")
Back to Journals » International Journal of Nanomedicine » Volume 14
Advances in nanocarriers as drug delivery systems in Chagas disease
Authors Quijia Quezada C , Azevedo CS , Charneau S , Santana JM , Chorilli M , Carneiro MB , Bastos IMD
Received 22 February 2019
Accepted for publication 31 May 2019
Published 9 August 2019 Volume 2019:14 Pages 6407—6424
DOI https://doi.org/10.2147/IJN.S206109
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Thomas Webster
Christian Quijia Quezada,1,2 Clênia S Azevedo,1 Sébastien Charneau,3 Jaime M Santana,1 Marlus Chorilli,2 Marcella B Carneiro,4 Izabela Marques Dourado Bastos1
1Pathogen-Host Interface Laboratory, Department of Cell Biology, Institute of Biology, University of Brasilia, Brasília, Brazil; 2Department of Drugs and Medicines, São Paulo State University (UNESP), Araraquara, São Paulo, Brazil; 3Laboratory of Protein Chemistry and Biochemistry, Department of Cell Biology, Institute of Biology, University of Brasilia, Brasília, Brazil; 4Electron Microscopy Laboratory, Department of Cell Biology, Institute of Biology, University of Brasilia, Brasília, Brazil
Abstract: Chagas disease is one of the most important public health problems in Latin America due to its high mortality and morbidity levels. There is no effective treatment for this disease since drugs are usually toxic with low bioavailability. Serious efforts to achieve disease control and eventual eradication have been unsuccessful to date, emphasizing the need for rapid diagnosis, drug development, and a reliable vaccine. Novel systems for drug and vaccine administration based on nanocarriers represent a promising avenue for Chagas disease treatment. Nanoparticulate systems can reduce toxicity, and increase the efficacy and bioavailability of active compounds by prolonging release, and therefore improve the therapeutic index. Moreover, nanoparticles are able to interact with the host’s immune system, modulating the immune response to favour the elimination of pathogenic microorganisms. In addition, new advances in diagnostic assays, such as nanobiosensors, are beneficial in that they enable precise identification of the pathogen. In this review, we provide an overview of the strategies and nanocarrier-based delivery systems for antichagasic agents, such as liposomes, micelles, nanoemulsions, polymeric and non-polymeric nanoparticles. We address recent progress, with a particular focus on the advances of nanovaccines and nanodiagnostics, exploring new perspectives on Chagas disease treatment.
Keywords: delivery systems, nanobiosensors, nanodiagnostics, nanoparticle systems, nanovaccine
Introduction
Chagas disease, caused by the protozoan Trypanosoma cruzi, is endemic in Latin America, where between five and eight million people are infected. The disease is spreading to non-endemic countries, such as Australia, Canada, Japan, Spain and United States of America (USA).1 It is mainly transmitted by faecal contamination of Reduviidae insects through insect bites or another injury of the skin2 parasite can also be spread by blood transfusion, organ transplantation, congenital contamination, and consumption of contaminated food and drinks.3 The T. cruzi biological cycle is comprised of three fundamental forms: (1) infective trypomastigotes found in mammalian blood and in the hindgut of triatomine bugs as metacyclic forms, (2) epimastigotes, the proliferative form located in the bug’s midgut, and (3) amastigotes that multiply by binary fission inside mammalian host cells, causing disruption and the release of new trypomastigotes into the bloodstream capable of invading any nucleated cell to begin a new reproductive cycle.4 Regarding clinical symptoms, Chagas disease can manifest both an acute phase which is asymptomatic in most cases and a chronic phase that is depicted by digestive and/or cardiac lesions.5 Treatment is based on the nitroheterocyclic compounds benznidazole and nifurtimox. However, in long-term therapy, both of the aforementioned induce serious side effects and cross-resistance. To date, research into the production of Chagas disease vaccines has been conducted. The main difficulties have been finding a protective antigen and generating attenuated parasites that will not trigger pathology in the long-term.6 Therefore, in the absence of vaccines, control measures for Chagas disease remain limited to diagnosis and treatment.5
Nanocarriers has demonstrated important results in terms of increasing the efficacy and decreasing the toxicity of drugs, antigens, and adjuvants currently used against some diseases. The use of nanocarriers against pathogens provides a greater ability to overcome biological barriers, maintenance of drug integrity in biological media, with higher specificity to target cells and tissues together with prolonged drug release in comparison to conventional drugs.7 Moreover, nanodiagnostic systems have been developed to improve the accuracy of sample preparation and detection of infectious pathogens by means of a simple, quick, accurate and inexpensive technique.8
In this review, we provide an overview of synthetic methods, physical characteristics and delivery systems based on nanocarriers for various antitrypanosomal agents, through liposomes, micelles, mesoporous silica nanoparticles, polymeric and non-polymeric nanoparticles (Figure 1). In this context, we highlight new perspectives and innovative strategies on the treatment of Chagas disease, as well as a general overview of nanovaccines and nanodiagnostics.
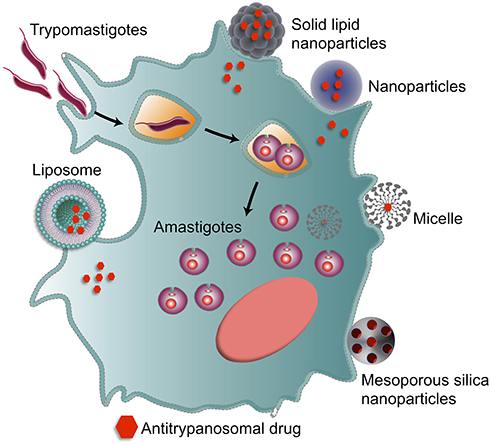 |
Figure 1 Nanomaterials used against Chagas disease. Strategies and application of nanocarrier-based drug delivery systems, such as liposomes, micelles, mesoporous silica nanoparticles, polymeric and non-polymeric nanoparticles to optimize the delivery of antitrypanosomal agents. |
General aspects
Physical characteristics of the nanocarriers
Some physical characteristics can influence nanocarrier functions and thus can be strategically explored to control the Chagas disease, especially against intracellular parasites, which are the most difficult forms to eliminate. These main characteristics are: 1) size of the particles: this feature improves greater bioavailability and circulation time (release, aggregation and transport of the drug), ranging from 1 to 100 nm or micrometric size (μm).9–11 2) Solubility: nanostructures can encapsulate insoluble or poorly soluble drugs, and the use of them can eliminate the need for toxic organic solvents.12 3) Shape: it can influence interactivity, cellular uptake, retention, biocompatibility, transport and drug loading capacity. The main shapes are spheres, discs, hemispheres, cylinders, cones, and tubes.10,11 4) Surface area: provides more reactivity as the particle size decreases in comparison to larger molecules (bulk solids), in addition, they are responsible for the improved water solubility and bioavailability.10 5) Net charge: it can influence the retention in the tissue, the circulation half-life and the uptake. In fact, cationic particles can have better uptake in the electrostatic interaction with the membrane,13,14 compared to particles with negative charges that have longer circulation half-life.15
The interactions of nanoparticles with their biological environment (molecules, cells, organs) depend on a complex interaction between controllable particle properties and largely uncontrollable properties of the surrounding media. Particle size, surface charge, polydispersity index, shape, and surface chemistry are key factors that determine the performance criteria, including the degree of protein adsorption, cell absorption, biodistribution patterns and mechanisms.16 Regarding Chagas disease, these nanomaterials must show a high volume of distribution and longer circulation time in the body compared to the available drugs (Nifurtimox or Benznidazole). It has been shown that highly cationic nanoparticles are eliminated from the circulation more rapidly than highly anionic nanoparticles.17 In contrast, neutral nanoparticles, as well as those with a slight negative charge, show significantly longer circulating half-lives, since the negative nature of cell membranes allows them to be highly absorbed by cells.18–22 Similarly, the size of the nanoparticle, with an average of less than 200 nm, enables a longer lifetime in the bloodstream, by avoiding interactions with the mononuclear phagocyte system and controlling drug delivery.23
Polymeric nanoparticles
Polymeric nanoparticles are solid colloidal particles used as adjuvants in vaccines or in therapeutic agent delivery systems that can be dissolved, entrapped, encapsulated or adsorbed onto the constituent polymer matrix. They are formed from natural or synthetic polymers such as: poly(lactide-co-glycolide) (PLGA); polylactide (PLA); polyglycolide (PGA); polycaprolactone (PCL); poly(D, L-lactide); chitosan, and PLGA-polyethylene glycol (PEG).24 Considering their biodegradability and biocompatibility, these polymers are among the few synthetic polymers approved by the U. S. Food and Drug Administration (FDA).25,26 Over the last 20 years, a variety of experimental administration systems with polymeric nanoparticles have been developed. An extensive compilation of detailed information about nanomaterials is presented in Table 1.
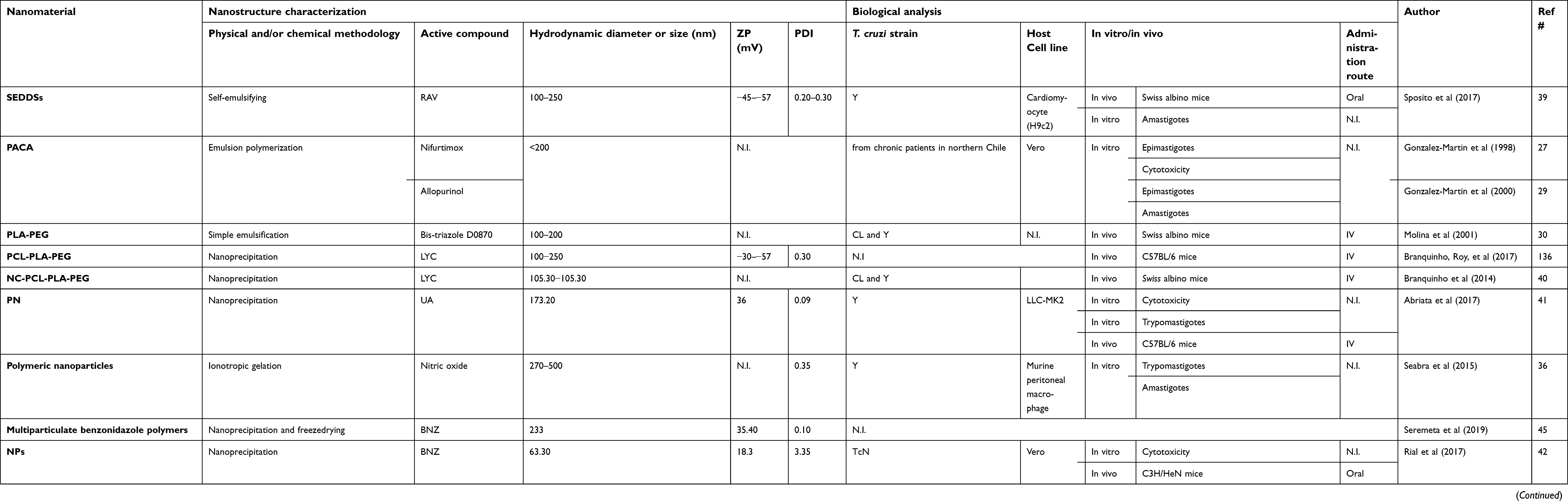 |
Table 1 Polymeric and liposomal nanomaterials against Chagas disease in preclinical trials |
The first studies with polymeric nanoparticles against Chagas disease were developed by Gonzalez-Martin et al27 and involved preparation of poly(alkyl cyanoacrylate) nanoparticles (PACA) containing nifurtimox. PACAs are generally formed from longer alkyl chains so degradation is slower and less cytotoxic.28 They obtained nanoparticles smaller than 200 nm, with 33.4% absorption to nifurtimox and a 65.4% drug release rate, after 6 hrs of incubation at pH 7.4. In vitro assays using nifurtimox-loaded nanoparticles and parasites isolated from chronic patients (from Northern Chile) showed the anti-epimastigote activity of 98.9% at a concentration of 0.2 μg/mL, while the same concentration of free nifurtimox in standard solution was approximately 40%. In addition, trypanocidal activity showed an IC50 of 0.683 (±0.269) μg/mL for the nifurtimox standard solution and 0.0015±0.0006 μg/mL for nifurtimox-loaded nanoparticles. Furthermore, electron microscopy studies revealed that intracellular parasites underwent a series of degenerative processes with an accumulation of electrodense nuclear chromatin, parasite cytoplasm autolysis, kinetoplast degeneration, and parasite membrane lysis. Finally, cell cultures previously infected with metacyclic parasites showed that after only 2 h of treatment with 0.001% nifurtimox-loaded nanoparticle suspension, parasitism was reduced by 87–94%. The degenerative processes observed by electron microscopy in the treated free amastigote and intracellular parasites were not observed in the host cells.
