")
Back to Journals » OncoTargets and Therapy » Volume 13
Acetyl-11-Keto-β-Boswellic Acid Exerts the Anti-Cancer Effects via Cell Cycle Arrest, Apoptosis Induction and Autophagy Suppression in Non-Small Cell Lung Cancer Cells
Authors Lv M , Shao S, Zhang Q, Zhuang X , Qiao T
Received 28 October 2019
Accepted for publication 9 January 2020
Published 23 January 2020 Volume 2020:13 Pages 733—744
DOI https://doi.org/10.2147/OTT.S236346
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Minghe Lv, Shali Shao, Qi Zhang, Xibing Zhuang, Tiankui Qiao
Center for Tumor Diagnosis and Therapy, Jinshan Hospital, Fudan University, Shanghai 201508, People’s Republic of China
Correspondence: Tiankui Qiao
Center for Tumor Diagnosis and Therapy, Jinshan Hospital, Fudan University, Jinshan District, Shanghai 201508, People’s Republic of China
Email [email protected]
Objective: Acetyl-11-keto-β-boswellic acid (AKBA) is a triterpenoid, which is the main component of boswellic acid from Boswellia Serrata, a medicinal plant that has shown immense potential in anti-cancer therapy. This study aims to explore the roles and molecular mechanisms of AKBA on cell behavior in non-small cell lung cancer (NSCLC) cells.
Materials and Methods: The effects of AKBA on the cell viability in A549, H460, H1299, and BEAS-2B cells were determined by the CCK-8 assay. The colony formation assay was used to identify the effects of AKBA on cell proliferation. Potential roles of AKBA in regulating the cell cycle, apoptosis, and autophagy in A549 were evaluated by flow cytometry, Western blotting, reverse transcription-polymerase chain reaction (PCR) and immunofluorescence (IF).
Results: AKBA reduced cell viability in A549, H460, H1299, and BEAS-2B. In A549 cells, AKBA suppressed the clone formation, arrested the cell cycle at the G0/G1 phase, induced cellular apoptosis. We found that AKBA suppressed the formation of autolysosome, and decreased the expression levels of Beclin-1, LC3A/B-I, and LC3A/B-II proteins. Furthermore, AKBA also inhibited the expression levels of PI3K/Akt signaling pathway proteins.
Conclusion: AKBA exerts the anti-cancer effects via cell cycle arrest, apoptosis induction, and autophagy suppression in NSCLC cells. This body of evidence supports the potential of AKBA as a promising drug in the treatment of NSCLC.
Keywords: Acetyl-11-keto-β-boswellic acid, cell cycle, apoptosis, autophagy, non-small cell lung cancer
Introduction
Lung cancer is the most common cause of malignancies cancer-related deaths worldwide.1 Non-small cell lung cancer (NSCLC) is the most commonly diagnosed type of lung cancer, accounting for approximately 85% of all cases.2 According to the latest cancer statistical analysis,3 the new cases and deaths from lung cancer rank the first among all cancers. A large proportion of lung cancer patients are diagnosed with advanced-stage diseases and have lost the chance for surgical operation when they report to the hospital for therapy. Usually, traditional chemotherapy and radiotherapy play an irreplaceable role in the whole therapy for lung cancer;4 however, just 70% of the patients benefit from these due to chemotherapy and radiotherapy resistance. Therefore, it is vital to find new therapy measures for improving the survival quality of lung cancer patients.
Traditional Chinese medicine (TCM), is popular in healthcare systems among Chinese mainland and East Asian populations. TCM has been commonly used to improve the adverse effects of conventional therapy in patients with lung cancer,5 esophageal cancer,6 and liver cancer,7 especially those with NSCLC who received combined chemotherapy and radiotherapy.8 Hence, TCM has become a research focus because it has a broad application prospect in anti-tumor.
Acetyl-11-keto-β-boswellic acid (AKBA) is a pentacyclic triterpene, which is the main component of boswellic acid from Boswellia Serrata that promotes blood circulation to remove blood stasis. Boswellia Serrata is a medicinal plant that has been proved to reveal the immense potential in combating cancer, extensively known as Indian olibanum. Boswellic acid promotes blood circulation and removes wind, relieving muscle pain and swelling; thus, it is widely used in the treatment of rheumatoid arthritis and osteoarthritis.9 As an anti-inflammatory agent, boswellic acid down-regulates the TNF-α expression and suppresses the activity of active human recombinant GST-IKKα and His-IKKβ.10 Boswellic acid also inhibits the growth factors, proinflammatory interleukins,11 NF-ƙB, and NF- ƙB-regulated gene expression.12 Meanwhile, boswellic acid has been shown to noncompetitively inhibit 5-lipoxygenase and topoisomerase I and II.13,14 However, whether AKBA can exert the anti-cancer effects in NSCLC cell lines is unknown. Here, we aimed to explore in-depth the potential role and the mechanism of AKBA in combating NSCLC lines.
Materials and Methods
Reagents
Purified AKBA was supplied by the Duma Biotechnology (Shanghai, China), dissolved in dimethyl sulfoxide (DMSO, Sigma, Louis, Missouri, USA) at 20 mg/mL as a stock solution stored at −20°C until use. The DMSO concentration of each treatment group was less than or equal to 0.1%.
Cell Lines and Cell Culture
The human NSCLC cell line A549 was purchased from the Cell Bank of the China Science Academy (Shanghai, China). The normal human lung epithelium cell line BEAS-2B, and the human NSCLC cell lines H460 and H1299 were purchased from Cell Research (Shanghai, China). A549, H460, and H1299 were maintained in RPMI-1640 medium (Sigma, Louis, Missouri, USA) containing 10% fetal bovine serum (Biological Industries, Israel), and all cells were cultured at 37°C under 5% CO2. BEAS-2B was cultured with complete medium for bronchial epithelial cells (Cell Research, Shanghai, China).
Cell Proliferation Assay
The cells were seeded into 96 well-plates at a density of 5×103 cells per well. Cell viability was determined at 24 h, 48 h, and 72 h using the Cell Counting Kit-8 Assay Kit (Do Jindo Laboratories, Kumamoto, Japan). The experiments were conducted according to the manufacturer’s protocol of a cell cytotoxicity assay kit.
Clone Formation Experiment
The cells were seeded into 6-well plates at 2×105 cells per well. After each group accepted corresponding treatment for 48 h, A549 cells were digested with Trypsin (Keygen, Biotech, China) to reseed to 6-well plates at 500 cells per well and continually cultured for 8–10 days.The colonies were stained with crystal violet staining solution (Beyotime Biotechnology, Shanghai, China)
Flow Cytometric Determination of the Cell Cycle Stage
To determine the cell cycle distribution, about 2×105 cells were collected in a flow tube after drug treatment and fixed in 70% ethyl alcohol at −20°C overnight. Then, the cells were incubated with PI RNase A (BD Biosciences, Franklin, NJ, USA) for 15 mins. The fractions of the cells in the G0/G1, S and G2/M phases were analyzed by flow cytometry.
