen species may play a central role in the fibrosis component of systemic sclerosis.
The hypochlorous mouse model is a newly developed inducible animal model, and provides further important insights into the initiating signals derived from the reactive oxygen species that promote fibrosis. The hypochlorous mouse model is induced by repeated intradermal injections of hypochlorous acid, which causes generation of hydroxyl radicals leading to increased synthesis of collagen in the skin and lung tissues. In addition, this model mimics the pathological changes observed in the systemic sclerosis kidney and induces anti-topoisomerase antibodies.71 The downside of this model at the moment is that it is currently only used by two laboratories to study the initiating events of fibrosis,72–74 and it is not fully understood how hypochlorous acid induces fibrosis. It has been shown that overproduction of reactive oxygen species in this model activates ADAM17, which is involved in NOTCH signaling.73 However, this model, like the bleomycin model, has proved to be useful in the testing of various candidate molecules for the prevention of fibrosis.73,74
Sclerodermatous GVHD model
This model has been used to understand further some of the initiating factors that lead to systemic sclerosis, and is derived from the initial observations of dermal fibrosis induced in patients with chronic GVHD. Thus, this mouse model was developed and then characterized. The model demonstrates increased collagen synthesis as in systemic sclerosis, and the fibrotic changes were found to be driven by TGF-β because inhibition of TGF-β abrogated the progression of fibrosis.75 Unlike systemic sclerosis, mice with sclerodermatous GVHD have a significant inflammatory infiltrate consisting of T-cells and monocytes/macrophages, and the numbers of inflammatory cells in the lesions are directly dependent on TGF-β.76 Patients with systemic sclerosis generally do not present with such an infiltrate, and the few T-cells found in the lesions tend to be perivascular.77
A modified model of GVHD was then developed and found to exhibit all the major aspects of systemic sclerosis. GVHD was developed in the RAG-2 knockout mouse by the transfer of B10.D2 spleen cells.78 GVHD develops over time, and the model shows progressive skin thickening with disease duration. The kidneys and large and small intestines were found to be fibrotic. Vasoconstriction was reported, increased α-smooth muscle actin and endothelin-1 were found in the vessels of the skin and kidney, and anti-Scl70 antibodies were found in the serum. This model characterizes many of the features of systemic sclerosis, and of note, renal crisis was one of the pathologies observed. TGF-β was found to be elevated, and neutralization of TGF-β with antibodies reduced skin thickness and the ensuing vasculopathy.78
Angiotensin II-inducible model of fibrosis
This is a recently developed model that uses angiotensin II to induce vascular constriction, leading to inflammation and fibrosis.79 Using Alzet osmotic pumps, angiotensin II was found to induce dermal fibrosis by activating the TGF-β pathway, and increases were found in TGF-β2 and TGF-β3 but not in TGF-β1 mRNA. Angiotensin II was also found to increase the numbers of myofibroblasts and fibrocytes in the skin.79 Increased numbers of vascular endothelial (VE)-cadherin and fibroblast-specific protein-1-positive cells were observed around the small vessels in this mouse model, suggesting increased endothelial to mesenchymal transition. The model also includes cardiovascular remodeling.80 However, it is unknown whether these mice develop autoantibodies specific for systemic sclerosis.79
The hypochlorous mouse model was used to test the hypothesis that angiotensin II receptor blockade could ameliorate fibrosis. Irbesartan, an angiotensin II receptor inhibitor, was found to reduce dermal thickness and total collagen content, with less inflammation, and to reduce anti-topoisomerase I antibodies.81 This suggests that angiotensin levels can be regulated by oxidative stress and play a critical role in the fibrosis component of systemic sclerosis, and it has been found that a subset of patients have elevated angiotensin II signaling.82
Model induced by DNA topoisomerase I and Freund’s complete adjuvant
Autoimmunity is a significant feature in systemic sclerosis, and yet few investigators have developed models that specifically investigate this specific aspect of its pathology. Yoshizaki et al83 demonstrated that subcutaneous administration of DNA topoisomerase I and Freund’s complete adjuvant for 8 weeks promoted dermal and lung fibrosis. Adjuvant lacking mycobacterial components (Freund’s incomplete adjuvant) did not induce fibrosis. Cytokine perturbations included increased levels of IL-4, interferon-gamma, IL-10, TGF-β, and tumor necrosis factor-alpha.83 Bronchoalveolar lavage fluid was analyzed and the investigators found that the T-cell profile was skewed toward Th2 and Th17 in mice receiving DNA topoisomerase I and Freund’s complete adjuvant, and in the mice receiving DNA topoisomerase I and Freund’s incomplete adjuvant, and in the IL-6 deficient mice it was skewed toward Th1 and regulatory T-cells. This study raises the provocative possibility that an antibody directed against topoisomerase I could promote fibrosis.
Transgenic models of fibrosis
These models are important for understanding the role of the specific genes that contribute to fibrosis. Many of these models have been utilized from investigators outside the field of systemic sclerosis who study other forms of fibrosis. However, this should not detract from the importance of these models and what they tell us about the role of a particular gene in regulating collagen synthesis.
Endothelin-1 mouse model
Damage to endothelial cells causes increased expression of endothelin-1. The effects of overexpression of endothelin-1 were found to be deleterious to the vasculature and to contribute to hypertrophy of the vessels and vascular permeability, leading to vasoconstriction.84 Endothelin-1 also plays a role in increased cell proliferation, fibrosis, and inflammation.85,86 The transgenic mouse develops glomerulosclerosis and interstitial fibrosis.87,88 Patients with systemic sclerosis have increased tissue and circulatory endothelin-1 levels as well as increased vasculopathy, and these findings underscore the importance of this protein in the pathology of systemic sclerosis.89,90
Bosentan is an endothelin-1 antagonist and has been used successfully to treat pulmonary hypertension in systemic sclerosis.91,92 Tsk1/+ mice were found to have impaired aortic relaxation, suggesting a role for endothelin-1,93 and this led to the study of bosentan in these mice.94 With bosentan, there was partial restoration of the relaxation capability of the vessels in Tsk1/+ mice, suggesting that these mice have endothelial dysfunction.94 More specifically, endothelin-1 promoted increased expression of endothelin receptor B, but not endothelin receptor A, in Tsk1/+ mice.
Fos-related antigen-2 mouse model
The transgenic mouse model overexpressing the Fos-related antigen-2 (FRA-2) gene demonstrates many of the important factors resulting in the vasculopathy of systemic sclerosis. The FRA-2 transgenic animal displays microangiopathy in addition to progressive skin fibrosis. Overall, it was found that this pathology was preceded by apoptosis of endothelial cells.95 FRA-2 is a transcription factor activator protein and belongs to the activator protein-1 family. This family is induced by cellular stress, and as a result, controls many of the downstream responses in the cell, such as cell proliferation, apoptosis, inflammation, tumorigenesis, and wound healing. In light of this, it was found that expression of FRA-2 was independent of the expression of major cytokines such as TGF-β1 and PDGF, and both are important cytokines driving the pathology of systemic sclerosis. However, overexpression of FRA-2 is apparent in the skin lesions of systemic sclerosis, suggesting there may be a direct causal involvement in fibrosis95 and that it may also play a role in vasculopathy.96
Type I TGF-β receptor transgenic model
A model was developed to specifically upregulate the type I TGF-β receptor in fibroblasts on a Cre-ER transgenic background. In this model, with progressing age, there were increasing levels of collagen deposition in the skin of the mouse. In addition, the ultrastructure of collagen was found to be disorganized. Vascular changes similar to systemic sclerosis were apparent, with hypertrophy of smooth muscle cells and neointimal thickening of vessel walls. Mice had increased levels of von Willebrand factor, which is similar to what is observed in systemic sclerosis, leading to the hypothesis that sustained activation of TGF-β signaling in fibroblasts can lead to endothelial damage.97 Vascular smooth muscle cells from this mouse demonstrate enhanced remodeling of collagen gel lattices with altered endothelin-1 and endothelin receptor signaling. This model further replicates other features of the pathology of systemic sclerosis in that it has myocardial fibrosis and altered aortic dynamics. As expected, activation of the TGF-β pathway caused increased phosphorylation of Smad2/3 in fibroblasts, leading to increased phosphorylation of ERK1/2 and p38.97 This model suggests that chronic expression of TGF-β in fibroblasts could cause the pathology observed in systemic sclerosis; however, some investigators believe that damage to endothelial cells is one of the leading culprits in initiating and driving the disease.98–100
Kinase-deficient type II TGF-β receptor transgenic model
This model was developed when a fibroblast-specific transcriptional enhancer was identified upstream of the COL1A2 collagen gene.101 The enhancer was used to specifically express a kinase-deficient mutant type II TGF-β receptor that can engage TGF-β but cannot lead to phosphorylation of TGF-β receptor 1 and its downstream signaling pathways.102 The directed expression of the kinase-deficient type II TGF-β receptor resulted in increased fibrosis in the lung and skin. The transgenic fibroblasts were found to have a TGF-β activated phenotype with sustained induced expression of connective tissue growth factor and increased phosphorylation of Smad2/3,102 and increased myofibroblast numbers.103 Although this transgenic model allows for the study of TGF-β signaling, caveats are the lack of immune activation and the fact that autoantibodies are not present. In addition, the transgenic model does not have vasculopathy; however, when a vascular endothelial growth factor receptor inhibitor is administered to these mice, they develop pulmonary arterial hypertension pathology similar to that observed in systemic sclerosis.104
PDGF receptor-α transgenic model of fibrosis
PDGF has been implicated in the development of the fibrosis and vasculopathy seen in systemic sclerosis.105,106 Olson and Soriano investigated the role of the PDGF receptor-α in fibrosis.107 They designed transgenic mice to carry conditionally expressed PDGF receptor-α activating mutations that promoted fibrosis in the skin and internal organs. The importance of PDGF signaling during fibrosis was further recapitulated utilizing sunitinib and sorafenib,108 two tyrosine kinase inhibitors that act to block the bioactivity of PDGF receptors and the vascular endothelial growth factor (VEGF)/VEGF receptor pathway. This study demonstrated that inhibition of phosphorylation of PDGF receptor-β with sunitinib effectively reduced skin and lung fibrosis. Sorafenib was not as effective for reduction of fibrosis; however, it significantly reduced the DNA topoisomerase I autoantibodies, IL-6 and TGF-β.108 As discussed previously, imatinib, a TGF-β and PDGF pathway inhibitor, effectively reduced Tsk1/+ dermal fibrosis,25 adding further credence to the value of tyrosine kinase inhibitors.