Gonzalez-Martin et al29 synthesized the same allopurinol-loaded polyether cyclic nanoparticles and showed that the pyrazolopyrimidine allopurinol has significant activity against T. cruzi, with less toxicity than nifurtimox. The maximum amount of allopurinol trapped in the nanoparticles was 62 (±1.9) μg/mg. Furthermore, their analysis of pH effects showed that after 6 h of incubation at pH 7.4, the release of allopurinol was 7.4%, whereas only 3.1% were released after 4–6 h at pH 1.2. Results of in vitro anti-T. cruzi activity experiments (epimastigotes) were considerably superior for allopurinol-loaded nanoparticles compared to an allopurinol standard solution (91.5% versus 45.9%) with 16.7 μg/mL of allopurinol. Cytotoxicity tests in Vero cells with allopurinol-loaded nanoparticles showed an IC50 80-fold lower than the value obtained for the free drug. The allopurinol-loaded nanoparticles significantly increased trypanocidal activity, thus these nanomaterials could prove to be an effective drug carrier against T. cruzi.
In 2001, Molina et al30 used sterols in polyethylene glycol-polylactide (PEG-PLA) nanospheres produced by the simple emulsification method with polymerization. Among nanoparticle characteristics, they showed a monodisperse size distribution of between 100 and 200 nm, and the drug encapsulation efficiency was 90, 87 and 92% for bis-triazole D0870, itraconazole and ketoconazole, respectively. In vivo assays were performed with Swiss albino mice infected intraperitoneally with CL or Y strains. Treatment over 30 consecutive days with D0870-loaded nanospheres, intravenous injection (IV), showed that the significant cure rate reached 60 and 90% for 1.5 and 3 mg/kg/day, respectively for both strains, whereas discharged nanoparticles demonstrated no trypanocidal activity. Finally, no induced healing or anti-T. cruzi activity was observed for ketoconazole or itraconazole, not even in the encapsulated form.
It has been well documented that endogenous nitric oxide (NO) plays a key role in the immune system against several intracellular pathogens.31 After infection, activated macrophages and other immune cells such as dendritic cells, crucial to connecting the innate and adaptive immune responses against T. cruzi,32 inhibit pathogen replication by releasing several effector molecules, including NO.33,34 As a free radical, NO is unstable and can be easily inactivated in the biological system. In order to increase the NO half-life, several authors described the preparation of low molecular weight S-nitrosothiols (RSNO), an important class of NO donors.35 However, RSNOs are thermally unstable in aqueous solutions and lead to the release of free NO.34 In this context, Seabra et al36 prepared and evaluated antiparasitic activity by incorporating biocompatible and biodegradable polymer nanoparticles to promote sustained NO release. An ionotropic gelation method was used to prepare chitosan/tripolyphosphate (TPP) containing S-nitroso-mercaptosuccinic acid (S-nitroso-MSA), a RSNO, which acts as a spontaneous NO donor. The antitrypanosomal activity showed great inhibitory effects against Y strain epimastigotes. A dose-dependent antitrypanosomal effect was observed in trypomastigotes with an IC50 value of 252 μg/mL and an EC50 of 91 μg/mL, suggesting the inhibitory effect is due to the presence of S-nitroso-MSA in the chitosan nanoparticles since the MSA nanoparticles and sodium nitrite were non-toxic. The S-NO groups act as a spontaneous NO donor due to the homolytic division of S-N with the release of free NO. The results obtained from murine peritoneal macrophage showed an antitrypanosomal activity of 200 μg/mL with NO-releasing nanoparticles decreasing the number of intracellular amastigotes. Furthermore, the treatment conditions tested during and after-infection indicated that NO release can interfere with adhesion, invasion and/or T. cruzi replication in mammalian cells. The authors claimed that this toxic effect against different T. cruzi lifecycle forms makes NO donor S-nitroso-MSA encapsulation an interesting approach for combating and preventing Chagas disease.
Another method of drug administration is the self-emulsifying drug delivery system (SEDDS) that contains a mixture of drug, oil, surfactant and occasionally solvent.37 The purpose of SEDDS is to improve drugs that have poor hydro-solubility and ultimately increase their oral absorption.38 Ravuconazole (RAV), an antifungal triazole drug with potent anti-T. cruzi activity (epimastigote and amastigote) was formulated by SEDDS with isotropic mixtures of anhydrous oil and surfactant.39 In vitro tests demonstrated more than 70% anti-amastigote activity, approximately 1.8 times more in comparison with free RAV with an equivalent IC50 dose (0.1 nM). In addition, in vivo toxicity tests using Swiss mice, treated with RAV-SEDDS (10%, v/v) for 20 days, showed no weight loss for animals treated with RAV-SEDDS or free RAV, indicating no additional drug toxicity. The authors concluded that RAV in the SEDDS dosage form is a strategy that merits further in vivo experiments and preclinical studies as a potential treatment of human T. cruzi infections.
Lychnopholide (LYC) is a bioactive compound found in plants from the Asteraceae family that has been shown to be a potent trypanocidal agent, but its use is limited by its low aqueous solubility and high lipophilicity. Therefore, Branquinho et al40 developed LYC-loaded nanocapsules (NC) using poly-ɛ-caprolactone (PCL) and poly (lactic acid) polyethylene glycol (PLA-PEG) polymers that were subsequently tested in Swiss mice infected intraperitoneally with T. cruzi. The mice infected with CL or Y strain were treated with 10 and 20 doses, respectively, of free drug LYC, Benznidazole (BNZ), LYC-PCL-NC or LYC-PLA-PEG-NC (2 mg/kg/day). The animals infected with the Y strain treated with LYC-PLA-PEG-NC in the prepatent period (with treatment initiated on day 7 after infection) for 20 days achieved 100% cure, while those treated with BNZ and LYC-PCL-NC achieved less than 75% and 62.5% cure, respectively. This study indicated that LYC-PLA-PEG-NC efficacy was attributed to the NCs ability to maintain LYC release for longer periods in biological media and when a higher number of trypomastigotes in the blood were exposed to the drug. Furthermore, PLA-PEG simultaneously performed a sustained and passive directed-delivery.
Abriata et al41 developed ursolic acid-loaded nanoparticles with poly-Ɛ-caprolactone (PN-UA) as a potential option for Chagas disease treatment, however, it was found to have low bioavailability. The PN-UA formulation had a size of 173.2 nm (±7.28), a polydispersity of 0.09 (±0.03) and a zeta potential of −36 mV (±3.34), which are important for particle stability, together with a maximum efficiency of encapsulation equivalent to 94.1% (±1.31). Cytotoxicity studies with fibroblast cells (LLC-MK2) showed no toxicity with PN-UA (120 μM) both in 24 hrs and 48 hrs of incubation whereas showed a 50.00±2.90% inhibition against trypomastigotes (Y- strain) at the same concentration. It was observed that PN-UA reduced parasitemia in male C57BL/6 mice, up to 3.5 times more in comparison with benznidazole. Thus, these nanomaterials may contribute to future clinical trials to treat Chagas disease.
Rial et al42 evaluated C3H/HeN mice infected with T. cruzi in the acute phase using the Nicaragua (TcN) strain by administering benznidazole nanoparticles (BNZ-nps) orally with 10, 25 or 50 mg/kg/day. Mice treated with these nanoparticles survived until euthanasia (92 days after infection, dpi). Histological analysis of cardiac tissue showed a significant decrease in inflammation, with lower levels of T. cruzi-specific antibodies. Additionally, Vero cells treated with BNZ-nps led to a significant increase in ROS production, suggesting these BNZ-nps could reduce the administration frequency while providing a significant increase in the pharmacological action related to the greater solubility together with a reduction in the number undesirable effects associated with BNZ.
A recent study developed by Tessarolo et al43 designed calcium carbonate nanoparticles loaded with benznidazole (BNZ@CaCO3). The @CaCO3 nanoparticles are biodegradable, biocompatible and have low toxicity in LLCMK2 cells.44 BNZ@CaCO3 was not only able to reduce epimastigote, trypomastigote and amastigote (Y-strain) viability showing that the amount of BNZ was up to 25 times lower when loaded with CaCO3 nanoparticles, they also had an improved selectivity index (SI =2.38).
Seremeta et al45 formulated nano- and multiparticulate benznidazole polymers, by means of nanoprecipitation/lyophilization based on Acrylates/Ammonium Methacrylate Copolymer, aiming to increase the dissolution rate of the drug. The obtained nanoparticles had a size range of 200–300 nm, yield of 86%, encapsulation efficiency of 78%, while the load capacity was 24% p/p and a zeta potential of 35.4±1.7 that was able to maintain during the lyophilization process. In addition, in vitro drug release assay showed that the nanoparticles had a higher rate compared to free drugs. Therefore, these nanocarriers would help in the pediatric treatments for Chagas disease, and the powder formulation would allow the dose flexibility to adapt to the dosage requirements depending on age groups.
Liposomes
Liposomes are vesicular nanomaterials formed by a lipid bilayer composed of phospholipids and cholesterol molecules, characterized by extended, two-dimensional and clearly separated hydrophilic and hydrophobic regions.46 These systems can deliver molecules such as ribosomes, proteins and/or peptides, DNA, immunological adjuvants or drugs while providing greater therapeutic efficacy with reduced toxicity and biocompatibility in biological systems.47–49 Over the last three decades, a variety of experimental lipid administration systems have been developed (Table 1).
