")
Back to Journals » Drug Design, Development and Therapy » Volume 14
A Synthetic Peptide 2Abz23S29 Reduces Bacterial Titer and Induces Pro-Inflammatory Cytokines in a Murine Model of Urinary Tract Infection
Authors Moazzezy N , Asadi Karam MR, Rafati S , Bouzari S , Oloomi M
Received 25 April 2020
Accepted for publication 6 July 2020
Published 17 July 2020 Volume 2020:14 Pages 2797—2807
DOI https://doi.org/10.2147/DDDT.S259937
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Georgios Panos
Neda Moazzezy,1 Mohammad Reza Asadi Karam,1 Sima Rafati,2 Saeid Bouzari,1 Mana Oloomi1
1Molecular Biology Department, Pasteur Institute of Iran, Tehran, Iran; 2Immunotherapy and Leishmania Vaccine Research Department, Pasteur Institute of Iran, Tehran, Iran
Correspondence: Saeid Bouzari; Mana Oloomi Email [email protected]; [email protected]
Introduction: A urinary tract infection (UTI), which is often caused by uropathogenic E. coli (UPEC) strains, affects many people worldwide annually. UPEC causes the production of pro-inflammatory cytokines by the bladder epithelial cells; however, it has been proven that the UPEC can inhibit the early activation of the innate immune system.
Methods: This study aimed to examine the antibacterial and immunomodulatory effects of different doses of truncated alpha-defensins (human neutrophil peptide (HNP)-1) analog 2Abz23S29 on the mouse UTI model. Experimentally uropathogenic E. coli CFT073-infected mice were treated with low-dose 2Abz23S29 (250μg/mL), high-dose 2Abz23S29 (750μg/mL), ciprofloxacin (cip) (800μg/mL), or high-dose 2Abz23S29plus cip once a day 24 h post-infection. The 2Abz23S29 and cip treatment were given for two consecutive days.
Results: The in vivo results showed that fewer UPEC were recovered from the bladders of mice treated transurethrally with 2Abz23S29. Moreover, low-dose 2Abz23S29 significantly decreased the level of the interleukin-6 (IL-6), whereas high-dose 2Abz23S29 increased pro-inflammatory cytokines including IL-6, macrophage inflammatory protein/2 (MIP/2), tumor necrosis factor-α (TNF-α), and interleukin-1β (IL-1β) in infected bladders of mice. Besides, the levels of cytokines IL-6 and MIP/2 in infected mice treated with a combination of high-dose 2Abz23S29 and cip were significantly higher than the untreated mice. In contrast, CFT073-infected mice treated with a combination of high-dose 2Abz23S29 and cip showed no changes in cytokines TNF-α and IL-1β levels, indicating that ciprofloxacin may play an anti-inflammatory role.
Conclusion: Collectively, apart from the direct antibacterial role of 2Abz23S29, our data illustrated that 2Abz23S29 modulates pro-inflammatory cytokine production of bladder in a dose-dependent manner, which has implications for the development of new anti-infective agents.
Keywords: urinary tract infection, pro-inflammatory cytokines, human neutrophil peptide (HNP)-1
Introduction
One of the most prevalent sites of bacterial infection in humans is the urinary tract. It has been estimated that nearly half of women and 12% of men will experience at least once urinary tract infection (UTI) during their lifetimes that a quarter of these people are at risk of becoming infected again. The establishment of ascending colonization of uropathogenic E. coli (UPEC) in the bladder or kidneys is the most common causative agent of UTIs.1–4 Colonization and invasion of UPEC in the bladder lead to produce pro-inflammatory cytokines and chemokines, which can enhance inflammatory responses and have a principal role in the first line of defense against microbial invasion. These cytokines are important for the clearance of UPEC and the recruitment of neutrophils.5 Human C-X-C ligand 8 (IL-8) is the major chemokine for neutrophil recruitment during UTI, which is secreted by the urothelial cells.6 The level of IL-8 was also raised in the urine of humans suffering from acute UTI. In UPEC infected mice, the functional mouse homologous of IL-8, macrophage inflammatory protein (MIP)-2, can be found in the urine, bladders, and kidneys.7 In addition to chemokines, levels of cytokines IL-1β, IL-6, and tumor necrosis factor-alpha (TNF-α) also were upregulated in UTI-89 infected C57BL/6 mice bladders.8,9 Antibiotics are the most common treatment for UTI, but their widespread overuse has led to the rapid incidence of antibiotic resistance, which suggests that novel alternative drug agents should be replaced to control bacterial infection. Antimicrobial peptides (AMPs) are produced by a wide variety of organisms and are among the essential components of the innate immune system. AMPs have a broad-spectrum effect against Gram-negative and Gram-positive bacteria, viruses, and fungi. Host defense antimicrobial peptides (HDPs) have immunomodulatory and adjuvant actions. HDPs act as chemotactic factors on immune cells to secret cytokines and chemokines.10 The critical point is that HDPs have been exposed to pathogens for over millions of years, but they still retain their antibacterial properties. This may be because bacterial resistance mechanisms can hardly activate against AMPs.11 Among the AMPs, Human Neutrophil Peptide 1 (HNP-1, also known as α-defensin) has been shown to possess direct antibacterial effects and immunomodulatory properties, including chemotaxis, phagocytosis, and cytokine induction depending on the used concentration.11–15 2Abz23S29 peptide is truncated HNP-1 analog lacking disulfide bridges. This short peptide was modeled on the C-terminal region of HNP-1 (R15-C30). 2Abz23S29 has bend-forming mimetic 2Abz (2-aminobenzoyl derivative) group substituting G23 and S substituting C29. Substitution with 2Abz was designed to introduce conformational constrain in the β-hairpin bend region of the C-terminal sequence of HNP-1. It has mostly antibacterial properties similar to its original form.16
In the present study, the efficacy and immune-modulatory effects of synthetic 2Abz23S29 peptide were compared with the ciprofloxacin, to treat uropathogenic E. coli CFT073 strain-infected UTI BALB/c mice. The results of our study displayed that 2Abz23S29 peptide is a potential molecule that can trigger the immune response as well as decrease the bacterial load in a dose-dependent manner.
