")
Back to Journals » Drug Design, Development and Therapy » Volume 9
Anesthetic drug midazolam inhibits cardiac human ether-à-go-go-related gene channels: mode of action
Authors Vonderlin N, Fischer F, Zitron E, Seyler C, Scherer D, Thomas D , Katus HA, Scholz E
Received 15 August 2014
Accepted for publication 3 December 2014
Published 16 February 2015 Volume 2015:9 Pages 867—877
DOI https://doi.org/10.2147/DDDT.S72765
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Shu-Feng Zhou
Nadine Vonderlin,1 Fathima Fischer,1 Edgar Zitron,1,2 Claudia Seyler,1 Daniel Scherer,1 Dierk Thomas,1,2 Hugo A Katus,1,2 Eberhard P Scholz1
1Department of Internal Medicine III, University Hospital Heidelberg, 2German Centre for Cardiovascular Research, Partner Site Heidelberg/Mannheim, Heidelberg, Germany
Abstract: Midazolam is a short-acting benzodiazepine that is in wide clinical use as an anxiolytic, sedative, hypnotic, and anticonvulsant. Midazolam has been shown to inhibit ion channels, including calcium and potassium channels. So far, the effects of midazolam on cardiac human ether-à-go-go-related gene (hERG) channels have not been analyzed. The inhibitory effects of midazolam on heterologously expressed hERG channels were analyzed in Xenopus oocytes using the double-electrode voltage clamp technique. We found that midazolam inhibits hERG channels in a concentration-dependent manner, yielding an IC50 of 170 µM in Xenopus oocytes. When analyzed in a HEK 293 cell line using the patch-clamp technique, the IC50 was 13.6 µM. Midazolam resulted in a small negative shift of the activation curve of hERG channels. However, steady-state inactivation was not significantly affected. We further show that inhibition is state-dependent, occurring within the open and inactivated but not in the closed state. There was no frequency dependence of block. Using the hERG pore mutants F656A and Y652A we provide evidence that midazolam uses a classical binding site within the channel pore. Analyzing the subacute effects of midazolam on hERG channel trafficking, we further found that midazolam does not affect channel surface expression. Taken together, we show that the anesthetic midazolam is a low-affinity inhibitor of cardiac hERG channels without additional effects on channel surface expression. These data add to the current understanding of the pharmacological profile of the anesthetic midazolam.
Keywords: midazolam, anesthetics, human ether-à-go-go-related gene, potassium channels
Introduction
Midazolam is a short-acting benzodiazepine that is in wide clinical use for premedication as well as during the perioperative period.1 Due to its anticonvulsant activity, midazolam is further effective in the treatment of generalized seizures and status epilepticus.2 Even in psychiatric and behavioral emergencies, midazolam has proven to be a useful treatment option that can be applied intramuscularly when intravenous access is not readily available. Midazolam belongs to the group of imidazobenzodiazepines, a newer class of benzodiazepines.3 Similar to classic benzodiazepines, it enhances the inhibitory activity of the amino acid neurotransmitter gamma-aminobutyric acid by increasing the flow of chloride ions, which, in turn, potentiates the inhibitory effect of gamma-aminobutyric acid.2 When applied intravenously, midazolam is rapidly distributed throughout the body, with a distribution half-time of 6–15 minutes. Drowsiness starts approximately 15 minutes after oral administration and 3 minutes after intravenous application.4 When compared with other benzodiazepines, midazolam exhibits the shortest recovery time and a high metabolic clearance.5 Midazolam is known to be relatively safe in clinical practice, and the effects on cardiac repolarization have been analyzed in several studies.6–9 Interestingly, one of the studies revealed a significant increase of the QTc interval, while another study observed a tendency towards a QT interval prolongation.6,9
The human ether-à-go-go-related gene (hERG) encodes the α-subunit of IKr, which represents one of the most important repolarizing ion currents in the human ventricle.10 IKr is thought to be responsible for the termination of the plateau phase in humans.11 A decrease of IKr has been connected with an inherited and an acquired form of the long QT syndrome.12–14 It is well recognized that hERG channel inhibition may critically prolong ventricular repolarization, thereby favoring the induction of torsade de pointes tachycardia, especially in patients with a reduced repolarization reserve.
So far, midazolam has been shown to exert inhibitory effects on calcium as well as potassium channels.15–17 However, the inhibitory effects of midazolam on cardiac hERG channels have not been studied as yet. Here we show that midazolam is a low-affinity inhibitor of cardiac hERG channels without additional effects on channel trafficking.
Materials and methods
Solutions and drug administration
Double-electrode voltage clamp experiments were performed in Xenopus oocytes using a standard external solution containing (in mM) 5 KCl, 100 NaCl, 1.5 CaCl2, 2 MgCl2, and 10 HEPES (pH adjusted to 7.4 with NaOH). Electrodes were back-filled with 3 M KCl solution. Midazolam was purchased from Roche (Grenzach, Germany), and stored and handled according to the manufacturer’s specifications. On the day of the experiment, stock solution was further diluted to the desired concentration with external solution. Having obtained a control measurement, electrodes were carefully removed from the oocyte and cells were placed in 10 mL plastic dishes for drug incubation. After a period of 30 minutes, cells were again placed into the recording chamber and the experiment was repeated. Only for the analysis of the time course of block development, midazolam was added to the bath solution while the cells were continuously attached to the micropipettes. Whole cell patch-clamp experiments were performed in a HEK 293 cell line stably expressing hERG channels using an a standard external solution containing (in mM) 140 NaCl, 5 KCl, 1 MgCl2, 10 HEPES, 1.8 CaCl2, 10 glucose (pH adjusted to 7.4 with NaOH) and a pipette solution containing (in mM) 100 K-aspartate, 10 KCl, 2 MgCl2, 1 CaCl2, 10 EGTA, 10 HEPES, and 40 glucose (pH adjusted to 7.2 with KOH). All electrophysiological experiments were performed at room temperature (22°C).
Heterologous expression of hERG channel subunits in Xenopus oocytes
The hERG clone (GenBank accession number u04270) was a generous gift from MT Keating (Boston, MA, USA). Complementary hERG RNA was prepared from hERG complementary DNA with the mMESSAGE mMACHINE in vitro transcription kit (Ambion, Spitfire Close, Huntingdon, UK) by use of SP6 polymerase after linearization with EcoRI (Roche, Mannheim, Germany). Generation of the hERG pore mutations Y652A and F656A has been described elsewhere.18 Only stage V and VI defolliculated oocytes were used for RNA injection. The volume of injected complementary RNA was 50 nL per oocyte. Measurements were carried out 1–3 days after complementary RNA injection. The investigation was approved by the regional administrative council and conforms to the Guide for the Care and Use of Laboratory animals published by the US National Institutes of Health (publication No 85-23, revised 1996).
