")
Back to Journals » International Journal of General Medicine » Volume 14
An Update in Our Understanding of the Relationships Between Gene Polymorphisms and Chemotherapy-Induced Nausea and Vomiting
Authors Jin Y, Li X, Jiang C, Zhao J, Liu G, Li H, Jin G, Li Q
Received 13 July 2021
Accepted for publication 1 September 2021
Published 18 September 2021 Volume 2021:14 Pages 5879—5892
DOI https://doi.org/10.2147/IJGM.S329257
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Scott Fraser
Yilan Jin,* Xiaorong Li,* Caihong Jiang,* Jun Zhao, Guang Liu, Hui Li, Gaowa Jin, Quanfu Li
Department of Medical Oncology, Ordos Central Hospital, Ordos, 017000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Quanfu Li; Gaowa Jin
Department of Medical Oncology, Ordos Central Hospital, 23th Yinjihuoluo Western Road, Ordos, 017000, People’s Republic of China
Tel +86 0477-8363279
Fax +86 0477-8363158
Email [email protected]; [email protected]
Abstract: The occurrence and severity of chemotherapy-induced nausea and vomiting (CINV) are influenced by many factors; this includes therapeutic factors, such as the dose, administration mode, and chemotherapeutic agent emetogenicity, as well as patient-related risk factors, such as the gender, age, alcohol consumption history, and anxiety level. However, these factors cannot fully explain the individual CINV differences. In recent years, the correlation between gene polymorphism and CINV has been a hot research topic; the present paper reviews current research on CINV-related gene polymorphisms, and the results indicate that the use of gene polymorphism for the optimization of CINV efficacy is of important clinical significance.
Keywords: chemotherapy-induced nausea and vomiting, gene polymorphisms, prevention and treatment, antiemetic
Introduction
Nausea and vomiting are common adverse reactions to chemotherapy. If untreated, chemotherapy-induced nausea and vomiting (CINV) can affect 70–80% of patients undergoing chemotherapy; this may lead to complications, such as water and electrolyte disturbance, quality of life reduction, and treatment interruption.1,2 CINV is influenced by complex metabolic and gene-environment interactions, which leads to inter-individual differences in its occurrence and severity. It is influenced by a variety of factors; this includes therapeutic factors, such as the chemotherapeutic agent emetogenicity, dose, administration route, and various environmental triggers, as well as patient-related factors, such as the gender, age, alcohol consumption history, and anxiety level.3 With the development of technology, gene polymorphism detection has become possible. New studies and discoveries based on gene polymorphism suggest that it may be an independent CINV influencing factor in patients.4 All gene polymorphisms involved in CINV and related pharmacogenetic mechanisms, such as genetic drug transporter variation, drug metabolic enzymes, and drug receptors, may affect patient CINV occurrence and severity. The present study aims to briefly review several relatively well-established gene polymorphism findings associated with their different, sometimes paradoxical, effects on CINV.
Methods
Literature Search
Pieces of electronic literature published from July 2020 to July 2021 were obtained from the following databases: PubMed®, Excerpta Medica Database, China National Knowledge Infrastructure, Wanfang Data, China Science and Technology Journal Database, and Chinese BioMedical Literature Database. The reference list of related articles was consulted to find more qualified studies. The keyword combination was chemotherapy or cancer AND nausea or vomiting or antiemetics or chemotherapy-induced nausea and vomiting or chemotherapy-induced vomiting or chemotherapy-induced nausea and gene or genes or genetics or polymorphism or candidate genes or gene expression or pharmacogenetics.
The inclusion criteria were: (1) studies involving patients with cancer receiving chemotherapy agents; (2) studies with patients assessed for nausea and/or vomiting outcomes; (3) studies with genotyped patients; and (4) studies evaluating the association between genetic polymorphisms and chemotherapy-induced nausea (CIN) and/or chemotherapy-induced vomiting (CIV), and/or antiemetic effectiveness.
The exclusion criteria were: (1) studies investigating patients with post-operative nausea and vomiting; (2) studies investigating patients with radiotherapy-induced nausea and vomiting; and (3) studies with genotype associations only evaluated in the context of the pharmacokinetics.
After excluding articles with a lack of pharmacogenomic information or articles published before 2000 (the human genome was sequenced in 2000), 29 original research articles focusing on genetic polymorphisms and CINV were included in the present study.
Data Synthesis
These 29 studies were summarized using the following prespecified evaluation criteria: Author, country, year; emetogenicity of the chemotherapy regimen; sample characteristics (ie, sample size, age, gender, diagnosis, and antiemetic treatment); CINV assessment (ie, instrument[s] and CINV assessment timing); gene(s) and associated polymorphism(s) classified by function; major study outcome(s); major findings; sample characteristics; CINV assessments; CINV assessments timing; genotyping method types; specific evaluated polymorphisms; chemotherapy drug types; and the results. The above-listed data are summarized in tabular and narrative forms.
The CINV Mechanism and the Antiemetic Mechanism and Classification
Depending on the time of first chemotherapeutic drug use, CINV can be divided into acute-phase CINV and delayed-phase CINV. This is a complicated process involving neuroanatomy, central and peripheral pathways, neurotransmitters, and receptors.5 The peripheral pathway is usually characterized by acute-phase CINV;6 its mechanism is that the chemotherapeutic agents act on the enterochromaffin cells in the gastrointestinal tract mucosa, causing the release of a large amount of serotonin (5-hydroxytryptamine 3 [5-HT3]) to act on the adjacent vagus fiber corresponding receptors and leading to an afferent stimulus that terminates in the dorsal brain stem (primarily in the nucleus tractus solitarius) and subsequently activates the central pattern generator.5–8 The central pathway occurs mainly in centers of the higher central nervous system located in limbic forebrain structures, such as the amygdala, which is usually characterized by drug-induced delayed vomiting.6 Substance P is the main neurotransmitter that activates neurokinin-1 (NK-1) in the central pathway. The 5-HT3 receptors are cross-linked with the NK-1 receptors, but the mechanisms involved are not fully understood.9 The mechanism of nausea may not be identical to the mechanism of vomiting; the exact mechanism remains unclear. However, the clinical management of nausea and vomiting usually occur simultaneously.10
The interaction between different pathways and neurotransmitters leads to the development of different CINV types. Thus, to achieve maximum effectiveness, its prevention and treatment require the use of drugs targeting these pathways and transmitters.11 The most commonly used antiemetics are currently generally classified into 5-HT3 receptor antagonists (5-HT3RA), NK-1 receptor antagonists, glucocorticoids, NK-1/5-HT3 compounds, atypical antipsychotics, benzodiazepines, phenothiazines, and cannabinoid antiemetics.12–14 Among them, in addition to the 5-HT3RA (eg, ondansetron, tropisetron, palonosetron, etc.), antipsychotic drug olanzapine, NK-1 receptor antagonists (intravenous: Such as fosaprepitant, oral: Such as aprepitant), and glucocorticoids (generally refer to dexamethasone), which are standard CINV therapeutic drugs, olanzapine has been receiving an increasing amount of attention.15
CINV and Drug-Receptor Pathway-Related Gene Polymorphisms
Serotonin Pathway
Of the 5-HT receptors (5-HTRs), only 5-HTR3, which is a ligand-gated ion channel, plays a role in CINV pathogenesis.16 It is distributed in the central and peripheral systems and ultimately exerts a role in intestinal motility.17 It is composed of five subunits (5-HT3A-E) encoded by the HTR3A-E gene; these receptor subtypes cause CINV under different circumstances.18 Different subunit compositions contribute to the 5HTR3 system’s complexity.19 Cell culture experiments and clinical studies show that genetic variants of the 5-HTR3 gene are associated with altered protein function and clinical outcomes in patients with CINV.16 The standard drug for the prevention of acute CINV is 5-HT3RA; its action mechanism is binding to and selectively inhibiting 5-HT3R excitation. This inhibition occurs in the chemosensory trigger zone at the central site and in the peripheral region, thereby attenuating CINV.20 Data suggest that variants in the HTR3A-D gene may be associated with CINV as well as the therapeutic effect of 5-HT3RA in patients.16,17,19,21–29
Kaiser et al sequenced and performed a haplotype analysis of the HTR3A gene in 233 patients on highly emetogenic chemotherapy regimens; the results showed that HTR3A CT haplotype carriers (unreported rs IDs) were less likely to develop CINV after prophylactic antiemetic treatment (Chi-squared test, p = 0.01).21 Certain studies analyzed the relationship between some common HTR3A loci (rs1062613, rs1176722, rs1176719, rs2276303, rs909411, and rs1176713) and CINV; no correlation was discovered.22,23 It would appear that due to the paucity of relevant studies and the unclear nature of the predictive gene, the single genotype of 5-HTR3A cannot be used as a genetic predictor for the antiemetic efficacy of 5-HT3RA.