Flow Cytometric Determination of Apoptotic Cells
The percentage of apoptosis was measured by Annexin V-FITC Apoptosis Detection Kit (Do Jindo Laboratories, Kumamoto, Japan), according to the manufacturer’s protocol. The cells (2×105) were seeded into 6-well plates and grown to 70% confluence. Then, cells were treated with different concentrations of AKBA for 48 h, washed twice with 1× binding buffer, and stained with Annexin V-FITC antibody and propidium iodide (PI) in the dark on the ice for 15 mins. After antibody incubation, cells were resuspended in 400 μL of 1× binding buffer and were analyzed using a flow cytometer.
Western Blot Analysis
The cells were seeded into 6-well plates and treated with AKBA for the indicated time. Then, the liquid supernatant was taken out and the cells were washed three times on the surface of the 6-well plate. Cell lysates were prepared, and the concentration of protein was quantified using a BCA protein assay kit (ThermoFisher, USA). Equal amounts of cellular protein from each sample were applied to 8% to 12% sodium dodecyl sulfate (SDS)-polyacrylamide gels, transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA, USA), and probed with specific antibodies followed by exposure to a horseradish peroxidase–conjugated goat anti-mouse or goat anti-rabbit antibody and secondary antibodies (Cell Signaling Technology, Danvers, MA, USA). The immune complexes were visualized using a horseradish peroxidase-conjugated antibody, followed by a chemiluminescence reagent (Millipore, Bedford, MA, USA) and detected on photographic film.
Reverse Transcription-Polymerase Chain Reaction (PCR)
RNA isolation was performed using the RNA Purification Kit (Yi Shan Biotechnology Company, Shanghai, China). cDNA was prepared using the 5× Reverse Transcription Master Mix (Takara, Osaka, Japan) and was performed according to the manufacturer’s protocol. Primers used in these experiments were as follows: β-Actin, forward 5ʹ-CATTGCCGACAGGATGCAG-3ʹ, reverse 5ʹ-CTCGTCATACTCCTGCTTGCTG-3ʹ; P21, forward 5ʹ-CATGTGGACCTGTCACTGTCTTGTA-3ʹ, reverse 5ʹ-GAAGATCAGCCGGCGTTG-3ʹ; P27, forward 5ʹ-CAATGCCGGTTCTGTGGAG-3ʹ, reverse 5ʹ-TCCATTCCATGAAGTCAGCGATA-3ʹ. After reverse transcription, the cDNA product was amplified by PCR with Taq DNA polymerase (Takara, Osaka, Japan).
Immunofluorescence (IF)
To detect the formation of autolysosome, the cells were seeded into 24-well plates at 2 × 104 per well, and treated with AKBA for 48 h. Then cells were washed with RPMI-1640 medium (Sigma, Louis, Missouri, USA), and treated with DALGreen Working Solution (Do Jindo Laboratories, Kumamoto, Japan) for 30 mins. After treatment with DALGreen Working Solution, cells were washed with RPMI-1640 medium again, and cultured with autophagy inducer rapamycin (Ji Kai Gene Technology Company, Shanghai, China) for 24 h. The cells were observed under a confocal microscope.
Statistical Analysis
GraphPad Prism 7 (GraphPad Software, San Diego, CA, USA) and Microsoft Office Excel 2017(Microsoft, USA) were used for data analysis. Statistical significance (P <0.05) was determined by a one-way analysis of variance (ANOVA) and Tukey’s test. At least three independent samples were analyzed in each experiment.
Results
AKBA Inhibited the Proliferation of Human NSCLC Cell Lines and Normal Human Lung Epithelial Cells
AKBA reduced the cell viability of the NSCLC cell lines, A549, H460, and H1299, and inhibited the proliferation in a dose-dependent manner (Figure 1A–C). Fifty percent inhibitory concentration (IC50) values for AKBA on A549 at 24 h, 48 h, and 72 h were 11.52 μg/mL, 9.03 μg/mL, and 7.41 μg/mL. IC50 values for AKBA on H460 at 24 h, 48 h, and 72 h were 63.08 μg/mL, 33.25 μg/mL, and 22.3 μg/mL. IC50 values for AKBA on H1299 at 24 h, 48 h, and 72 h were 204.6 μg/mL, 31.62 μg/mL, and 25.17 μg/mL. By comparing the cell viability of AKBA on A549, H460, and H1299, we found that AKBA can inhibit the cell viability for A549 more significantly than other NSCLC lines. To demonstrate the cytotoxicity of AKBA, we similarly used CCK8 assay to test the viability of normal human lung epithelial cell line BEAS-2B treated with AKBA at 0, 5, 10, 15, 18, and 20 μg/mL (Figure 1D–F). We found that AKBA at equal concentrations had a greater lethality in A549 rather than BEAS-2B, which suggested that AKBA might be an effective drug for adenocarcinoma in NSCLC. In addition, we found that AKBA at 5, 10, and 20 μg/mL all suppressed the cell viability of A549 in a time-dependent manner, and AKBA at 20 μg/mL also showed time-dependence in H460, H1299, and BEAS-2B (Figure 1G–I). Therefore, AKBA could exert anti-cancer effects in NSCLC cell lines.
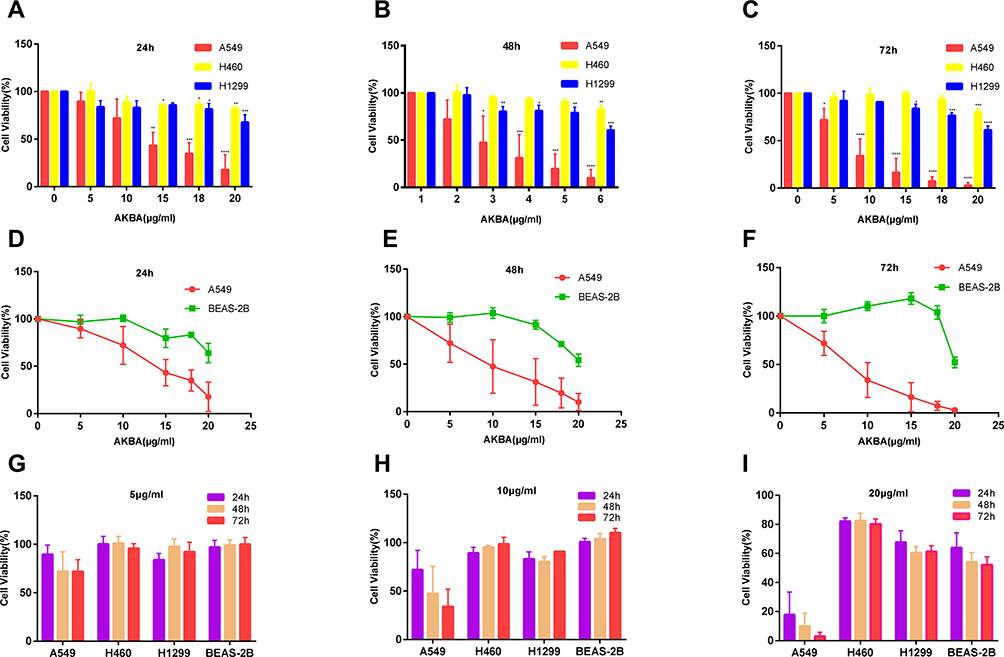 |
Figure 1 Effects of AKBA on the cell viability in human NSCLC cell lines A549, H460, and H1299 and normal human lung epithelium cell line BEAS-2B. (A–C) CCK8 assay showed that AKBA inhibited the cell viability of A549, H460, and H1299 in a dose-dependent manner. (D–F) Cytotoxicity test showed the comparison of cell viability between A549 and BEAS-2B. (G–I) The cell viability of A549, H460, H1299, and BEAS-2B was suppressed after treatment with AKBA at 5, 10, and 20 μg/mL in a time-dependent or independent manner. The experiments were performed in triplicate, and the data were presented as mean ± SD, *P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001, vs the AKBA-untreated control group (0 μg/mL). |
AKBA Suppressed the Clone Formation and Altered the Cell Morphology in Human NSCLC Cell Line A549
According to the previous results, we selected NSCLC cell line A549 for further researches. AKBA treatment resulted in a clone formation inhibition in a dose-dependent manner compared with the AKBA-untreated control group in A549 (Figure 2A) and decreased the clone number (Figure 2B). With the increasing treatment dose of AKBA, the cellular morphology was altered, and the cellular numbers were decreased (Figure 2C). These data indicated that AKBA inhibited the proliferation of A549.