Heterologous expression of hERG channel subunits in a HEK cell line
The HEK cell line stably expressing hERG channels was generously provided by Barbara A Wible (Cleveland, OH, USA). Cells were cultured in Dulbecco’s Modified Eagle’s Medium (Invitrogen, Waltham, MA, USA) supplemented with 10% fetal bovine serum, 100 U/mL penicillin G sodium, 100 μg/mL streptomycin sulfate, and 100 μg/mL gentamicin in an atmosphere of 95% humidified air and 5% CO2 at 37°C. Cells were passaged regularly with trypsin/ethylenediaminetetraacetic acid and subcultured in 3 mL dishes prior to treatment.
Electrophysiological experiments and data analysis
In Xenopus oocytes, hERG currents were obtained using the double-electrode voltage clamp technique as reported before.19 Tip resistances of microelectrodes ranged from 1 to 5 MΩ. Data were low-pass filtered at 1–2 kHz (−3 dB, four-pole Bessel filter) before digitalization at 5–10 kHz. Recordings were performed using a commercially available amplifier (OC-725A, Warner Instruments, Hamden, CT, USA) and the pCLAMP software package (Molecular Devices, Sunnyvale, CA, USA). For control patch-clamp experiments, cells were transferred from the incubator into a recording chamber which was continuously superfused with bath solution. Single cells were selected for measurements. For studying the effect of midazolam on hERG, HEK cells were incubated with different midazolam concentrations for 30 minutes before they were transferred to the recording chamber. Pipettes with resistances of 3–4 MΩ and a RK-400 amplifier (Bio-Logic SAS, Claix, France) were used for current measurements. Data were analyzed using the pCLAMP software package. Concentration-response curves were fitted with a Hill function: Y = A1+ {[A2− A1]/[1+10^ log(X0 − X)*nH]} with A1 being the bottom asymptote, A2 being the top asymptote, logX0 being the center, and nH being the Hill slope. Activation and inactivation curves were fitted with a Boltzmann function: Y = {{(A1− A2)/[1+ (e^(X − X0)/k)]} + A2}, with A1 being the initial value, A2 being the final value, X0 representing the half-maximal activation potential, Y being the degree of activation, and k being the slope factor.
Statistical data are expressed as the mean ± standard error, with n representing the number of experiments performed. The independent Student’s t-test or analysis of variance using the Bonferroni correction were used to evaluate statistical significance. Differences were considered to be statistically significant when the P-value was <0.05.
Western blot analysis
The effects of midazolam on hERG channel trafficking were analyzed as reported before.20 Briefly, a HEK cell line stably expressing hERG channels was used for analysis. Increasing midazolam concentrations (1, 3, 10, 30, and 100 μM) as well as 100 μM As2O3, which served as a positive control, were diluted in Dulbecco’s Modified Eagle’s Medium (Invitrogen). Cells were incubated with each solution at 37°C for 24 hours prior to protein extraction. For control measurements, pure Dulbecco’s Modified Eagle’s Medium was used. Cells were washed with phosphate-buffered saline and subsequently harvested with RIPA buffer (50 mM Tris, 150 mM NaCl, 1 mM ethylenediaminetetraacetic acid, and 1% Triton) supplemented with 1 mM phenylmethylsulfonyl fluoride and complete protease puffer (Roche, Germany). Using the bicinchoninic acid (BCA) technique (Pierce BCA Protein Assay Kit, Thermo Scientific, Waltham, MA, USA), the individual protein concentration was quantified. Proteins (20 μg per sample) were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (8% polyacrylamide) and transferred to a polyvinylidene fluoride membrane. For detecting hERG bands, blots were probed with a rabbit polyclonal anti-Kv11.1 antibody (Alomone Labs, Jerusalem, Israel, 1:400) and signals were detected by chemiluminescence. Blots were quantified using the gel analysis feature of ImageJ (National Institutes of Health, Bethesda, MD, USA).
Results
Midazolam inhibits heterologously expressed hERG channels
To investigate the inhibitory effects of midazolam on cloned hERG potassium channels, double-electrode voltage clamp experiments were performed in Xenopus oocytes heterologously expressing hERG channels. From a holding potential of −80 mV, variable test pulses from −70 mV to +70 mV in 10 mV increments (2,000 msec) were applied. This first variable voltage step was followed by a constant return pulse to −50 mV (2,000 msec) eliciting typical outward tail currents. Having obtained a control measurement (Figure 1A), cells were incubated with 200 μM midazolam and the measurement was repeated (Figure 1B). Application of 200 μM midazolam significantly reduced the hERG activation current by 43.0%±3.4% and the hERG tail current amplitude by 52.1%±1.9% (n=6). Figure 1C depicts the current-voltage relationship (I–V curve) established at the end of the first pulse (at first dashed line in Figure 1A and B). In Xenopus oocytes, hERG channel activation occurred at a membrane potential of approximately −30 mV. The hERG activation current reached its maximum at approximately +20 mV. As a measure of channel activation, Figure 1D displays the current voltage relationship of peak tail current amplitude (established at the second dashed line in Figure 1A and B) versus the potential of the preceding voltage step. In order to obtain the half-maximal activation voltage, a Boltzmann function was fitted to the data, yielding a half-maximal activation voltage of −9.2±0.59 mV for control conditions and −12.6±1.0 mV after midazolam incubation. There was a small shift of the half-maximal activation potential towards more negative potentials (P<0.05, n=6).