Materials and Methods
Ethics Statement
All mice investigations including keeping, handling and euthanizing were approved by Institutional Animal Care and Research Advisory Committee of Pasteur Institute of Iran with ethical code IR.RII.REC.1398.057, based on the Specific National Ethical Guidelines for Biochemical Research issued in 2005 by the Research and Technology Deputy of Ministry of Health and Medical Education of Iran.
Peptide and Antibiotic
The test peptide 2Abz23S29 with sequences of (N-RYGTC(Acm)IYQ2AbzRLWAFS-C) was synthesized by biomatik (Canada) in trifluoroacetate (TFA) salt, and high-performance chromatography (HPLC) purified to > 96% with water and acetonitrile. 2Abz23S29 was dissolved in distilled water and was stored in dark at −80°C until use. Ciprofloxacin (cip) was also used in this study (Sigma-Aldrich). Stock solution from dry powder was prepared and stored in dark at −20°C until use.
Minimum Inhibitory Concentration (MIC) Assay
The MICs of 2Abz23S29 and ciprofloxacin (cip) were determined by broth microdilution assay, according to a previously reported protocol.17 In general, bacteria were grown overnight in LB for 24 h and were diluted to approximately 5 × 105 colony-forming units (CFU/mL) in Mueller Hinton Broth (MHB, Difco, Becton Dickinson Co., Sparks, MD, USA). Then, 50 µL of the bacterial suspension was added to a 96-well U-bottom polystyrene microplate (Grainer, Germany) containing 50 µL of serially diluted peptides or antibiotics. The microplates were incubated at 37°C for 18 to 24 h. MICs were defined as the lowest concentration of the antimicrobial agent that entirely inhibited visible growth.
Checkerboard Titration Assay
Interactions between ciprofloxacin (cip) and 2Abz23S29 were studied by a broth microdilution checkerboard titration assay. Serial dilutions of ciprofloxacin were tested in the presence of serial dilutions of 2Abz23S29. Then, the suspension of 1 × 106 CFU/mL CFT073 was incubated with antimicrobial agents. The plate was incubated for 24 h at 37◦C. The fractional inhibitory concentration (FIC) index was calculated as follows: FIC of cip=MIC of cip in combination with 2Abz23S29/MIC of cip alone, FIC of 2Abz23S29=MIC of 2Abz23S29 in combination with cip/MIC of 2Abz23S29 alone, and FIC index= FIC of cip + FIC of 2Abz23S29. The obtained value is classified as FIC≤0.5 as synergistic, FIC>0.5–1.0 as an additive, FIC>1.0 but <4.0 as indifferent, and FIC≥4.0 as antagonistic.18
Time-Kill Assay
The time-dependent results of 2Abz23S29 peptide on uropathogenic E. coli CFT073 viability were estimated by measuring the reduction in the numbers of CFU/mL for 3 h. The exponentially growing bacteria CFT073 was adjusted to 5×105 CFU/mL and then was incubated with 2Abz23S29 and ciprofloxacin (cip) at a certain concentration (1 × MIC) in MHB at 37°C in a final volume of 100 µL. Aliquots of 20 μL were removed at various time intervals (0, 30, 60, 90, 120, and 180 min), serially diluted in sterile saline and plated onto LB agar for 18–24 h at 37°C in the presence of 5% CO2. The bacterial growth was monitored by counting viable colonies and was shown by plotting the log10 CFU/mL against incubation time. A growth control group that had not received any treatment was included as a negative control.
In vitro Cytotoxicity Assay
Cell Culture
Human uroepithelial cell line 5637 (ATCC® HTB-9™) was obtained from the National Cell Bank of Iran (NCBI; Tehran, Iran) and maintained in RPMI 1640 (Gibco, Carlsbad, CA) media which was supplemented with 10% heat-inactivated fetal bovine serum (FBS). The cells were incubated at 37°C in a humidified 5% CO2 incubator.
MTT Assay
Effects of 2Abz23S29 peptide on the viability of 5637 human bladder cancer cells was assessed by measuring mitochondrial activity using MTT [3-(4, 3-dimethylthiazole-2-yl)-2, 5-diphenyltetrazolium bromide] assay.19 Briefly, the epithelial cells were seeded at a density of 1 × 105 cells/mL in each well of 96 well plates containing 100 μL of fresh growth medium per well. After 20–24 h of incubation at 37°C and 5% CO2, cells were treated with peptide at different concentrations and incubated for another 24 h. After incubation, each well was added by 20 μL of MTT (5 mg/mL) and incubated for 4 h at 37°C. After removing the medium, 100 μL isopropanol was rapidly added to well for dissolving the formazan crystals. The absorbance was measured in a microplate reader at 570 nm. The toxicity percentage was calculated according to the following formula.
Toxicity%= (1-mean OD Sample/mean OD control) × 100
In vivo Infection Experiments
Female BALB/c mice, 18–20 g between 6 and 8 weeks of age were obtained from the breeding station of Pasteur Institute, Karaj, Iran, and were held on a 12 h light/dark cycle, at a constant temperature at 21°C to 25°C with 40–60% humidity and with free access to water and standard dietary food. Animals were randomly distributed into different treatment groups, and a detailed overview of the animal groups is shown in Figure 1A and B.