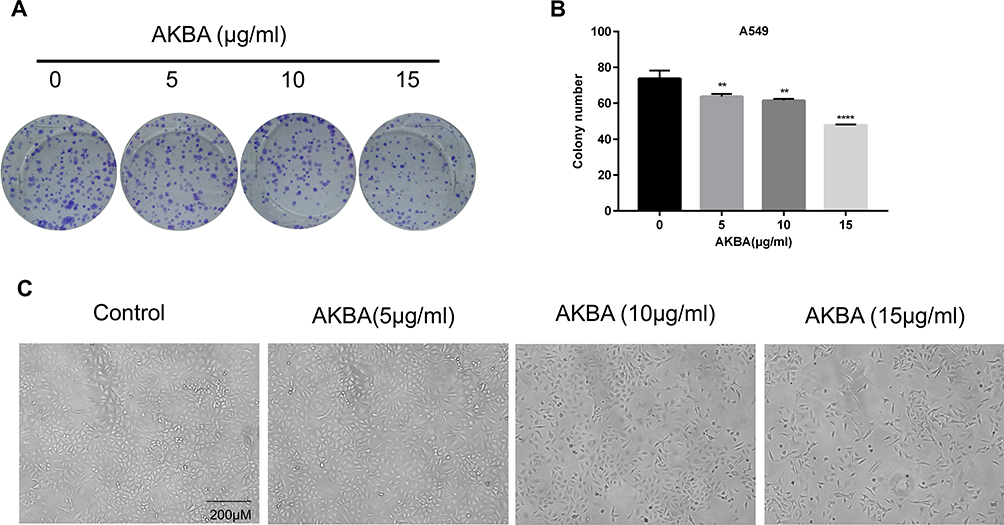 |
Figure 2 Effects of AKBA on the clone formation and the morphology in human NSCLC cell line A549. (A) AKBA inhibited the clone formation of A549 in a dose-dependent manner. (B) Statistical analysis of clone number in A549. (C) Morphology of A549 cells treated with AKBA at 0, 5, 10, and 15 μg/mL after 48 h. Scale bar=200 μM. The experiments were performed in triplicate, and the data were presented as mean ± SD, **P<0.01, ****P<0.0001 vs the AKBA-untreated control group (0 μg/mL). |
AKBA Induced the Cell Cycle Arrest at G0/G1 Phase in Human NSCLC Cell Line A549
As we all know, cell cycle arrest can lead to proliferation inhibition. In this study, to examine the effects of AKBA on cell cycle, we used flow cytometry to show the distribution of the cell cycle after treatment with AKBA at 0, 5, 10 μg/mL in A549 (Figure 3A). Interestingly, we found that AKBA increased the percent of G0/G1 phase (Figure 3B) and decreased the percent of S phase at 10 μg/mL (Figure 3C), but rarely changed the percent of G2/M (Figure 3D). These results suggested that AKBA suppressed the growth of tumor cells via arresting the cell cycle at the G0/G1 phase.
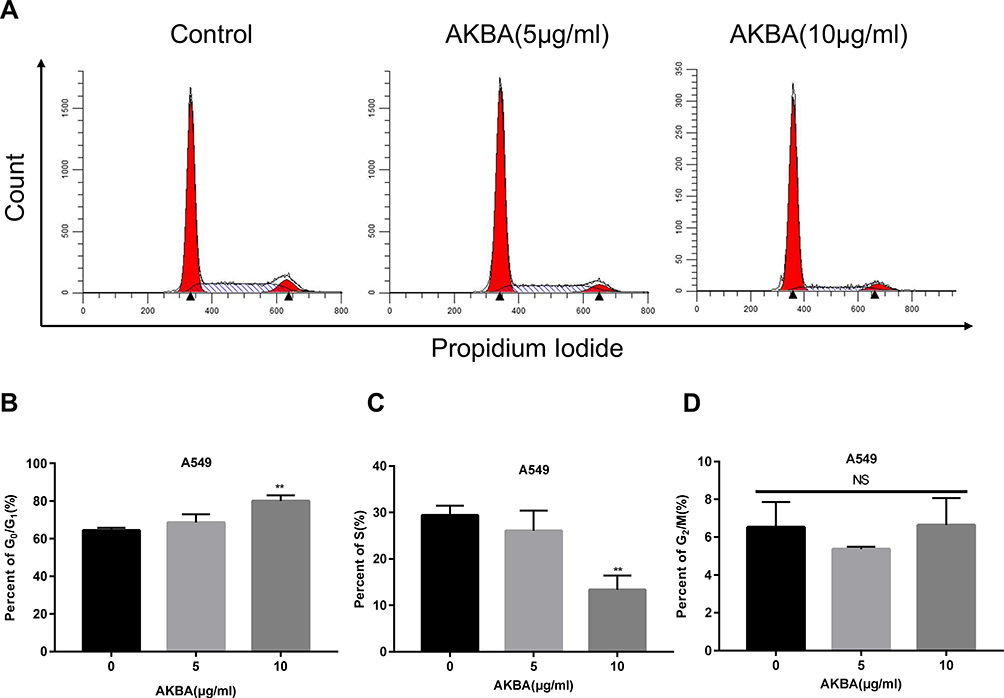 |
Figure 3 Effects of AKBA on the cell cycle in A549. (A) Flow cytometry showed that AKBA arrested the A549 cell cycle in the G0/G1 phase. (B) Statistical analysis of the cell cycle of A549 in the G0/G1 phase. (C) Statistical analysis of the cell cycle of A549 in the S phase. (D) Statistical analysis of the cell cycle of A549 in the G2/M phase. The experiments were performed in triplicate, and the data were presented as mean ± SD, **P<0.01, vs the AKBA-untreated control group (0 μg/mL). |
AKBA Suppressed the Expressions of the Cell Cycle Proteins in Human NSCLC Cell Line A549
Cell cycle arrest is correlated with the cell cycle regulator. To investigate the effects of AKBA on cell cycle regulator, we used Western blotting assay to show the expressions of the cell cycle proteins (Figure 4A). In this study, AKBA downregulated the expression levels of cyclin A2 and cyclin E1 (Figure 4B and C). Especially, the expression of p-cdc2 was significantly decreased after treatment with AKBA (Figure 4D). These results of the Western blotting analysis were in agreement with the results of flow cytometry, further demonstrating that AKBA inhibited the proliferation of A549.