Knockout models of fibrosis
These models study the specific contribution of a gene in fibrosis. In some instances, the gene is a negative regulator of collagen synthesis and its deletion promotes fibrosis, whereas other genes are required for induction of the fibrotic process.
Caveolin-1
Caveolin is found to be expressed within the lipid rafts of the caveolae in the cell membranes and functions to modulate the functioning of various signaling molecules.109 It was noted that caveolin-1-deficient mice had increased alveolar septa and lung fibrosis. In addition, these mice were found to have altered vasculopathy due to endothelial proliferation.110 On closer examination of the systemic sclerosis phenotype and the phenotypic changes, these mice were observed to have increased collagen and fibronectin accumulation, in addition to increased numbers of myofibroblasts.111 It is now known that caveolin-1 regulates TGF-β signaling by interacting with the type I TGF-β receptor and regulating Smad signaling pathways by suppression of phosphorylation of Smad2112 and inhibiting ERK signaling.113 However, TGF-β decreases expression of caveolin-1.114 Further, it has been found that loss of caveolin-1 can lead to oxidative stress115 and this is a pathology that has been associated with systemic sclerosis.116,117
Early growth response protein-1 knockout mouse
Early growth response protein-1 (Egr-1) is a transcription factor that is a zinc finger binding protein and is involved in cell growth, differentiation, and survival. It is induced by stress signals.118 Egr-1-deficient mice were injected with bleomycin and skin fibrosis was assessed. There were decreased numbers of infiltrating inflammatory cells in the skin in addition to decreased dermal thickness and collagen expression.119 The signaling events downstream from Egr-1 play a role in collagen fibril formation because mice deficient in this protein have disorganized collagen fibers. Furthermore, bleomycin-induced lung fibrosis was attenuated in the absence of Egr-1, as was the ability to induce wound healing, and the absence of Egr-1 inhibited wound closure in in vitro scratch wound assays.119 In confirmation of the importance of Egr-1 in wound healing, an Egr-1 transgenic mouse was tested and found to enhance wound healing. This further demonstrates the importance of this transcription factor in the synthesis of collagen, wound healing, and fibrosis.
Friend leukemia integration factor-1 conditional knockout mouse
This model recapitulates many of the features of systemic sclerosis in that it has abnormalities of the vasculature.120 Friend leukemia integration factor-1 (Fli1) is constitutively expressed in fibroblasts and plays a pivotal role in inhibiting the expression of extracellular matrix proteins, such as connective tissue growth factor and type I collagen.121 However, more recently, Fli1 was demonstrated to modulate endothelial cell function and to be involved during vascular development. The Fli1 conditional knockout mouse has increased vascular permeability, and this closely mimics the permeability observed in patients with systemic sclerosis due to reduced expression of VE-cadherin.122 Collagen fiber formation is dependent on Fli1123 and this model further mimics the abnormalities in collagen fibril formation in systemic sclerosis, further confirming the importance of Fli1 in the pathogenesis of systemic sclerosis. Phosphorylation of Fli1 and its interaction with p300 transcription factor is also dependent on TGF-β, thus underscoring further the importance of this cytokine in the pathology of systemic sclerosis.
Macrophage chemoattractant protein-1 mouse model
The involvement of basic fibroblast growth factor and connective tissue growth factor in conjunction with macrophage chemoattractant protein-1 (MCP-1) is required for tissue fibrosis.124 MCP-1-deficient mice show abrogated fibrotic responses when basic fibroblast growth factor and connective tissue growth factor are injected into the skin. In addition, this model has reduced macrophage recruitment, fewer mast cells, and fewer CD4+ T-cell numbers.124 Connective tissue growth factor alone does not induce fibrosis, but when administered with basic fibroblast growth factor, there is increased autocrine signaling along with persistent fibrosis. MCP-1, basic fibroblast growth factor, and connective tissue growth factor are proteins that are regulated by TGF-β.125–127 The correlation and interaction between MCP-1, basic fibroblast growth factor, and connective tissue growth factor indicates the importance of the interplay between these three proteins in fibrosis.
Microsomal prostaglandin E2 synthase-1 knockout model
Microsomal prostaglandin E2 synthase-1 (mPGES-1) is an inducible enzymatic protein that acts downstream of cyclo oxygenase. It specifically causes the conversion of prostaglandin H2 to prostaglandin E2. mPGES-1 was found to be overexpressed in systemic sclerosis fibroblasts, suggesting that this pathway is dysregulated.128 Mice deficient in mPGES-1 are resistant to bleomycin-induced fibrosis, have decreased levels of α-smooth muscle actin, and reduced numbers of macrophages,128 further underscoring the importance of this gene in fibrosis.
Peroxisome proliferator-activated receptor-gamma
Peroxisome proliferator-activated receptor-gamma (PPARγ) has been associated with the pathology of numerous diseases, including obesity, diabetes, atherosclerosis, and cancer. More recently, decreased PPARγ has been shown to be associated with fibrosis.129 PPARγ is a nuclear receptor ligand-activated transcription factor that is decreased by TGF-β1 signaling. PPARγ inhibits collagen synthesis,130 whereas TGF-β inhibits PPARγ expression,131 and is thus involved in the fibrotic response in systemic sclerosis. Kapoor et al further explored the role of PPARγ in fibrosis and found that conditional PPARγ-deficient mice developed greater skin thickening with bleomycin when compared with wild-type mice.132 Skin thickening was assessed by analysis of total collagen content, increased α-smooth muscle actin expression, and increased inflammation.132 In confirmation of this finding, rosiglitazone, a PPARγ agonist, was found to decrease fibrosis in the bleomycin mouse model.129 Further, rosiglitazone was also effective in reducing already established fibrosis.129 Inhibition of PPARγ with rosiglitazone and the PPARγ knockout mouse have helped to define the role of this protein in collagen synthesis. It is unknown if this model sees increased vascular remodeling, as it was not reported, but PPARγ is known to play a role in this process.133
PTEN conditional knockout mouse model
Phosphatase and tensin homolog (PTEN) is a protein lipid phosphatase that activates Akt via phosphorylation, and the phosphoinositide 3-kinase-Akt signaling axis has been implicated in the production of collagen.134 Akt phosphorylation is elevated in systemic sclerosis fibroblasts135 and PTEN levels are decreased.136 Regulation of PTEN is dependent on TGF-β; however, PTEN knockout mice have increased collagen deposition and thickened skin, in addition to increased numbers of proliferating cell nuclear antigen-positive fibroblasts, increased numbers of α-smooth muscle actin-bearing fibroblasts, and increased numbers of connective tissue growth factor-positive fibroblasts.136 The data from this mouse model suggest that PTEN normally suppresses fibrogenesis and its absence spontaneously promotes collagen synthesis.