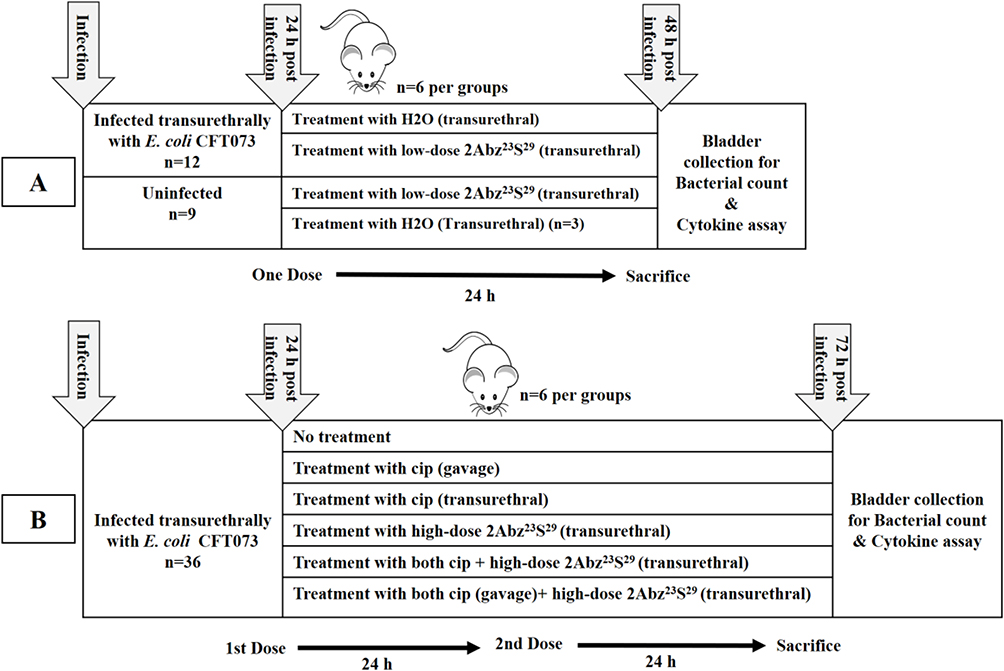 |
Figure 1 Schematic overview of the experimental set up used for in vivo UTI experiment. At the beginning of the experiment (A), mice were subdivided into four groups. Two infected groups and two uninfected groups that received H2O or 2Abz23S29 treatment (n=6 per group, except uninfected group that received H2O n=3). A transurethral injection of 50 µL of H2O or 2Abz23S29 at a low dosage (250µg/mL) was performed after 24 h post-infection, only once. All mice were euthanized on the third day, and bladders were harvested for bacterial count and evaluation of pro-inflammatory cytokines. In the second part of the study (B), mice were randomly subdivided into six groups (n=6 per group), infected untreated group: no treatment, the ciprofloxacin (cip) groups: treated with oral or transurethral cip, the 2Abz23S29group: treated with transurethral 2Abz23S29, and the cip plus 2Abz23S29 groups: treated with oral or transurethral cip and transurethral 2Abz23S29. A single transurethral high-dose of 2Abz23S29and ciprofloxacin was 750 µg/mL, and 40 µg in 50 µL PBS, respectively. For a better comparison between groups, ciprofloxacin was also injected transurethrally. Treatments were performed once a day for two consecutive days. All mice were euthanized on the fourth day, and bladders were harvested for bacterial count and evaluation of pro-inflammatory cytokines. |
The mouse bladder was emptied of urine before the challenge of urinary tract infection (UTI) was performed. Mice were anesthetized with ketamine/xylazine (70 mg/kg + 5 mg/kg) (Alfasan, Netherland) and inoculated transurethrally (administered in ~2–3 sec) with 1 ×107 CFU per mouse suspension of the UPEC CFT073 by using a UV-sterilized polyethylene catheter PE-10 (inner diameter 0.28 mm; outer diameter 0.61 mm) (B&D, USA). The inoculation volume was 50 µL. After inoculation, the mice were kept free of food and water for several hours (4–5 h) to ensure the colonization of bacteria in the bladder, and then they were given water and food again. Alternatively, mice were treated 24 h and 48 h after the bactericidal challenge. Ciprofloxacin (cip) was also given by oral gavage that was accomplished using a 20 gauge-feeding needle 1465LL (Acufirm Ernst Kratz, Dreieich, Germany). In this study, the dose selection of cip (40 mg/kg) as control positive was based on the typical concentration prescribed to human patients, scaled down for treatment in mice.20 At 48 h and 72 h post-infection, all mice were sacrificed to remove bladders, which were homogenized for CFU titration and cytokine assay.
Cytokine ELISAs
Cytokine levels within homogenized bladder supernatants were measured using ELISA 48 h and 72 h post-infection in mice. Levels of mouse cytokines MIP/2, IL-6, 1L-1β, and TNF-α were detected using ELISA kits following the manufacture’s protocol (R&D Systems). Assays were performed in triplicates for each experiment.
Statistical Analysis
All statistics were performed using the nonparametric, Mann–Whitney U-test using Graph Pad Prism 6 (Graph Pad Software, Inc., La Jolla, CA, USA). For all analyses, a P value ˂0.05 was considered statistically significant.
Results
Antimicrobial Activity of the Antimicrobial Agents Alone and Combination
The MIC of 2Abz23S29 and ciprofloxacin against E. coli CFT073 strain were found to be 125 µg/mL and 0.05 µg/mL, respectively. Interaction between 2Abz23S29 and ciprofloxacin was interpreted indifference for E. coli CFT073 strain, with FIC =1.25.
Bacterial Killing Kinetics
2Abz23S29 indicated fast bactericidal activity and was capable of eliminating a great starting inoculum of E. coli CFT073 with approximately 5×105 CFU/mL within 120 mins at 1 × MIC (Figure 2). In contrast to 2Abz23S29, ciprofloxacin displayed slower killing kinetics at 1 × MIC, and complete clearance was not shown after 180 min of exposure.
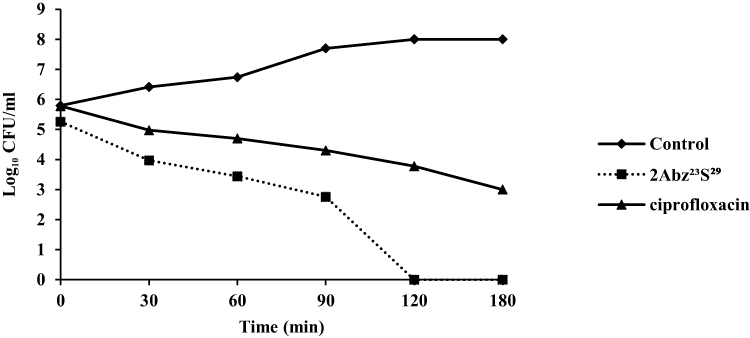 |
Figure 2 Killing kinetic curve of E. coli CFT073 at 1×MIC for ciprofloxacin and 2Abz23S29. Bacteria (5×105 CFU/mL) were cultured in the presence of 2Abz23S29 or ciprofloxacin for various times at 37°C. The untreated bacteria represented as a control. 2Abz23S29 has shown faster bactericidal effect than ciprofloxacin. |
MTT Assay
From Figure 3, it was observed that the test 2Abz23S29 failed to show any cytotoxic effect on human bladder carcinoma cell line 5637 at MIC (125 μg/mL). Based on the toxicity/concentration chart, the half-maximal inhibitory concentration IC50 was determined > 750 μg/mL.