A common polymorphism of the 5-HT3B subunit is −100_-102AAG deletion. Tremblay et al hypothesized that HTR3B gene polymorphisms might have an impact on acute CINV and the therapeutic effects of 5HT3RA. They sequenced the entire 5-HTR3B and found that among patients with CINV, homozygous patients with −100_-120AAG deletion had a higher frequency of vomiting than other patients in the recessive model (Chi-squared test, n = 242, p < 0.02).24 However, only 1.2% of the patients were homozygotes for this allele; thus, only a small proportion of treatment failures could be explained by this variant.19 Kang et al reached the same conclusion in Korean patients in the dominant model who were prophylactically administered ramosetron (Mann–Whitney U-test, n = 50, p = 0.02).17 Furthermore, domestic researchers Cao Jiayan et al found that the TT genotype incidence at rs3758987 was significantly lower than the non-TT genotype incidence (Logistic regression, n = 212, p = 0.014).25 However, a 2021 research by Tsuji et al found no connection between the two.26 Other studies failed to find any correlation between CINV and the loci of HTR3B rs1176744, rs4938058, rs7943062, rs12795805, or rs2276305.17–19,21,24,25,27,30,33 Although there are more studies on the polymorphism of the HTR3B gene than on any other subunit gene, the heterogeneity of research on ethnicity and polymorphism distribution may lead to different results. Hence, the meta-analysis of the primary and secondary outcomes in certain studies has little significance.32
Fasching et al analyzed the common non-synonymous single nucleotide polymorphism (SNP) of 5HTR3 and found that a rare C allele homozygote at the HTR3C rs6766410 locus was correlated with acute phase vomiting (Chi-squared tests, n = 110, p = 0.01).16 In contrast, Pud et al reported that, in a recessive model, a rare N allele homozygote at the same locus was correlated with a reduction in the severity of delayed CIN (Cochran-Mantel-Haenszel test, n = 110, p = 0.04).27 This SNP causes a change in the amino acid sequence from lysine to arginine, which may alter the HTR3C structure. However, a study by Ward et al and Tsuji et al found no correlation between the seven SNPs (ie, 1651C>T, 3885C>T, 3894C>A, 6342C>T, 7051G>A, 7082C>T, and 7142G>C, 1214C>G) or rs6766410 with CINV.19,26 Although the 4 studies included the same locus, three different results emerged. This may have been caused by the smaller sample size available for evaluation, as there is a low number of relevant studies due to the lack of focus on this particular locus. Larger scale studies are needed to clarify the correlation.
Hammer et al showed that rs6443930, a rare C allele homozygote of HTR3D, might be correlated with enhanced antiemetic efficacy for 5-HT3RA (Kaplan Meier log rank test, n = 110, p = 0.048).22 However, Tsuji et al did not find any correlation between the two.26 At the time that the present review was written, no other articles investigating the correlation between HTR3D and CINV were identified.
The serotonin transporter (SERT) can terminate the action of 5-HT through its rapid uptake from a corresponding site. In a retrospective analysis of 166 patients with colorectal cancer conducted in Southern China, Wei Yisheng et al found that moderate–severe CINV was associated with the rs956304 genotype of the SERT promoter polymorphic locus (Chi-squared test, n = 166, p = 0.011), which was independent of the rs1176744, rs12795805, and rs2276305 loci,30 and that the rs956304 AG+GG genotype was an independent risk factor for the development of moderate–severe CINV in patients with colorectal cancer. This study provided a new perspective and concept for the study of CINV-related gene polymorphism in China.
NK-1 Receptor Pathway
Aprepitant is a NK-1 receptor antagonist that acts on the area postrema of the brain and is mainly effective against delayed CINV. Recently, an association between the tachykinin receptor 1 (TACR1) gene-encoding NK1 receptor and CINV has been considered.26 There are 2 studies analyzing the relationship between certain TACR1 loci (rs2111375, rs3771836, rs6715729, rs3821313, rs3755468, Y192H, and rs17838409) and CINV.23,26 Tsuji et al found that rs3755468TT was a significant complete response (CR) determinant in delayed-phase CINV (Logistic regression, n = 166, p = 0.014).26
CINV and Drug Metabolism Pathway-Related Gene Polymorphisms
Cytochrome P4502D6 (CYP2D6) is one of the most important drug-metabolizing enzymes in the liver. Most antiemetic drugs, including the 5-HT3RA and dopamine receptor antagonists, are metabolized by the CYP2D6 system, and its variation can lead to significant changes in the metabolism of antiemetic drugs and may ultimately affect the drug action and the occurrence of adverse reactions.33,34 The activity of CYP2D6 varies widely within the same race and may include the following patient genotypes: Ultrafast metabolizer (UM), extensive metabolizer (EM), intermediate metabolizer (IM), and poor metabolizer (PM). The distribution of CYP2D6 in different ethnic groups is also very different, resulting in different PM, IM, EM, and UM percentages in specific populations. For example, CYP2D6 UM is rare among the Chinese, only accounting for approximately 1% of the total population.35
Kaiser et al investigated the influence of the CYP2D6 genotype on the therapeutic effects of 5-HT3RA and found that the serum concentration of tropisetron was higher in patients with the CYP2D6 PM allele (p < 0.03) and that the incidence of acute CIV was increased in UM allele carriers (Chi-squared analysis, n = 270, p < 0.001, p < 0.03 for the tropisetron and ondansetron groups, respectively);36 the difference was more significant in the tropisetron group than in the ondansetron group. The researchers considered PM allele carriers to have the highest 5-HT3RA blood concentration and, therefore, the best protection against CINV. Meanwhile, the UM allele carriers were considered to have the least protection against CINV when given the standard doses. Another study found that UM allele carriers had an increased risk of developing severe CINV (Chi-squared test, n = 242, p = 0.03),24 while a different study found a decrease in the CINV complete control rate in patients carrying three active CYP2D6 alleles (ie, UMs) who received prophylactic ondansetron therapy (Chi-squared test, n = 202, p < 0.05).31 However, no significant correlation was found between the other phenotypes (rs16947, rs3892097, and rs1065852) and CINV. Despite the result discrepancy in the above-listed studies, there is still some guidance, with respect to the UM allele, for clinical practice concerning CINV.