Relaxin knockout mouse
Relaxin is a small dimeric peptide that is generally associated with the female reproductive tract, and is known to be a vasodilator but also has antifibrotic effects.137,138 Mice deficient in relaxin were found to have increased collagen deposition in the skin over time. Administration of human recombinant relaxin via osmotic minipumps abrogated fibrosis in this model.139 This knockout mouse has been used to study other fibrotic diseases, such as liver fibrosis due to administration of carbon tetrachloride,140 and cardiac fibrosis,141 and silica-induced pulmonary fibrosis.142 In all cases, relaxin was found to ameliorate collagen synthesis. This suggests that relaxin regulates collagen synthesis and could be used as a therapy for fibrotic diseases.
Using mouse models for preclinical testing
There are no formal guidance documents from the US Food and Drug Administration (FDA) requiring the use of animal models of efficacy for preclinical studies of a particular drug candidate prior to human clinical testing. However, the FDA does require various toxicological and pharmacological safety studies to ensure that the drug candidate and the final formulated drug product is reasonably safe before proceeding to human clinical testing. These toxicological and pharmacological safety studies are usually performed in rodent and canine species, but may include other species (porcine for example) if physiologically relevant to the drug candidate being tested or therapeutic area being targeted. Certainly from this perspective, a model replicating all or some of the aspects of disease is not required. However, in light of this, the efficacy of a potential drug candidate in preclinical studies has usually been required by pharmaceutical companies prior to investment of significant financial resources in clinical trials and in bringing the product to market. In addition, animal models have been used in proof-of-concept studies to investigate the role of a particular protein or pathway, or to test inhibitors or reagents identified more often than not at the coal face by academic researchers. Even so, success in an animal model has not always translated into a successful treatment for the patient (Table 2). Due to the polymorphic nature of the disease, until the cause (or causes) of systemic sclerosis are identified, a drug that treats all symptoms of the disease may not be realistic. Indeed, it may be that multiple treatment regimens are required to manage symptoms.
| Table 2 Preclinical testing of drugs in mice and their outcome in systemic sclerosis |
Anakinra is an IL-1R antagonist and has shown efficacy in treating IL-1-related diseases that are driven by inflammasome activation. Anakinra was effective at blocking bleomycin-induced pulmonary fibrosis,143 and clinical trials are currently underway to determine whether it will be effective in systemic sclerosis. Systemic sclerosis was not originally thought of as an IL-1-mediated disease until recently;55 however, IL-1 expression can drive TGF-β signaling, and inhibition of this innate immune signaling pathway may also lead to inhibition of fibrosis in systemic sclerosis. Rilonacept is another drug that interferes with IL-1 signaling. This drug was never studied in preclinical models; however, a clinical trial is currently underway in patients with systemic sclerosis although no results are available.
Several immunosuppressants have been tested in preclinical models and in systemic sclerosis clinical trials with mixed success. Cyclosporine demonstrated some efficacy in preventing bleomycin-induced lung fibrosis;144,145 however, although some patients with systemic sclerosis tolerated the drug,146 there is a significant risk of renal toxicity.147 Likewise, prednisone can promote renal toxicity in patients and their renal function has to be closely monitored.148
Mycophenolate mofetil is an immunosuppressant that prevents the purine biosynthesis necessary for development of T-cells and B-cells. It is currently used to treat other autoimmune diseases. This drug successfully prevented bleomycin-induced fibrosis149 and was successful in a small study of patients with systemic sclerosis;150 however, larger studies are currently underway and it is currently unknown as to whether this drug will hold up to close scrutiny.
Thalidomide has emerged as an immunomodulating therapeutic and is thought to modulate the Th1/Th2 profile by inhibiting tumor necrosis factor-alpha, IL-6, IL-10, and IL-12, and by enhancing the production of IL-2, IL-4, and IL-5 by immune cells.151 It has also been reported to increase the lymphocyte count, induce costimulation of T-cells, and alter the cytotoxicity of natural killer cells.151 Thalidomide and its analogs could abrogate fibrosis in mouse models, paving the way for this drug to be trialed in patients. In a small pilot study of patients with systemic sclerosis, thalidomide was found to enhance the healing of digital ulcers.152 However, to date, there has been some difficulty in recruiting patients for a long-term clinical trial and the study was terminated.
Colchicine, a drug that inhibits mitosis, was demonstrated to afford some protection against bleomycin-induced pulmonary fibrosis.153 However, when used to treat systemic sclerosis, there was mixed success in the clinical trials.154,155 Inhibition of tumor necrosis factor-alpha by etanercept was effective in treating bleomycin-induced dermal fibrosis;156 however, when trialed in systemic sclerosis, the drug appeared to effectively reduce the inflammatory joint disease seen in these patients157 but did not appear to be efficacious against fibrosis,158 and is not without risk for fatal fibrosing alveolitis.159
Imatinib, a tyrosine kinase inhibitor, was initially promising as a therapeutic and was found to reduce the fibrosis in the Tsk1/+ mouse and the inducible bleomycin model;25 however, when used in patients, adverse events were frequently observed and tolerability was poor.160 Iloprost, a prostacyclin agonist, also effectively reduced bleomycin-induced fibrosis;161 however, there were heterogeneous responses in small patient cohorts and this has made the efficacy of iloprost hard to assess.162,163
Halofuginone has been shown to interfere with collagen synthesis. It does so by inhibiting TGF-β signaling through a process that prevents phosphorylation of Smad3.164 The drug has been used in Tsk1/+ mice and in mice with chronic GVHD, and both models demonstrated a significant reduction in fibrosis.165 However, when halofuginone was used topically in a small pilot study on 12 patients with systemic sclerosis, five patients responded by demonstrating a significant reduction in skin score whereas seven did not respond.165 A larger follow-up clinical trial has not been performed.
Bosentan was found to effectively reduce moderate vasculopathy in the Tsk1/+ mouse94 and was effective in preventing bleomycin-induced pulmonary fibrosis.166,167 Bosentan has been used to ameliorate the microcirculation, helping to reduce Raynaud’s phenomenon. Losartan, an angiotensin II receptor inhibitor, was also effective for the treatment of Raynaud’s phenomenon, but to date the studies have not been formally published. However, losartan was found to reduce bleomycin-induced pulmonary fibrosis.168,169 Both drugs lend credence to targeting abnormalities in the vasculature as valid therapeutics for systemic sclerosis. Relaxin is a small dimeric peptide that regulates vascular tone and also has an antifibrotic mode of action. The relaxin-deficient mouse suggested that this molecule could be used to treat fibrosis and this was confirmed with bleomycin studies; however, when trialed in systemic sclerosis, there were mixed results and overall relaxin was not effective at reducing fibrosis.170
Much controversy is centered on the antibiotic minocycline as to whether it is effective in treating systemic sclerosis. One small clinical study demonstrated that four of eleven patients had complete remission of disease,171 which sparked huge interest in minocycline as a treatment for patients with systemic sclerosis. However, minocycline failed a larger open-label clinical trial.172 Like many other drugs, minocycline may be of value to a small number of patients, and that perhaps may be determined by the etiology of their disease. Because systemic sclerosis is very polymorphic in nature, its mode of onset may have many different origins.
Methotrexate failed as a drug for the treatment of adult-onset systemic sclerosis and did not significantly alter the overall skin score;173 however, it was successful in treating juvenile localized systemic sclerosis.174 Methotrexate was not shown to be effective in the bleomycin-induced mouse model of fibrosis.175 Cyclophosphamide was found to have enhanced toxicity in the lung, especially when used in conjunction with bleomycin, where mice were found to have enhanced progression of lung fibrosis,176 whereas cyclophosphamide appeared to protect against interstitial lung disease in patients with systemic sclerosis, with improvement of lung function and quality of life.177
Future directions for animal models replicating systemic sclerosis pathology
In recent years, animal models have been developed that replicate more faithfully many of the important aspects of systemic sclerosis.35,45,78,120 However, until the cause of systemic sclerosis is identified, an accurate model is not possible. Even so, once the cause has been identified, the use of animals may still preclude accurate development of the disease in an animal model, for the simple reason that animals are not humans. Recent research suggests that infection of fibroblasts by Epstein-Barr virus might drive the pathology of systemic sclerosis178 and if this proves to be true, the development of an animal model will be very difficult. The natural host for the virus is man, but under specific experimental conditions, a small number of new world monkey species can be infected. In light of this, a humanized Epstein-Barr virus mouse has already been developed, but the mouse has not been reported to display fibrosis,179 and the model may only be possible if mouse fibroblasts are specifically designed to allow infection with virus.
Due to the polymorphic and systemic nature of systemic sclerosis and the poor replicability of many of the models utilized in studies published to date, the issue arises as to whether more than one model should be used to study drug efficacy. This would be best from an idealistic point of view; however, it may not always be practical, due to the inherent expense of breeding and housing of animals for research. If more than one model is to be used, then the models should be carefully chosen. For example, in studying the efficacy of a drug on the TGF-β pathway, investigators might select animal models that target TGF-β but from different angles, such as receptor signaling using the kinase-deficient type II TGF-β receptor model and the type I TGF-β receptor transgenic model. The hope is that both models will together recapitulate and confirm present findings. However, disparate study findings can preclude a drug from entering clinical trials, and this drug may have been a successful therapeutic that is never realized, because animals are not humans.