Morilla et al50 showed that multilamellar liposome vesicles (MLV) loaded with benznidazole (BNZ) presented three times higher accumulation in the mice liver, in addition to 30% lower blood BNZ concentrations (1.1 g/mL), compared to free BNZ treatment. However, increased hepatic uptake of BNZ under the conditions tested had no effect on parasitemia levels in T. cruzi pantropic/reticulotropic RA strain-infected mice. This strain is generally more studied in mice, because they show high virulence and develop an early peak of parasitaemia during acute infection. In addition, they can induce a transient deterioration of the microbicidal activity of macrophages and the lymphoproliferative response induced by mitogens in vitro.51
Hydrogenated trypanocidal etanidazole (ETZ) is a drug with trypanocidal activity, but its hydrophilic structure causes slow diffusion through the membrane. In this context, in 2005, Morilla et al52 used pH-sensitive liposomes to encapsulate ETZ (L-ETZ). Murine macrophage-like cell line J774 treated with L-ETZ showed an anti-amastigote activity of 72% (RA strain), while the same dose of free ETZ produced no activity. In vivo experiments in female BALB/c mice, showed a significant decrease in parasitemia levels. In contrast, inoculation of a 180-fold higher dose of free ETZ failed to reduce the number of trypomastigotes in the bloodstream and failed to achieve complete parasitological cure with L-ETZ treatment. The limitations of L-ETZ can be attributed to the fact that this nanomaterial is not directed to several cells such as hepatic macrophages, spleen cells, nerve cells, muscle, considering that these liposomes are pH-sensitive. In conclusion, this study should be conducted with charge modifications made to the surface of this nanomaterial in order for it to assist immune system evasion and have a greater penetration capacity in most cells.
Amphotericin B (AmB) is a macrolide polyene chemotherapy that exists in three different aggregation states: monomer, dimer, and poly-aggregate.53 Formulations of parenteral AmB, either as the original micelle formulation with sodium deoxycholate (Fungizone®) or the less nephrotoxic and hemolytic liposomal formulation (AmBisome®), have been used as effective treatments for visceral leishmaniasis (VL).54 However, although AmB trypanocidal activity in infections was reported for the first time in 1962 by Actor et al,55 and there are several tests illustrating in vitro nanomolar trypanocidal activity for amphotericin B-deoxycholate and lipid formulations of AmB (amphotericin B colloidal dispersion (Amphocil®) and hemolytic liposomal formulation),56 only relatively few reports described the in vivo effects of AmB liposomes in mice infected with T. cruzi,57 with no AmB formulation authorized in the market. However, when used against T. cruzi, high parenteral doses (>25 mg/kg) of AmB were required for a prolonged period. AmB has both low solubility and permeability in the gastrointestinal epithelium, resulting in a low oral bioavailability (<0.9%).58 In this context, studies carried out by Cencig et al59 using amphotericin B liposomes showed efficacy in BALB/cJ mice infected with Tulahuen strain trypomastigotes during acute and/or chronic stages. Amphotericin B liposomes administration by 6 intraperitoneal injections at a dose of 25 mg/kg, at different times, showed that it prevented the mice from having fatal problems in the acute phase. In addition, parasitemia was reduced in acute and undetectable phase in chronic infection. Quantitative PCR analyses showed a significant reduction in parasite load in the heart, liver, spleen, skeletal muscle and adipose tissues in the acute and chronic phases. This study indicated that amphotericin B liposomes does not completely cure T. cruzi infected mice, it merely prevents mortality and significantly reduces parasite load in most tissues.
Ideally, nanoparticulate systems must be efficient in both acute stage and in the more challenging chronic stage of the disease that is characterized by the presence of intracellular amastigotes in different parts of the organism such as cardiac, skeletal, smooth muscle and glial cells.60 Therefore, nanoparticulate systems must have broad and selective targeting to infected cells (intracellular sites of amastigotes), reducing parasite viability. The use of pH-sensitive particles, which allow the drug release in the cytoplasm and nests of amastigotes, has been explored, however, reaching cellular internalization and endosomal escape is a considerable barrier. The endosomal compartmentation of internalized nanoparticles, subjected to a low pH environment and enzymes, is unfavourable to drug loading, especially for encapsulated drugs.61 Consequently, these nanomaterials must function at different pH levels to obtain a more drastic reduction in parasitemia levels, but so far, few studies relating to these particles have been published (Figure 2).
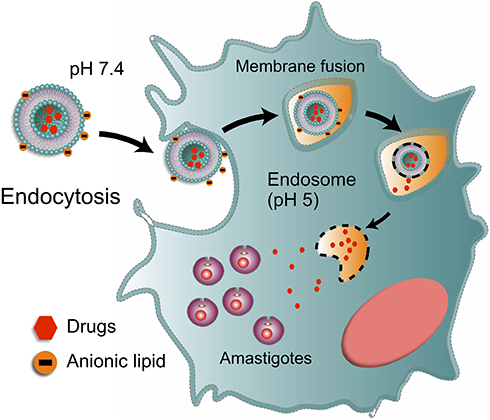 |
Figure 2 Hypothetical mechanisms of the internalization of pH-sensitive liposomes. |
Other types of nanomaterials
A variety of experimental administration systems have been developed with other types of nanomaterials such as solid lipid nanoparticles, metal nanoparticles, micelle and nanoemulsions (Table 2).
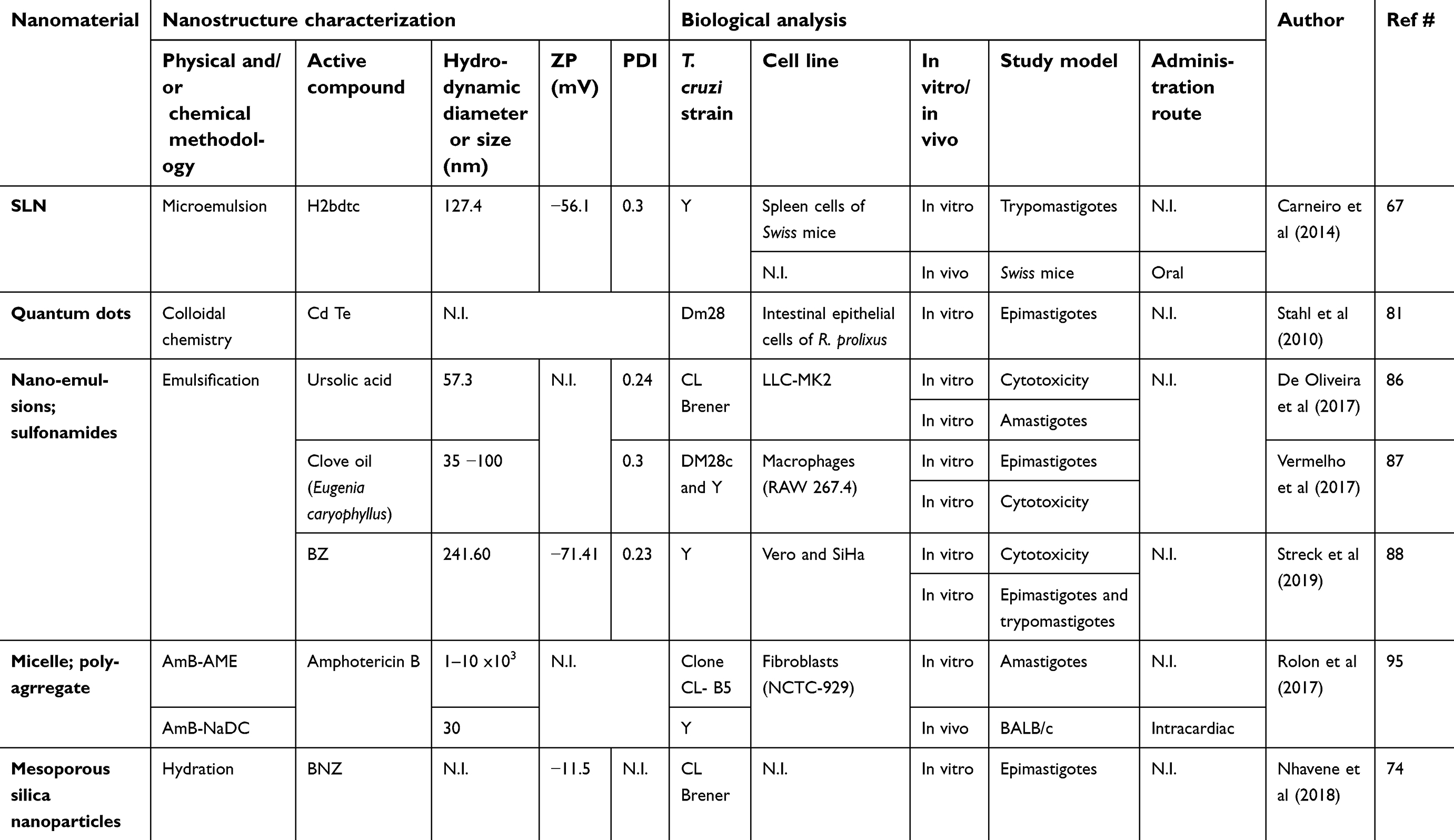 |
Table 2 Other types of nanomaterials against Chagas disease in preclinical trials |
Solid lipid nanoparticles
Solid lipid nanoparticles (SLN) are nanoparticles composed of a solid and a liquid lipid mixture in which the lipid matrix is solid at room and body temperature. They are stable and have the ability to encapsulate drugs with controlled-release. In addition, SLN has good biocompatibility, biodegradability, high bioavailability, and also the possibility of large-scale production.62–65
5-Hydroxy-3-methyl-5-phenyl-pyrazoline-1- (S-benzyl dithiocarbazate) (H2bdtc) belongs to the class of S-dithiocarbazates. Previous studies have shown the effectiveness of H2bdtc against Chagas disease, but their interference in subsequent studies are due to its low solubility.66 In this context, Carneiro et al67 synthesized H2bdtc-loaded with SLN. The authors showed that H2bdtc-SLN had an effective parasitemia reduction at concentrations 100 times lower than those normally used for benznidazole (clinically applied at 400 µM/kg/day). In addition, it decreased inflammation and damage to the liver and heart resulted in 100% survival of mice infected with the T. cruzi Y strain. Finally, H2bdtc-SLN overcame problems inherent to the low water solubility of the compound and it could be more accessible to the parasite.
Immunostimulant complexes (ISCOM),68 are lipids composed of Quillaja saponaria in a pentagonal dodecahedral shape of uniform size (40 nm) used as adjuvants for vaccines,69 such as DNA vaccines, antigen carriers for systemic, oral and intranasal administration.70,71 The basis of ISCOM structure is the interaction between saponin molecules, cholesterol, phospholipids and hydrophilic and hydrophobic antigens associated in a single nanomaterial. When combined, they form stable rings in aqueous solutions. These particles trigger a balanced Th1 and Th2 response and stimulate a cytotoxic cellular response that is rarely obtained with other adjuvants.72 Encapsulation of actinomycin D in ISCOM functionalized with antibodies produced dose-dependent effects,73 and 100% effectiveness at 25.47×10–2 ng/mL or 196.8 times less than the antibiotic alone, which kills the parasite within 24 h at 50 ng/mL. ISCOM functionalized with compounds containing vinyl sulfone and a hydrophobic chain are capable of binding proteins or ligands and are suitable for delivery systems in directional therapy in which antibodies or specific ligands are required to drive them to their target. Thus, this system increases drug effectiveness while drastically reducing dosage and toxicity, which is often disadvantage in conventional therapy.