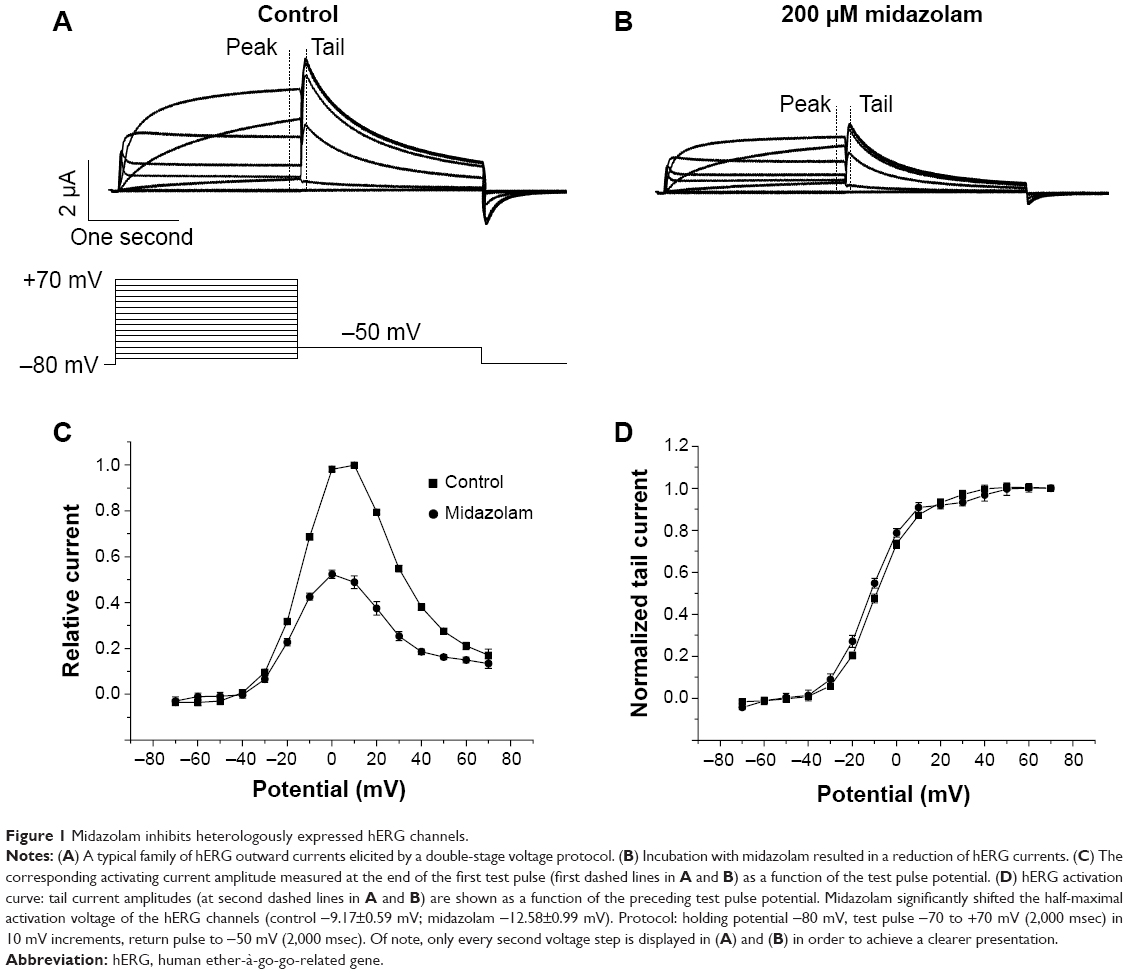 | Figure 1 Midazolam inhibits heterologously expressed hERG channels. |
Concentration dependence of midazolam-induced hERG block
To study the concentration dependence of block, cells were incubated with increasing midazolam concentrations. Starting from a holding potential of −80 mV, a variable test pulse to between −70 mV and +70 mV (10 mV increment, 400 msec) was applied. Each pulse was followed by a constant return pulse to −120 mV (400 msec) eliciting large inward tail currents. An exemplary current trace (step to +70 mV) is displayed in Figure 2A. Increasing midazolam concentrations (10, 30, 100, 200, 300, and 1,000 μM) resulted in a reduction of peak hERG current by 0.01%±9.12%, 20.89%±15.5%, 41.1%±18.66%, 51.13%±11.41%, 66.91%±9.18%, and 87.57%±3.45%, respectively (n=6–9; Figure 2B). When fitted with a Hill function, the IC50 was 170±22.3 μM (nH =1.0). In order to analyze the physiological relevance of our findings, we next analyzed the inhibitory effects of midazolam on hERG channels stably expressed in a mammalian cell line (HEK). Using the whole-cell patch clamp technique, current density was obtained. Starting from a holding potential of −80 mV, cells were depolarized by a first voltage step (400 msec, 90 mV) that was followed by a return pulse to −120 mV (400 msec) to elicit large tail currents. Again, inhibitory effects of increasing midazolam concentrations (1, 10, 30, and 100 μM) were analyzed. For instance, 10 and 30 μM midazolam reduced the hERG tail current by 48.69%±3.68% and 63.26%±2.41%, respectively n=6–7; Figure 2C). A representative current trace under control conditions and after incubation with 10 μM midazolam is displayed in the inset. In the HEK cell line, the IC50 of inhibition yielded 13.6±30 μM (nH =0.4±0.36, n=6–7).
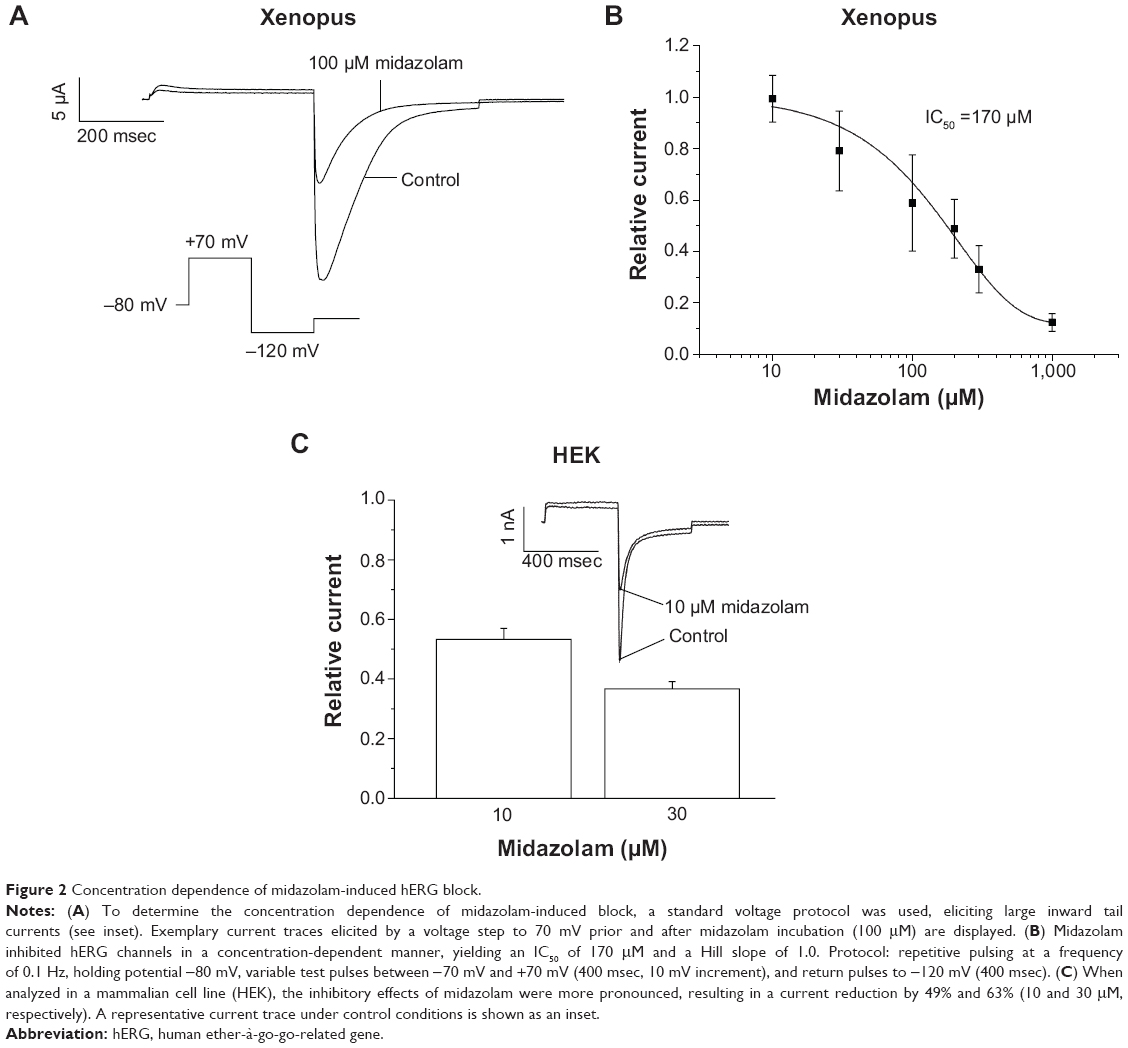 | Figure 2 Concentration dependence of midazolam-induced hERG block. |
Effects of midazolam on channel inactivation
The effects of midazolam on channel inactivation were analyzed using a double-step voltage protocol as reported previously.21,22 A first depolarizing step to +40 mV served to fully activate and inactivate the channels. This step was followed by a variable return pulse to potentials between −140 mV and +80 mV (500 msec duration, 20 mV increment; Figure 3A, inset). During this second voltage step, channels quickly redistributed to their activated and inactivated states (Figure 3A and B). In order to correct for channel deactivation, single exponentials or double exponentials were fitted to the deactivating current traces and extrapolated to the beginning of the second voltage step (see dashed lines and arrows in Figure 3A). Figure 3C displays the current-voltage relationship of corrected tail current amplitudes under control conditions and after incubation with 200 μM midazolam. When dividing the current amplitude by the corresponding electrochemical driving force, steady-state inactivation curves could be established.22 Inactivation curves for control conditions and after midazolam incubation are displayed in Figure 3D. When fitted with a Boltzmann function, the half-maximal inactivation voltage could be obtained, yielding a V1/2 of −29.2±3.8 mV for control conditions and −31.3±13.4 mV for midazolam incubation. There was no significant difference between these values (n=6, P<0.05).