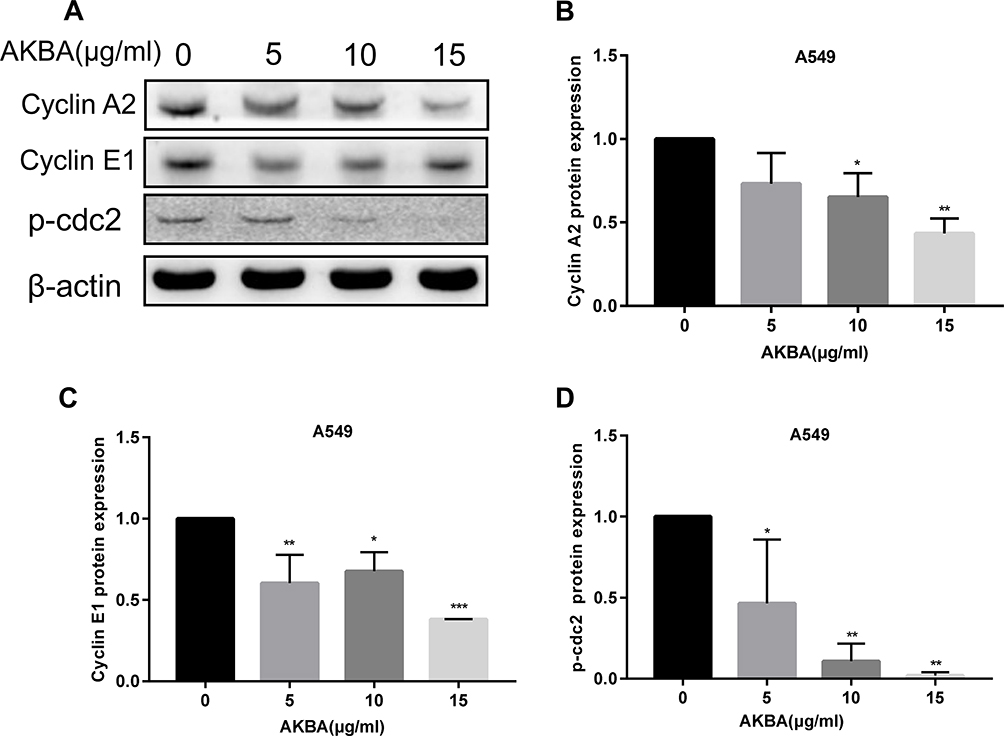 |
Figure 4 Effects of AKBA on cell cycle proteins in A549. (A) Western blotting analysis showed that AKBA decreased the protein expression of the cell cycle in A549 cells. (B) Statistical analysis of cyclin A2 in A549 cells. (C) Statistical analysis of cyclin E1 in A549. (D) Statistical analysis of p-cdc2 in A549 cells. The experiments were performed in triplicate, and the data were presented as mean ± SD, *P<0.05, **P<0.01, and ***P<0.001 vs the AKBA-untreated control group (0 μg/mL). |
AKBA Promoted the mRNA Expressions of p27 and p21 in a Dose-Dependent Manner in Human NSCLC Cell Line A549
P27, as well as p21, is a cyclin-dependent kinase inhibitor (CKI) that negatively regulates cell cycle proteins, thereby affecting the cell cycle process. To further investigate the effects of AKBA on the cell cycle, we used quantitative real-time fluorescence PCR to show the relative mRNA expressions of p27 and p21 in A549. In this study, AKBA remarkedly increased the relative mRNA level of p27 (Figure 5A). Furthermore, we found that the mRNA expression of p21 in A549 was upregulated in a dose-dependent manner (Figure 5B). these results demonstrated that AKBA promoted the mRNA expressions of p27 and p21, thereby suppressing the expressions of cell cycle proteins.
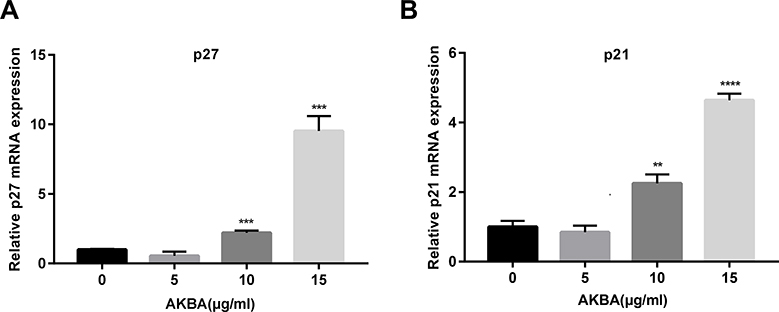 |
Figure 5 Effects of AKBA on the mRNA levels of p27 and p21 in A549. (A) The relative mRNA level of p27. (B) The relative mRNA level of p21. The experiments were performed in triplicate, and the data were presented as mean ± SD, **P<0.01, ***P<0.001, and ****P<0.0001 vs the AKBA-untreated control group (0 μg/mL). |
AKBA Upregulated the Expression Levels of p27 and p21 Protein in Human NSCLC Cell Line A549
According to the previous results about p27 and p21 in mRNA level, we further studied the effects of AKBA on the protein expressions of p27 and p21 by using Western blotting assay (Figure 6A). We found that p27 and p21 protein expression levels were markedly increased after treatment with AKBA at 10 μg/mL (Figure 6B and C). These data proved that AKBA at 10 ug/mL upregulated the protein expressions of p27 and p21 in A549. By analyzing the previous results about p27 and p21, we found that the effects of AKBA on p27 and p21 in protein and mRNA levels were not exactly the same, suggesting that AKBA at 10 μg/mL could exert more comprehensive effects.
 |
Figure 6 Effects of AKBA on the protein levels of p27 and p21 in A549. (A) Western blotting analysis showed that AKBA increased the expression levels of p27 and p21 protein. (B) Statistical analysis of p27 protein. (C) Statistical analysis of p21 protein. The experiments were performed in triplicate, and the data were presented as mean ± SD, *P<0.05, **P<0.01 vs the AKBA-untreated control group (0 μg/mL). |
AKBA Induced the Cellular Apoptosis in Human NSCLC Cell Line A549
Considering the previous results about the effects of A549 on proliferation and cell cycle, we employed flow cytometry to demonstrate the impact on cellular apoptosis in A549 (Figure 7A). Compared with the AKBA-untreated control group, the percentages of apoptotic cells were notably aggrandized after treatment with AKBA at 10 μg/mL (Figure 7B). Moreover, the Western blotting analysis showed that the relevant apoptotic proteins Bcl-xl and Bax were changed after AKBA treatment (Figure 7C). The expression of the negative regulatory protein of apoptosis Bcl-xl was obviously downregulated after treatment with AKBA in a dose-dependent manner, but the expression of the positive regulatory protein of Bax was slightly increased after AKBA treatment (Figure 7D and E). Therefore, these data indicated that AKBA induced cellular apoptosis in NSCLC cell line A549, which suggested that AKBA might be an effective drug to treat NSCLC by promoting cancer cell apoptosis.