Conclusion
Animal models are very useful in studying the initiating events, genes, and various factors required to establish fibrosis as often these events or features cannot be studied in the patient. With the exception of stiff skin syndrome, which is an inheritable variant of systemic sclerosis, all other systemic sclerosis variants are inducible, given that onset of the disease is usually later in life. Animal models, by their very nature, do not replicate all aspects of systemic sclerosis. Many of the recently developed animal models replicating various features of systemic sclerosis have provided insights into the vasculopathy, the role of proteins regulating fibrosis, and the role of the innate immune system in initiating fibrotic events. All these models have proved useful in increasing our understanding of the pathogenesis of systemic sclerosis that one day may lead to new paradigm shifts in the etiology of the disease. The animal models have led to the development of various drugs; however, many of these drugs have not translated well into therapeutics to treat systemic sclerosis, due in part to the fact that mice are not humans.
Disclosure
The author reports no conflicts of interest in this work.
References
Mayes MD, Lacey JV Jr, Beebe-Dimmer J, et al. Prevalence, incidence, survival, and disease characteristics of systemic sclerosis in a large US population. Arthritis Rheum. 2003;48:2246–2255. | |
Varga J, Abraham D. Systemic sclerosis: a prototypic multisystem fibrotic disorder. J Clin Invest. 2007;117:557–567. | |
Fanning GC, Welsh KI, Bunn C, Du Bois R, Black CM. HLA associations in three mutually exclusive autoantibody subgroups in UK systemic sclerosis patients. Br J Rheumatol. 1998;37:201–207. | |
Denton CP, Lapadula G, Mouthon L, Muller-Ladner U. Renal complications and scleroderma renal crisis. Rheumatology. 2009;48 Suppl 3: iii32–iii35. | |
Green MC, Sweet HO, Bunker LE. Tight-skin, a new mutation of the mouse causing excessive growth of connective tissue and skeleton. Am J Pathol. 1976;82:493–512. | |
Chatterjee S, Mark ME, Wooley PH, Lawrence WD, Mayes MD. Increased dermal elastic fibers in the tight skin mouse. Clin Exp Rheumatol. 2004;22:617–620. | |
Gardi C, Martorana PA, de Santi MM, van Even P, Lungarella G. A biochemical and morphological investigation of the early development of genetic emphysema in tight-skin mice. Exp Mol Pathol. 1989;50: 398–410. | |
Wong AK, Osborn TG, Miller JG, Wickline SA. Quantification of ventricular remodeling in the tight-skin mouse cardiomyopathy with acoustic microscopy. Ultrasound Med Biol. 1993;19:365–374. | |
Siracusa LD, McGrath R, Ma Q, et al. A tandem duplication within the fibrillin 1 gene is associated with the mouse tight skin mutation. Genome Res. 1996;6:300–313. | |
Saito S, Nishimura H, Brumeanu TD, et al. Characterization of mutated protein encoded by partially duplicated fibrillin-1 gene in tight skin (TSK) mice. Mol Immunol. 1999;36:169–176. | |
Murai C, Saito S, Kasturi KN, Bona CA. Spontaneous occurrence of anti-fibrillin-1 autoantibodies in tight-skin mice. Autoimmunity. 1998;28:151–155. | |
Tan FK, Arnett FC, Antohi S, et al. Autoantibodies to the extracellular matrix mircofibrillar protein, fibrillin-1, in patients with scleroderma and other connective tissue diseases. J Immunol. 1999;163:1066–1072. | |
Tan FK, Arnett FC, Reveille JD, et al. Autoantibodies to fibrillin 1 in systemic sclerosis: ethnic differences in a recognition and lack of correlation with specific clinical features and HLA alleles. Arthritis Rheum. 2000;43:2464–2471. | |
Tan FK, Strivers DN, Foster MW, et al. Association of microsatellite markers near the fibrillin 1 gene on human chromosome 15q with scleroderma in a native American population. Arthritis Rheum. 1998;41:1729–1737. | |
Tan FK, Wang N, Kuwana M, et al. Association of fibrillin 1 single-nucleotide polymorphisms haplotypes with systemic sclerosis in Chotaw and Japanese populations. Arthritis Rheum. 2001;44: 893–901. | |
Brinckmann J, Hunzelmann N, El-Hallous E, et al. Absence of autoantibodies against correctly folded recombinant fibrillin-1 protein in systemic sclerosis patients. Arthritis Res Ther. 2005;7: R1221–R1226. | |
Wipff J, Giraud M, Sibilia J, et al. Polymorphic markers of the fibrillin-1 gene and systemic sclerosis in European Caucasian patients. J Rheumatol. 2008;35:643–649. | |
Wallis DD, Tan FK, Kielty CM, Kimball MD, Arnett FC, Milewicz DM. Abnormalities in fibrillin 1-containing microfibrils in dermal fibroblast cultures from patients with systemic sclerosis (scleroderma). Arthritis Rheum. 2001;44:1855–1864. | |
Loeys BL, Gerber EE, Riegert-Johnson D, et al. Mutations in fibrillin-1 cause congenital scleroderma: stiff skin syndrome. Sci Trans Med. 2010;2(23):23ra20. | |
Gerber EE, Gallo EM, Fontana SC, et al. Integrin-modulating therapy prevents fibrosis and autoimmunity in mouse models of scleroderma. Nature. 2013;503:126–130. | |
Sassi-Gaha S, Loughlin DT, Kappler F, et al. Two dicarbonyl compounds, 3-deoxyglucosone and methylglyoxal, differentially modulate dermal fibroblasts. Matrix Biol. 2010;29:127–134. | |
Loughlin DT, Artlett CM. 3-Deoxyglucosone-collagen alters human dermal fibroblast migration and adhesion: implications for impaired wound healing in patients with diabetes. Wound Repair Regen. 2009;17: 739–749. | |
Loughlin DT, Artlett CM. Modification of collagen by 3-deoxyglucosone alters wound healing through differential regulation of p38 MAP kinase. PLoS One. 2011;6:e18676. | |
Loughlin DT, Artlett CM. Precursor of advanced glycation end products mediates ER-stress-induced caspase-3 activation of human dermal fibroblasts through NAD(P)H oxidase 4. PLoS One. 2010;5: e11093. | |
Akhmetshina A, Venalis P, Dees C, et al. Treatment with imatinib prevents fibrosis in different preclinical models of systemic sclerosis and induces regression of established fibrosis. Arthritis Rheum. 2009;60:219–224. | |
Badea I, Virtanen C, Verrall RE, Rosenberg A, Foldvari M. Effect of topical interferon-gamma gene therapy using gemini nanoparticles on pathophysiological markers of cutaneous scleroderma in Tsk/+ mice. Gene Ther. 2012;19:978–987. | |
Yoshizaki A, Yanaba K, Ogawa A, et al. The specific free radical scavenger edaravone suppresses fibrosis in the bleomycin-induced and tight skin mouse models of systemic sclerosis. Arthritis Rheum. 2011;63:3086–3097. | |
Yoshizaki A, Yanaba K, Yoshizaki A, et al. Treatment with rapamycin prevents fibrosis in tight-skin and bleomycin-induced mouse models of systemic sclerosis. Arthritis Rheum. 2010;62:2476–2487. | |
Peters J, Ball ST. Tight skin 2 (Tsk2/+). Mouse News Letter. 1986;74: 91–92. | |
Christner PJ, Siracusa LD, Hawkins DF, et al. A high resolution linkage map of the tight skin 2 (tsk2) locus: a mouse model for scleroderma (SSc) and other cutaneous fibrotic diseases. Mamm Genome. 1996;7:610–612. | |
Christner PJ, Peters J, Hawkins D, Siracusa LD, Jimenez SA. The tight skin 2 mouse. An animal model of scleroderma displaying cutaneous fibrosis and mononuclear cell infiltration. Arthritis Rheum. 1995;38:1791–1798. | |
Gentiletti J, McCloskey LJ, Artlett CM, Peters J, Jimenez SA, Christner PJ. Demonstration of autoimmunity in the tight skin-2 mouse: a model for scleroderma. J Immunol. 2005;175:2418–2426. | |