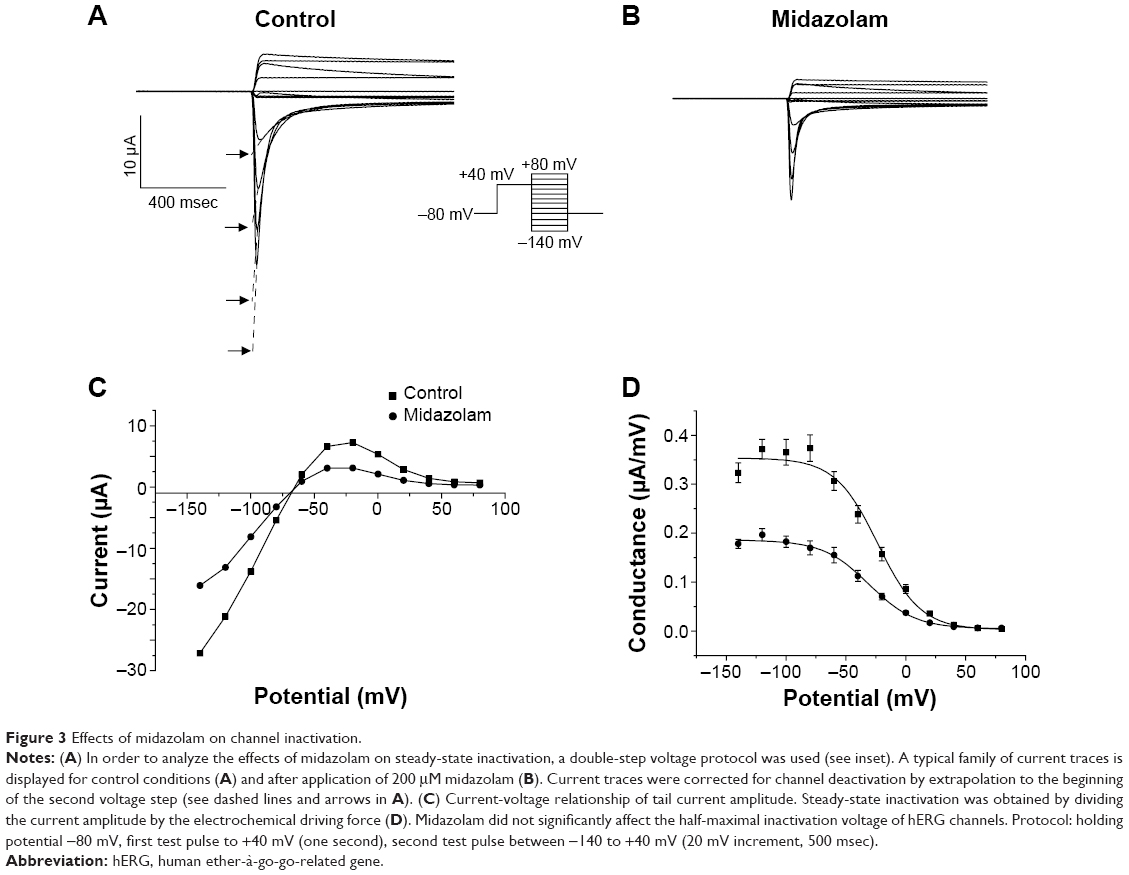 | Figure 3 Effects of midazolam on channel inactivation. |
State dependence of midazolam-induced hERG block
For most drugs tested so far, open channel block of hERG has been suggested.23 As published previously, two different voltage protocols were applied in order to qualitatively analyze the state dependence of hERG channel inhibition.20,24,25 For each voltage protocol, a measurement before and after drug incubation was performed, and development of block was calculated by division. First, using a long depolarizing voltage step to 0 mV (6 seconds), cells were depolarized (Figure 4A). Having obtained a control measurement, cells were kept in their closed state at a holding potential of −80 mV. Cells were then incubated with 200 μM midazolam for 30 minutes and the protocol was repeated. The time course of block development was calculated by division and evaluated qualitatively (Figure 4B, representative experiment of n=7). The time course of inhibition appeared in a typical time-dependent manner characteristic for an open channel inhibitor.