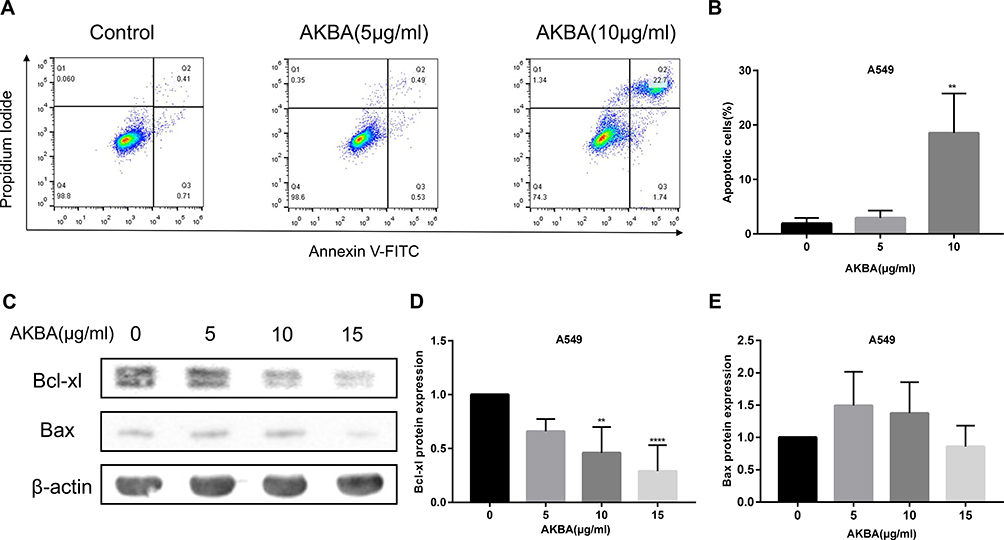 |
Figure 7 AKBA induced cellular apoptosis in A549. (A) Apoptosis was determined by flow cytometry with FITC/PI gating strategy in A549. (B) Statistical analysis of the percentages of apoptotic cells in A549. (C) The protein expressions of Bcl-xl and Bax by Western blotting in A549. (D) Statistical analysis of Bcl-xl in A549. (E) Statistical analysis of Bax in A549. The experiments were performed in triplicate, and the data were presented as mean ± SD, **P<0.01, ****P<0.0001 vs the AKBA-untreated control group (0 μg/mL). |
AKBA Suppressed the Protein Expressions of Autophagy in Human NSCLC Cell Line A549
Autophagy is commonly considered a survival pathway, and inhibiting autophagy in cancer cells might be an effective method of suppressing the proliferation of cancer cells, which might be a method of treating cancer. The autophagy proteins Beclin-1, LC3A/B-I, and LC3A/B-II were detected by Western blotting assay (Figure 8A). Statistical analysis results showed that the protein expressions of Beclin-1 and LC3A/B-II were obviously decreased after treatment with AKBA at 48 h (Figure 8B and C). In addition, NF- ƙB protein as a protein that can affect autophagy, in this study, it and its phosphorylated protein were determined by Western blotting assay (Figure 8D). We also found that AKBA led to the downregulation of NF-ƙB p65 and p- NF-ƙB p65 (Figure 8E and F). The data of this part revealed that AKBA exerted the anti-cancer effects due to the suppression of autophagy in A549.
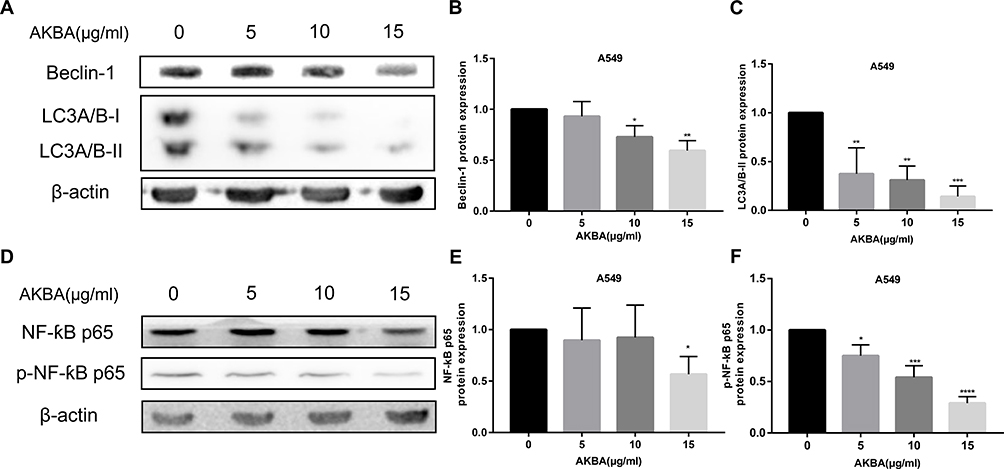 |
Figure 8 Effects of AKBA on autophagy proteins in A549. (A) The protein expressions of Beclin-1, LC3A/B-I, and LC3A/B-II by Western blotting in A549. (B) Statistical analysis of Beclin-1 in A549. (C) Statistical analysis of LC3A/B-II in A549. (D) The protein expressions of NF-ƙB p65, and p- NF-ƙB p65 by Western blotting in A549. (E) Statistical analysis of NF-ƙB p65 in A549. (F) Statistical analysis of p- NF-ƙB p65 in A549. The experiments were performed in triplicate, and the data were presented as mean ± SD, *P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001 vs the AKBA-untreated control group (0 μg/mL). |
AKBA Inhibited the Formation of Autolysosome in Human NSCLC Cell Line A549
To further explore the effects of AKBA on autophagy, we examined the autolysosome by immunofluorescence in A549. In the control group (AKBA at 0 μg/mL), we found that a mass of autolysosomes was developed by rapamycin (autophagy inducer) (Figure 9A). Interestingly, the number of autolysosomes was markedly decreased after treatment with AKBA at 10 μg/mL (Figure 9B), indicating that AKBA suppressed autophagy by decreasing the formation of autolysosome.