Christner PJ, Hitraya EG, Peters J, McGrath R, Jimenez SA. Transcriptional activation of the alpha1(I) procollagen gene and up-regulation of alpha1(I) and alpha1(III) procollagen messenger RNA in dermal fibroblasts from tight skin 2 mice. Arthritis Rheum. 1998;41: 2132–2142. | |
Wooley PH, Sud S, Langendorfer A, et al. T cells infiltrating the skin of Tsk2 scleroderma-like mice exhibit T cell receptor bias. Autoimmunity. 1998;27:91–98. | |
Gruschwitz MS, Moormann S, Kromer G, et al. Phenotypic analysis of skin infiltrates in comparison with peripheral blood lymphocytes, spleen cells and thymocytes in early avian scleroderma. J Autoimmun. 1991;4:577–593. | |
Nguyen VA, Sgonc R, Dietrich H, Wick G. Endothelial injury in internal organs of University of California at Davis line 200 (UCD 200) chickens, an animal model for systemic sclerosis (scleroderma). J Autoimmun. 2000;14:143–149. | |
Gershwin ME, Abplanalp H, Castles JJ, et al. Characterization of a spontaneous disease of white leghorn chickens resembling progressive systemic sclerosis (scleroderma). J Exp Med. 1981;153: 1640–1659. | |
Worda M, Sgonc R, Dietrich H, et al. In vivo analysis of the apoptosis-inducing effect of anti-endothelial cell antibodies in systemic sclerosis by the chorionallantoic membrane assay. Arthritis Rheum. 2003;48: 2605–2614. | |
Ek W, Sahlqvist AS, Crooks L, et al. Mapping QTL affecting a systemic sclerosis-like disorder in a cross between UCD-200 and red jungle fowl chickens. Dev Comp Immunol. 2012;38:352–359. | |
Asano Y, Ihn H, Yamane K, Kubo M, Tamaki K. Impaired Smad7-Smurf-mediated negative regulation of TGF-beta signaling in scleroderma fibroblasts. J Clin Invest. 2004;113:253–264. | |
Brissett M, Veraldi KL, Pilewski JM, Medsger TA, Feghali-Bostwick CA. Localized expression of tenascin in systemic sclerosis-associated pulmonary fibrosis and its regulation by insulin-like growth factor binding protein 3. Arthritis Rheum. 2012;64:272–280. | |
Radstake TR, Gorlova O, Rueda B, et al. Genome-wide association study of systemic sclerosis identifies CD247 as a new susceptibility locus. Nat Genet. 2010;42:426–429. | |
Fujimoto M, Tsutsui H, Xinshou O, et al. Inadequate induction of suppressor of cytokine signaling-1 causes systemic autoimmune diseases. Int Immunol. 2004;16:303–314. | |
Fischer FR, Santambrogio L, Luo Y, Berman MA, Hancock WW, Dorf ME. Modulation of experimental autoimmune encephalomyelitis: effect of altered peptide ligand on chemokine and chemokine receptor expression. J Neuroimmunol. 2000;110:195–208. | |
Gruschwitz MS, Shoenfeld Y, Krupp M, et al. Antinuclear antibody profile in UCD line 200 chickens: a model for progressive systemic sclerosis. Int Arch Allergy Immunol. 1993;100:307–313. | |
Raman V, Quillen K, Sloan JM. Acquired Glanzmann thrombasthenia associated with Hodgkin lymphoma: rapid reversal of functional platelet defect with ABVD (adriamycin/bleomycin/ vinblastine/dacarbazine) chemotherapy. Clin Lymphoma Myeloma Leuk. 2014;14:e51–e54. | |
Lewis TG, Nydorf ED. Intralesional bleomycin for warts: a review. J Drugs Dermatol. 2006;5:499–504. | |
Inaoki M, Kawabata C, Nishijima C, Yoshio N, Kita T. Case of bleomycin-induced scleroderma. J Dermatol. 2012;39:482–484. | |
Mountz JD, Downs Minor MB, Turner R, Thomas TB, Richards F, Pisko E. Bleomycin-induced cutaneous toxicity in the rat: analysis of histopathology and ultrastructure compared with progressive systemic sclerosis (scleroderma). Br J Dermatol. 1983;108:679–686. | |
Finch WR, Rodnan GP, Buckingham RB, Prince RK, Winkelstein A. Bleomycin-induced scleroderma. J Rheumatol. 1980;7:651–659. | |
Yamamoto T, Takagawa S, Katayama I, et al. Animal model of sclerotic skin. Local injections of bleomycin induces sclerosis skin mimicking scleroderma. J Invest Dermatol. 1999;112:456–462. | |
Yamamoto T, Kuroda M, Nishioka K. Animal model of sclerotic skin. III: histopathological comparison of bleomycin-induced scleroderma in various mice strains. Arch Dermatol Res. 2000;292:535–541. | |
Yamamoto T, Katayama I. Vascular changes in bleomycin-induced scleroderma. Int J Rheumatol. 2011;2011:270938. | |
Clements PJ, Hurwitz EL, Wong WK, et al. Skin thickness score as a predictor and correlate of outcome in systemic sclerosis: high-dose versus low-dose penicillamine trial. Arthritis Rheum. 2000;43: 2445–2454. | |
Artlett CM, Sassi-Gaha S, Rieger JL, Boesteanu AC, Feghali-Bostwick CA, Katsikis PD. The inflammasome activating caspase-1 mediates fibrosis and myofibroblast differentiation in systemic sclerosis. Arthritis Rheum. 2011;63:3563–3574. | |
Mouratis MA, Aidinis V. Modeling pulmonary fibrosis with bleomycin. Curr Opin Pulm Med. 2011;17:355–361. | |
Santos-Silva MA, Pires KM, Trajano ET, et al. Redox imbalance and pulmonary function in bleomycin-induced fibrosis in C57BL/6, DBA/2, and BALB/c mice. Toxicol Pathol. 2012;40:731–741. | |
Lee R, Reese C, Bonner M, et al. Bleomycin delivery by osmotic mini-pump: similarity to human scleroderma interstitial lung disease. Am J Physiol Lung Cell Mol Physiol. 2014;306:L736–L748. | |
Heid ME, Keyel PA, Kamga C, Shiva S, Watkins SC, Salter RD. Mitochondrial reactive oxygen species induces NLRP3-dependent lysosomal damage and inflammasome activation. J Immunol. 2013;191: 5230–5238. | |
Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature. 2011;469:221–225. | |
Liu R, Chen H, Bai H, et al. Suppression of nuclear factor erythroid 2-related factor 2 via extracellular signal-regulated kinase contributes to bleomycin-induced oxidative stress and fibrogenesis. Toxicol Lett. 2013;220:15–25. | |
Cort A, Ozdemir E, Timur M, Ozben T. Effects of N-acetyl-L-cysteine on bleomycin induced oxidative stress in malignant testicular germ cell tumors. Biochimie. 2012;94:2734–2739. | |
Brar SS, Meyer JN, Bortner CD, Van Houten B, Martin WJ 2nd. Mitochondrial DNA-depleted A549 cells are resistant to bleomycin. Am J Physiol Lung Cell Mol Physiol. 2012;303:L413–L424. | |
Zhou CF, Yu JF, Zhang JX, et al. N-acetylcysteine attenuates subcutaneous administration of bleomycin-induced skin fibrosis and oxidative stress in a mouse model of scleroderma. Clin Exp Dermatol. 2013;38:403–409. | |
Feldmeyer L, Werner S, French LE, Beer HD. Interleukin-1, inflammasomes and the skin. Eur J Cell Biol. 2010;89:638–644. | |
Mortaz E, Henricks PA, Kraneveld AD, Givi ME, Garssen J, Folkerts G. Cigarette smoke induces the release of CXCL-8 from human bronchial epithelial cells via TLRs and induction of the inflammasome. Biochim Biophys Acta. 2011;1812:1104–1110. | |
Ishikawa H, Takeda K, Okamoto A, Matsuo S, Isobe K. Induction of autoimmunity in a bleomycin-induced murine model of experimental systemic sclerosis: an important role for CD4+ T cells. J Invest Dermatol. 2009;129:1688–1695. | |
Sambo P, Jannino L, Candela M, et al. Monocytes of patients with systemic sclerosis (scleroderma) spontaneously release in vitro increased amounts of superoxide anion. J Invest Dermatol. 1999;112:78–84. | |
Sambo P, Baroni SS, Luchetti M, et al. Oxidative stress in scleroderma: maintenance of scleroderma fibroblast phenotype by the constitutive up-regulation of reactive oxygen species generation through the NADPH oxidase complex pathway. Arthritis Rheum. 2001;44:2653–2664. | |
Bhandary B, Marahatta A, Kim HR, Chae HJ. An involvement of oxidative stress in endoplasmic reticulum stress and its associated diseases. Int J Mol Sci. 2012;14:434–456. | |