In vitro studies have shown that cytochrome P4503A (CYP3A) enzymes are involved in the metabolism of palonosetron and aprepitant.33,37 Yokoyama et al found no correlation between CINV and CYP3A5 rs776746 in a triple antiemetic regimen consisting of palonosetron, aprepitant, and dexamethasone that is used in Japanese patients with breast cancer (Chi-squared test, n = 125, p = 1.00).29
CINV and Drug Transport Pathway-Related Gene Polymorphisms
The gene polymorphisms of drug transport proteins may be a source of variation in drug configuration and response.38 The polymorphisms of the ATP-binding cassette transporter protein B1 (ABCB1), ATP-binding cassette transporter protein G2 (ABCG2), ATP-binding cassette transporter protein C1 (ABCC1), copper transporter P-type ATP gene (ATP7B), and organic cation transporter 1 (OCT1) may alter the uptake and efflux rates of chemotherapeutic agents entering the blood–barrier, resulting in CINV differences.
ABCB1
ABCB1, or the multidrug resistance gene, encodes the P-glycoprotein, which is a trans-membrane protein that is considered an important transporter protein in limiting the accumulation of drugs, including many chemotherapeutic agents and antiemetics.39,40 Babaoglu et al determined the ABCB1 3435C>T genotype in patients and analyzed its relationship with the antiemetic effect of 5-HT3RA.39 The results showed that the rare T allele homozygote or heterozygote was associated with the reduced occurrence and severity of acute-phase CIV compared with the common C allele homozygote or heterozygote (Chi-squared test, n = 216, p = 0.04). He and Zoto reached a similar conclusion when they found that CINV severity and incidence were reduced in patients with the rare T allele homozygote compared with patients with a homozygous or heterozygous common C allele (Logistic regression, n = 215 and n = 239, p = 0.002 and p = 0.016, respectively).40,41 A meta-analysis showed that the risk of acute CINV was significantly lower in the recessive model but not significantly different in the dominant model.32 These results are different from the results of Jin et al in the 2021 study of the Chinese population. Jin et al found that the ABCB1 3435C>T TT genotype in the olanzapine group was associated with a lower, delayed TP rate in patients using the olanzapine-based triple antiemetic regimen (Chi-squared test, n = 210, p = 0.006). Meanwhile, in the aprepitant group it was associated with patients with the 2677G>T/A GG genotype (p = 0.008).42 In 2013, Tsuji et al evaluated 64 patients with breast cancer being treated with adriamycin in combination with cyclophosphamide; they concluded that the acute-phase CR rate was not correlated with 3435C>T (Cochran-Mantel-Haenszel test, n = 64, p = 0.19); however, there was a significant correlation between ABCB1 2677G>T/A polymorphism and patients achieving CR within 24 hours of chemotherapy (p = 0.047).43 In contrast, a 2017 study of 156 patients on cisplatin conducted by the same researchers yielded dissimilar results: The CR rate was significantly higher in patients with the ABCB1 3435TT genotype than in patients with the non-TT genotype (Logistic regression, p = 0.022); the CR rate was also higher in patients with the ABCB1 2677TT genotype than in other genotypes (p = 0.063).44 However, no significant CR rate difference was found in ABCB1 1236C>T gene carriers. This is consistent with the results of Jin et al.44 The results of Zoto et al, on the other hand, suggest a positive correlation between 1236C>T and CIV incidence (p = 0.027).41 Another study reported a higher incidence of grade 3/4 delayed-phase CINV in carriers of the ABCB1 gene CTG haplotype (C3435T, C1236T, and G2677T) than in other haplotypes (p < 0.05).31 A multivariate analysis conducted by He et al concluded that the CG haplotype carrier was correlated with a risk of grade 3/4 nausea and was an independent prognostic marker.40
As ABCB1 is one of the most investigated gene polymorphisms related to CINV and drug transport pathways, the discrepancy between the results of ABCB1-related studies may be caused by the high bias risk in some studies as well as the synergistic or antagonistic effects between different loci within the gene.
ABCC1, ABCG2, and ATP7B
The proteins encoded by the ABCC1 and ABCG2 genes are trans-membrane proteins; they are part of the blood–brain barrier and can cause the efflux of some chemotherapeutic agents.4 One study found that patients with missense mutations in ABCG2 rs2231142 had an increased risk of CIN; this was possibly caused by the SNPs reducing the ABCG2 exocytosis activity.45 A multivariate analysis by Yokoyama et al showed a significant correlation between the ABCG2 rs2231142 AA genotype and acute severe nausea (p = 0.049).29 However, Lamba et al failed to find any correlations among ABCC1 rs246240, rs2238476, and CINV.4
The ATP7B gene encodes the ATP7B enzyme. Higher levels of mRNA expression of ATP7B are correlated with higher rates of efflux as well as the accumulation of chemotherapeutic agents in the blood.46 Lamba et al conducted an exploratory study on SNPs in ATP7A and ATP7B and found a correlation between ATP7B rs1801244 and an increased CIN risk (Cox-proportion hazards models, n = 90, p = 0.027).4
OCT1
As one of the most abundantly expressed drug transport proteins in the human liver, OCT1 is important for its antiemetic effects. It is highly polymorphic throughout different ethnic groups, which can lead to differences in transporter function and variation in response to antiemetics. However, this remains controversial.47
One study assessed the correlation between five common OCT1 haplotype loss-of-function polymorphisms and the pharmacokinetics and therapeutic effects of 5-HTRA.47 The in vitro experiments showed that the amino acid substitutions (ie, R61C, C88R, G401S, M420del, and G465R) OCT1 polymorphisms might inhibit the tropisetron uptake. Patients with complete active OCT1 allele deletion had the highest plasma concentrations of tropisetron, at 3 hours and 6 hours after administration, and ondansetron, at 3 hours after administration (Logistic regression, n = 270, p < 0.05). Patients with active OCT1 allele deletion exhibited a higher CR rate (p = 0.007). The researchers concluded that patients with complete active OCT1 allele deletion had an impaired cellular uptake and reduced biological inactivation, resulting in prolonged drug residence time in the circulation system and better therapeutic effects.46
The above-mentioned studies suggest that polymorphisms of this drug transport pathway gene may be a CINV influencing factor. However, while our understanding of the gene is increasing, further studies are still needed to provide stronger evidence. It is hoped that OCT1 will play a key role in CINV treatment in the future.