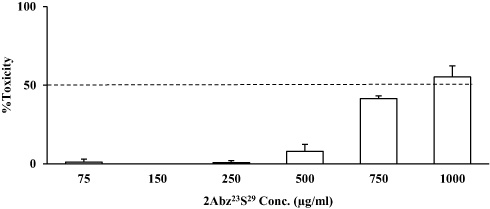 |
Figure 3 Toxicity of 2Abz23S29 in human bladder carcinoma cell line 5637. Cytotoxicity assay representing the mean percentage absorbance at 570 nm after incubation human bladder carcinoma cell line 5637 with different concentrations of 2Abz23S29. Cell viability was measured by MTT assay. All data are displayed as means ± SD from three independent experiments performed in duplicate. |
Efficacy of 2Abz23S29peptide in Mice Model of UTI Infection
The antimicrobial activity of 2Abz23S29 was assessed in a murine model infected with uropathogenic E. coli CFT073 through transurethral injection. In the first step, different concentrations of 2Abz23S29 (below the 50% lethal concentration from 2 × MIC (250 µg/mL) to 6 × MIC (750 µg/mL)) were locally used by transurethral injection 24 h after bacterial challenge to evaluate effectiveness and toxicity. All mice were sacrificed 24 h after treatment, and bladders were homogenized for bacterial enumeration. As a result, the CFU count displayed a significant reduction at all tested concentrations, and no health problems were shown with a transurethral therapeutic route (data not shown).
According to Figure 1A, when mice were treated transurethrally with a low-dose of 2Abz23S29 (250 µg/mL), 24 h post-infection, the mean bacterial bladder load remarkably reduced compared to the H2O treated group, 48 h post-infection (1.46 log reduction, p=0.004) (Figure 4A).
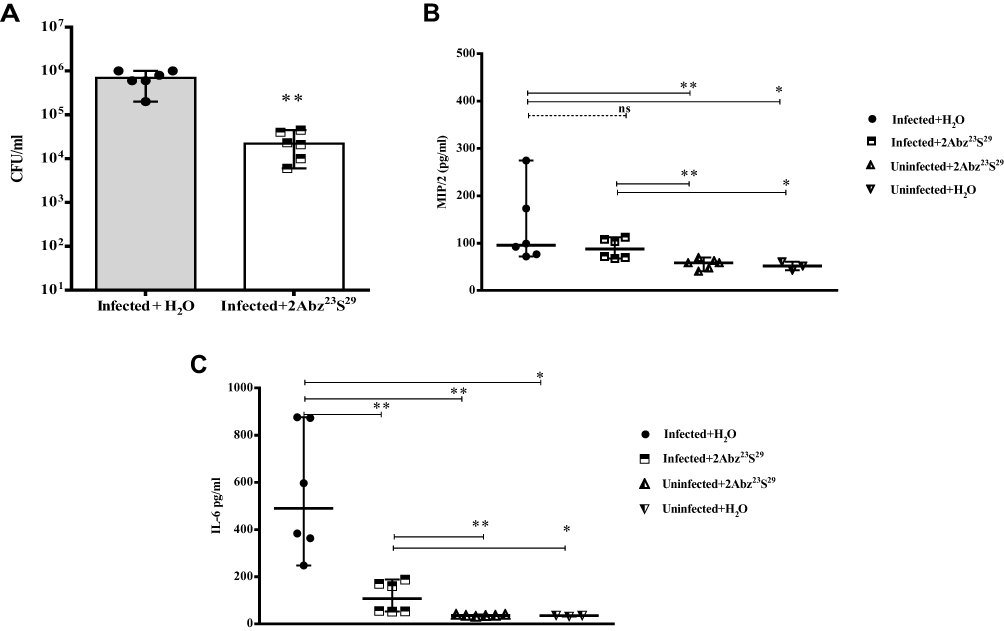 |
Figure 4 Efficacy of low-dose 2Abz23S29 peptide in bacterial load (A) and level of pro-inflammatory cytokines (B and C) in the E. coli-infected mouse model. Female BALB/c mice were infected with 1 × 107 CFU E. coli strain CFT073 by transurethral injection. After 24 h, infected and uninfected mice were treated with single low dose of 2Abz23S29 (250 µg/mL) or H2O administered. The mice were sacrificed, and the bladders were homogenized 48 h post-infection. Bladder homogenate supernatant was examined for bacterial load and ELISA detection of pro-inflammatory cytokine MIP/2 and IL-6. Data are representative of at least two independent experiments (n=6 per group, except H2O treated without challenge group n=3). Bars represent the median values. ns indicates not significant, *p<0.05, **p<0.01, as determined by Mann–Whitney U-test. |
Since cytokines can play a pivotal role in the development and spread of the immune response to bacterial infections, in this study, we also examined the levels of pro-inflammatory cytokines including, MIP/2, IL-6, TNF-α, and IL-1β in different groups of mice. As shown in Figure 4B and C in the groups of infected with CFT073, unlike the MIP/2, the level of IL-6 was significantly decreased in 2Abz23S29 treated group (p=0.004). In contrast, in the uninfected groups, 2Abz23S29 did not significantly alter the production of MIP/2 and IL-6. We also examined pro-inflammatory cytokines TNF-α and IL-1β in infected and uninfected groups 48 post-infection but did not show a significant amount (data not shown). Thus, the 2Abz23S29 at the concentration of 250 µg/mL, in addition to reducing the number of bacteria in the bladder, also affects the immune system by decreasing the level of IL-6 pro-inflammatory cytokine.
Next, according to Figure 1B, the 24 h after CFT073 challenge, we examined the effect of a high-dose of 2Abz23S29 (750 µg/mL) alone or in combination with ciprofloxacin (cip). All treatments were administered once daily for two consecutive days. On the fourth day, the bladder of mice was investigated for bacterial load and pro-inflammatory cytokine levels.
As presented in Figure 5A, all treatments significantly decreased the mean bacterial counts in bladders compared to the infected untreated group (p˂0.01). The infected groups treated with oral cip, transurethral cip, and 2Abz23S29 alone showed 3.47, 2.58, and 3.86 log reduction in bacterial load, respectively. The infected groups treated with both transurethral cip and 2Abz23S29 or oral cip and transurethral 2Abz23S29 generated 3.87 and 4.32 log reduction in bacterial load, respectively. These results indicate that double doses of 750 µg/mL of 2Abz23S29 injected transurethrally can effectively diminish the rate of bacteria in CFT073 bladder infection (more than 99.9% reduction) as the same as double doses of 800 µg/mL of ciprofloxacin. There were not any significant differences between infected treated groups. Even though the combination of 2Abz23S29 and cip reduced the bacterial count more than the cip treated groups, but this difference was not statistically significant.