Servettaz A, Goulvestre C, Kavian N, et al. Selective oxidation of DNA topoisomerase 1 induces systemic sclerosis in the mouse. J Immunol. 2009;182:5855–5864. | |
Servettaz A, Kavian N, Nicco C, et al. Targeting the cannabinoid pathway limits the development of fibrosis and autoimmunity in a mouse model of systemic sclerosis. Am J Pathol. 2010;177:187–196. | |
Batteux F, Kavian N, Servettaz A. New insights on chemically induced animal models of systemic sclerosis. Curr Opin Rheumatol. 2011;23:511–518. | |
Bagnato G, Bitto A, Irrera N, et al. Propylthiouracil prevents cutaneous and pulmonary fibrosis in the reactive oxygen species murine model of systemic sclerosis. Arthritis Res Ther. 2013;15:R120. | |
McCormick LL, Zhang Y, Tootell E, Gilliam AC. Anti-TGF-á treatment prevents skin and lung fibrosis in murine sclerodermatous graft-versus-host disease: a model for human scleroderma. J Immunol. 1999;163:5693–5699. | |
Zhang Y, McCormick LL, Desai SR, Wu C, Gilliam AC. Murine sclerodermatous graft-versus-host disease, a model for human scleroderma: cutaneous cytokines, chemokines, and immune cell activation. J Immunol. 2002;168:3088–3098. | |
Mavalia C, Scaletti C, Romagnani P, et al. Type 2 helper T-cell predominance and high CD30 expression in systemic sclerosis. Am J Pathol. 1997;151:1751–1758. | |
Ruzek MC, Jha S, Ledbetter S, Richards SM, Garman RD. A modified model of graft-versus-host-induced systemic sclerosis (scleroderma) exhibits all major aspects of the human disease. Arthritis Rheum. 2004;50:1319–1331. | |
Stawski L, Han R, Bujor AM, Trojanowska M. Angiotensin II induces skin fibrosis: a novel mouse model of dermal fibrosis. Arthritis Res Ther. 2012;14:R194. | |
Zhao Q, Ishibashi M, Hiasa K, Tan C, Takeshita A, Egashira K. Essential role of vascular endothelial growth factor in angiotensin II-induced vascular inflammation and remodeling. Hypertension. 2004;44:264–270. | |
Marut W, Kavian N, Servettaz A, et al. Amelioration of systemic fibrosis in mice by angiotensin II receptor blockade. Arthritis Rheum. 2013;65:1367–1377. | |
Kawaguchi Y, Takagi K, Hara M, et al. Angiotensin II in the lesional skin of systemic sclerosis patients contributes to tissue fibrosis via angiotensin II type 1 receptors. Arthritis Rheum. 2004;50:216–226. | |
Yoshizaki A, Yanaba K, Ogawa A, Asano Y, Kadono T, Sato S. Immunization with DNA topoisomerase I and Freund’s complete adjuvant induces skin and lung fibrosis and autoimmunity via interleukin-6 signaling. Arthritis Rheum. 2011;63:3575–3585. | |
Abraham D, Dashwood M. Endothelin – role in vascular disease. Rheumatology. 2008;47 Suppl 5:v23–v24. | |
Bouallegue A, Daou GB, Srivastava AK. Endothelin-1-induced signaling pathways in vascular smooth muscle cells. Curr Vasc Pharmacol. 2007;5:45–52. | |
Marasciulo FL, Montagnani M, Potenza MA. Endothelin-1: the yin and yang on vascular function. Curr Med Chem. 2006;13:1655–1665. | |
Hocher B, Thone-Reineke C, Rohmeiss P, et al. Endothelin-1 transgenic mice develop glomerulosclerosis, interstitial fibrosis, and renal cysts but not hypertension. J Clin Invest. 1997;99:1380–1389. | |
Hocher B, Schwarz A, Fagan KA, et al. Pulmonary fibrosis and chronic lung inflammation in ET-1 transgenic mice. Am J Respir Cell Mol Biol. 2000;23:19–26. | |
Vancheeswaran R, Magoulas T, Efrat G, et al. Circulating endothelin-1 levels in systemic sclerosis subsets – a marker of fibrosis or vascular dysfunction? J Rheumatol. 1994;21:1838–1844. | |
Kawaguchi Y, Suzuki K, Hara M, et al. Increased endothelin-1 production in fibroblasts derived from patients with systemic sclerosis. Ann Rheum Dis. 1994;53:506–510. | |
Rubin LJ, Badesch DB, Barst RJ, et al. Bosentan therapy for pulmonary arterial hypertension. N Engl J Med. 2002;346:896–903. | |
Joglekar A, Tsai FS, McCloskey DA, Wilson JE, Seibold JR, Riley DJ. Bosentan in pulmonary arterial hypertension secondary to scleroderma. J Rheumatol. 2006;33:61–68. | |
Marie I, Beny JL. Endothelial dysfunction in murine model of systemic sclerosis: tight-skin mice 1. J Invest Dermatol. 2002;119: 1379–1387. | |
Richard V, Solans V, Favre J, et al. Role of endogenous endothelin in endothelial dysfunction in murine model of systemic sclerosis: tight skin mice 1. Fundam Clin Pharmacol. 2008;22:649–655. | |
Maurer B, Busch N, Jungel A, et al. Transcription factor fos-related antigen-2 induces progressive peripheral vasculopathy in mice closely resembling human systemic sclerosis. Circulation. 2009;120: 2367–2376. | |
Maurer B, Distler JH, Distler O. The Fra-2 transgenic mouse model of systemic sclerosis. Vasc Pharmacol. 2013;58:194–201. | |
Sonnylal S, Denton CP, Zheng B, et al. Postnatal induction of transforming growth factor beta signaling in fibroblasts of mice recapitulates clinical, histologic, and biochemical features of scleroderma. Arthritis Rheum. 2007;56:334–344. | |
Rabquer BJ, Koch AE. Angiogenesis and vasculopathy in systemic sclerosis: evolving concepts. Curr Rheumatol Rep. 2012;14:56–63. | |
Fleming JN, Nash RA, Mahoney WM Jr, Schwartz SM. Is scleroderma a vasculopathy? Curr Rheumatol Rep. 2009;11:103–110. | |
Koenig M, Joyal F, Fritzler MJ, et al. Autoantibodies and microvascular damage are independent predictive factors for the progression of Raynaud’s phenomenon to systemic sclerosis: a twenty-year prospective study of 586 patients, with validation of proposed criteria for early systemic sclerosis. Arthritis Rheum. 2008; 58:3902–3912. | |
Bou-Gharios G, Garrett LA, Rossert J, et al. A potent far-upstream enhancer in the mouse pro alpha 2(I) collagen gene regulates expression of reporter genes in transgenic mice. J Cell Biol. 1996;134: 1333–1344. | |
Denton CP, Zheng B, Evans LA, et al. Fibroblast-specific expression of a kinase-deficient type II transforming growth factor beta (TGFbeta) receptor leads to paradoxical activation of TGFbeta signaling pathways with fibrosis in transgenic mice. J Biol Chem. 2003;278: 25109–25119. | |
Denton CP, Lindahl GE, Khan K, et al. Activation of key profibrotic mechanisms in transgenic fibroblasts expressing kinase-deficient type II transforming growth factor-{beta} receptor (T{beta}RII{delta}k). J Biol Chem. 2005;280:16053–16065. | |
Derrett-Smith EC, Dooley A, Gilbane AJ, et al. Endothelial injury in a transforming growth factor beta-dependent mouse model of scleroderma induces pulmonary arterial hypertension. Arthritis Rheum. 2013;65:2928–2939. | |
Overbeek MJ, Boonstra A, Voskuyl AE, et al. Platelet-derived growth factor receptor-beta and epidermal growth factor receptor in pulmonary vasculature of systemic sclerosis-associated pulmonary arterial hypertension versus idiopathic pulmonary arterial hypertension and pulmonary veno-occlusive disease: a case-control study. Arthritis Res Ther. 2011;13:R61. | |
Trojanowska M. Role of PDGF in fibrotic diseases and systemic sclerosis. Rheumatology. 2008;47 Suppl 5:v2–v4. | |
Olson LE, Soriano P. Increased PDGFRalpha activation disrupts connective tissue development and drives systemic fibrosis. Dev Cell. 2009;16:303–313. | |
Kavian N, Servettaz A, Marut W, et al. Sunitinib inhibits the phosphorylation of platelet-derived growth factor receptor beta in the skin of mice with scleroderma-like features and prevents the development of the disease. Arthritis Rheum. 2012;64:1990–2000. | |
Razani B, Woodman SE, Lisanti MP. Caveolae: from cell biology to animal physiology. Pharmacol Rev. 2002;54:431–467. | |