Mesoporous silica nanoparticles
A recent study developed by Nhavene et al74 combined mesoporous silica nanoparticles, solid inorganic material and chitosan coating, and effectively showed the anchoring of benzonidazol molecules. These silica nanoparticles have a network of ordered pores and uniform sizes that help to functionalize the antichagasic agent and other components. In transmission electron microscopy, these nanoparticles showed a uniform shape and sizes over 3.3 nm, and zeta potential of −11.5±0.5 mV. In vitro assays showed that these particles were thirty times more active against CL-Brener epimastigotes than free BZ, revealing their potential as nanoplatforms for drug delivery.
Metallic nanoparticles
Metallic nanoparticles (MNPs) are clusters of metal atoms with optoelectronic and physicochemical properties that are strongly dependent on their size, shape, crystallinity, and structure. MNPs are used for a wide range of applications such as molecular diagnostics, electronics, catalysis, together with the delivery of drugs, antibodies, and other ligands. However, potential toxicity and other properties that might influence toxic responses must also be studied and understood. A study conducted by Eger and Soares (2012),75 monitored intracellular endocytosis in epimastigotes (strain Dm28c) with gold nanoparticles marked with transferrin (iron-specific transport protein in the plasma of humans and mammals) by confocal microscopy (633nm) and evidenced two sites of parasite that represent the lower part of the cytopharynx and reservosomes, being able to visualize the basic aspects of endocytosis. This study opens up promising routes for immunolocalization research with secondary antibodies labeled with Au.
Quantum dots
Quantum dots (QD) have unique autofluorescence properties and thus have great potential in biomedicine for diagnosis, drug delivery and imaging.76 These nanoparticles can have adverse effects in vitro and in vivo and pose potential risks to human health. QDs can be distributed to all body systems and they can be added in some tissues and organs.77,78 QDs deposited in cells can alter cell viability and morphology, including induction of chromatin condensation and apoptosis.79,80
In 2010, Stahl et al81 evaluated the possible effects of quantum dots of cadmium telluride (CdTe) in T. cruzi epimastigotes (clone Dm28). Using high doses of QD (20 and 200 μM) led to a decrease in T. cruzi growth patterns. Additionally, the concentrations used to label T. cruzi and intestinal epithelial cells from Rhodnius prolixus were 2 and 10 μM of CdTe and CdSe, respectively, encouraging further study into the interaction between parasites and vertebrate host cells. They showed that after 72 h with 200 μM CdTeQD, important morphological alterations were induced in T. cruzi, such as DNA damage, the formation of blisters in the plasma membrane and mitochondrial swelling. However, parasites incubated with CdTe QDs at 2 μM continued to proliferate after seven days. In conclusion, a low concentration of CdTe QDs (2 μM) is optimal for bioimaging, while a higher concentration (200 μM CdTe) can be toxic to cells due to the resulting morphological changes.
Nanoemulsions
Nanoemulsions (NEs) are emulsions of nanometric particles ranging between 10 and 600 nm,82 consisting of a mixture of insoluble liquids in two phases in which vesicles in the dispersed phase are surrounded by a continuous phase. This nanomaterial is ideal for improving the solubility of hydrophobic drugs. Different types of surfactants are used to stabilize the emulsion in order to prevent the dispersed phase from fusing in a macroscopic phase.83 The advantage of these nanoemulsions is their mutual compatibility - solubility in aqueous media and the ability to protect drugs from hydrolysis and enzymatic degradation.84,85
Oliveira et al86 used ursolic acid, a molecule with trypanocidal action, in nanoemulsions which were stable for 90 days, 57.3 nm in size and had a polydispersity index of 0.24. The dissolution profile in alkaline medium showed a release of 75% ursolic acid after 5 mins. Biological studies showed that the developed formulation was non-toxic in LLC-MK2 fibroblast cells (monkey kidney) and possessed anti-amastigote activity in the CL Brener strain, with a Selective Index (SI) of 21.66 μM and an IC50 of 18 μM.
Vermelho et al87 synthesized NEs with sulfonamides and clove oil (Eugenia caryophyllus). Previous studies have shown that sulfonamide carbonic anhydrase inhibitors targeting the α-class enzyme from T. cruzi have high trypanocidal activity at 4 µM. Growth inhibitions of 57% and 43.51% were observed in DM28c and Y strain epimastigotes, respectively. In addition, cellular toxicity against RAW 267.4 macrophages had an SI value ranging from 1 to 3, compared to the reference drug benznidazole (SI =5–5.8). These effects are probably due to greater enzyme inhibitor penetration through nanoemulsion formulation that had average sizes of 35–100 nm and a PDI of less than 0.3, indicating that the size distribution is homogeneous and monomodal.
A nanosystem was recently developed to release benznidazole from oil nanoemulsions and showed efficacy against Trypanosoma cruzi parasites (Y strain).88 This nanostructure was established with a transition from laminar liquid crystal to nanoemulsions using cosolvents. They showed a diameter of 241.60±17.10 nm, PdI of 0.23 and a zeta potential of −71.40±4.9 mV, and these characteristics favored the improved aqueous solubility of the drug and slow its release rate. In addition, the in vitro cellular biocompatibility of the formulations was evaluated in Vero and SiHa cell lines, showing slight cytotoxicity up to 200 μg/mL for both cells, whereas IC50 on epimastigotes was ~0.30 μg/mL and 0.53 for trypomastigotes. The obtained results indicating these nanoemulsions are potential colloidal drug delivery system candidates.
Micelles
Micelles are composed of amphiphilic polymers that self-assemble into nanomaterial ranging from 20–200 nm in size,89 which are generally in the form of spherical vesicles. They contain a hydrophobic core into which hydrophobic substances, such as pharmacological compounds, can be introduced for release in different parts of the body. These nanomaterials provide greater solubility and stability of hydrophobic drugs,90 especially in vivo when compared with the free drug. Furthermore, they are able to i) improve cellular uptake of drug-loaded micelles, and ii) provide an alternative form of internalization (endosomes).91–94
Another strategy to release poly-aggregate amphotericin B (AmB) was developed by Rolon et al95 which involved two types of formulation: 1) AmB polyamide encapsulated in albumin microspheres (AmB-AME), and 2) dimeric AmB-sodium deoxycholate micelles (AmB-NaDC). The dimeric AmB-NaDC exhibited a promising selectivity index (SI =3164) for T. cruzi amastigotes, which was much higher than those obtained for the licensed drugs - benznidazole and nifurtimox. In vivo experiments administered AmB-NaDC orally at 10–15 mg/kg/day for 10 days in BALB/c mice infected with the T. cruzi Y strain, with a 75% reduction of parasitemia and prolonged survival observed in the acute phase. Similarly, AmB-AME at 5 mg/kg increased survival and reduced parasitemia by 3.6 times after 7 days of infection in the acute phase, compared with the AmB-NaDC form. However, toxicity associated with parenteral administration was observed with AmB-AME that can be attributed to cardiomyopathy,57 which renders them more susceptible to the side effects associated with AmB infusion. This study suggested further AmB-NaDC pharmacokinetic investigations for the clinical development of a cost-effective and orally bioavailable AmB treatment for Chagas disease.
Nanocarriers for photodynamic
Currently, many studies are being carried out with photodynamic therapy in several areas of medicine such as cancer, skin diseases, macular degeneration, psoriasis, atherosclerosis, myopia and others. In addition, they can be applied against microorganisms, such as fungi, bacteria, viruses and protozoa.96–98 Photodynamic inactivation was tested in vitro on trypomastigotes (Y strain) using the water-soluble photosensitizer hypericin, formulated with ethanol/water, loaded in different systems of aqueous nanocarriers, such as polymeric micelles Poloxamers (Pluronic ™ surfactants) and liposomal phospholipid vesicles 2-dipalmitoyl-sn-glycero-3-phosphocholine.99 These systems had activity to eradicate the trypomastigote forms in the dark, however with a stronger activity under light. For instance, the pluronic micelles showed EC50 around 6–8 μmol/L tested in the dark and 0.31–0.36 μmol/L under light whereas using the lipid vesicles the values were 10.84±0.47 μmol/L in the dark and ~5 fold less (2.22±0.52 μmol/L) under light. Although the results are promising, further investigation is needed to assess whether these systems have photo-efficacy in vivo.
Nanodiagnostics
Some nanodiagnostic tools are based on nanomaterials such as liposomes, nanopores, nanowires and metallic nanoparticles, which are combined with conventional methods such as fluorescence, potentiometry, and voltammetry. It incorporates biological recognition elements or biomimetics, including nucleic acids, enzymes, antibodies, receptors, tissue, cells, inflammatory proteins from macrophages, aptamers and peptides.100,101 The interaction between the compound or microorganism of interest and the recognition element produces a variation in one or more physicochemical properties (such as: pH, electron transfer, heat, potential, mass, optical properties, etc.) that are detected by a transducer. The resulting electronic signal indicates the presence of an analyte of interest and its concentration in the sample. Therefore, nanobiosensors are revolutionizing the in vitro diagnosis of diseases and have important implications for human health.
Biosensors based on electrically active field effect transistors (FETs) offer direct, rapid and highly sensitive detection capabilities without labeling several biomolecules of specific interest. Nanowires (NW) are more efficient bioelectrochemical transducers, compared to their thin film analogs, so they are used as efficient detection biosensors.102
An integrated microfluidic system coupled to screen-printed carbon electrode (SPCE) has been developed to quantify specific IgG antibodies present in serum samples from Chagas disease patients.103 The principle is based on an SPCE coupled to a microfluidic device modified by the electrodeposition of gold nanoparticles (AuNP) functionalized with T. cruzi proteins from epimastigote membranes. A microfluidic immunosensor was used to detect specific anti-T. cruzi IgG antibodies that reacted immunologically with the immobilized T. cruzi antigen. The calculated detection limit was 3.065 ng/mL and the intra-assay and inter-assay variation coefficients were lower than 6.95%. These values demonstrated this immunosensor could be used as a serological method for the diagnosis of congenital Chagas disease in addition to donor blood screening and monitoring antibody reduction during the treatment of chagasic patients.103
Early and non-invasive diagnosis for congenital Chagas disease concentrates, preserves and detects T. cruzi antigens from the patient’s urine (Figure 3).104 The Chagas urine nanoparticle assay (Chunap) was effective for a single urine sample from a one-month-old baby with more than 90% sensitivity and more than 95% specificity. Nanoparticles were functionalized with polyunsaturated N-isopropylacrylamide and trypan blue, which can capture and concentrate T. cruzi analytes from different chemical structures, including proteins (H49 and 1F8), glycoproteins (TESA) and lipophosphoglycans.