Other Related Gene Polymorphisms and CINV
Excision Repair Cross Complementation Gene
Platinum compounds act by binding with DNA; this leads to intra-strand cross-links that disrupt DNA structure. The excision repair cross complementation gene (ERCC) is a key nucleotide excision repair pathway component, and a high expression of ERCC proteins and genes is correlated with cisplatin resistance and chemotherapy toxicity.48 Yokoi et al conducted a study of 156 Japanese patients treated with cisplatin during chemotherapy; the multifactorial logistic regression analysis showed the ERCC1 8092C>A gene AA as a significant acute-phase CINV predictor (Logistic regression, p = 0.011).28 The researchers concluded that the ERCC1 8092A allele causes the decreased expression and increased injury of this enzyme in normal gastrointestinal tissues. Thus, it promotes the small intestinal cell dysfunction induced by cisplatin but does not decrease its effect; hence, CINV continues to occur. However, Lamba et al found no correlation between the ERCC4 rs744154 and CINV.4
Dihydropyrimidine Dehydrogenase Gene
Dihydropyrimidine dehydrogenase (DPYD), encoded by the DPYD gene, is the first rate-limiting enzyme in the 5-fluorouracil catabolic cascade. It plays a key role in the fluorouracil catabolism, and a DPYD deficiency affects fluorouracil-related side effects.49 One study found an increased CINV risk in patients with the DPYD*2A rs3918290 splice donor variant (Logistic regression, n = 2594, p = 0.007).50 Madi et al found that the rare DPYD mutation, Asp949Val variant, was correlated with increased emetogenic toxicity in a large sample of 1456 patients with advanced colorectal cancer (Logistic regression, p = 0.002).51 However, there was no correlation between CINV and the DPYD variants of Cys29Arg, Val732Ile, Met166Val, Lys259Glu, Ser534Asn, Ile543Val, and IVS14+1G>A.
Methylenetetrahydrofolate Reductase Gene
Methylenetetrahydrofolate reductase (MTHFR) is a key enzyme in the folate metabolism; it plays an important role in DNA methylation and gene regulation.54 The MTHFR activity level affects the methylation of genomic DNA. There are several MTHFR gene mutation types; the most common is the C677T point mutation.53 Gao Changming et al investigated the relationship between MTHFR polymorphisms and 5-fluorouracil-based chemotherapy regimen efficacy and toxicity in 57 Chinese patients with advanced gastric cancer, and the CINV incidence was significantly higher in patients with the MTHFR C677T TT, TC, and A1298C AA genotypes than in patients with other genotypes (Chi-squared test, p = 0.04 and p = 0.00, respectively).54 In contrast, Kou Junyan et al’s study of patients with non-small-cell lung cancer treated with platinum in combination with gemcitabine found no correlation between the MTHFR C677T genotype and CINV (Chi-squared test, n = 105, p = 0.773).53
X-Ray Repair Cross-Complementary Gene 1
The X-ray repair cross-complementary gene 1 (XRCC1) is a widely studied DNA repair gene known to play an integral role in base excision repair. Yang Mudan et al investigated 159 patients with colorectal cancer in China and found that the CINV incidence was significantly higher in patients with the XRCC1 G28152A AA or AG genotype than in patients with the GG genotype (Chi-squared test, p < 0.01).55
Generic Transcription Factor IIE Subunit 1
The generic transcription factor IIE1, encoded by the generic transcription factor IIE subunit 1 (GTF2E1) gene, is essential for transcription initiation through direct interaction with DNA repair factors.56 Lamba et al conducted an SNP typing of the candidate genes associated with platinum and paclitaxel in the DNA samples of 90 adult male patients diagnosed with advanced non-small-cell lung cancer; the correlation with survival and toxicity was analyzed,4 and the results revealed a correlation between the transcription factor GTF2E1 rs447978 (specific allele not reported) and a reduced risk of CIN (p = 0.004). Platinum acts on inter-strand and intra-strand DNA cross-links, and the genetic variation in this gene may have implications for platinum’s pharmacokinetic and pharmacodynamic pathways or lead to differences in response and tolerability between patients.4
Dopamine D2 Receptor Gene
Mukoyama et al investigated the rs6277 and rs1076560 of the dopamine D2 receptor gene and the rs4680 of the catechol-O-methyltransferase gene; the results showed that only C homozygote patients with rs1076560 had an increased CINV incidence (Logistic regression, n = 60, p = 0.016).23 This study also investigated the same loci; however, no association was found.42
Acetaldehyde Dehydrogenase 2 (ALDH2)
It has been reported that habitual alcohol consumption is negatively correlated with CINV59 and that the ALDH2 gene might be the main factor affecting the alcohol metabolism. Toshitaka et al hypothesized that the alcohol metabolism enzyme gene may affect the metabolism of emetic chemotherapeutic drugs; they conducted a research study but found no correlation between ADLH2 rs671 and CINV.58
Fas Cell Surface Death Receptor Gene, Lysophosphatidic Acid Receptor 6, and CC Chemokine Ligand Gene 2
Oliva et al examined 48 SNPs in 43 genes and analyzed the correlation of each SNP with patient-reported nausea and vomiting frequency and intensity; the study included 114 females with breast cancer who received chemotherapy with 5-fluorouracil, epirubicin, and cyclophosphamide.59 The following three SNPs were found to be correlated with an increased risk of nausea: Fas cell surface death receptor gene (FAS) rs2234978 (Chi-squared test, p = 0.03), lysophosphatidic acid receptor 6 (LPAR6) rs2854344 (p = 0.03), and chemokine (C-C motif) ligand 2 (CCL2) rs2530797 (p = 0.01). FAS, LPAR6, and CCL2 genes are important for the control of cellular homeostasis. CCL2 is a chemokine gene involved in immune regulation and inflammatory processes,60 LPAR6 is a key component of the cell cycle control pathway,59 and FAS is a death receptor system gene that mediates apoptosis induction to maintain immune homeostasis. In addition, these genes are important for the patient’s immune response as well as the elimination of abnormal and cancerous cells.61 Researchers speculatively suggest that inflammation and cell death may also be correlated with CINV.
Gene polymorphisms other than the three most studied pathways of serotonin receptor genes, drug metabolism, and drug transport are equally clinically important for exploring and providing individualized CINV treatment and optimizing clinical antiemetic regimens. Thus, they provide new directions for future studies.
Discussion
The basic characteristics and results of the above-listed studies are summarized in Tables 1–3 for a brief analysis of the correlation between CIN/CIV and gene polymorphisms. As shown in Table 2, a total of 23 loci of 16 genes are associated with CIN occurrence or severity in 21 studies; as shown in Table 3, a total of 22 loci of 15 genes are associated with CIV occurrence or severity in 25 studies. It would seem that CINV partially depends on the differences between the genes involved in DNA repair, cell function, drug metabolism and transport, drug targets, serotonin receptors, NK-1 receptors, cell cycle processes, and cell death processes. Taken together, the reasons for the lack of consistency in the results of the above-mentioned studies include the sample characteristic heterogeneity, small sample sizes, gene frequency differences across ethnic populations, uncontrolled differences in phenotypic characteristics, and different CINV assessment tools.62 To date, there are few candidate gene studies based on different perspectives, and it is difficult to draw clear conclusions from such data due to the limitations mentioned above. Therefore, a larger scale, more diverse populations, and more rigorous validation studies (eg, genome-wide association studies) are needed to provide comprehensive evidence for the utility assessment of genetic polymorphisms as CINV predictors.