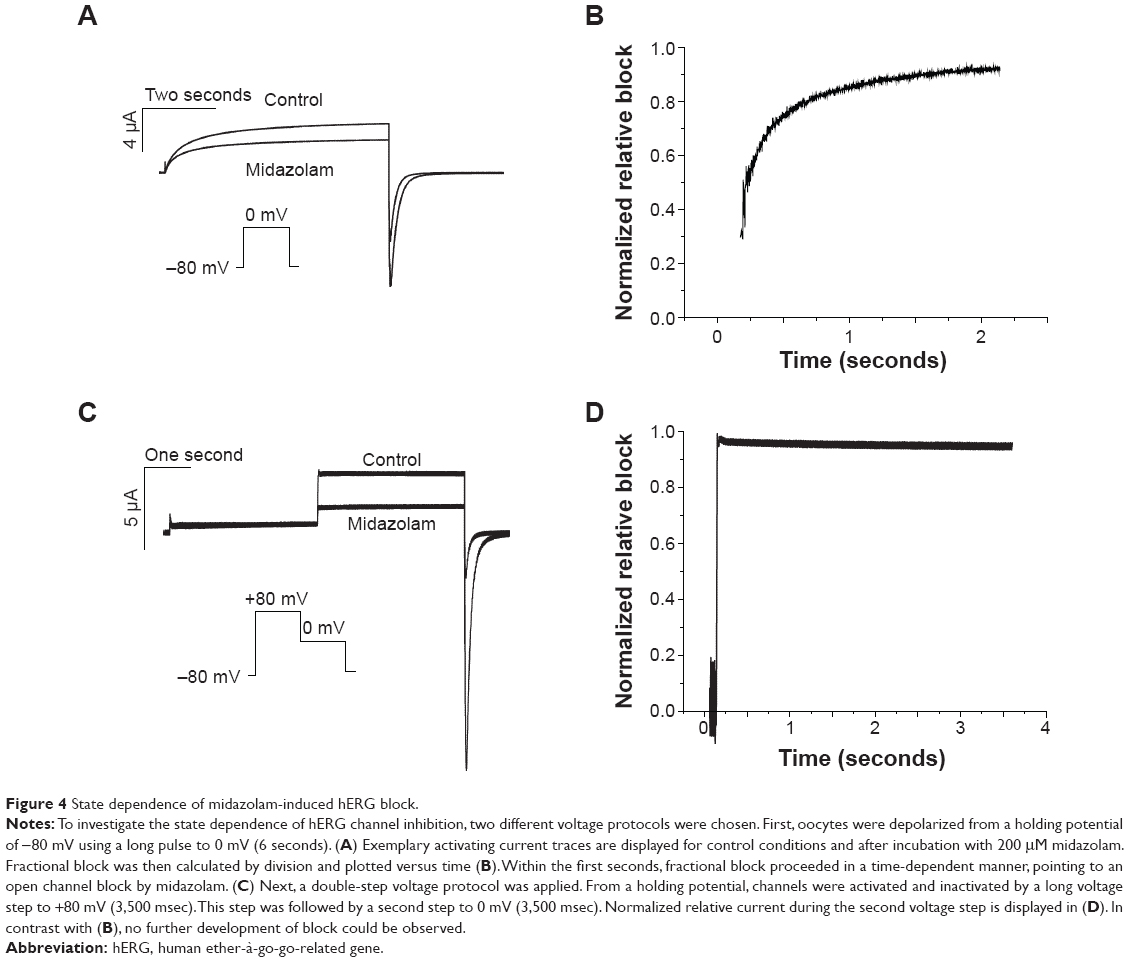 | Figure 4 State dependence of midazolam-induced hERG block. |
Next, a second voltage protocol was applied to analyze whether midazolam is able to inhibit hERG channels in their inactivated states. Cells were first inactivated by a long voltage step to +80 mV (3.5 seconds). A return pulse to 0 mV (3.5 seconds) then served to reactivate channels. A typical current trace before and after incubation with 200 μM midazolam is displayed in Figure 4C (representative experiment of n=8). Again, fractional block (during the second voltage step) was calculated by division and evaluated by qualitative assessment (Figure 4D). Interestingly, no time dependence of block development could be observed during the process of channel reopening, thereby underlining the hypothesis that midazolam blocks hERG channels in the open and inactivated but not closed states.
Onset and frequency dependence of block
To obtain the time course of block development under midazolam application, test pulses to +30 mV (400 msec) were applied at a frequency of 0.033 Hz, each followed by a return pulse to −60 mV (400 msec) to elicit outward tail currents. Peak tail current amplitude was followed to quantify the time dependence of block. After a control period of 1–2 minutes, 300 μM midazolam was applied (Figure 5A). Onset of block was rapidly reaching half maximal inhibition approximately 4 minutes after drug application (Figure 5A). These results underline that the chosen incubation period of 30 minutes is well suited for analysis of the inhibitory effects of midazolam. To further analyze the frequency dependence of block, cells were subject to repetitive pulsing at different frequencies (1, 0.5, and 0.33 Hz) for 60 seconds. From a holding pulse of −80 mV, a depolarizing voltage step to +20 mV (300 msec) served to activate hERG channels. A return pulse to −40 mV (300 msec) then followed to elicit tail currents. Having obtained a control measurement, the oocyte was exposed to 200 μM midazolam for 30 minutes and the protocol was repeated. Peak tail currents before and after incubation were quantified (n=5). A representative trace before and after midazolam incubation is shown as an inset in Figure 5B. Relative current after midazolam inhibition was calculated for every fifth second. Midazolam-induced inhibition did not exhibit significant frequency dependence (n=5, P>0.05).
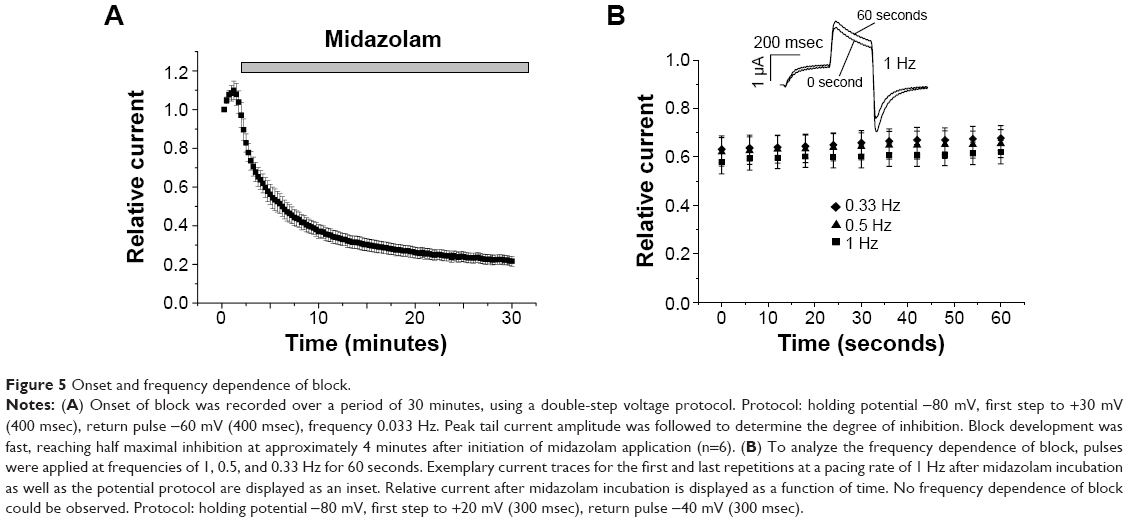 | Figure 5 Onset and frequency dependence of block. |
S6 domain mutations F656A and Y652A attenuate inhibitory effects of midazolam
The two aromatic amino acids F656 and Y652 located in the S6 domain of hERG play a major role in the binding properties of most hERG inhibitors.18,26 The inhibitory effects of 200 μM midazolam on wild-type hERG and on both hERG mutants were compared using a standard double-step voltage protocol (see inset in Figure 6A). Under control conditions (incubation with standard external solution) a run up to 117.89%±18.42% could be observed (Figure 6D, n=7). In wild-type hERG, the effect of midazolam caused a current reduction to 48.87%±4.03% of initial tail current amplitude (Figure 6A, D, n=9). As expected, inhibition was significantly attenuated for Y652A and F656A hERG, yielding a reduction of only 18.65%±5.18% for Y652A and even a run up of 4.2%±9.88% for F656A (Figure 6B–D, n=6).