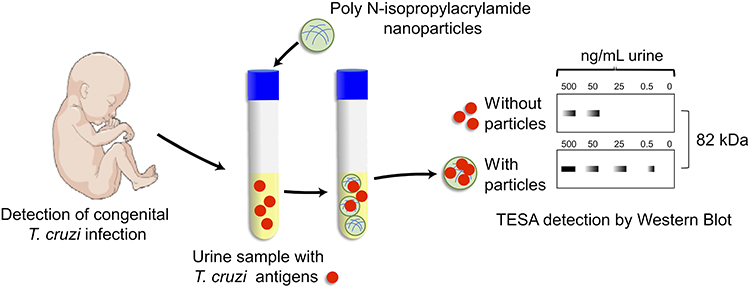 |
Figure 3 Chunap assay for the early diagnosis of congenital Chagas disease. Chunap (Chagas urine nanoparticle assay) is to perform an early and non-invasive diagnosis for congenital Chagas disease, by concentrating, preserving and detecting antigens secreted/excreted by trypomastigote forms (TESA) from urine by Western Blot. |
Surface functionalization and covalent immobilization of biomarkers, using biocompatible ethanolamine and polyethylene glycol derivatives, substantially increases the sensitivity and molecular selectivity of biosensors based on field effect transistors (nano-FET).105 The authors achieved significantly improved binding specificity, biomarker density and capture efficiency of target biomolecules for DNA as well as pathogen proteins. The developed devices provided ultra-high sensitivity capable of detecting approximately ~1 fM for specific DNA sequences and 6 fM for a protein marker of Chagas disease (IBMP8-1), measured by the net change in electrical resistance of the device. The developed InP nanowire biosensor provides a qualified tool for the detection of the chronic stage, leading to better diagnosis and spreading control.
Currently, there are few conventional tests for Chagas disease diagnosis, causing a limitation in parasite detection in acute and chronic phases. Providing detection of the different stages of infection using nanobiosensors would lead to better diagnosis and spreading control. Another advantage within the limit of detection based on nanomaterials is the use of a wide range of biological fluids such as blood, saliva or urine.106 Therefore, they could improve the monitoring of patients’ treatment in terms of speed, small sample volume, ease of use, real-time detection and low cost compared to current conventional tests.107 That said, more research efforts are still needed to develop these types of nanodiagnostics.
Nanovaccine
Studies have shown the importance of CD4+ and CD8+ T cell responses against T. cruzi infection.108–110 Attention is directed toward antigens that elicit cell-mediated immune responses and would result in immune recognition of host cells infected with parasites.111,112 A favourable immune response would have the ability to trigger the production of the cytokine associated with IFN-γ induced Th1 response by CD8+ T cells. Depletion of CD8+ T cells in mice results in uncontrolled parasitemia and high mortality.113,114 A therapeutic vaccine for Chagas disease has potential advantages, such as reduction in side effects and the potential to be used as a replacement for current therapies or in parallel with low doses of chemotherapy.115
Adjuvants
Adjuvants are used to increase the immune response against vaccine antigens, achieve homogeneous responses in vaccinated individuals, decrease the amount of antigen used, reduce the number of vaccine doses and direct a beneficial response profile.116 This is achieved using molecules that stimulate pattern recognition receptors (PRRs), using strategies that improve the uptake and presentation of antigens by dendritic cells, promoting the antigen transport by lymph node drainage or by deposit formations.117
A study conducted by Bertona et al118 evaluated ISPA immunostimulatory nanoparticles and developed an adjuvant based on cage-like particle composed of Q. saponaria, phospholipids (PL), cholesterol (CHOL) and hydrophobic antigens associated in a single nanostructure. The size of the nanomaterial was 42.1 nm and the zeta potential was −1.96 mV. The authors compared it to different adjuvants such as Freund Adjuvant (FA), aluminum hydroxide (AH), Montanide TM ISA 206 (I206) and the commercial form ISCOMATRIX™ (IMX) aiming to determine the level, kinetics and humoral response profile against the parasite (Tulahuen strain) used female BALB/c mice. Unlike the individual effect of these adjuvants, combined ISPA particles improved the immune response against specific antigens, reduced the number of doses, provided a homogenized response among individuals and a balanced Th1/Th2 response. Moreover, mice immunized with ISPA and trans-sialidase (TS) generated a complete protective immune response when challenged with T. cruzi parasites.
Proteoliposomes
The reconstitution of membrane proteins in liposomes has proved to be a useful tool for preparing antigenic components to induce immunity in experimental models.119,120 Regarding cellular immune responses, Mesa et al121 showed that very small proteoliposomes can act as effective adjuvants for Th1 polarization and dendritic cell activation.
In this context, proteoliposomes for T. cruzi transport proteins were prepared from sonicated trypomastigotes and amastigotes (Y strain),122 and mixed with sodium dodecyl sulfate (SDS) to achieve maximum solubilization of parasite membranes, without destroying protein antigenicity. The resulting proteoliposomes, with a mean diameter of 199 nm, were used to immunize BALB/c mice, which were further infected with trypomastigotes. Intraperitoneal macrophages collected one week after the in vivo infection, were infected in vitro and showed a significant decrease of intracellular parasites 72 h later, in comparison with the controls. Despite the generated antibodies, macrophage activation and delayed animal death, no difference was noted with regards to survival rate between the groups on day 30 after infection. In conclusion, although these proteoliposomes do not protect mice from challenge with a virulent strain of T. cruzi, the improvement of these membrane mimic systems as immunogenic vehicles for parasite molecules could eventually contribute to the development of an effective vaccine.
PLGA nanoparticles
Considered a promising antigen target,123 Tc24 is a calcium-acyl exchange protein located in the parasite flagellar pocket which is expressed at all stages of T. cruzi development.124 Prophylactic immunization with Tc24 is protective against T. cruzi infection125,126 and induces high levels of antigen-specific IFN-γ CD8+ cells, protecting against both parasitemia and cardiac pathology. In humans, recombinant Tc24 has been used as an antigen for Chagas disease serodiagnosis and as a tool to monitor treatment success.127,128 A PLGA nanoparticle-based vaccine containing oligodeoxynucleotides with the CpG motif (CpG ODN) as an immunomodulatory adjuvant has been designed which acts as a TLR-9 agonist. CpG ODNs have been described as potent immune response stimulators of Th1-mediated CD8+ T cells.129,130
PLGA nanoparticles produced with CpG ODN and recombinant Tc24 protein provide a deposit effect similar to Aluminum hydroxide Gel (Alhydrogel®) in immunized mice, allowing the prolonged release of Tc24 antigen. In addition, the vaccine produced a skewed Th1 immune response, demonstrating a significant increase in antigenic splenocytes producing specific IFN-γ, IgG2a titers, and CD8+ T cell proliferative capacity. This study provided important evidence of the principle that PLGA nanoparticles can be used as a delivery system to improve both the immunogenicity and efficacy of a protein-based vaccine for Chagas disease.131
In summary, the desirable features of vaccine against T. cruzi infection are the ability to decrease the parasitic burden, inflammation, and damage of heart tissue or increase host survival.132–135 It has been shown that nanostructural systems or immunogenic vehicles are better than conventional therapies, showing a system of directed-administration, such as the use of specific antibodies or ligands to transport drugs directly to their targets. Thus, a successful nanovaccine should induce a strong cellular immune response, with the activation of CD8+ T cells to effectively control the parasites.108–110
Conclusion
Nowadays, there are not enough data to predict whether the nanoparticles will offer improved effective antichagasic treatments. Preclinical data analyzed herein has not yet clearly shown the successive elimination of intracellular parasites such as in chronic asymptomatic cases, or non-rodent immunized animal models.30,36,41,42,50,59,95,136,137 Moreover, the only nanoparticulate system approved for clinical trials is liposomal amphotericin B (LAMB), however, it has been shown to have little anti-T. cruzi activity.138,139
Trypanocidal activity on different forms of T. cruzi would be essential for an excellent nanoparticulate system. However, neither amastigotes nor trypomastigotes are as effective in uptaking nanoparticles as epimastigotes.140 Ideally, nanoparticles must have a size smaller than 250 nm and the surface charges must depend on the established goal. For the acute phase of the disease, a neutral or slightly negative nanoparticle surface charge is desired for intravenous administration since nanoparticles reduce the adsorption of serum proteins, resulting in longer half-lives in the circulation141 and therefore, promoting a controlled drug release in the body. For indeterminate and chronic phases, nanoparticles with a positive surface charge would be desirable to reach infection sites since they have a higher rate of non-specific uptake in most cells.142 In addition, nanomaterial must function at different pH’s, allowing massive and selective targeting to infected cells, and a more drastic reduction in parasitemia levels.137 Finally, this review provided an overview of recent advances in development and design of new strategies to synthesize nanoparticulated systems for treatment against Chagas disease.
Acknowledgments
This work was supported by the Brazilian Federal Agency for the Support and Evaluation of Graduate Education (CAPES, grant 923/18 CAPES-COFECUB) and the Brazilian National Council for Scientific and Technological Development (CNPq, grants 433208/2016-3 and INCT - MCTI/CNPq/CAPES/FAPs 16/2014).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Echeverria LE, Morillo CA. American trypanosomiasis (Chagas disease). Infect Dis Clin. 2019;33(1):119–134. doi:10.1016/j.idc.2018.10.015
2. Andrade DV, Gollob KJ, Dutra WO. Acute Chagas disease: new global challenges for an old neglected disease. PLoS Negl Trop Dis. 2014;8(7):e3010. doi:10.1371/journal.pntd.0003010
3. Hotez PJ, Molyneux DH, Fenwick A, et al. Control of neglected tropical diseases. N Engl J Med. 2007;357(10):1018–1027. doi:10.1056/NEJMra064142
4. Prata A. Clinical and epidemiological aspects of Chagas disease. Lancet Infect Dis. 2001;1(2):92–100. doi:10.1016/S1473-3099(01)00065-2
5. Pérez-Molina JA, Molina I. Chagas disease. Lancet. 2018;391(10115):82–94. doi:10.1016/S0140-6736(17)31612-4
6. Sánchez-Valdéz FJ, Pérez Brandán C, Ferreira A, Basombrío MÁ. Gene-deleted live-attenuated Trypanosoma cruzi parasites as vaccines to protect against Chagas disease. Expert Rev Vaccines. 2015;14(5):681–697. doi:10.1586/14760584.2015.989989
7. Barratt GM. Therapeutic applications of colloidal drug carriers. Pharm Sci Technolo Today. 2000;3(5):163–171. doi:10.1016/S1461-5347(00)00255-8
8. Damhorst GL, Murtagh M, Rodriguez WR, Bashir R. Microfluidics and nanotechnology for detection of global infectious diseases. Proc IEEE. 2015;103(2):150–160. doi:10.1109/JPROC.2014.2385078
9. Galvin P, Thompson D, Ryan KB, et al. Nanoparticle-based drug delivery: case studies for cancer and cardiovascular applications. Cell Mol Life Sci. 2012;69(3):389–404. doi:10.1007/s00018-011-0856-6
10. Godin B, Sakamoto JH, Serda RE, Grattoni A, Bouamrani A, Ferrari M. Emerging applications of nanomedicine for the diagnosis and treatment of cardiovascular diseases. Trends Pharmacol Sci. 2010;31(5):199–205. doi:10.1016/j.tips.2010.01.003
11. Seigneuric R, Markey L, Nuyten DS, et al. From nanotechnology to nanomedicine: applications to cancer research. Curr Mol Med. 2010;10(7):640–652.