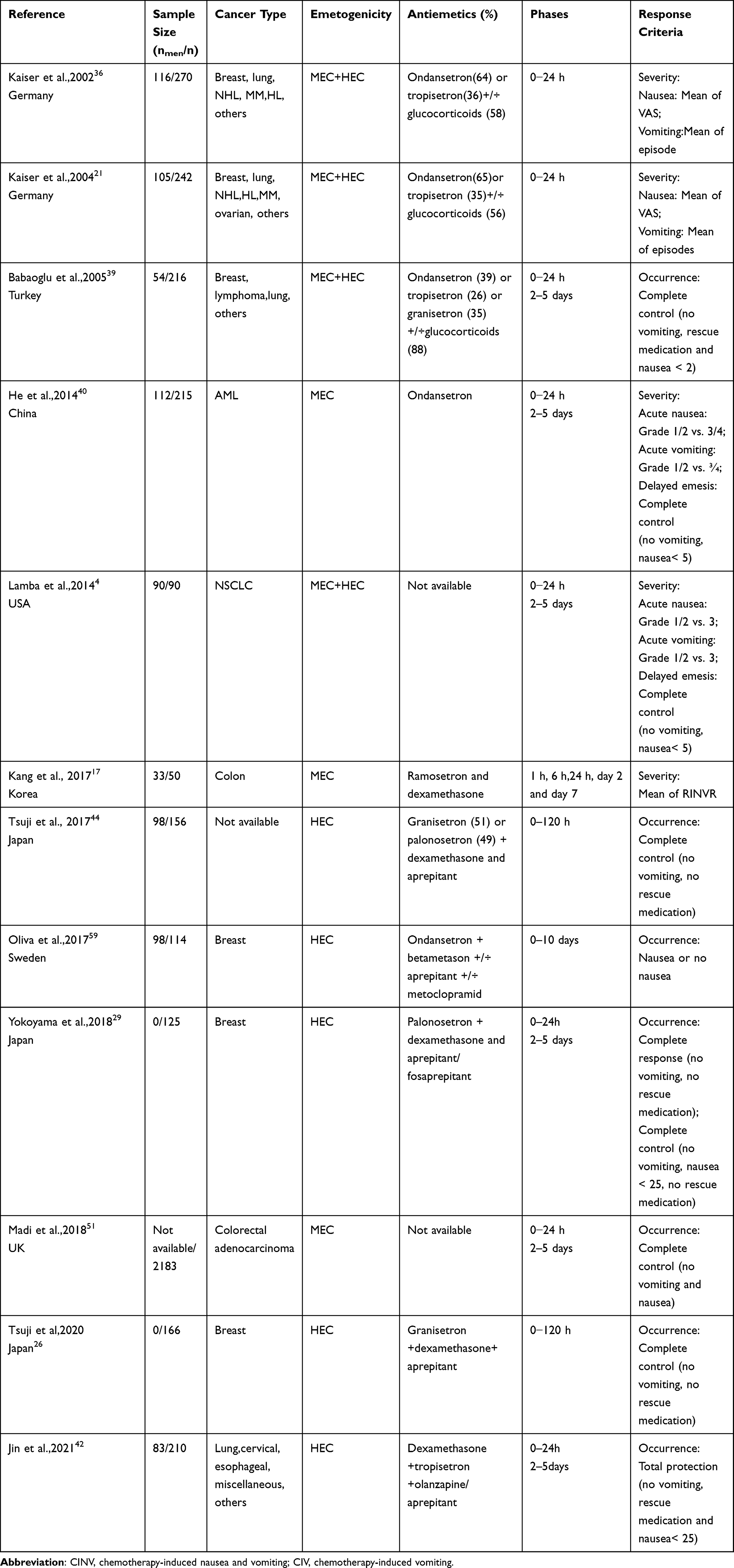 |
Table 1 Basic Characteristics of Clinical Trials of CINV-Related Gene Polymorphisms |
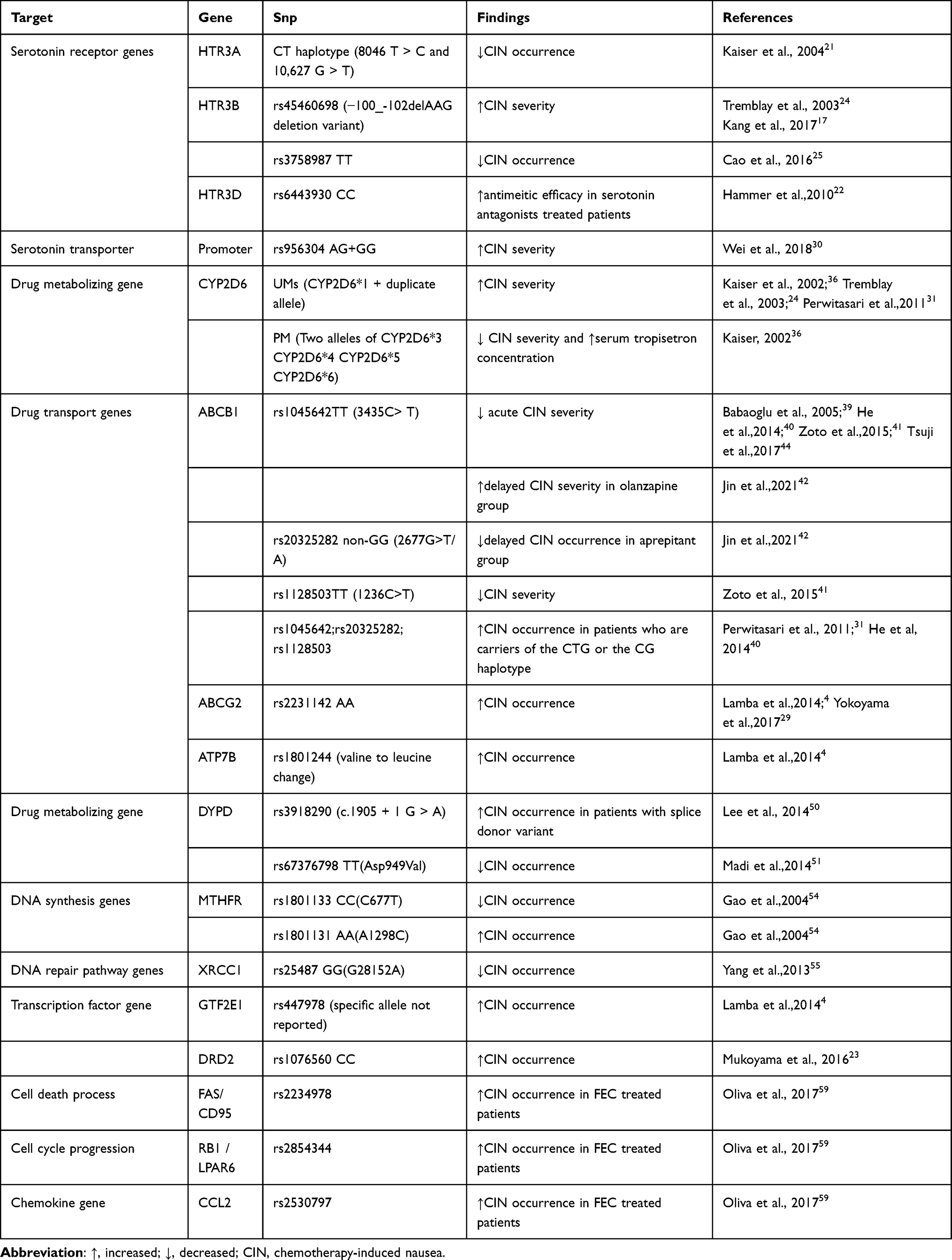 |
Table 2 Summary of Findings on Associations Between CIN and Candidate Gene Polymorphisms |
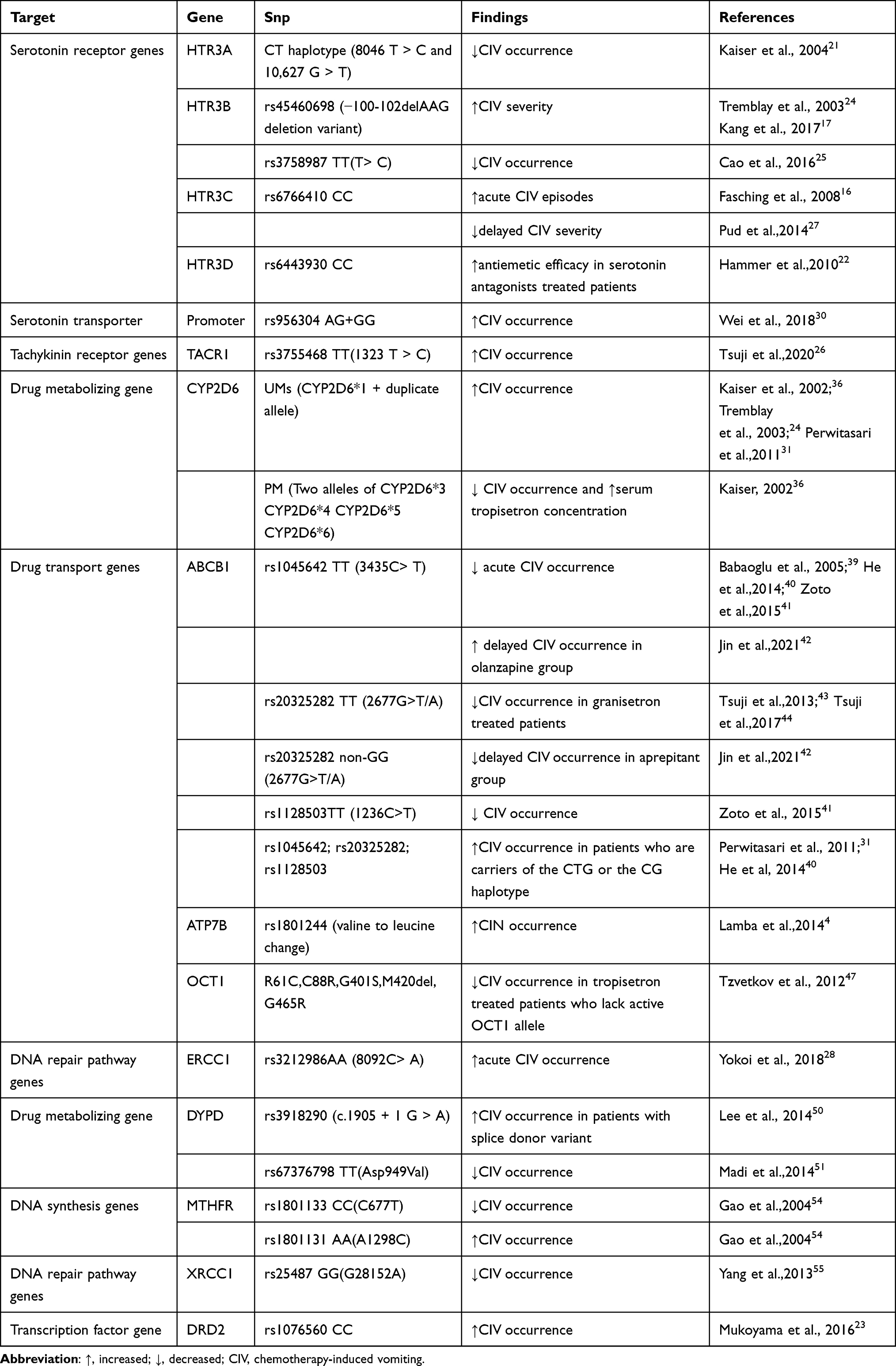 |
Table 3 Summary of Findings on Associations Between CIV and Candidate Gene Polymorphisms |
Genotype-based antiemetic prescriptions may become increasingly practical as the cost of genotyping decreases and Clinical Laboratory Improvement Amendments-certified genotyping becomes more readily available.15 The use of genetic polymorphisms to optimize the CINV effectiveness is beginning to receive attention, and individualized CINV prevention guided by a combination of patient clinical characteristics and genotyping is the direction of future development. It is expected that truly tailored antiemetic protocols will be available in clinical practice soon.
Acknowledgments
We would like to acknowledge the hard and dedicated work of all the staff that implemented the intervention and evaluation components of the study.
Lanzhen Zhao, MD, Lenggaowa Da, MD, Feng Chen, MM, Ying Jiang, MM, Gang Chen, MD, Yun An, MD, Shuqin Deng, MD, Wenjuan Wang, MM, Yungaowa Wu, MM,
Department of Medical Oncology, Ordos Central Hospital, Ordos 017000, China.
Funding
This study was supported by Science and technology project of Inner Mongolia Medical University.