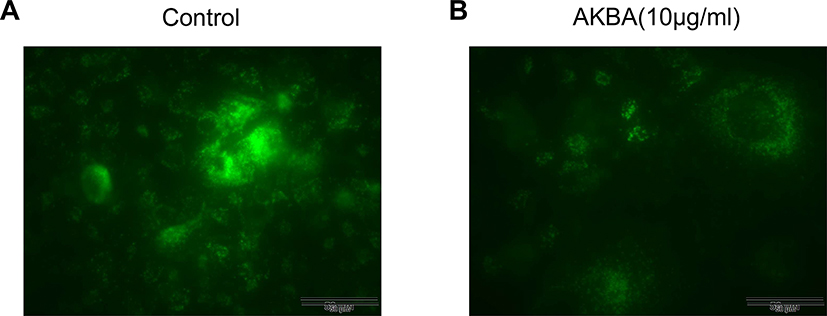 |
Figure 9 Effects of AKBA on autolysosome by immunofluorescence in A549. (A) The formation of autolysosome in the control group. (B) AKBA at 10 μg/mL suppressed the formation of autolysosome in A549. Scale bar=50 μM. |
AKBA Regulated PI3K/Akt Signaling Pathway in Human NSCLC Cell Line A549
PI3K/Akt has been reported to be a key signaling pathway, having an important impact on cell proliferation and apoptosis. In this study, the PI3K and p-PI3K proteins were tested by Western blotting assay (Figure 10A). We found that AKBA downregulated the protein expression levels of PI3K and p-PI3K (Figure 10B and C). Interestingly, the effects of AKBA on p-PI3K protein at 10 ug/mL were more significant than at other concentrations. Akt is the downstream protein of PI3K protein. We used Western blotting assay to detect the expressions of Akt and p-Akt (Figure 10D), finding that AKBA decreased the expression of Akt and p-Akt (Figure 10E and F). In summary, AKBA regulated PI3K/Akt signaling pathway by resulting in the expression inhibition of PI3K, p-PI3K, Akt, and p-Akt in A549.
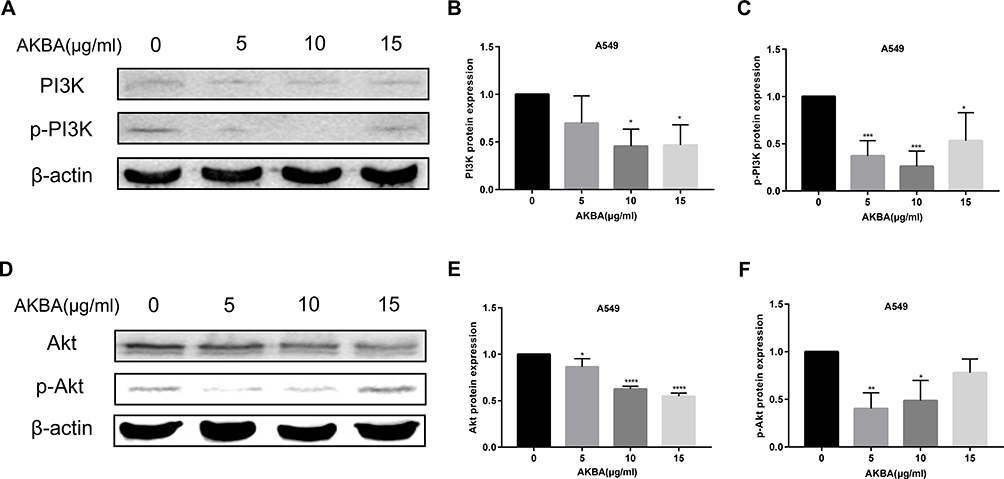 |
Figure 10 AKBA regulated PI3K/Akt signaling pathway in A549 cells. (A) The protein expressions of PI3K and p-PI3K by Western blotting. (B) Statistical analysis of PI3K. (C) Statistical analysis of p-PI3K. (D) The protein expressions of Akt and p-Akt by Western blotting. (E) Statistical analysis of Akt. (F) Statistical analysis of p-Akt. The experiments were performed in triplicate, and the data were presented as mean ± SD, *P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001 vs the AKBA-untreated control group (0 μg/mL). |
Discussion
The identification of effective anti-cancer drugs from plant-derived natural products play an important role in cancer treatment. AKBA has recently gained considerable attention because of its potential antitumor activity. By preliminary investigation, we found that AKBA can reduce the viability of A549, H1299, and H460. Therefore, we attempted to elucidate the anti-tumor effects of AKBA on NSCLC cell lines and the possible mechanisms of action in this study.
Cell proliferation is defined by several genetically defined checkpoints to ensure its coordinated progression through different stages of the cell cycle and to monitor DNA integrity.15 Cell cycle arrest in stress response is the key to maintaining genomic integrity. The regulatory mechanism of inhibiting cell cycle transformation or inducing the apoptosis signaling pathway after cell stress is called cell cycle checkpoint.16 In this study, we used flow cytometry to investigate cycle distribution. The data showed that the percentages of G0/G1 were increased in a dose-dependent manner, suggesting that AKBA arrested the cell cycle in the G0/G1 phase. Cell cycle progression is highly regulated by a series of cell cycle checkpoint proteins, such as the cyclins, CDKs, and CKI. Among these proteins, cyclin A and E, together with CDK2, play major roles in DNA replication and mitosis by regulating the G0/G1 phase of the cell cycle.17 p27 and p21, two types of cyclin-dependent kinase inhibitors, can inhibit the combination of cyclin E or cyclin A with CDK2.18,19 Our results showed that AKBA suppressed the protein expression levels of cyclin E and cyclin A by Western blotting analysis and upregulated the relative mRNA levels of p27 and p21 by using quantitative real-time fluorescence PCR, revealing the particular mechanism by which AKBA regulated the cell cycle in NSCLC cell line A549. These results demonstrated that AKBA exerted the anti-tumor effects via inducing cell cycle arrest.
Apoptosis, also known as programmed cell death, and cell cycle arrest are two important events involved in anticancer drug treatment. Apoptosis, easily triggered by many commonly used chemopreventive agents,20 is the result of a highly complex series of events involving cell chromatin condensation, DNA fragmentation, and cell shrinkage,21 including two major signaling pathways controlling apoptosis, the extrinsic apoptosis pathway (death receptor) and the intrinsic apoptosis pathway (mitochondrial-dependent).22,23 Mitochondria play a crucial role in apoptotic regulation and the mitochondrial dysfunction that occurs during apoptosis.24 In this study, the results of flow cytometry demonstrated that AKBA increased the percentage of apoptotic cells. We used Western blotting analysis to show that AKBA decreased the expression levels of anti-apoptotic protein Bcl-xl and upregulated pro-apoptotic protein Bax in A549, indicating that AKBA may induce the mitochondrial dysfunction caused during apoptosis. It has been reported that the fast Bax retrotranslocation to the cytosol mediated by Bcl-xl avoids Bax accumulation at the mitochondria and ensures cell survival,25 which is in agreement with our results regarding apoptosis. Hence, AKBA could exert anti-cancer role by increasing the cellular apoptosis in NSCLC cell line A549.