12. Bharali DJ, Mousa SA. Emerging nanomedicines for early cancer detection and improved treatment: current perspective and future promise. Pharmacol Ther. 2010;128(2):324–335. doi:10.1016/j.pharmthera.2010.07.007
13. Dellian M, Yuan F, Trubetskoy V, Torchilin V, Jain R. Vascular permeability in a human tumour xenograft: molecular charge dependence. Br J Cancer. 2000;82(9):1513.
14. He C, Hu Y, Yin L, Tang C, Yin C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials. 2010;31(13):3657–3666. doi:10.1016/j.biomaterials.2010.01.065
15. Sankaranarayanan J, Mahmoud EA, Kim G, Morachis JM, Almutairi A. Multiresponse strategies to modulate burst degradation and release from nanoparticles. ACS Nano. 2010;4(10):5930–5936. doi:10.1021/nn100968e
16. Nel AE, Mädler L, Velegol D, et al. Understanding biophysicochemical interactions at the nano–bio interface. Nat Mater. 2009;8(7):543. doi:10.1038/nmat2442
17. Arvizo RR, Miranda OR, Moyano DF, et al. Modulating pharmacokinetics, tumor uptake and biodistribution by engineered nanoparticles. PLoS One. 2011;6(9):e24374. doi:10.1371/journal.pone.0024374
18. Gratton SE, Ropp PA, Pohlhaus PD, et al. The effect of particle design on cellular internalization pathways. Proc Natl Acad Sci. 2008;105(33):11613–11618. doi:10.1073/pnas.0801763105
19. Miller CR, Bondurant B, McLean SD, McGovern KA, O’Brien DF. Liposome− cell interactions in vitro: effect of liposome surface charge on the binding and endocytosis of conventional and sterically stabilized liposomes. Biochemistry. 1998;37(37):12875–12883. doi:10.1021/bi980096y
20. Osaka T, Nakanishi T, Shanmugam S, Takahama S, Zhang H. Effect of surface charge of magnetite nanoparticles on their internalization into breast cancer and umbilical vein endothelial cells. Colloids Surf B Biointerfaces. 2009;71(2):325–330. doi:10.1016/j.colsurfb.2009.03.004
21. Perrie Y, Mohammed AR, Kirby DJ, McNeil SE, Bramwell VW. Vaccine adjuvant systems: enhancing the efficacy of sub-unit protein antigens. Int J Pharm. 2008;364(2):272–280. doi:10.1016/j.ijpharm.2008.04.036
22. Serda RE, Gu J, Bhavane RC, et al. The association of silicon microparticles with endothelial cells in drug delivery to the vasculature. Biomaterials. 2009;30(13):2440–2448. doi:10.1016/j.biomaterials.2009.01.019
23. Valente I, Celasco E, Marchisio D, Barresi A. Nanoprecipitation in confined impinging jets mixers: production, characterization and scale-up of pegylated nanospheres and nanocapsules for pharmaceutical use. Chem Eng Sci. 2012;77:217–227. doi:10.1016/j.ces.2012.02.050
24. Prabhu RH, Patravale VB, Joshi MD. Polymeric nanoparticles for targeted treatment in oncology: current insights. Int J Nanomedicine. 2015;10:1001–1018. doi:10.2147/IJN.S56932
25. Hans M, Lowman A. Biodegradable nanoparticles for drug delivery and targeting. Curr Opin Solid St Mate Sci. 2002;6(4):319–327. doi:10.1016/S1359-0286(02)00117-1
26. Hu X, Zou G. Biodegradable polymeric nanoparticles as drug delivery systems. Amino Acids Biotic Resour. 2003;2(1–2):1–20.
27. Gonzalez-Martin G, Merino I, Rodriguez-Cabezas MN, Torres M, Nunez R, Osuna A. Characterization and trypanocidal activity of nifurtimox-containing and empty nanoparticles of polyethylcyanoacrylates. J Pharm Pharmacol. 1998;50(1):29–35.
28. Müller RH, Lherm C, Herbert J, Couvreur P. In vitro model for the degradation of alkylcyanoacrylate nanoparticles. Biomaterials. 1990;11(8):590–595.
29. Gonzalez-Martin G, Figueroa C, Merino I, Osuna A. Allopurinol encapsulated in polycyanoacrylate nanoparticles as potential lysosomatropic carrier: preparation and trypanocidal activity. Eur J Pharm Biopharm. 2000;49(2):137–142.
30. Molina J, Urbina J, Gref R, Brener Z, Rodriguez JM. Cure of experimental Chagas’ disease by the bis-triazole DO870 incorporated into ‘stealth’ polyethyleneglycol-polylactide nanospheres. J Antimicrob Chemother. 2001;47(1):101–104. doi:10.1093/jac/47.1.101
31. GFP DS, Yokoyama-Yasunaka JK, Seabra AB, Miguel DC, de Oliveira MG, Uliana SRB. Leishmanicidal activity of primary S-nitrosothiols against Leishmania major and Leishmania amazonensis: implications for the treatment of cutaneous leishmaniasis. Nitric Oxide. 2006;15(3):209–216. doi:10.1016/j.niox.2006.01.011
32. Gil-Jaramillo N, Motta FN, Favali CB, Bastos I, Santana JM. Dendritic cells: a double-edged sword in immune responses during Chagas disease. Front Microbiol. 2016;7:1076. doi:10.3389/fmicb.2016.01076
33. Gutierrez FR, Mineo TW, Pavanelli WR, Guedes PM, Silva JS. The effects of nitric oxide on the immune system during Trypanosoma cruzi infection. Mem Inst Oswaldo Cruz. 2009;104(2):236–245. doi:10.1590/s0074-02762009000900030
34. Seabra AB, Da Silva R, De Souza GF, De Oliveira MG. Antithrombogenic Polynitrosated Polyester/Poly (methyl methacrylate) blend for the coating of blood‐contacting surfaces. Artif Organs. 2008;32(4):262–267. doi:10.1111/j.1525-1594.2008.00540.x
35. Amadeu TP, Seabra AB, De Oliveira MG, Costa AM. S‐nitrosoglutathione‐containing hydrogel accelerates rat cutaneous wound repair. J Eur Acad Dermatol Venereol. 2007;21(5):629–637. doi:10.1111/j.1468-3083.2006.02032.x
36. Seabra AB, Kitice NA, Pelegrino MT, et al. Nitric oxide-releasing polymeric nanoparticles against Trypanosoma cruzi. J Phys: Conf Ser. 2015;617:012020. doi:10.1088/1742-6596/617/1/012020
37. Kohli K, Chopra S, Dhar D, Arora S, Khar RK. Self-emulsifying drug delivery systems: an approach to enhance oral bioavailability. Drug Discov Today. 2010;15(21–22):958–965. doi:10.1016/j.drudis.2010.08.007
38. Pouton CW. Lipid formulations for oral administration of drugs: non-emulsifying, self-emulsifying and ‘self-microemulsifying’drug delivery systems. Eur J Pharm Sci. 2000;11(2):S93–S98.
39. Sposito PA, Mazzeti AL, Faria CD, et al. Ravuconazole self-emulsifying delivery system: in vitro activity against Trypanosoma cruzi amastigotes and in vivo toxicity. Int J Nanomedicine. 2017;12:3785–3799. doi:10.2147/IJN.S133708
40. Branquinho RT, Mosqueira VCF, JCV DO-S, Simoes-Silva MR, Saude-Guimaraes DA, de Lana M. Sesquiterpene lactone in nanostructured parenteral dosage form is efficacious in experimental Chagas disease. Antimicrob Agents Chemother. 2014;58(4):2067–2075. doi:10.1128/AAC.00617-13
41. Abriata JP, Eloy JO, Riul TB, Campos PM, Baruffi MD, Marchetti JM. Poly-epsilon-caprolactone nanoparticles enhance ursolic acid in vivo efficacy against Trypanosoma cruzi infection. Mater Sci Eng C Mater Biol Appl. 2017;77:1196–1203. doi:10.1016/j.msec.2017.03.266
42. Rial MS, Scalise ML, Arrua EC, Esteva MI, Salomon CJ, Fichera LE. Elucidating the impact of low doses of nano-formulated benznidazole in acute experimental Chagas disease. PLoS Negl Trop Dis. 2017;11(12):e0006119. doi:10.1371/journal.pntd.0006119
43. Tessarolo LD, Mello CP, Lima DB, et al. Nanoencapsulation of benznidazole in calcium carbonate increases its selectivity to Trypanosoma cruzi. Parasitology. 2018;145(9):1–8. doi:10.1017/S0031182017001858
44. Fujiwara M, Shiokawa K, Morigaki K, Zhu Y, Nakahara Y. Calcium carbonate microcapsules encapsulating biomacromolecules. Chem Eng J. 2008;137(1):14–22. doi:10.1016/j.cej.2007.09.010
45. Seremeta KP, Arrúa EC, Okulik NB, Salomon CJ. Development and characterization of benznidazole nano-and microparticles: a new tool for pediatric treatment of Chagas disease? Colloids Surf B Biointerfaces. 2019;177:169–177. doi:10.1016/j.colsurfb.2019.01.039
46. Huang SL. Liposomes in ultrasonic drug and gene delivery. Adv Drug Deliv Rev. 2008;60(10):1167–1176. doi:10.1016/j.addr.2008.03.003
47. Crommelin DJ, Schreier H. Liposomes. In: Kreuter J, editor. Colloidal Drug Delivery Systems. Boca Raton: CRC Press; 2014:85–159.
48. Mody N, Tekade RK, Mehra NK, Chopdey P, Jain NK. Dendrimer, liposomes, carbon nanotubes and PLGA nanoparticles: one platform assessment of drug delivery potential. AAPS PharmSciTech. 2014;15(2):388–399. doi:10.1208/s12249-014-0073-3
49. Samad A, Sultana Y, Aqil M. Liposomal drug delivery systems: an update review. Curr Drug Deliv. 2007;4(4):297–305.
50. Morilla MJ, Montanari JA, Prieto MJ, Lopez MO, Petray PB, Romero EL. Intravenous liposomal benznidazole as trypanocidal agent: increasing drug delivery to liver is not enough. Int J Pharm. 2004;278(2):311–318. doi:10.1016/j.ijpharm.2004.03.025
51. Soto CDA, Mirkin GA, Solana ME, Cappa SMG. Trypanosoma cruzi infection modulates in vivo expression of major histocompatibility complex class II molecules on antigen-presenting cells and T-cell stimulatory activity of dendritic cells in a strain-dependent manner. Infect Immun. 2003;71(3):1194–1199. doi:10.1128/iai.71.3.1194-1199.2003
52. Morilla MJ, Montanari J, Frank F, et al. Etanidazole in pH-sensitive liposomes: design, characterization and in vitro/in vivo anti-Trypanosoma cruzi activity. J Control Release. 2005;103(3):599–607. doi:10.1016/j.jconrel.2004.12.012
53. Espada R, Valdespina S, Alfonso C, Rivas G, Ballesteros MP, Torrado JJ. Effect of aggregation state on the toxicity of different amphotericin B preparations. Int J Pharm. 2008;361(1–2):64–69. doi:10.1016/j.ijpharm.2008.05.013
54. Torrado J, Espada R, Ballesteros M, Torrado-Santiago S. Amphotericin B formulations and drug targeting. J Pharm Sci. 2008;97(7):2405–2425. doi:10.1002/jps.21179
55. Actor P, Wind S, Pagano J. Potentiation of amphotericin B activity against trypanosoma congolense in mice. Proc Soc Exp Biol Med. 1962;110(3):409–412. doi:10.3181/00379727-110-27532
56. Rolon M, Seco EM, Vega C, et al. Selective activity of polyene macrolides produced by genetically modified Streptomyces on Trypanosoma cruzi. Int J Antimicrob Agents. 2006;28(2):104–109. doi:10.1016/j.ijantimicag.2006.02.025
57. Yardley V, Croft SL. In vitro and in vivo activity of amphotericin B-lipid formulations against experimental Trypanosoma cruzi infections. Am J Trop Med Hyg. 1999;61(2):193–197. doi:10.4269/ajtmh.1999.61.193
58. Torrado JJ, Serrano DR, Uchegbu IF. The oral delivery of amphotericin B. Ther Deliv. 2013;4(1):9–12.
59. Cencig S, Coltel N, Truyens C, Carlier Y. Parasitic loads in tissues of mice infected with trypanosoma cruzi and treated with AmBisome. PLoS Negl Trop Dis. 2011;5(6):e1216. doi:10.1371/journal.pntd.0001370
60. Tarleton R. Trypanosoma cruzi and Chagas disease: cause and effect. In: Tyler KM, Miles MA, editors. American Trypanosomiasis. World Class Parasites, vol 7. Boston: Springer; 2003:107–115.