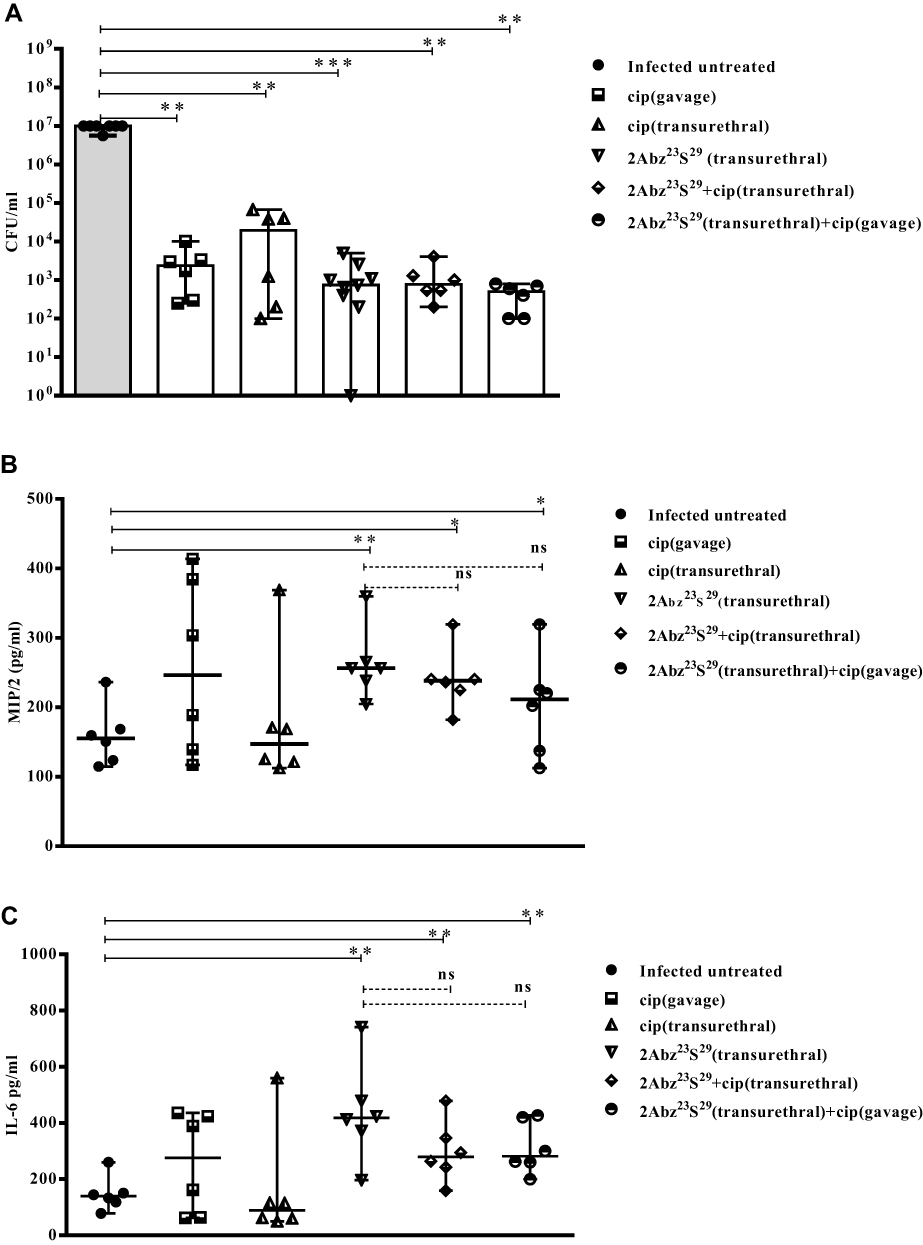 |
Figure 5 Continued. |
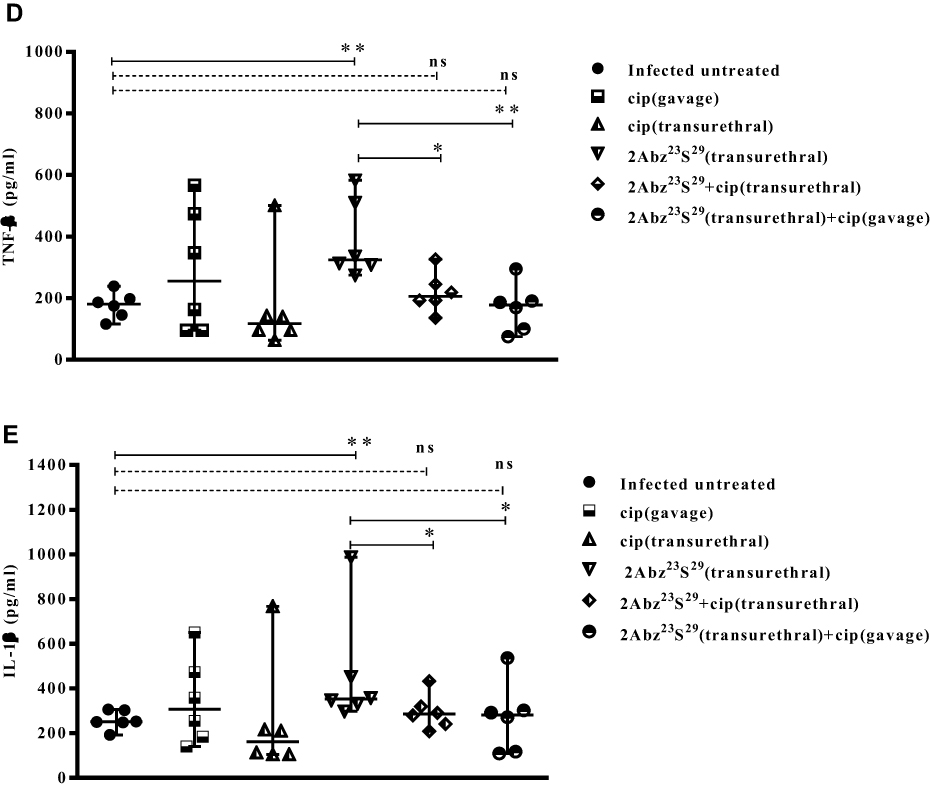 |
Figure 5 Efficacy of high-dose 2Abz23S29 peptide, ciprofloxacin, or a combination of both in bacterial load (A) and level of pro-inflammatory cytokines (B–E) in the E. coli-infected mouse model. Female BALB/c mice were infected with 1 × 107 CFU E. coli strain CFT073 by transurethral injection, and double dose of 2Abz23S29 peptide and/or ciprofloxacin administered after 24 h and 48 h, once a day. The mice were sacrificed, and the bladders were homogenized 72 h post-infection. Bladder homogenate supernatant was examined for bacterial load and ELISA detection of pro-inflammatory cytokine MIP/2, IL-6, TNF-α, and IL-1β. Data are representative of at least two independent experiments (n=6 per group). Bars represent the median values. ns indicates not significant, *p<0.05, **p<0.01, as determined by Mann–Whitney U-test. |
As shown in Figure 5B–E, in the 2Abz23S29 treated group, the level of all pro-inflammatory cytokines (MIP/2, IL-6, TNF-α, and IL-1β) was significantly increased compared to the infected untreated group (p=0.006, 0.006, 0.004, and 0.010, respectively). No statistically significant difference was observed between the level of all pro-inflammatory cytokines (MIP/2, IL-6, TNF-α, and IL-1β) in groups treated with cip alone and the infected untreated mice.