Disclosure
The authors declare that they have no competing interests.
References
1. Sommariva S, Pongiglione B, Tarricone R. Impact of chemotherapy-induced nausea and vomiting on health-related quality of life and resource utilization: a systematic review. Crit Rev Oncol Hematol. 2016;99:13–36. doi:10.1016/j.critrevonc.2015.12.001
2. Koth SM, Kolesar J. New options and controversies in the management of chemotherapy-induced nausea and vomiting. Am J Health Syst Pharm. 2017;74(11):812–819. doi:10.2146/ajhp160227
3. Navari RM. Treatment of breakthrough and refractory chemotherapy-induced nausea and vomiting. Biomed Res Int. 2015;2015:1–6. doi:10.1155/2015/595894
4. Lamba JK, Fridley BL, Ghosh TM, et al. Genetic variation in platinating agent and taxane pathway genes as predictors of outcome and toxicity in advanced non-small-cell lung cancer. Pharmacogenomics. 2014;15(12):1565–1574. doi:10.2217/pgs.14.107
5. Navari RM, Aapro M. Antiemetic prophylaxis for chemotherapy-induced nausea and vomiting. N Engl J Med. 2016;374(14):1356–1367. doi:10.1056/NEJMra1515442
6. Aapro M. CINV: still troubling patients after all these years. Support Care Cancer. 2018;26(Suppl 1):5–9. doi:10.1007/s00520-018-4131-3
7. Jiang W, Ba Y, Feng J, et al. Chinese expert consensus on prevention and treatment of nausea and vomiting related to oncology drug therapy (2019 Edition). Chinese J Front Med. 2019;11(11):16–26.