Drab M, Verkade P, Elger M, et al. Loss of caveolae, vascular dysfunction, and pulmonary defects in caveolin-1 gene-disrupted mice. Science. 2001;293:2449–2452. | |
Castello-Cros R, Whitaker-Menezes D, Molchansky A, et al. Scleroderma-like properties of skin from caveolin-1-deficient mice: implications for new treatment strategies in patients with fibrosis and systemic sclerosis. Cell Cycle. 2011;10:2140–2150. | |
Razani B, Zhang XL, Bitzer M, von Gersdorff G, Bottinger EP, Lisanti MP. Caveolin-1 regulates transforming growth factor (TGF)-beta/SMAD signaling through an interaction with the TGF-beta type I receptor. J Biol Chem. 2001;276:6727–6738. | |
Tourkina E, Gooz P, Pannu J, et al. Opposing effects of protein kinase Cà and protein kinase Cî on collagen expression by human lung fibroblasts are mediated via MEK/ERK and caveolin-1 signaling. J Biol Chem. 2005;280:13879–13887. | |
Wang XM, Zhang Y, Kim HP, et al. Caveolin-1: a critical regulator of lung fibrosis in idiopathic pulmonary fibrosis. J Exp Med. 2006;203: 2895–2906. | |
Pavlides S, Tsirigos A, Vera I, et al. Loss of stromal caveolin-1 leads to oxidative stress, mimics hypoxia and drives inflammation in the tumor microenvironment, conferring the “reverse Warburg effect”: a transcriptional informatics analysis with validation. Cell Cycle. 2010;9:2201–2219. | |
Avouac J, Borderie D, Ekindjian OG, Kahan A, Allanore Y. High DNA oxidative damage in systemic sclerosis. J Rheumatol. 2010;37(12):2540–2547. | |
Erre GL, De Muro P, Dellaca P, et al. Iloprost therapy acutely decreases oxidative stress in patients affected by systemic sclerosis. Clin Exp Rheumatol. 2008;26:1095–1098. | |
Liang SH, Zhang W, McGrath BC, Zhang P, Cavener DR. PERK (eIF2alpha kinase) is required to activate the stress-activated MAPKs and induce the expression of immediate-early genes upon disruption of ER calcium homoeostasis. Biochem J. 2006;393:201–209. | |
Wu M, Melichian DS, de la Garza M, et al. Essential roles for early growth response transcription factor Egr-1 in tissue fibrosis and wound healing. Am J Pathol. 2009;175:1041–1055. | |
Asano Y, Stawski L, Hant F, et al. Endothelial Fli1 deficiency impairs vascular homeostasis: a role in scleroderma vasculopathy. Am J Pathol. 2010;176:1983–1998. | |
Nakerakanti SS, Kapanadze B, Yamasaki M, Markiewicz M, Trojanowska M. Fli1 and Ets1 have distinct roles in connective tissue growth factor/CCN2 gene regulation and induction of the profibrotic gene program. J Biol Chem. 2006;281:25259–25269. | |
Fleming JN, Nash RA, Mahoney WM Jr, Schwartz SM. Is scleroderma a vasculopathy? Curr Rheumatol Rep. 2009;11:103–110. | |
Asano Y, Markiewicz M, Kubo M, Szalai G, Watson DK, Trojanowska M. Transcription factor Fli1 regulates collagen fibrillogenesis in mouse skin. Mol Cell Biol. 2009;29:425–434. | |
Chujo S, Shirasaki F, Kondo-Miyazaki M, Ikawa Y, Takehara K. Role of connective tissue growth factor and its interaction with basic fibroblast growth factor and macrophage chemoattractant protein-1 in skin fibrosis. J Cell Physiol. 2009;220:189–195. | |
Wang X, Li X, Ye L, Chen W, Yu X. Smad7 inhibits TGF-beta1-induced MCP-1 upregulation through a MAPK/p38 pathway in rat peritoneal mesothelial cells. Int Urol Nephrol. 2013;45:899–907. | |
Jones SG, Morrisey K, Williams JD, Phillips AO. TGF-beta1 stimulates the release of pre-formed bFGF from renal proximal tubular cells. Kidney Int. 1999;56:83–91. | |
Tall EG, Bernstein AM, Oliver N, Gray JL, Masur SK. TGF-beta-stimulated CTGF production enhanced by collagen and associated with biogenesis of a novel 31-kDa CTGF form in human corneal fibroblasts. Invest Ophthalmol Vis Sci. 2010;51:5002–5011. | |
McCann MR, Monemdjou R, Ghassemi-Kakroodi P, et al. mPGES-1 null mice are resistant to bleomycin-induced skin fibrosis. Arthritis Res Ther. 2011;13:R6. | |
Wu M, Melichian DS, Chang E, Warner-Blankenship M, Ghosh AK, Varga J. Rosiglitazone abrogates bleomycin-induced scleroderma and blocks profibrotic responses through peroxisome proliferator-activated receptor-gamma. Am J Pathol. 2009;174:519–533. | |
Ghosh AK, Bhattacharyya S, Lakos G, Chen SJ, Mori Y, Varga J. Disruption of transforming growth factor beta signaling and profibrotic responses in normal skin fibroblasts by peroxisome proliferator-activated receptor gamma. Arthritis Rheum. 2004;50:1305–1318. | |
Wei J, Ghosh AK, Sargent JL, et al. PPARgamma downregulation by TGFβ in fibroblast and impaired expression and function in systemic sclerosis: a novel mechanism for progressive fibrogenesis. PLoS One. 2010;5:e13778. | |
Kapoor M, McCann M, Liu S, et al. Loss of peroxisome proliferator-activated receptor gamma in mouse fibroblasts results in increased susceptibility to bleomycin-induced skin fibrosis. Arthritis Rheum. 2009;60:2822–2829. | |
Cheang WS, Fang X, Tian XY. Pleiotropic effects of peroxisome proliferator-activated receptor gamma and delta in vascular diseases. Circulation J. 2013;77:2664–2671. | |
Lu Y, Azad N, Wang L, et al. Phosphatidylinositol-3-kinase/akt regulates bleomycin-induced fibroblast proliferation and collagen production. Am J Resp Cell Mol Biol. 2010;42:432–441. | |
Shi-Wen X, Chen Y, Denton CP, et al. Endothelin-1 promotes myofibroblast induction through the ETA receptor via a rac/phosphoinositide 3-kinase/Akt-dependent pathway and is essential for the enhanced contractile phenotype of fibrotic fibroblasts. Mol Biol Cell. 2004;15: 2707–2719. | |
Parapuram SK, Shi-wen X, Elliott C, et al. Loss of PTEN expression by dermal fibroblasts causes skin fibrosis. J Invest Dermatol. 2011;131: 1996–2003. | |
Tang ML, Samuel CS, Royce SG. Role of relaxin in regulation of fibrosis in the lung. Ann N Y Acad Sci. 2009;1160:342–347. | |
Huang X, Gai Y, Yang N, et al. Relaxin regulates myofibroblast contractility and protects against lung fibrosis. Am J Pathol. 2011;179: 2751–2765. | |
Samuel CS, Zhao C, Yang Q, et al. The relaxin gene knockout mouse: a model of progressive scleroderma. J Invest Dermatol. 2005;125: 692–699. | |
Bennett RG, Heimann DG, Singh S, Simpson RL, Tuma DJ. Relaxin decreases the severity of established hepatic fibrosis in mice. Liver Int. 2013;34:416–426. | |
Sassoli C, Chellini F, Pini A, et al. Relaxin prevents cardiac fibroblast-myofibroblast transition via notch-1-mediated inhibition of TGF-beta/Smad3 signaling. PLoS One. 2013;8:e63896. | |
Li XF, Liao J, Xin ZQ, Lu WQ, Liu AL. Relaxin attenuates silica-induced pulmonary fibrosis by regulating collagen type I and MMP-2. Int Immunopharmacol. 2013;17:537–542. | |
Gasse P, Mary C, Guenon I, et al. IL-1R1/MyD88 signaling and the inflammasome are essential in pulmonary inflammation and fibrosis in mice. J Clin Invest. 2007;117:3786–3799. | |
Sendelbach LE, Lindenschmidt RC, Witschi HP. The effect of cyclosporin A on pulmonary fibrosis induced by butylated hydroxytoluene, bleomycin and beryllium sulfate. Toxicol Lett. 1985;6: 169–173. | |
Lossos IS, Or R, Goldstein RH, Conner MW, Breuer R. Amelioration of bleomycin-induced pulmonary injury by cyclosporin A. Exp Lung Res. 1996;22:237–249. | |
Basso M, Filaci G, Cutolo M, et al. Long-term treatment of patients affected by systemic sclerosis with cyclosporin A. Ann Ital Med Int. 2001;16:233–239. | |
Manno R, Boin F. Immunotherapy of systemic sclerosis. Immunotherapy. 2010;2:863–878. | |
Guillevin L, Berezne A, Seror R, et al. Scleroderma renal crisis: a retrospective multicentre study on 91 patients and 427 controls. Rheumatology. 2012;51:460–467. | |