In the groups treated with both 2Abz23S29 and cip, the levels of cytokines MIP/2 and IL-6 were significantly higher than those of the infected untreated group. In contrast, there was not any significant difference between the levels of TNF-α and IL-1β in the groups treated with both 2Abz23S29 and cip and infected untreated group. The 2Abz23S29 treated mice had significantly higher levels of cytokines TNF-α and IL-1β than mice treated with both 2Abz23S29 and cip, while no statistically significant difference was observed in the levels of MIP/2 and IL-6 between 2Abz23S29 treated group and mice treated with both 2Abz23S29 and cip groups.
Discussion
In recent years, the rapid increase in antibiotic resistance as the most common treatment for urinary tract infections (UTIs) has led researchers to pay particular attention to cationic human antimicrobial peptides (AMPs) and their derivatives. Functions of human neutrophil antimicrobial peptides HNPs as strong immune-modulatory molecules may vary based on the local inflammatory conditions, which is strictly connected with their concentration.21 In the present study, the use of low-dose 2Abz23S29 (250 µg/mL) markedly reduced bacterial load in the bladder, and it also significantly decreased the level of pro-inflammatory cytokine IL-6. The decrease in bacterial levels and the peptide’s anti-inflammatory effect at this concentration may indicate a direct antibacterial effect and indirect immune-modulating activity of the peptide. This shows that low-dose 2Abz23S29 may reduce the amount of IL-6 cytokine by producing anti-inflammatory cytokines or decreasing pro-inflammatory cytokine synthesis. In support of this, Miles et al reported that systemic administration of HNP-1 protects peritonitis mice and plays a translation-based anti-inflammatory role in macrophages.22 HNP-1 may impact the translation of pro-inflammatory cytokines in two ways, an effect on mRNA stability or inhibition of the pro-inflammatory transcription factor NF-κB. It was previously shown that intraperitoneal injection of low-dose HNP-1 (5 µg/day) ameliorated intestinal inflammation via diminished colonic expression of pro-inflammatory cytokines IL-6 and IL-1β in BALB/c mice dextran sulfate sodium (DSS)-induced colitis.23 In another study, HNP-1 showed an anti-inflammatory effect and attenuated IL-6 cytokine responses to recombinant hemagglutinin B (rHagB) from Porphyromonas gingivalis.24 A study using UTI-89-infected C57BL/6 mice displayed that the levels of IL-6 and IL-1β peaked at 24 h after bacterial challenge and then started to decline for up to 2 weeks in infected mouse bladders.8 Rapid activation of pro-inflammatory cytokines and their presence for up to 2 weeks indicate that enhanced cytokines can be utilized for the clearance of the bacteria. In our study, the levels of cytokines IL-1β and TNF-α at 72 h post-infection were significantly raised compared to 48 hrs post-infection. This cytokines-activated delay may be related to the fact that uropathogenic E. coli (UPEC) strain CFT073 causes milder bladder inflammation than UPEC strain UTI89.25 In the case of a UPEC strain CFT073, a factor called TcpC can bind to the host adaptor proteins Mal and MyD88, then clogs up the Toll-like receptor (TLR) signaling mechanism and inhibit TNF-α and IL-1β production, therefore limits the host defense response.26,27
Our results indicate that 2Abz23S29 at the high dose decreased more bacterial bladder load compared to low dose and increased secretion of the all tested pro-inflammatory cytokines (MIP/2, IL-6, TNF-α, and IL-1β) in infected mice groups. A previous study has shown that high doses of synthetic HNP-1 (100 μg/day) in BALB/c mice with colitis induced by DSS exacerbate the disease by increasing macrophage-derived cytokines TNF-α and IL-1β28 Recently it has been shown that mice inoculated transurethrally with UPEC enhanced the IL-6 release, urothelial stat 3 phosphorylation, and triggered antimicrobial peptides expression through activation of TLR4 by bacterial lipopolysaccharide (LPS).29 CFT073 bacteria tend to attenuate the innate immune system by reducing the early uroepithelial pro-inflammatory cytokines production to gains access to intracellular bacterial communities (IBCs).30 It has also been shown that the deletion of IL-6 can raise IBCs.31–33 The higher numbers of IBCs enhance the risk of recurrent UTI. High-dose 2Abz23S29 can induce pro-inflammatory cytokines production, so it may act to decrease UTI recurrence by reducing IBCs. The basis for the complexity of the innate immune response and alteration in cytokine expression may be related to E. coli strains. The basis for this diversity may be related to different sets of virulence genes between strains. One study reported that anti-inflammatory cytokine IL-10 upregulated after infection with CFT073 in the bladder.34 In contrast, IL-10 was not found during a 2-week time course of UTI89 infection.26
One of the main limitations of the promotion of AMPs for clinical applications is toxicity to host tissues. In our study, we measured the potential cytotoxic effect of 2Abz23S29 peptide on 5637 bladder cell lines. Our results revealed that test peptide has a safety profile at our usage concentrations. However, the cytotoxicity effect of test peptide on a primary urothelial cell line should be further investigated in future studies.