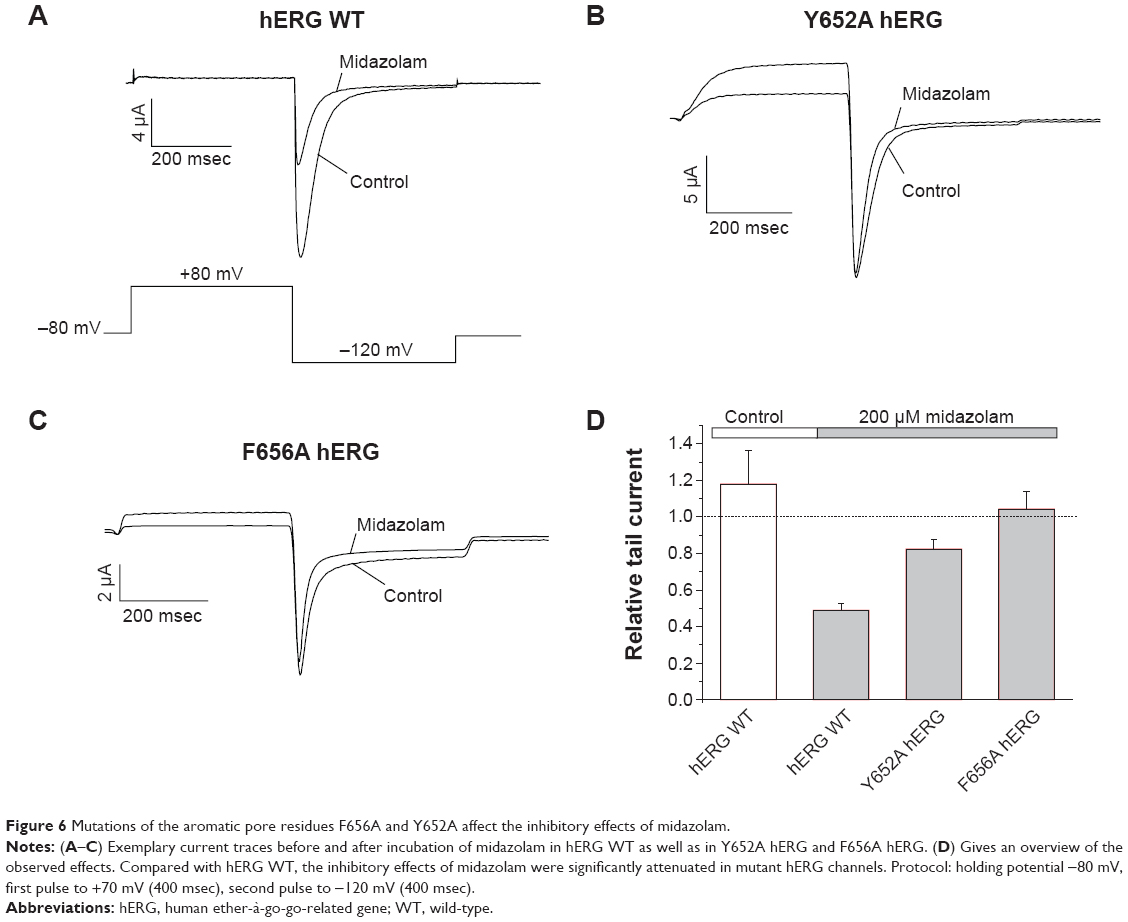 | Figure 6 Mutations of the aromatic pore residues F656A and Y652A affect the inhibitory effects of midazolam. |
Midazolam does not affect hERG channel cell surface expression
Several inhibitors of hERG channels have been associated with a reduction in the surface expression of hERG channels.20,27 To analyze the effects of midazolam on hERG channel trafficking, we used the Western blot technique. HEK cells stably expressing hERG channels were incubated for 24 hours with either control medium, 100 μM As2O3 as a positive control, or increasing midazolam concentrations (1, 3, 10, 30, 100 μM). Figure 7 (upper panel) shows a representative result of Western blot analysis. Under control conditions, two protein bands could be observed: a first band at 135 kDa representing the core glycosylated hERG protein and a second band at 155 kDa representing complex glycosylated hERG protein. To determine the effect of midazolam on hERG channel surface expression, the amount of complex glycosylated hERG protein was divided by the amount of core glycosylated hERG protein. Compared with control conditions, treatment with As2O3 (100 μM) resulted in a strong reduction of channel surface expression by 77.83%±3.4% (Figure 7, lane 1, n=3 blots). In contrast, incubation with increasing midazolam concentrations (1, 3, 10, 30, 100 μM) did not result in a significant reduction of the 155 kDa hERG (Figure 7, lines 3–8, n=3, P<0.05, analysis of variance).
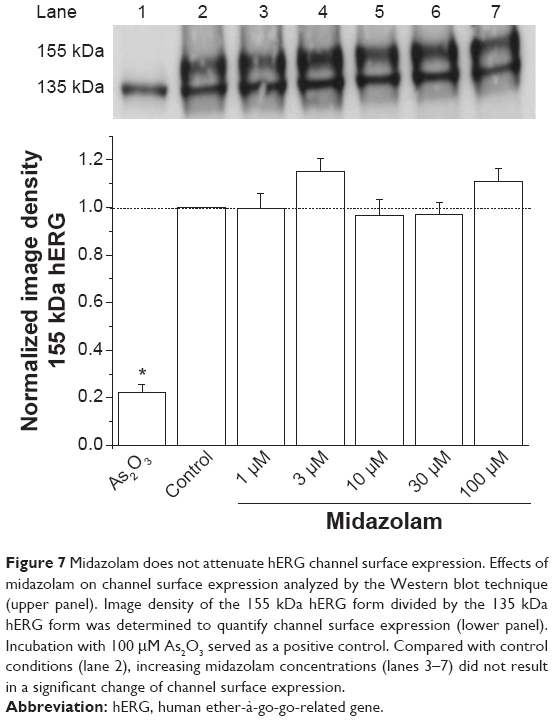 | Figure 7 Midazolam does not attenuate hERG channel surface expression. Effects of midazolam on channel surface expression analyzed by the Western blot technique (upper panel). Image density of the 155 kDa hERG form divided by the 135 kDa hERG form was determined to quantify channel surface expression (lower panel). Incubation with 100 μM As2O3 served as a positive control. Compared with control conditions (lane 2), increasing midazolam concentrations (lanes 3–7) did not result in a significant change of channel surface expression. |
Discussion
To the best of our knowledge, this is the first study analyzing the acute and subacute effects of the anesthetic midazolam on heterologously expressed hERG channels. Midazolam exhibits characteristics of a typical open channel inhibitor without relevant effects on hERG channel gating kinetics. Further, channel surface expression was not significantly affected by midazolam. These results add to the understanding of the pharmacological profile of midazolam.