Ozgen M, Koca SS, Dagli AF, Gundogdu B, Ustundag B, Isik A. Mycophenolate mofetil and daclizumab targeting T lymphocytes in bleomycin-induced experimental scleroderma. Clin Exp Dermatol. 2012;37:48–54. | |
Mendoza FA, Nagle SJ, Lee JB, Jimenez SA. A prospective observational study of mycophenolate mofetil treatment in progressive diffuse cutaneous systemic sclerosis of recent onset. J Rheumatol. 2012;39:1241–1247. | |
Singhal S, Mehta J. Thalidomide in cancer. Biomed Pharmacother. 2002;56:4–12. | |
Oliver SJ, Moreira A, Kaplan G. Immune stimulation in scleroderma patients treated with thalidomide. Clin Immunol. 2000;97: 109–120. | |
Zhang L, Zhu Y, Luo W, Xi P, Yan Y. The protective effect of colchicine on bleomycin-induced pulmonary fibrosis in rats. Chin Med Sci J. 1992;7:58–60. | |
Guttadauria M, Diamond H, Kaplan D. Colchicine in the treatment of scleroderma. J Rheumatol. 1977;4:272–276. | |
Frayha RA. Colchicine therapy in scleroderma. Preliminary results. Dermatologica. 1979;159:78–81. | |
Koca SS, Isik A, Ozercan IH, Ustundag B, Evren B, Metin K. Effectiveness of etanercept in bleomycin-induced experimental scleroderma. Rheumatology. 2008;47:172–175. | |
Lam GK, Hummers LK, Woods A, Wigley FM. Efficacy and safety of etanercept in the treatment of scleroderma-associated joint disease. J Rheumatol. 2007;34:1636–1637. | |
Jackson RM, Fell CD. Etanercept for idiopathic pulmonary fibrosis: lessons on clinical trial design. Am J Resp Crit Care Med. 2008;178: 889–891. | |
Allanore Y, Devos-Francois G, Caramella C, Boumier P, Jounieaux V, Kahan A. Fatal exacerbation of fibrosing alveolitis associated with systemic sclerosis in a patient treated with adalimumab. Ann Rheum Dis. 2006;65:834–835. | |
Prey S, Ezzedine K, Doussau A, et al. Imatinib mesylate in scleroderma-associated diffuse skin fibrosis: a phase II multicentre randomized double-blinded controlled trial. Br J Dermatol. 2012;167: 1138–1144. | |
Aytemur ZA, Hacievliyagil SS, Iraz M, et al. Effects of iloprost on bleomycin-induced pulmonary fibrosis in rats compared with methyl-prednisolone. Rev Port Pneumol. 2012;18:272–277. | |
Bali G, Schwantzer G, Aberer F, Kraenke B, Aberer E. Discontinuing long-term Iloprost treatment for Raynaud’s phenomenon and systemic sclerosis: a single-center, randomized, placebo-controlled, double-blind study. Acta Dermatovenerol Alp Panonica Adriat. 2011;20: 13–21. | |
Tingey T, Shu J, Smuczek J, Pope J. Meta-analysis of healing and prevention of digital ulcers in systemic sclerosis. Arthritis Care Res. 2013;65:1460–1471. | |
Nelson EF, Huang CW, Ewel JM, Chang AA, Yuan C. Halofuginone down-regulates Smad3 expression and inhibits the TGFbeta-induced expression of fibrotic markers in human corneal fibroblasts. Mol Vis. 2012;18:479–487. | |
Pines M, Snyder D, Yarkoni S, Nagler A. Halofuginone to treat fibrosis in chronic graft-versus-host disease and scleroderma. Biol Blood Marrow Transplant. 2003;9:417–425. | |
Penn H, Quillinan N, Khan K, et al. Targeting the endothelin axis in scleroderma renal crisis: rationale and feasibility. QJM. 2013;106: 839–848. | |
Guiducci S, Bellando Randone S, Bruni C, et al. Bosentan fosters microvascular de-remodelling in systemic sclerosis. Clin Rheumatol. 2012;31:1723–1735. | |
Molina-Molina M, Serrano-Mollar A, Bulbena O, et al. Losartan attenuates bleomycin induced lung fibrosis by increasing prostaglandin E2 synthesis. Thorax. 2006;61:604–610. | |
Yao HW, Zhu JP, Zhao MH, Lu Y. Losartan attenuates bleomycin-induced pulmonary fibrosis in rats. Respiration. 2006;73:236–242. | |
Khanna D, Clements PJ, Furst DE, et al. Recombinant human relaxin in the treatment of systemic sclerosis with diffuse cutaneous involvement: a randomized, double-blind, placebo-controlled trial. Arthritis Rheum. 2009;60:1102–1111. | |
Le CH, Morales A, Trentham DE. Minocycline in early diffuse scleroderma. Lancet. 1998;352:1755–1756. | |
Mayes MD, O’Donnell D, Rothfield NF, Csuka ME. Minocycline is not effective in systemic sclerosis: results of an open-label multicenter trial. Arthritis Rheum. 2004;50:553–557. | |
Krishna Sumanth M, Sharma VK, Khaitan BK, Kapoor A, Tejasvi T. Evaluation of oral methotrexate in the treatment of systemic sclerosis. Int J Dermatol. 2007;46:218–223. | |
Zulian F, Vallongo C, Patrizi A, et al. A long-term follow-up study of methotrexate in juvenile localized scleroderma (morphea). J Am Acad Dermatol. 2012;67:1151–1156. | |
Ozgen M, Koca SS, Isik A, Ozercan IH. Methotrexate has no antifibrotic effect in bleomycin-induced experimental scleroderma. J Rheumatol. 2010;37:678–679. | |
Saba, Khan S, Parvez S, et al. Ellagic acid attenuates bleomycin and cyclophosphamide-induced pulmonary toxicity in Wistar rats. Food Chem Toxicol. 2013;58:210–219. | |
Ando K, Motojima S, Doi T, et al. Effect of glucocorticoid monotherapy on pulmonary function and survival in Japanese patients with scleroderma-related interstitial lung disease. Respir Investig. 2013;51: 69–75. | |
Farina A, Cirone M, York M, et al. Epstein-Barr virus infection induces aberrant TLR activation pathway and fibroblast-myofibroblast conversion in scleroderma. J Invest Dermatol. 2014;134:954–964. | |
Fujiwara S. Reproduction of Epstein-Barr virus infection and pathogenesis in humanized mice. Immune Netw. 2014;14:1–6. | |
Medina FA, de Almeida CJ, Dew E, et al. Caveolin-1-deficient mice show defects in innate immunity and inflammatory immune response during Salmonella enterica serovar Typhimurium infection. Infect Immun. 2006;74:6665–6674. | |
Park SH, Saleh D, Giaid A, Michel RP. Increased endothelin-1 in bleomycin-induced pulmonary fibrosis and the effect of an endothelin receptor antagonist. Am J Resp Crit Care Med. 1997;156:600–608. | |
Alarcon-Segovia D, Ramos-Niembro F, Ibanez de Kasep G, Alcocer J, Tamayo RP. Long-term evaluation of colchicine in the treatment of scleroderma. J Rheumatol. 1979;6:705–712. | |
Trivedi R, Redente EF, Thakur A, Riches DW, Kompella UB. Local delivery of biodegradable pirfenidone nanoparticles ameliorates bleomycin-induced pulmonary fibrosis in mice. Nanotechnology. 2012;23:505101. | |
Kehrer JP, Margolin SB. Pirfenidone diminishes cyclophosphamide-induced lung fibrosis in mice. Toxicol Lett. 1997;90:125–132. | |
Antoniu SA. Pirfenidone for the treatment of idiopathic pulmonary fibrosis. Expert Opin Investig Drugs. 2006;15:823–828. | |
Seibold JR, Korn JH, Simms R, et al. Recombinant human relaxin in the treatment of scleroderma. A randomized, double-blind, placebo-controlled trial. Ann Intern Med. 2000;132:871–879. | |
Jordan S, Distler JH, Maurer B, et al; on behalf of the EUSTAR Rituximab Study Group. Effects and safety of rituximab in systemic sclerosis: an analysis from the European Scleroderma Trial and Research (EUSTAR) group. Ann Rheum Dis. Epub 2014 January 17. | |
Dees C, Akhmetshina A, Zerr P, et al. Platelet-derived serotonin links vascular disease and tissue fibrosis. J Exp Med. 2011;208:961–972. | |
Tabata C, Tabata R, Kadokawa Y, et al. Thalidomide prevents bleomycin-induced pulmonary fibrosis in mice. J Immunol. 2007;179: 708–714. | |
Weingartner S, Zerr P, Tomcik M, et al. Pomalidomide is effective for prevention and treatment of experimental skin fibrosis. Ann Rheum Dis. 2012;71:1895–1899. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.