Ciprofloxacin is a second-generation fluoroquinolone (FQ) and commonly used antibiotic to treat UTI. Fluoroquinolones target bacterial DNA gyrase and topoisomerase IV, especially in gram-negative bacteria. Several recent independent studies have shown that ciprofloxacin has anti-inflammatory properties, apart from the antibacterial activity. In the present study, the levels of cytokines IL-6, and MIP/2 in mice treated with a combination of 2Abz23S29 and ciprofloxacin, were significantly higher than the mice untreated. By contrast, the levels of cytokines TNF-α, and especially IL-1β in the 2Abz23S29 plus ciprofloxacin treated mice were not significantly different from the untreated; suggesting that ciprofloxacin at this concentration might have anti-inflammatory activity and reduce the pro-inflammatory effect of 2Abz23S29. Ogino et al reported that the effect of ciprofloxacin (100 µg/mL) on lipopolysaccharide (LPS)-induced pro-inflammatory cytokines production has been revealed strong inhibitory activity on TNF-α production in vivo.35 It has been shown that ciprofloxacin has immunomodulatory effects at different concentrations in bladder epithelial cell lines and on urothelial tissues in murine models of UTI. A study on recurrent UTI infection with UTI89 concluded that the sub-inhibitory concentration of ciprofloxacin has an anti-inflammatory effect and suppressed the pro-inflammatory mediator IL-1β, which produces by activated macrophage. Alternatively, ciprofloxacin dramatically increased both IL-10 and GC-SF during UTI that leads to a reduction of macrophage activity.36 According to a recent study, ciprofloxacin attenuated inflammation by decreasing the LPS-induced release of TNF-α and IL-1β via the TLR4/NF-κB signaling pathway in the microglia cells.37 Finally, it should be mentioned that HNP-1 as an immunostimulatory molecule is chemotactic for naive CD45RA/CD4 T cells, CD8 T cells, immature dendritic cells, and monocytes.38 So the capacity for 2Abz23S29 to increase one cytokine and decrease another cytokine might depend on the accumulation of immune populations and their activity within the bladder during UTI infection. In the future, the antimicrobial and immune-modulatory effect of the test peptide can be strengthened by the addition of some type of comparison of the inflammatory cell population in the urine and bladder. Additionally, if a significant percentage of bladders are completely cleared of UPEC after 48 or 72 hrs, it would indicate that this peptide could be a viable candidate for the treatment of patients who need clean intermittent catheterization.
Conclusion
In conclusion, 2Abz23S29 decreased bacterial load, in the bladder infected mice, due to the direct antibacterial activity. Additionally, this peptide also altered pro-inflammatory cytokines in mice infected in a dose-dependent manner. Both aspects together confirm that a truncated analog of HNP-1 2Abz23S29 has the potential to be an effective agent candidate for the treatment of UTI. In future studies, the monotherapy and combination therapy of 2Abz23S29 with antibiotics that have the synergistic interaction can be considered to combat with UTI caused by MDR E. coli.
Abbreviations
UTI, urinary tract infection; UPEC, uropathogenic E. coli; HNP-1, human neutrophil peptide −1; cip, ciprofloxacin; IL-6, interleukin-6; MIP/2, macrophage inflammatory protein/2; TNF-α, tumor necrosis factor-α; IL-1β, interleukin-1β; AMPs, antimicrobial peptides; HDPs, Host defense antimicrobial peptides; MIC, Minimum inhibitory concentration; FIC, fractional inhibitory concentration; TLR, toll-like receptor; IBC, intracellular bacterial communities.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Foxman B. Urinary tract infection syndromes: occurrence, recurrence, bacteriology, risk factors, and disease burden. Infect Dis Clin North Am. 2014;28:1–13. doi:10.1016/j.idc.2013.09.003
2. Terlizzi ME, Gribaudo G, Maffei ME. UroPathogenic Escherichia coli (UPEC) infections: virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front Microbiol. 2017;8:1566. doi:10.3389/fmicb.2017.01566
3. Walters MS, Lane MC, Vigil PD, et al. Kinetics of uropathogenic Escherichia coli metapopulation movement during urinary tract infection. mBio. 2012;3(1):
4. Flores-Mireles AL, Walker JN, Caparon M, et al. Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat Rev Microbiol. 2015;13:269–284. doi:10.1038/nrmicro3432
5. Engelsöy U, Rangel I, Demirel I. Impact of proinflammatory cytokines on the virulence of uropathogenic Escherichia coli. Front Microbiol. 2019;10:1051. doi:10.3389/fmicb.2019.01051
6. Ching C, Schwartz L, Spencer JD, et al. Innate immunity and urinary tract infection. Pediatr Nephrol. 2020;35:1183–1192. doi:10.1007/s00467-019-04269-9
7. Billips BK, Forrestal SG, Rycyk MT, et al. Modulation of host innate immune response in the bladder by uropathogenic Escherichia coli. Infect Immun. 2007;75:5353–5360. doi:10.1128/IAI.00922-07
8. Ingersoll MA, Kline KA, Nielsen HV, et al. G-CSF induction early in uropathogenic Escherichia coli infection of the urinary tract modulates host immunity. Cell Microbiol. 2008;10:2568–2578. doi:10.1111/j.1462-5822.2008.01230.x
9. Sivick KE, Mobley HL. Waging war against uropathogenic Escherichia coli: winning back the urinary tract. Infect Immun. 2010;78:568–585. doi:10.1128/IAI.01000-09
10. Pachón-Ibáñez ME, Smani Y, Pachón J, et al. Perspectives for clinical use of engineered human host defense antimicrobial peptides. FEMS Microbiol Rev. 2017;41:323–342. doi:10.1093/femsre/fux012