8. Rapoport BL. Delayed chemotherapy-induced nausea and vomiting: pathogenesis, incidence, and current management. Front Pharmacol. 2017;8:19. doi:10.3389/fphar.2017.00019
9. Rojas C, Raje M, Tsukamoto T, et al. Molecular mechanisms of 5-HT3 and NK1 receptor antagonists in prevention of emesis. Eur J Pharmacol. 2014;722:26–37. doi:10.1016/j.ejphar.2013.08.049
10. Yu S, Yin J, Qin S, et al. Guidelines for prevention and treatment of vomiting related to tumor treatment (2014 Edition). Chinese Clin Oncol. 2014;19(03):263–273.
11. Natale JJ. Overview of the prevention and management of CINV. Am J Manag Care. 2018;24(18 Suppl):S391–S397.
12. Roila F, Molassiotis A, Herrstedt J, et al. MASCC and ESMO guideline update for the prevention of chemotherapy- and radiotherapy-induced nausea and vomiting and of nausea and vomiting in advanced cancer patients. Ann Oncol. 2016;27(suppl 5):v119–v133. doi:10.1093/annonc/mdw270
13. Ng TL, Hutton B, Clemons M. Chemotherapy-induced nausea and vomiting: time for more emphasis on nausea? Oncologist. 2015;20(6):576–583. doi:10.1634/theoncologist.2014-0438
14. Adel N. Overview of chemotherapy-induced nausea and vomiting and evidence-based therapies. Am J Manag Care. 2017;23(14 Suppl):S259–S265.
15. Puri S, Hyland KA, Weiss KC, et al. Prediction of chemotherapy-induced nausea and vomiting from patient-reported and genetic risk factors. Supportive Care Cancer. 2018;26(8):2911–2918. doi:10.1007/s00520-018-4120-6
16. Fasching PA, Kollmannsberger B, Strissel PL, et al. Polymorphisms in the novel serotonin receptor subunit gene HTR3C show different risks for acute chemotherapy-induced vomiting after anthracycline chemotherapy. J Cancer Res Clin Oncol. 2008;134(10):1079–1086. doi:10.1007/s00432-008-0387-1
17. Kang G, Kim K, Shim H, et al. Effect of the allelic variants ofABCB1, CYP2D6 andHTR3B on response of ramosetron to prevent chemotherapy-induced nausea and vomiting in Korean cancer patients. Asia Pac J Clin Oncol. 2017;13(1):53–60. doi:10.1111/ajco.12575
18. Celli J, Rappold G, Niesler B. The human serotonin type 3 receptor gene (HTR3A-E) allelic variant database. Hum Mutat. 2017;38(2):137–147. doi:10.1002/humu.23136
19. Ward MB, Kotasek D, Mckinnon RA. Investigation ofHTR3C mutations for association with 5HT3 receptor antagonist anti-emetic efficacy. Pharmacogenomics. 2008;9(8):1027–1033. doi:10.2217/14622416.9.8.1027
20. Kovac AL. Comparative pharmacology and guide to the use of the serotonin 5-HT(3) receptor antagonists for postoperative nausea and vomiting. Drugs. 2016;76(18):1719–1735. doi:10.1007/s40265-016-0663-3
21. Kaiser R, Tremblay PB, Sezer O, et al. Investigation of the association between 5-HT3A receptor gene polymorphisms and efficiency of antiemetic treatment with 5-HT3 receptor antagonists. Pharmacogenetics. 2004;14(5):271–278. doi:10.1097/00008571-200405000-00001
22. Hammer C, Fasching PA, Loehberg CR, et al. Polymorphism inHTR3D shows different risks for acute chemotherapy-induced vomiting after anthracycline chemotherapy. Pharmacogenomics. 2010;11(7):943–950. doi:10.2217/pgs.10.67
23. Mukoyama N, Yoshimi A, Goto A, et al. An Analysis of Behavioral and Genetic Risk Factors for Chemotherapy-Induced Nausea and Vomiting in Japanese Subjects. Biol Pharm Bull. 2016;39(11):1852–1858. doi:10.1248/bpb.b16-00440
24. Tremblay P, Kaiser R, Sezer O, et al. Variations in the 5-Hydroxytryptamine Type 3B Receptor Gene as Predictors of the Efficacy of Antiemetic Treatment in Cancer Patients. J Clin Oncol. 2003;21(11):2147–2155. doi:10.1200/JCO.2003.05.164
25. Cao J, Chen C, Zhang R, et al. Relationship between HTR3B gene polymorphism and nausea and vomiting after chemotherapy in 212 patients with head and neck cancer. Shandong Med. 2016;56(40):89–91.
26. Tsuji D, Matsumoto M, Kawasaki Y, et al. Analysis of pharmacogenomic factors for chemotherapy-induced nausea and vomiting in patients with breast cancer receiving doxorubicin and cyclophosphamide chemotherapy. Cancer Chemother Pharmacol. 2021;87(1):73–83. doi:10.1007/s00280-020-04177-y
27. Pud D, Har-Zahav G, Laitman Y, et al. Association between variants of 5-hydroxytryptamine receptor 3C (HTR3C) and chemotherapy-induced symptoms in women receiving adjuvant treatment for breast cancer. Breast Cancer Res Treat. 2014;144(1):123–131. doi:10.1007/s10549-014-2832-y
28. Yokoi M, Tsuji D, Suzuki K, et al. Genetic risk factors for chemotherapy-induced nausea and vomiting in patients with cancer receiving cisplatin-based chemotherapy. Support Care Cancer. 2018;26(5):1505–1513.
29. Yokoyama S, Tamaru S, Tamaki S, et al. Genetic risk factors associated with antiemetic efficacy of palonosetron, aprepitant, and dexamethasone in Japanese breast cancer patients treated with anthracycline-based chemotherapy. Clin Breast Cancer. 2018;18(2):e157–e165. doi:10.1016/j.clbc.2017.05.013
30. Wei Y, Zhou Y, Wang Q, et al. Serotonin transporter promoter single nucleotide polymorphism susceptibility to chemotherapy-induced nausea and vomiting in colorectal cancer. J Practical Med. 2018;34(06):949–952.
31. Perwitasari DA, Gelderblom H, Atthobari J, et al. Anti-emetic drugs in oncology: pharmacology and individualization by pharmacogenetics. Int J Clin Pharm. 2011;33(1):33–43. doi:10.1007/s11096-010-9454-1
32. Eliasen A, Dalhoff K, Mathiasen R, et al. Pharmacogenetics of antiemetics for chemotherapy-induced nausea and vomiting: a systematic review and meta-analysis. Crit Rev Oncol Hematol. 2020;149:102939. doi:10.1016/j.critrevonc.2020.102939
33. Yang Y, Botton MR, Scott ER, et al. Sequencing the CYP2D6 gene: from variant allele discovery to clinical pharmacogenetic testing. Pharmacogenomics. 2017;18(7):673–685. doi:10.2217/pgs-2017-0033
34. Trammel M, Roederer M, Patel J, et al. Does pharmacogenomics account for variability in control of acute chemotherapy-induced nausea and vomiting with 5-hydroxytryptamine type 3 receptor antagonists? Curr Oncol Rep. 2013;15(3):276–285. doi:10.1007/s11912-013-0312-x
35. Zhou SF. Polymorphism of human cytochrome P450 2D6 and its clinical significance: part I. Clin Pharmacokinet. 2009;48(11):689–723.