Biophysical properties of hERG block by the anesthetic midazolam
We found that midazolam inhibits cloned hERG channels in a dose-dependent manner, yielding an IC50 of 170 μM in Xenopus oocytes. This concentration may appear quite high. However, it has to be considered that concentrations used in Xenopus oocytes are known to be up to 30 times higher than those needed in mammalian cells in order to achieve comparable effects. This phenomenon is well appreciated and has been mainly attributed to diffusion barriers caused by the vitteline membrane and yolk sac of Xenopus oocytes.28 To further underline the physiological relevance of our findings, we analyzed the effects of midazolam on hERG channels heterologously expressed in a mammalian cell line (HEK). As expected, the inhibitory effect was more pronounced in these cells, yielding an IC50 of 13.6 μM. Our results are in line with previously published data regarding the pharmacological effects of midazolam on other ion channels. Yamakage et al found that midazolam inhibits calcium channels with an IC50 of 10 μM.15 These data correspond well to the observed effects of midazolam on the transient outward potassium current IKto (23%±3% reduction by 60 μM midazolam).16
By analyzing the pharmacological properties of inhibition, we found that midazolam causes a significant shift of the half-maximal activation voltage of hERG channels towards more negative potentials. However, this effect was small and therefore unlikely to be of physiological relevance.
There was no significant effect on the half-maximal inactivation voltage of hERG channels. Midazolam-induced hERG blockade was state-dependent, with inhibition occurring in the open and inactivated but not closed states. State dependence of block is a common finding among hERG channel inhibitors.18,27,29 Further, it has been shown by Mitcheson et al that most hERG inhibitors bind to distinct aromatic residues located within the pore cavity of hERG.18,26 For midazolam, we found that the inhibitory effects are significantly attenuated when exchanging the residues Y652 and F656 to alanine. To further analyze the exact binding properties of midazolam, an in silico docking analysis might be helpful. A novel powerful algorithm has just been published by Anwar-Mohamed et al.30 These newer computational models might serve as powerful tools to analyze and even predict hERG liability. Taken together, our results show that midazolam acts as a typical open channel inhibitor of hERG channels. Analyzing the subacute effects of midazolam on channel surface expression of hERG channels, we further found that channel trafficking is not significantly attenuated.
Clinical implications
It is well recognized that intravenous anesthetics may have cardiovascular side effects.31–34 For example, application of midazolam reduces systolic and diastolic blood pressure and increases heart rate.35 The effects of midazolam on the corrected QT interval have been analyzed in several clinical studies. Whereas Michaloudis et al did not observe any effect on the QTc interval, others, including Saarnivaara et al found that midazolam prolonged repolarization.6–8 Owczuk et al found a small increase in the QTc interval, but this was not significant.9 The inhibitory effects on cardiac hERG channels have already been observed for other anesthetics, eg, ketamine (IC50 =12.05±1.38 μM) and articaine (IC50 =224±6 μM).36–38 However, the inhibitory effects of midazolam on heterologously expressed hERG channels have not previously been described.
In clinical practice, the midazolam dose recommended for induction of anesthesia is 0.2–0.4 mg/kg by intravenous application. Crevat-Pisano et al reported that peak concentrations of up to 1.1 μg/L can be observed, which correspond to 3.4 μM midazolam.39 Nevertheless, it has to be taken into account that peak plasma concentrations during induction vary widely depending on individual factors, such as metabolism, age, and sex.3 Midazolam is rapidly distributed after intravenous injection, with a distribution half-time ranging between 6 and 15 minutes.40 Due to its lipophilic properties, midazolam tends to accumulate in adipose tissue. As a result, a prolonged elimination half-life has been observed in obese patients.41 Also, for reasons that are still unknown, a small portion of the healthy population exhibits a prolonged elimination half-time of more than 7 hours.42 We found that midazolam inhibited heterologously expressed cardiac hERG channels in a mammalian cell line (HEK) in a dose-dependent manner, yielding an IC50 of 13.6 μM. Considering the above-mentioned modulating factors, midazolam might result in a small inhibition of hERG channels at therapeutic dosages. Thus, from a clinical point of view, pronounced effects of midazolam on cardiac repolarization seem unlikely. These data correspond well to the reported variable and rather small effect of midazolam on the QTc interval. However, within this context, it needs to be mentioned that midazolam exerts inhibitory effects on a variety of ion channels that might counterbalance or even aggravate its pharmacological effects on cardiac electrophysiology.
Conclusion
In the present study, we analyzed the inhibitory effects of the anesthetic midazolam on cloned hERG channels. We found that midazolam acts as a low-affinity inhibitor of hERG channels without additional effects on channel surface expression. These findings add to the current understanding of the pharmacological profile of this compound.
Acknowledgments
This work was supported by grants from DFG Zi1177/1-1, Scho1350/1-1, and Scho1350/2-1 (to EZ and EPS). FF is supported by the Hartmut Hoffmann-Berling International Graduate School of Molecular and Cellular Biology. The skilled assistance of Christine Jeckel is gratefully acknowledged.
Disclosure
The authors report no conflicts of interest in this work.
References
Kain ZN, Mayes LC, Bell C, Weisman S, Hofstadter MB, Rimar S. Premedication in the United States: a status report. Anesth Analg. 1997;84(2):427–432. | ||
Nordt SP, Clark RF. Midazolam: a review of therapeutic uses and toxicity. J Emerg Med. 1997;15(3):357–365. | ||
Reves JG, Fragen RJ, Vinik HR, Greenblatt DJ. Midazolam: pharmacology and uses. Anesthesiology. 1985;62(3):310–324. | ||
Kanto JH. Midazolam: the first water-soluble benzodiazepine. Pharmacology, pharmacokinetics and efficacy in insomnia and anesthesia. Pharmacotherapy. 1985;5(3):138–155. | ||
Saari TI, Uusi-Oukari M, Ahonen J, Olkkola KT. Enhancement of GABAergic activity: neuropharmacological effects of benzodiazepines and therapeutic use in anesthesiology. Pharmacol Rev. 2011;63(1):243–267. | ||