11. Becknell B, Schwaderer A, Hains DS, et al. Amplifying renal immunity: the role of antimicrobial peptides in pyelonephritis. Nat Rev Nephrol. 2015;11:642–655. doi:10.1038/nrneph.2015.105
12. Hancock R, Haney E, Gill E. The immunology of host defence peptides: beyond antimicrobial activity. Nat Rev Immunol. 2016;16:321–334. doi:10.1038/nri.2016.29
13. Mookherjee N, Hancock RE. Cationic host defence peptides: innate immune regulatory peptides as a novel approach for treating infections. Cell Mol Life Sci. 2007;64:922–933. doi:10.1007/s00018-007-6475-6
14. Dabirian S, Taslimi Y, Zahedifard F, et al. Human Neutrophil Peptide-1 (HNP-1): a new anti-leishmanial drug candidate. PLoS Negl Trop Dis. 2013;7(10):e2491. doi:10.1371/journal.pntd.0002491
15. Fruitwala S, El-Naccache DW, Chang TL. Multifaceted immune functions of human defensins and underlying mechanisms. Semin Cell Dev Biol. 2019;88:163–172. doi:10.1016/j.semcdb.2018.02.023
16. Lundy FT, Nelson J, Lockhart D, et al. Antimicrobial activity of truncated alpha-defensin (human neutrophil peptide (HNP)-1) analogues without disulphide bridges. Mol Immunol. 2008;45:190–193. doi:10.1016/j.molimm.2007.04.018
17. Wiegand I, Hilpert K, Hancock RE. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc. 2008;3:163–175. doi:10.1038/nprot.2007.521
18. Wang H, Dong B, Lou L. HNP-3 enhanced the antimicrobial activity of CIP by promoting ATP efflux from P. aeruginosa cells. Mol Biol Rep. 2011;38:2673–2678. doi:10.1007/s11033-010-0409-3
19. Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65:55–63. doi:10.1016/0022-1759(83)90303-4
20. Blango MG, Ott EM, Erman A, et al. Forced resurgence and targeting of intracellular uropathogenic Escherichia coli reservoirs. PLoS One. 2014;9:e93327. doi:10.1371/journal.pone.0093327
21. Prasad SV, Fiedoruk K, Daniluk T, Piktel E, Bucki R. Expression and function of host defense peptides at inflammation sites. Int J Mol Sci. 2020;21:104. doi:10.3390/ijms21010104
22. Miles K, Clarke DJ, Lu W, et al. Dying and necrotic neutrophils are anti-inflammatory secondary to the release of alpha-defensins. J Immunol. 2009;183:2122–2132. doi:10.4049/jimmunol.0804187
23. Maeda T, Sakiyama T, Kanmura S, et al. Low concentrations of human neutrophil peptide ameliorate experimental murine colitis. Int J Mol Med. 2016;38:1777–1785. doi:10.3892/ijmm.2016.2795
24. Kohlgraf KG, Ackermann A, Lu X, et al. Defensins attenuate cytokine responses yet enhance antibody responses to Porphyromonas gingivalis adhesins in mice. Future Microbiol. 2010;5:115–125. doi:10.2217/fmb.09.107
25. Lin AE, Beasley FC, Olson J, et al. Role of Hypoxia Inducible Factor-1α (HIF-1α) in innate defense against uropathogenic Escherichia coli infection. PLoS Pathog. 2015;11(4):e1004818. doi:10.1371/journal.ppat.1004818
26. Cirl C, Wieser A, Yadav M, et al. Subversion of Toll-like receptor signaling by a unique family of bacterial Toll/interleukin-1 receptor domain-containing proteins. Nat Med. 2008;14:399–406. doi:10.1038/nm1734
27. Hamilton C, Tan L, Miethke T, et al. Immunity to uropathogens: the emerging roles of inflammasomes. Nat Rev Urol. 2017;14:284–295. doi:10.1038/nrurol.2017.25
28. Hashimoto S, Uto H, Kanmura S, et al. Human neutrophil peptide-1 aggravates dextran sulfate sodium-induced colitis. Inflamm Bowel Dis. 2012;18:667–675. doi:10.1002/ibd.21855
29. Ching CB, Gupta S, Li B, et al. Interleukin-6/Stat3 signaling has an essential role in the host antimicrobial response to urinary tract infection. Kidney Int. 2018;93:1320–1329. doi:10.1016/j.kint.2017.12.006
30. Olson PD, Hunstad DA. Subversion of Host Innate Immunity by Uropathogenic Escherichia coli. Pathogens. Pathogens. 2016;5(1):
31. Hunstad DA, Justice SS, Hung CS, et al. Suppression of bladder epithelial cytokine responses by uropathogenic Escherichia coli. Infect Immun. 2005;73:3999–4006. doi:10.1128/IAI.73.7.3999-4006.2005
32. Hilbert DW, Pascal KE, Libby EK, et al. Uropathogenic Escherichia coli dominantly suppress the innate immune response of bladder epithelial cells by a lipopolysaccharide- and Toll-like receptor 4-independent pathway. Microbes Infect. 2008;10:114–121. doi:10.1016/j.micinf.2007.10.012
33. Lloyd AL, Smith SN, Eaton KA, et al. Uropathogenic Escherichia coli suppresses the host inflammatory response via pathogenicity island genes sisA and sisB. Infect Immun. 2009;77:5322–5333. doi:10.1128/IAI.00779-09
34. Duell BL, Carey AJ, Tan CK, et al. Innate transcriptional networks activated in bladder in response to uropathogenic Escherichia coli drive diverse biological pathways and rapid synthesis of IL-10 for defense against bacterial urinary tract infection. J Immunol. 2012;188:781–792. doi:10.4049/jimmunol.1101231
35. Ogino H, Fujii M, Ono M, et al. In vivo and in vitro effects of fluoroquinolones on lipopolysaccharide-induced pro-inflammatory cytokine production. J Infect Chemother. 2009;15:168–173. doi:10.1007/s10156-009-0680-1
36. Goneau LW, Hannan TJ, MacPhee RA, et al. Subinhibitory antibiotic therapy alters recurrent urinary tract infection pathogenesis through modulation of bacterial virulence and host immunity. mBio. 2015;6(2). doi:10.1128/mBio.00356-15.
37. Zusso M, Lunardi V, Franceschini D, et al. Ciprofloxacin and levofloxacin attenuate microglia inflammatory response via TLR4/NF-kB pathway. J Neuroinflammation. 2019;16(1):148. doi:10.1186/s12974-019-1538-9
38. Vordenbäumen S, Schneider M. Defensins: potential effectors in autoimmune rheumatic disorders. Polymers. 2011;3:1268–1281. doi:10.3390/polym3031268
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.