36. Kaiser R, Sezer O, Papies A, et al. Patient-Tailored Antiemetic Treatment With 5-Hydroxytryptamine Type 3 Receptor Antagonists According to Cytochrome P-450 2D6 Genotypes. J Clin Oncol. 2002;20(12):2805–2811. doi:10.1200/JCO.2002.09.064
37. Gilmore J, Bernareggi A. Complementary Pharmacokinetic Profiles of Netupitant and Palonosetron Support the Rationale for Their Oral Fixed Combination for the Prevention of Chemotherapy-Induced Nausea and Vomiting. J Clin Pharmacol. 2019;59(4):472–487. doi:10.1002/jcph.1338
38. Chen L, Takizawa M, Chen E, et al. Genetic polymorphisms in organic cation transporter 1 (OCT1) in Chinese and Japanese populations exhibit altered function. J Pharmacol Exp Ther. 2010;335(1):42–50. doi:10.1124/jpet.110.170159
39. Babaoglu M, Bayar B, Aynacioglu A, et al. Association of the ABCB1 3435C>T polymorphism with antiemetic efficacy of 5-hydroxytryptamine type 3 antagonists. Clin Pharmacol Ther. 2005;78(6):619–626. doi:10.1016/j.clpt.2005.08.015
40. He H, Yin JY, Xu YJ, et al. Association of ABCB1 polymorphisms with the efficacy of ondansetron in chemotherapy-induced nausea and vomiting. Clin Ther. 2014;36(8):1242–1252. doi:10.1016/j.clinthera.2014.06.016
41. Zoto T, Kilickap S, Yasar U, et al. Improved Anti-Emetic Efficacy of 5-HT3 Receptor Antagonists in Cancer Patients with Genetic Polymorphisms of ABCB1 (MDR1) Drug Transporter. Basic Clin Pharmacol Toxicol. 2015;116(4):354–360. doi:10.1111/bcpt.12334
42. Yilan J, Gaowa J, Jun Z, et al. Clinical Observation of Gene Polymorphism of Olanzapine or Aprepitant in Prevention of CINV. Pharmgenomics Pers Med. 2021;14:1–9.
43. Tsuji D, Kim Y, Nakamichi H, et al. Association of ABCB1 polymorphisms with the antiemetic efficacy of granisetron plus dexamethasone in breast cancer patients. Drug Metab Pharmacokinet. 2013;28(4):299–304. doi:10.2133/dmpk.DMPK-12-RG-084
44. Tsuji D, Yokoi M, Suzuki K, et al. Influence of ABCB1 and ABCG2 polymorphisms on the antiemetic efficacy in patients with cancer receiving cisplatin-based chemotherapy: a TRIPLE pharmacogenomics study. Pharmacogenomics J. 2017;17(5):435–440. doi:10.1038/tpj.2016.38
45. Morisaki K, Robey RW, Ozvegy-Laczka C, et al. Single nucleotide polymorphisms modify the transporter activity of ABCG2. Cancer Chemother Pharmacol. 2005;56(2):161–172. doi:10.1007/s00280-004-0931-x
46. Sprowl JA, Ness RA, Sparreboom A. Polymorphic transporters and platinum pharmacodynamics. Drug Metab Pharmacokinet. 2013;28(1):19–27. doi:10.2133/dmpk.DMPK-12-RV-073
47. Tzvetkov MV, Saadatmand AR, Bokelmann K, et al. Effects of OCT1 polymorphisms on the cellular uptake, plasma concentrations and efficacy of the 5-HT(3) antagonists tropisetron and ondansetron. Pharmacogenomics J. 2012;12(1):22–29. doi:10.1038/tpj.2010.75
48. Ning J, Jiao Y, Xie X, et al. miR1385p modulates the expression of excision repair crosscomplementing proteins ERCC1 and ERCC4, and regulates the sensitivity of gastric cancer cells to cisplatin. Oncol Rep. 2019;41(2):1131–1139.
49. Wigle TJ, Tsvetkova EV, Welch SA, et al. DPYD and Fluorouracil-Based Chemotherapy: mini Review and Case Report. Pharmaceutics. 2019;11(5). doi:10.3390/pharmaceutics11050199.
50. Lee AM, Shi Q, Pavey E, et al. DPYD Variants as Predictors of 5-fluorouracil Toxicity in Adjuvant Colon Cancer Treatment (NCCTG N0147). JNCI. 2014;106(12). doi:10.1093/jnci/dju298.
51. Madi A, Fisher D, Maughan TS, et al. Pharmacogenetic analyses of 2183 patients with advanced colorectal cancer; potential role for common dihydropyrimidine dehydrogenase variants in toxicity to chemotherapy. Eur J Cancer. 1990;2018(102):31–39.
52. Levin BL, Varga E. MTHFR: addressing Genetic Counseling Dilemmas Using Evidence-Based Literature. J Genet Couns. 2016;25(5):901–911. doi:10.1007/s10897-016-9956-7
53. Kou J, Hong W, Hong D, et al. Relationship between MTHFR C677T gene polymorphisms and chemotherapy side effects in advanced non-small cell lung cancer. China Oncol. 2014;24(03):197–202.
54. Gao C, Lu J, Toshiro T, et al. Polymorphism of methylenetetrahydrofolate reductase and sensitivity of stomach cancer to fluoropyrimidine- based chemotherapy. Chinese J Epidemiol. 2004;25(12):52–56.
55. Yang M, Liu X, Gao J. Effects of XRCC1 gene polymorphisms on efficacy and toxicity of FOLFOX4 chemotherapy for metastatic colorectal cancer. Chinese Remedies Clin. 2013;13(9):1127–1129.
56. Kuschal C, Botta E, Orioli D, et al. GTF2E2 mutations destabilize the general transcription factor complex TFIIE in individuals with DNA repair-proficient trichothiodystrophy. Am J Hum Genet. 2016;98(4):627–642. doi:10.1016/j.ajhg.2016.02.008
57. Mosa ASM, Hossain AM, Lavoie BJ, Yoo I. Patient-related risk factors for chemotherapy-induced nausea and vomiting: a systematic review. Front Pharmacol. 2020;1(11):329. doi:10.3389/fphar.2020.00329
58. Uomori T, Horimoto Y, Mogushi K, Matsuoka J, Saito M. Relationship between alcohol metabolism and chemotherapy-induced emetic events in breast cancer patients. Breast Cancer. 2017;24(5):702–707. doi:10.1007/s12282-017-0761-4
59. Oliva D, Nilsson M, Andersson B, et al. Single nucleotide polymorphisms might influence chemotherapy induced nausea in women with breast cancer. Clin Transl Radiation Oncol. 2017;2:1–6. doi:10.1016/j.ctro.2016.12.001
60. Kitamura T, Qian BZ, Soong D, et al. CCL2-induced chemokine cascade promotes breast cancer metastasis by enhancing retention of metastasis-associated macrophages. J Exp Med. 2015;212(7):1043–1059. doi:10.1084/jem.20141836
61. Peter ME, Hadji A, Murmann AE, et al. The role of CD95 and CD95 ligand in cancer. Cell Death Differ. 2015;22(5):885–886. doi:10.1038/cdd.2015.25
62. Singh KP, Dhruva AA, Flowers E, et al. A review of the literature on the relationships between genetic polymorphisms and chemotherapy-induced nausea and vomiting. Crit Rev Oncol Hematol. 2018;121:51–61. doi:10.1016/j.critrevonc.2017.11.012
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.