Saarnivaara L, Klemola UM, Lindgren L, Rautiainen P, Suvanto A. QT interval of the ECG, heart rate and arterial pressure using propofol, methohexital or midazolam for induction of anaesthesia. Acta Anaesthesiol Scand. 1990;34(4):276–281. | ||
Michaloudis DG, Kanakoudis FS, Petrou AM, Konstantinidou AS, Pollard BJ.The effects of midazolam or propofol followed by suxamethonium on the QT interval in humans. Eur J Anaesthesiol. 1996;13(4):364–368. | ||
Michaloudis DG, Kanakoudis FS, Xatzikraniotis A, Bischiniotis TS. The effects of midazolam followed by administration of either vecuronium or atracurium on the QT interval in humans. Eur J Anaesthesiol. 1995;12(6):577–583. | ||
Owczuk R, Twardowski P, Dylczyk-Sommer A, et al. Influence of promethazine on cardiac repolarisation: a double-blind, midazolam-controlled study. Anaesthesia. 2009;64(6):609–614. | ||
Curran ME, Splawski I, Timothy KW, Vincent GM, Green ED, Keating MT. A molecular basis for cardiac arrhythmia: HERG mutations cause long QT syndrome. Cell. 1995;80(5):795–803. | ||
Kiehn J, Lacerda AE, Brown AM. Pathways of HERG inactivation. Am J Physiol. 1999;277(1 Pt 2):H199–H210. | ||
Roden DM, Balser JR, George AL, Anderson ME. Cardiac ion channels. Annu Rev Physiol. 2002;64:431–475. | ||
Sanguinetti MC, Curran ME, Spector PS, Keating MT. Spectrum of HERG K+-channel dysfunction in an inherited cardiac arrhythmia. Proc Natl Acad Sci U S A. 1996;93(5):2208–2212. | ||
Viskin S. Long QT syndromes and torsade de pointes. Lancet. 1999;354(9190):1625–1633. | ||
Yamakage M, Matsuzaki T, Tsujiguchi N, Honma Y, Namiki A. Inhibitory effects of diazepam and midazolam on Ca2+ and K+ channels in canine tracheal smooth muscle cells. Anesthesiology. 1999;90(1):197–207. | ||
Buljubasic N, Marijic J, Berczi V, Supan DF, Kampine JP, Bosnjak ZJ. Differential effects of etomidate, propofol, and midazolam on calcium and potassium channel currents in canine myocardial cells. Anesthesiology. 1996;85(5):1092–1099. | ||
Vonderlin N, Fischer F, Zitron E, et al. Inhibition of cardiac Kv1.5 potassium current by the anesthetic midazolam: mode of action. Drug Des Devel Ther. 2014;8:2263–2271. | ||
Scholz EP, Zitron E, Kiesecker C, et al. Drug binding to aromatic residues in the HERG channel pore cavity as possible explanation for acquired long QT syndrome by antiparkinsonian drug budipine. Naunyn Schmiedebergs Arch Pharmacol. 2003;368(5):404–414. | ||
Fischer F, Vonderlin N, Zitron E, et al. Inhibition of cardiac Kv1.5 and Kv4.3 potassium channels by the class Ia anti-arrhythmic ajmaline: mode of action. Naunyn Schmiedebergs Arch Pharmacol. 2013;386(11):991–999. | ||
Scherer D, Hassel D, Bloehs R, et al. Selective noradrenaline reuptake inhibitor atomoxetine directly blocks hERG currents. Br J Pharmacol. 2009;156(2):226–236. | ||
Scholz EP, Niemer N, Hassel D, et al. Biophysical properties of zebrafish ether-à-go-go related gene potassium channels. Biochem Biophys Res Commun. 2009;381(2):159–164. | ||
Vandenberg JI, Varghese A, Lu Y, Bursill JA, Mahaut-Smith MP, Huang CL-H. Temperature dependence of human ether-a-go-go-related gene K+ currents. Am J Physiol, Cell Physiol. 2006;291(1):C165–C175. | ||
Redfern WS, Carlsson L, Davis AS, et al. Relationships between preclinical cardiac electrophysiology, clinical QT interval prolongation and torsade de pointes for a broad range of drugs: evidence for a provisional safety margin in drug development. Cardiovasc Res. 2003;58(1):32–45. | ||
Scholz EP, Konrad FM, Weiss DL, et al. Anticholinergic antiparkinson drug orphenadrine inhibits HERG channels: block attenuation by mutations of the pore residues Y652 or F656. Naunyn Schmiedebergs Arch Pharmacol. 2007;376(4):275–284. | ||
Scholz EP, Alter M, Zitron E, et al. In vitro modulation of HERG channels by organochlorine solvent trichlormethane as potential explanation for proarrhythmic effects of chloroform. Toxicol Lett. 2006;165(2):156–166. | ||
Mitcheson JS, Chen J, Lin M, Culberson C, Sanguinetti MC. A structural basis for drug-induced long QT syndrome. Proc Natl Acad Sci U S A. 2000;97(22):12329–12333. | ||
Fischer F, Vonderlin N, Seyler C, et al. Acute and subacute effects of the selective serotonin-noradrenaline reuptake inhibitor duloxetine on cardiac hERG channels. Naunyn Schmiedebergs Arch Pharmacol. 2013;386(9):795–804. | ||
Madeja M, Musshoff U, Speckmann EJ. Follicular tissues reduce drug effects on ion channels in oocytes of Xenopus laevis. Eur J Neurosci. 1997;9(3):599–604. | ||
Zitron E, Karle CA, Wendt-Nordahl G, et al. Bertosamil blocks HERG potassium channels in their open and inactivated states. Br J Pharmacol. 2002;137(2):221–228. | ||
Anwar-Mohamed A, Barakat KH, Bhat R, et al. A human ether-á-go-go-related (hERG) ion channel atomistic model generated by long supercomputer molecular dynamics simulations and its use in predicting drug cardiotoxicity. Toxicol Lett. 2014;230(3):382–392. | ||
De Hert SG, Vermeyen KM, Adriaensen HF. Influence of thiopental, etomidate, and propofol on regional myocardial function in the normal and acute ischemic heart segment in dogs. Anesth Analg. 1990;70(6):600–607. | ||
Gelman S, Reves JG, Harris D. Circulatory responses to midazolam anesthesia: emphasis on canine splanchnic circulation. Anesth Analg. 1983;62(2):135–139. | ||
Giese JL, Stockham RJ, Stanley TH, Pace NL, Nelissen RH. Etomidate versus thiopental for induction of anesthesia. Anesth Analg. 1985;64(9):871–876. | ||
Reves JG, Kissin I, Fournier S. Negative inotropic effects of midazolam. Anesthesiology. 1984;60(5):517–518. | ||
Forster A, Gardaz JP, Suter PM, Gemperle M. I.V. midazolam as an induction agent for anaesthesia: a study in volunteers. Br J Anaesth. 1980;52(9):907–911. | ||
Siebrands CC, Friederich P. Inhibition of HERG channels by the local anaesthetic articaine. Eur J Anaesthesiol. 2007;24(2):148–153. | ||
Sintra Grilo L, Carrupt P-A, Abriel H, Daina A. Block of the hERG channel by bupivacaine: electrophysiological and modeling insights towards stereochemical optimization. Eur J Med Chem. 2011;46(8):3486–3498. | ||
Zhang P, Xing J, Luo A, et al. Blockade of the human ether-a-go-go-related gene potassium channel by ketamine. J Pharm Pharmacol. 2013;65(9):1321–1328. | ||
Crevat-Pisano P, Dragna S, Granthil C, Coassolo P, Cano JP, Francois G. Plasma concentrations and pharmacokinetics of midazolam during anaesthesia. J Pharm Pharmacol. 1986;38(8):578–582. | ||
Allonen H, Ziegler G, Klotz U. Midazolam kinetics. Clin Pharmacol Ther. 1981;30(5):653–661. | ||
Greenblatt DJ, Abernethy DR, Locniskar A, Harmatz JS, Limjuco RA, Shader RI. Effect of age, gender, and obesity on midazolam kinetics. Anesthesiology. 1984;61(1):27–35. | ||
Dundee JW, Halliday NJ, Harper KW, Brogden RN. Midazolam. A review of its pharmacological properties and therapeutic use. Drugs. 1984;28(6):519–543. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.