61. Ferrari M. Frontiers in cancer nanomedicine: directing mass transport through biological barriers. Trends Biotechnol. 2010;28(4):181–188. doi:10.1016/j.tibtech.2009.12.007
62. Mehnert W, Mäder K. Solid lipid nanoparticles: production, characterization and applications. Adv Drug Deliv Rev. 2012;64(2–3):83–101. doi:10.1016/j.addr.2012.09.021
63. Müller RH, MaÈder K, Gohla S. Solid lipid nanoparticles (SLN) for controlled drug delivery–a review of the state of the art. Eur J Pharm Biopharm. 2000;50(1):161–177.
64. Tomiotto-Pellissier F, Miranda-Sapla MM, Machado LF, et al. Nanotechnology as a potential therapeutic alternative for schistosomiasis. Acta Trop. 2017;174:64–71. doi:10.1016/j.actatropica.2017.06.025
65. Wissing S, Kayser O, Müller R. Solid lipid nanoparticles for parenteral drug delivery. Adv Drug Deliv Rev. 2004;56(9):1257–1272. doi:10.1016/j.addr.2003.12.002
66. Muller RH. Solid lipid nanoparticles (SLN)-an alternative colloidal carrier system for controlled drug delivery. Eur J Pharm Biopharm. 1995;41(1):62–69.
67. Carneiro ZA, Maia PID, Sesti-Costa R, et al. In vitro and in vivo trypanocidal activity of H(2)bdtc-loaded solid lipid nanoparticles. PLoS Negl Trop Dis. 2014;8(5):e2847. doi:10.1371/journal.pntd.0002847
68. Morein B, Sundquist B, Höglund S, Dalsgaard K, Osterhaus A. Iscom, a novel structure for antigenic presentation of membrane proteins from enveloped viruses. Nature. 1984;308(5958):457. doi:10.1038/308457a0
69. Wilson NS, Duewell P, Yang B, et al. Inflammasome-dependent and -independent IL-18 production mediates immunity to the ISCOMATRIX adjuvant. J Immunol. 2014;192(7):3259–3268. doi:10.4049/jimmunol.1302011
70. Lendemans DG, Egert AM, Hook S, Rades T. Cage-like complexes formed by DOTAP, Quil-A and cholesterol. Int J Pharm. 2007;332(1–2):192–195. doi:10.1016/j.ijpharm.2006.09.029
71. Friede M, Hermand P, inventors; SmithKline Beechman Biologicals s.a., assignee. Vaccine Adjuvants. United States patent 6558670. 2003 May 6.
72. Sun HX, Xie Y, Ye YP. ISCOMs and ISCOMATRIX. Vaccine. 2009;27(33):4388–4401. doi:10.1016/j.vaccine.2009.05.032
73. Cruz-Bustos T, Gonzalez-Gonzalez G, Morales-Sanfrutos J, Megia-Fernandez A, Santoyo-Gonzalez F, Osuna A. Functionalization of immunostimulating complexes (ISCOMs) with lipid vinyl sulfones and their application in immunological techniques and therapy. Int J Nanomedicine. 2012;7(2):5941–5956. doi:10.2147/IJN.S35556
74. Nhavene EPF, Da Silva WM, Junior RRT, et al. Chitosan grafted into mesoporous silica nanoparticles as benznidazol carrier for Chagas diseases treatment. Microporous Mesoporous Mater. 2018;272:265–275. doi:10.1016/j.micromeso.2018.06.035
75. Eger I, Soares MJ. Endocytosis in Trypanosoma cruzi (Euglenozoa: kinetoplastea) epimastigotes: visualization of ingested transferrin-gold nanoparticle complexes by confocal laser microscopy. J Microbiol Methods. 2012;91(1):101–105. doi:10.1016/j.mimet.2012.07.013
76. Ho CC, Luo YH, Chuang TH, Yang CS, Ling YC, Lin P. Quantum dots induced monocyte chemotactic protein-1 expression via MyD88-dependent Toll-like receptor signaling pathways in macrophages. Toxicology. 2013;308:1–9. doi:10.1016/j.tox.2013.03.003
77. Hauck TS, Anderson RE, Fischer HC, Newbigging S, Chan WC. In vivo quantum-dot toxicity assessment. Small. 2010;6(1):138–144. doi:10.1002/smll.200900626
78. Wu T, Tang M. Toxicity of quantum dots on respiratory system. Inhal Toxicol. 2014;26(2):128–139. doi:10.3109/08958378.2013.871762
79. Chen N, He Y, Su Y, et al. The cytotoxicity of cadmium-based quantum dots. Biomaterials. 2012;33(5):1238–1244. doi:10.1016/j.biomaterials.2011.10.070
80. Nagy A, Hollingsworth JA, Hu B, et al. Functionalization-dependent induction of cellular survival pathways by CdSe quantum dots in primary normal human bronchial epithelial cells. ACS Nano. 2013;7(10):8397–8411. doi:10.1021/nn305532k
81. Stahl CV, Almeida DB, de Thomaz AA, et al. Studying nanotoxic effects of CdTe Quantum Dots in Trypanosoma cruzi. Colloidal Quantum Dots for Biomedical Applications V. Proc. SPIE 7575, Colloidal Quantum Dots for Biomedical Applications V, 757513; February 17; 2010; San Francisco, CA.
82. de Araujo SC, de Mattos AC, Teixeira HF, Coelho PM, Nelson DL, de Oliveira MC. Improvement of in vitro efficacy of a novel schistosomicidal drug by incorporation into nanoemulsions. Int J Pharm. 2007;337(1–2):307–315. doi:10.1016/j.ijpharm.2007.01.009
83. Sarker DK. Engineering of nanoemulsions for drug delivery. Curr Drug Deliv. 2005;2(4):297–310.
84. Jaiswal M, Dudhe R, Sharma P. Nanoemulsion: an advanced mode of drug delivery system. 3 Biotech. 2015;5(2):123–127. doi:10.1007/s13205-014-0214-0
85. Lovelyn C, Attama AA. Current state of nanoemulsions in drug delivery. J Biomater Nanobiotechnol. 2011;2(05):626. doi:10.4236/jbnb.2011.225075
86. de Oliveira ECV, Carneiro ZA, de Albuquerque S, Marchetti JM. Development and evaluation of a nanoemulsion containing ursolic acid: a promising trypanocidal agent. AAPS PharmSciTech. 2017;18(7):2551–2560. doi:10.1208/s12249-017-0736-y
87. Vermelho AB, Cardoso VD, Ricci E, Dos Santos EP, Supuran CT. Nanoemulsions of sulfonamide carbonic anhydrase inhibitors strongly inhibit the growth of Trypanosoma cruzi. J Enzyme Inhib Med Chem. 2017;33(1):139–146. doi:10.1080/14756366.2017.1405264
88. Streck L, Sarmento VH, de Menezes RP, Fernandes-Pedrosa MF, Martins AM, da Silva-Júnior AA. Tailoring microstructural, drug release properties, and antichagasic efficacy of biocompatible oil-in-water benznidazol-loaded nanoemulsions. Int J Pharm. 2019;555:36–48. doi:10.1016/j.ijpharm.2018.11.041
89. Torchilin VP. Micellar nanocarriers: pharmaceutical perspectives. Pharm Res. 2007;24(1):1. doi:10.1007/s11095-007-9271-y
90. Chiappetta DA, Poly SA. Poly (ethylene oxide)–poly (propylene oxide) block copolymer micelles as drug delivery agents: improved hydrosolubility, stability and bioavailability of drugs. Eur J Pharm Biopharm. 2007;66(3):303–317. doi:10.1016/j.ejpb.2007.03.022
91. Attia ABE, Ong ZY, Hedrick JL, et al. Mixed micelles self-assembled from block copolymers for drug delivery. Curr Opin Colloid Interface Sci. 2011;16(3):182–194. doi:10.1016/j.cocis.2010.10.003
92. Cabral H, Matsumoto Y, Mizuno K, et al. Accumulation of sub-100 nm polymeric micelles in poorly permeable tumours depends on size. Nat Nanotechnol. 2011;6(12):815. doi:10.1038/nnano.2011.166
93. Hu C-MJ, Zhang L. Therapeutic nanoparticles to combat cancer drug resistance. Curr Drug Metab. 2009;10(8):836–841.
94. Kwon GS, Kataoka K. Block copolymer micelles as long-circulating drug vehicles. Adv Drug Deliv Rev. 2012;64(2–3):237–245. doi:10.1016/j.addr.2012.09.016
95. Rolon M, Serrano DR, Lalatsa A, et al. Engineering oral and parenteral amorphous amphotericin B formulations against experimental trypanosoma cruzi infections. Mol Pharm. 2017;14(4):1095–1106. doi:10.1021/acs.molpharmaceut.6b01034
96. Huang L, Wei G, Sun X, et al. A tumor-targeted Ganetespib-zinc phthalocyanine conjugate for synergistic chemo-photodynamic therapy. Eur J Med Chem. 2018;151:294–303. doi:10.1016/j.ejmech.2018.03.077
97. Mai B, Gao Y, Li M, et al. Photodynamic antimicrobial chemotherapy for Staphylococcus aureus and multidrug-resistant bacterial burn infection in vitro and in vivo. Int J Nanomedicine. 2017;12:5915. doi:10.2147/IJN.S138185
98. Yuan Y, Liu Z-Q, Jin H, et al. Photodynamic antimicrobial chemotherapy with the novel amino acid-porphyrin conjugate 4I: in vitro and in vivo studies. PLoS One. 2017;12(5):e0176529. doi:10.1371/journal.pone.0176529
99. de Morais FAP, Enumo A, Gonçalves RS, et al. Hypericin photodynamic activity. Part III: in vitro evaluation in different nanocarriers against trypomastigotes of trypanosoma cruzi. Photochem Photobiol Sci. 2019;18(2):487–494. doi:10.1039/C8PP00444G
100. Jain KK. Nanodiagnostics: application of nanotechnology in molecular diagnostics. Expert Rev Mol Diagn. 2003;3(2):153–161. doi:10.1586/14737159.3.2.153
101. Singh RP, Oh BK, Choi JW. Application of peptide nucleic acid towards development of nanobiosensor arrays. Bioelectrochemistry. 2010;79(2):153–161. doi:10.1016/j.bioelechem.2010.02.004