Autophagy is a cellular process whereby cells eliminate misfolded intracellular proteins and damaged organelles through lysosomal degradation to recycle their nutrients.26 Emerging evidence indicates that autophagy is a self-protective cellular mechanism that provides energy through the degradation and recycling of cytoplasmic contents and promotes cell survival in response to a variety of stimuli.27 In a cancer cell, autophagy also acted as a survival mechanism to resist chemotherapeutic agents induced apoptosis.28 Hence, seeking new effective agents that can suppress autophagy in tumors may be a promising approach against cancer. Our results of Western blotting analysis demonstrated that AKBA might inhibit autophagy in NSCLC cell line A549 because the expression levels of Beclin-1, LC3A/B-I, and LC3A/B-II proteins were significantly decreased after treatment with AKBA. Recent studies showed that the expressions of autophagy-related proteins under oxidative stress were dependent on NF-κB p65 Ser536 phosphorylation.29 Otherwise, in a tumor cell, ANGPTL8 negatively regulates NF-κB activation by facilitating selective autophagic degradation, suggesting that autophagy inhibition is related to NF-κB inactivation.30 In this study, we found that AKBA suppressed the expression of NF-κB p65 and p- NF-κB p65, and led to the downregulation of the expression of autophagy-related proteins. Therefore, we predicted that AKBA could be a promising drug against autophagy, which deserved further investigation.
The PI3K/Akt pathway is one of the major signaling pathways regarding the molecular mechanisms by which this network regulates the activities of the cell cycle, cellular proliferation, growth, survival, apoptosis, drug resistance, protein synthesis, and glucose metabolism.31,32 Inhibiting the PI3K/Akt pathway could target cancer cell biology, suppress cell growth, and induce cell apoptosis.33 Akt is a key downstream effector of PI3K, and p-Akt, a phosphorylated protein of Akt, is downregulated in various cancers, including lung cancer34 and gastric cancer.35 PI3K phosphorylates Akt, and then Akt can further activate a variety of downstream targets. Our results demonstrated that the expressions of Akt and p-Akt were decreased after treatment of AKBA, and the upstream regulation proteins PI3K and p-PI3K were also suppressed by Western blotting analysis, indicating that the PI3K/Akt signaling was involved. The activation of PI3K/Akt signaling downregulated the p27 and p21 and upregulated cyclin E, cyclin A, or cyclin D, thereby contributing to promoting cell cycle progression.36–38 The previous results in this study have shown upregulation of p21 and p27, downregulation of cyclin E and cyclin A after treatment of AKBA for 48 h. In addition, the percentages of the G0/G1 phase were significantly increased after treatment with AKBA. Therefore, we predicted that AKBA exerted anti-cancer effects through inducing cell cycle arrest via possibly inactivating the PI3K/Akt pathway. Recently, many studies have shown that the PI3K/Akt signaling pathway plays a crucial role in inducing cell apoptosis,39,40 and suppression of PI3K/Akt signaling was involved in Bax -mediated apoptosis.41,42 In our study, flow cytometry assay showed AKBA increased the percentages of apoptotic cells in NSCLC cell line A549, and the results of Western blotting assay showed upregulation of Bax and downregulation of Bcl-xl, indicating that AKBA exerted the anti-cancer effects by inducing cell apoptosis via inhibition of PI3K/Akt pathway. However, these studies have not been conducted in animal experiments. Additionally, we need to add the activating agent of the PI3K/Akt pathway to further study the effects of AKBA on biological function in NSCLC cell lines.
Conclusion
AKBA exerts the anti-cancer effects via cell cycle arrest, apoptosis induction and autophagy suppression in non-small cell lung cancer cells. This body of evidence supports the potential of AKBA as a promising drug in the treatment of NSCLC.
Acknowledgments
This work was supported by the Jinshan District Health Administrative Authority (Grant Number: JSZK2015A04‐993) and the Jinshan Science and Technology Commission (Grant Number: 2018-3-5).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.21387
2. Gridelli C, Rossi A, Carbone DP, et al. Non-small-cell lung cancer. Nat Rev Dis Primers. 2015;1:15009. doi:10.1038/nrdp.2015.9
3. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.v68.6
4. Reck M, Heigener DF, Mok T, Soria JC, Rabe KF. Management of non-small-cell lung cancer: recent developments. Lancet. 2013;382(9893):709–719. doi:10.1016/S0140-6736(13)61502-0
5. Liao YH, Li CI, Lin CC, Lin JG, Chiang JH, Li TC. Traditional Chinese medicine as adjunctive therapy improves the long-term survival of lung cancer patients. J Cancer Res Clin Oncol. 2017;143(12):2425–2435. doi:10.1007/s00432-017-2491-6
6. Zhang YS, Shen Q, Li J. Traditional Chinese medicine targeting apoptotic mechanisms for esophageal cancer therapy. Acta Pharmacol Sin. 2016;37(3):295–302. doi:10.1038/aps.2015.116
7. Liao YH, Lin CC, Lai HC, Chiang JH, Lin JG, Li TC. Adjunctive traditional Chinese medicine therapy improves survival of liver cancer patients. Liver Int. 2015;35(12):2595–2602. doi:10.1111/liv.12847
8. Chen S, Flower A, Ritchie A, et al. Oral Chinese herbal medicine (CHM) as an adjuvant treatment during chemotherapy for non-small cell lung cancer: A systematic review. Lung Cancer. 2010;68(2):137–145. doi:10.1016/j.lungcan.2009.11.008
9. Shah BA, Qazi GN, Taneja SC. Boswellic acids: a group of medicinally important compounds. Nat Prod Rep. 2009;26(1):72–89. doi:10.1039/B809437N
10. Syrovets T, Buchele B, Krauss C, Laumonnier Y, Simmet T. Acetyl-boswellic acids inhibit lipopolysaccharide-mediated TNF-alpha induction in monocytes by direct interaction with IkappaB kinases. J Immunol. 2005;174(1):498–506. doi:10.4049/jimmunol.174.1.498
11. Barbarisi M, Barbarisi A, De Sena G, et al. Boswellic acid has anti-inflammatory effects and enhances the anticancer activities of Temozolomide and Afatinib, an irreversible ErbB family blocker, in human glioblastoma cells. Phytother Res. 2019;33(6):1670–1682. doi:10.1002/ptr.v33.6
12. Takada Y, Ichikawa H, Badmaev V, Aggarwal BB. Acetyl-11-keto-beta-boswellic acid potentiates apoptosis, inhibits invasion, and abolishes osteoclastogenesis by suppressing NF-kappa B and NF-kappa B-regulated gene expression. J Immunol. 2006;176(5):3127–3140. doi:10.4049/jimmunol.176.5.3127
13. Poeckel D, Werz O. Boswellic acids: biological actions and molecular targets. Curr Med Chem. 2006;13(28):3359–3369. doi:10.2174/092986706779010333
14. Bishnoi M, Patil CS, Kumar A, Kulkarni SK. Potentiation of antinociceptive effect of NSAIDs by a specific lipooxygenase inhibitor, acetyl 11-keto-beta boswellic acid. Indian J Exp Biol. 2006;44(2):128–132.