102. Patolsky F, Zheng G, Lieber CM. Nanowire-based Biosensors. Anal Chem. 2006;78(13):4260–4269.
103. Pereira SV, Bertolino FA, Fernandez-Baldo MA, et al. A microfluidic device based on a screen-printed carbon electrode with electrodeposited gold nanoparticles for the detection of IgG anti-Trypanosoma cruzi antibodies. Analyst. 2011;136(22):4745–4751. doi:10.1039/c1an15569e
104. Castro-Sesquen YE, Gilman RH, Galdos-Cardenas G, et al. Use of a novel Chagas urine nanoparticle test (Chunap) for diagnosis of congenital Chagas disease. PLoS Negl Trop Dis. 2014;8(10):e3211. doi:10.1371/journal.pntd.0003211
105. Janissen R, Sahoo PK, Santos CA, et al. InP nanowire biosensor with tailored biofunctionalization: ultrasensitive and highly selective disease biomarker detection. Nano Lett. 2017;17(10):5938–5949. doi:10.1021/acs.nanolett.7b01803
106. Vaculovicova M, Michalek P, Krizkova S, Macka M, Adam V. Nanotechnology-based analytical approaches for detection of viruses. Anal Methods. 2017;9(16):2375–2391. doi:10.1039/C7AY00048K
107. M-I R-G, L-J V-G, Beyssen D, Sarry F, Reyna M-A, Ibarra-Cerdeña C-N. Biosensors to diagnose chagas disease: a brief review. Sensors. 2017;17(11):2629. doi:10.3390/s17050968
108. Araújo AF, de Alencar BC, Vasconcelos JRC, et al. CD8+-T-cell-dependent control of Trypanosoma cruzi infection in a highly susceptible mouse strain after immunization with recombinant proteins based on amastigote surface protein 2. Infect Immun. 2005;73(9):6017–6025. doi:10.1128/IAI.73.9.6017-6025.2005
109. de Alencar BC, Araújo AF, Penido ML, Gazzinelli RT, Rodrigues MM. Cross-priming of long lived protective CD8+ T cells against Trypanosoma cruzi infection: importance of a TLR9 agonist and CD4+ T cells. Vaccine. 2007;25(32):6018–6027. doi:10.1016/j.vaccine.2007.05.022
110. Hoft DF, Schnapp AR, Eickhoff CS, Roodman ST. Involvement of CD4+ Th1 cells in systemic immunity protective against primary and secondary challenges with trypanosoma cruzi. Infect Immun. 2000;68(1):197–204. doi:10.1128/iai.68.1.197-204.2000
111. Low HP, Santos MA, Wizel B, Tarleton RL. Amastigote surface proteins of Trypanosoma cruzi are targets for CD8+ CTL. J Immunol. 1998;160(4):1817–1823.
112. Miller MJ, Wrightsman RA, Manning JE. Trypanosoma cruzi: protective immunity in mice immunized with paraflagellar rod proteins is associated with a T-helper type 1 response. Exp Parasitol. 1996;84(2):156–167. doi:10.1006/expr.1996.0101
113. Tarleton R. Depletion of CD8+ T cells increases susceptibility and reverses vaccine-induced immunity in mice infected with trypanosoma cruzi. J Immunol. 1990;144(2):717–724.
114. Tarleton RL, Sun J, Zhang L, Postan M. Depletion of T-cell subpopulations results in exacerbation of myocarditis and parasitism in experimental Chagas’ disease. Infect Immun. 1994;62(5):1820–1829.
115. Dumonteil E, Bottazzi ME, Zhan B, et al. Accelerating the development of a therapeutic vaccine for human Chagas disease: rationale and prospects. Expert Rev Vaccines. 2012;11(9):1043–1055. doi:10.1586/erv.12.85
116. Mohan T, Verma P, Rao DN. Novel adjuvants & delivery vehicles for vaccines development: a road ahead. Indian J Med Res. 2013;138(5):779.
117. Awate S, Babiuk LA, Mutwiri G. Mechanisms of action of adjuvants. Front Immunol. 2013;4:114. doi:10.3389/fimmu.2013.00114
118. Bertona D, Pujato N, Bontempi I, et al. Development and assessment of a new cage-like particle adjuvant. J Pharm Pharmacol. 2017;69(10):1293–1303. doi:10.1111/jphp.12768
119. Kita Y, Tanaka T, Yoshida S, et al. Novel recombinant BCG and DNA-vaccination against tuberculosis in a cynomolgus monkey model. Vaccine. 2005;23(17–18):2132–2135. doi:10.1016/j.vaccine.2005.01.057
120. Santos FR, Ferraz DB, Daghastanli KR, Ramalho-Pinto FJ, Ciancaglini P. Mimetic membrane system to carry multiple antigenic proteins from Leishmania amazonensis. J Membr Biol. 2006;210(3):173–181. doi:10.1007/s00232-006-0005-6
121. Mesa C, de León J, Rigley K, Fernández LE. Very small size proteoliposomes derived from Neisseria meningitidis: an effective adjuvant for Th1 induction and dendritic cell activation. Vaccine. 2004;22(23–24):3045–3052. doi:10.1016/j.vaccine.2004.02.010
122. Migliaccio V, Santos FR, Ciancaglini P, Ramalho-Pinto FJ. Use of proteoliposome as a vaccine against Trypanosoma cruzi in mice. Chem Phys Lipids. 2008;152(2):86–94. doi:10.1016/j.chemphyslip.2007.12.003
123. Engman D, Krause K, Blumin J, Kim K, Kirchhoff L, Donelson J. A novel flagellar Ca2+-binding protein in trypanosomes. J Biol Chem. 1989;264(31):18627–18631.
124. Krautz G, Kissinger J, Krettli A. The targets of the lytic antibody response against Trypanosoma cruzi. Parasitol Today. 2000;16(1):31–34.
125. Taibi A, Plumas-Marty B, Guevara-Espinoza A, et al. Trypanosoma cruzi: immunity-induced in mice and rats by trypomastigote excretory-secretory antigens and identification of a peptide sequence containing a T cell epitope with protective activity. J Immunol. 1993;151(5):2676–2689.
126. Taibi A, Espinoza AG, Ouaissi A. Trypanosoma cruzi: analysis of cellular and humoral response against a protective recombinant antigen during experimental Chagas’ disease. Immunol Lett. 1995;48(3):193–200.
127. Godsel LM, Tibbetts RS, Olson CL, Chaudoir BM, Engman DM. Utility of recombinant flagellar calcium-binding protein for serodiagnosis of Trypanosoma cruzi infection. J Clin Microbiol. 1995;33(8):2082–2085.
128. Guevara A, Taibi A, Alava J, Guderian R, Ouaissi A. Use of a recombinant Trypanosoma cruzi protein antigen to monitor cure of Chagas disease. Trans R Soc Trop Med Hyg. 1995;89(4):447–448. doi:10.1016/0035-9203(95)90052-7
129. Vabulas RM, Pircher H, Lipford GB, Häcker H, Wagner H. CpG-DNA activates in vivo T cell epitope presenting dendritic cells to trigger protective antiviral cytotoxic T cell responses. J Immunol. 2000;164(5):2372–2378. doi:10.4049/jimmunol.164.5.2372
130. Speiser DE, Liénard D, Rufer N, et al. Rapid and strong human CD8+ T cell responses to vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. J Clin Invest. 2005;115(3):739–746. doi:10.1172/JCI23373
131. Barry MA, Wang Q, Jones KM, et al. A therapeutic nanoparticle vaccine against Trypanosoma cruzi in a BALB/c mouse model of Chagas disease. Hum Vaccin Immunother. 2016;12(4):976–987. doi:10.1080/21645515.2015.1119346
132. Hyland KV, Leon JS, Daniels MD, et al. Modulation of autoimmunity by treatment of an infectious disease. Infect Immun. 2007;75(7):3641–3650. doi:10.1128/IAI.00423-07
133. Marin-Neto JA, Cunha-Neto E, Maciel BC, Simões MV. Pathogenesis of chronic Chagas heart disease. Circulation. 2007;115(9):1109. doi:10.1161/CIRCULATIONAHA.106.624296
134. Rocha M, Ribeiro A, Teixeira MM. Clinical management of chronic Chagas cardiomyopathy. Front Biosci. 2003;8(1):44–54. doi:10.2741/926
135. Tarleton RL. Parasite persistence in the aetiology of Chagas disease. Int J Parasitol. 2001;31(5–6):550–554.
136. Branquinho RT, Pound-Lana G, Milagre MM, et al. Increased body exposure to new anti-trypanosomal through nanoencapsulation. Sci Rep. 2017;7(1):8429. doi:10.1038/s41598-017-08469-x
137. Morilla MJ, Prieto MJ, Romero EL. Benznidazole vs benznidazole in multilamellar liposomes: how different they interact with blood components? Mem Inst Oswaldo Cruz. 2005;100(2):213–219. doi:10.1590/s0074-02762005000200017
138. Alderson NE. Is special FDA regulation of nanomedicine needed? A conversation with Norris E. Alderson. Interview by Barbara J Culliton. Health Aff (Millwood). 2008;27(4):w315–w317. doi:10.1377/hlthaff.27.4.w315
139. Clemons KV, Sobel RA, Martinez M, Correa-Oliveira R, Stevens DA. Lack of efficacy of liposomal amphotericin B against acute and chronic trypanosoma cruzi infection in mice. Am J Trop Med Hyg. 2017;97(4):1141–1146. doi:10.4269/ajtmh.16-0975
140. de Souza W, Sant’Anna C, Cunha-e-Silva NL. Electron microscopy and cytochemistry analysis of the endocytic pathway of pathogenic protozoa. Prog Histochem Cytochem. 2009;44(2):67–124. doi:10.1016/j.proghi.2009.01.001
141. Alexis F, Pridgen E, Molnar LK, Farokhzad OC. Factors affecting the clearance and biodistribution of polymeric nanoparticles. Mol Pharm. 2008;5(4):505–515. doi:10.1021/mp800051m
142. Gentile F, Chiappini C, Fine D, et al. The effect of shape on the margination dynamics of non-neutrally buoyant particles in two-dimensional shear flows. J Biomech. 2008;41(10):2312–2318. doi:10.1016/j.jbiomech.2008.03.021
143. Yoshihara E, Tachibana H, Nakae T. Trypanocidal activity of the stearylamine-bearing liposome in vitro. Life Sci. 1987;40(22):2153–2159. doi:10.1016/0024-3205(87)90005-1
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.