15. Brooks G, La Thangue NB. The cell cycle and drug discovery: the promise and the hope. Drug Discov Today. 1999;4(10):455–464. doi:10.1016/S1359-6446(99)01400-2
16. Flatt PM, Pietenpol JA. Mechanisms of cell-cycle checkpoints: at the crossroads of carcinogenesis and drug discovery. Drug Metab Rev. 2000;32(3–4):283–305. doi:10.1081/DMR-100102335
17. Vermeulen K, Berneman ZN, Van Bockstaele DR. Cell cycle and apoptosis. Cell Prolif. 2003;36(3):165–175. doi:10.1046/j.1365-2184.2003.00267.x
18. Chu IM, Hengst L, Slingerland JM. The Cdk inhibitor p27 in human cancer: prognostic potential and relevance to anticancer therapy. Nat Rev Cancer. 2008;8(4):253–267. doi:10.1038/nrc2347
19. Abbas T, Dutta A. p21 in cancer: intricate networks and multiple activities. Nat Rev Cancer. 2009;9(6):400–414. doi:10.1038/nrc2657
20. Jeyamohan S, Moorthy RK, Kannan MK, Arockiam AJ. Parthenolide induces apoptosis and autophagy through the suppression of PI3K/Akt signaling pathway in cervical cancer. Biotechnol Lett. 2016;38(8):1251–1260. doi:10.1007/s10529-016-2102-7
21. Debatin KM, Krammer PH. Death receptors in chemotherapy and cancer. Oncogene. 2004;23(16):2950–2966. doi:10.1038/sj.onc.1207558
22. Forbes-Hernandez TY, Giampieri F, Gasparrini M, et al. The effects of bioactive compounds from plant foods on mitochondrial function: a focus on apoptotic mechanisms. Food Chem Toxicol. 2014;68:154–182. doi:10.1016/j.fct.2014.03.017
23. Jin Z, El-Deiry WS. Overview of cell death signaling pathways. Cancer Biol Ther. 2005;4(2):139–163. doi:10.4161/cbt.4.2.1508
24. Hu W, Shen T, Wang MH. Cell cycle arrest and apoptosis induced by methyl 3,5-dicaffeoyl quinate in human colon cancer cells: involvement of the PI3K/Akt and MAP kinase pathways. Chem Biol Interact. 2011;194(1):48–57. doi:10.1016/j.cbi.2011.08.006
25. Pena-Blanco A, Garcia-Saez AJ. Bax, Bak and beyond - mitochondrial performance in apoptosis. FEBS J. 2018;285(3):416–431. doi:10.1111/febs.14186
26. He C, Klionsky DJ. Regulation mechanisms and signaling pathways of autophagy. Annu Rev Genet. 2009;43:67–93. doi:10.1146/annurev-genet-102808-114910
27. Lum JJ, Bauer DE, Kong M, et al. Growth factor regulation of autophagy and cell survival in the absence of apoptosis. Cell. 2005;120(2):237–248. doi:10.1016/j.cell.2004.11.046
28. Ozpolat B, Benbrook DM. Targeting autophagy in cancer management - strategies and developments. Cancer Manag Res. 2015;7:291–299. doi:10.2147/CMAR.S34859
29. Song C, Mitter SK, Qi X, et al. Oxidative stress-mediated NFkappaB phosphorylation upregulates p62/SQSTM1 and promotes retinal pigmented epithelial cell survival through increased autophagy. PLoS One. 2017;12(2):e171940.
30. Zhang Y, Guo X, Yan W, et al. ANGPTL8 negatively regulates NF-kappaB activation by facilitating selective autophagic degradation of IKKgamma. Nat Commun. 2017;8(1):2164. doi:10.1038/s41467-017-02355-w
31. O’Donnell JS, Massi D, Teng M, Mandala M. PI3K-AKT-mTOR inhibition in cancer immunotherapy, redux. Semin Cancer Biol. 2018;48:91–103. doi:10.1016/j.semcancer.2017.04.015
32. Mayer IA, Arteaga CL. The PI3K/AKT pathway as a target for cancer treatment. Annu Rev Med. 2016;67:11–28. doi:10.1146/annurev-med-062913-051343
33. Xie Q, Wen H, Zhang Q, et al. Inhibiting PI3K-AKt signaling pathway is involved in antitumor effects of ginsenoside Rg3 in lung cancer cell. Biomed Pharmacother. 2017;85:16–21. doi:10.1016/j.biopha.2016.11.096
34. Tripathi SC, Fahrmann JF, Celiktas M, et al. MCAM mediates chemoresistance in small-cell lung cancer via the PI3K/AKT/SOX2 signaling pathway. Cancer Res. 2017;77(16):4414–4425. doi:10.1158/0008-5472.CAN-16-2874
35. Luo Y, Zha L, Luo L, et al. [6]-Gingerol enhances the cisplatin sensitivity of gastric cancer cells through inhibition of proliferation and invasion via PI3K/AKT signaling pathway. Phytother Res. 2019;33(5):1353–1362. doi:10.1002/ptr.v33.5
36. Prasad SB, Yadav SS, Das M, et al. PI3K/AKT pathway-mediated regulation of p27(Kip1) is associated with cell cycle arrest and apoptosis in cervical cancer. Cell Oncol (Dordr). 2015;38(3):215–225. doi:10.1007/s13402-015-0224-x
37. Li D, Dai C, Yang X, et al. Critical role of p21 on olaquindox-induced mitochondrial apoptosis and S-phase arrest involves activation of PI3K/AKT and inhibition of Nrf2/HO-1pathway. Food Chem Toxicol. 2017;108(Pt A):148–160. doi:10.1016/j.fct.2017.07.054
38. Zheng L, Zhang Y, Liu Y, et al. MiR-106b induces cell radioresistance via the PTEN/PI3K/AKT pathways and p21 in colorectal cancer. J Transl Med. 2015;13:252. doi:10.1186/s12967-015-0592-z
39. Granato M, Rizzello C, Gilardini MM, et al. Quercetin induces apoptosis and autophagy in primary effusion lymphoma cells by inhibiting PI3K/AKT/mTOR and STAT3 signaling pathways. J Nutr Biochem. 2017;41:124–136. doi:10.1016/j.jnutbio.2016.12.011
40. Yang J, Pi C, Wang G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed Pharmacother. 2018;103:699–707. doi:10.1016/j.biopha.2018.04.072
41. Lin ML, Chen SS, Huang RY, et al. Suppression of PI3K/Akt signaling by synthetic bichalcone analog TSWU-CD4 induces ER stress- and Bax/Bak-mediated apoptosis of cancer cells. Apoptosis. 2014;19(11):1637–1653. doi:10.1007/s10495-014-1031-y
42. Lu X, Masic A, Li Y, Shin Y, Liu Q, Zhou Y. The PI3K/Akt pathway inhibits influenza A virus-induced Bax-mediated apoptosis by negatively regulating the JNK pathway via ASK1. J Gen Virol. 2010;91(Pt 6):1439–1449. doi:10.1099/vir.0.018465-0
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.