")
Back to Journals » Infection and Drug Resistance » Volume 13
A Novel Transposon, Tn6518, Mediated Transfer of mcr-3 Variant in ESBL-Producing Aeromonas veronii
Authors Wang X, Zhai W, Wang S , Shen Z, Wang Y, Zhang Q
Received 24 November 2019
Accepted for publication 11 January 2020
Published 25 March 2020 Volume 2020:13 Pages 893—899
DOI https://doi.org/10.2147/IDR.S239865
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Xiaoming Wang,1,2 Weishuai Zhai,3 Shaolin Wang,2 Zhangqi Shen,2 Yang Wang,2 Qidi Zhang3
1MOE Joint International Research Laboratory of Animal Health and Food Safety, College of Veterinary Medicine, Nanjing Agricultural University, Nanjing, People’s Republic of China; 2Beijing Advanced Innovation Center for Food Nutrition and Human Health, College of Veterinary Medicine, China Agriculture University, Beijing, People’s Republic of China; 3College of Veterinary Medicine, Qingdao Agricultural University, Qingdao, Shandong, People’s Republic of China
Correspondence: Qidi Zhang No. 700 Changcheng Road, Chengyang District, Qingdao City, Shandong Province 266109, People’s Republic of China
Email [email protected]
Purpose: The aim of this study was to determine the prevalence and transmission mechanism of mcr-3 in Aeromonas spp. isolated from chicken cloaca.
Materials and Methods: A. veronii w55 was isolated from chicken in 2008. PCR assay was used to detect mcr genes and putative circular intermediate. Susceptibility testing was identified by the microdilution method. WGS was performed to obtain the whole sequence. S1-PFGE and DNA southern hybridization were used to study the location of mcr-3.6.
Results: PCR-based analysis indicated that 1 out of 55 Aeromonas spp. isolates was mcr-3-positive. Whole-genome sequencing revealed that the strain A. veronii w55 belonged to novel sequence type ST514 and had two adjacent chromosomally located mcr variants, mcr-3.6 and mcr-3-like. The mcr-3.6 and mcr-3-like genes showed 93.67% and 82.84% nucleotide sequence identity, respectively, to original mcr-3 from E. coli. A. veronii w55 also exhibited resistance to extended-spectrum β-lactams and was positive for blaPER-3, and this is the first time to report blaPER-3 in A. veronii. Genetic environment analysis revealed that the segment of mcr-3.6-mcr-3-like-dgkA was flanked by five insertion sequence elements originated from Aeromonas species, and the structure of ISAs2-ISAhy2-ISAs20-mcr-3.6-mcr-3-like-dgkA-ISAs2 was designated as a novel transposon Tn 6518, in which an 8405-bp circular intermediate carrying two mcr-3 variants can be looped out.
Conclusion: This result suggested the mcr-3 variant genes could be disseminated between various Aeromonas species via transposon-mediated transmission.
Keywords: mcr-3.6, Tn 6158, Aeromonas veronii
Introduction
Aeromonas species are Gram-negative, rod-shaped bacteria with a broad host range. To date, they have been shown to infect a vast number of hosts, including fish, domestic chickens, lower and higher vertebrates, and humans.1–3 In animals, Aeromonas species cause illnesses ranging from furunculosis and diarrhea to septicemia.4 However, because of the wile use of broad-spectrum antibiotics in clinical settings, agriculture, and aquaculture, there has been an increase in antibiotic resistance among disease-causing Aeromonas species.
Colistin, a polycationic peptide, is considered as one of the last-resort drugs for the treatment of infections caused by multi-drug resistant Gram-negative bacteria.5 In 2016, the first plasmid-mediated colistin resistance gene mcr-1 was characterized in Enterobacteriaceae.6 To date, nine mobile colistin resistance determinants (mcr-1~mcr-9) have been reported in Enterobacteriaceae, Moraxella and Aeromonas.7–10 Subsequently, multiple mcr-3 variants and one mcr-5 gene were identified in Aeromonas species, implying Aeromonas species plays an important role in the dissemination of mobile colistin resistance genes.11–13 Among them mcr-3 was first found in Escherichia coli isolated from pig,14 then some papers illustrated that this gene emerged in Aeromonas spp.12 Subsequently, Ling et al study showed that mcr-3.3 and mcr-3-like have emerged in Aeromonas veronii isolated from retail chicken meat,8 and Eichhorn et al found four Aeromonas isolates were positive for mcr-3, each of the four mcr-3 genes showed amino acid identities of 95.0–98.0% to the original mcr-3 protein. These variants were designated mcr-3.6 (Aeromonas allosaccharophila from golden orfe), mcr-3.7 (Aeromonas media from), mcr-3.8 (Aeromonas jandaei from koi carp) and mcr-3.9 (Aeromonas hydrophila from koi carp).14 Herein, we investigated the presence of mcr-type genes in Aeromonas spp. from chicken, and identified a novel transposon Tn6518 carrying mcr-3 variant in ESBL-producing Aeromonas veronii.
Materials and Methods
Bacterial Isolates, Screening for mcr-Type, and Species Confirmation
Fifty-five Aeromonas spp. isolates were collected from commercial chickens in Sichuan Province, China, in 2008, and were screened for the presence of mcr-1 to mcr-9 genes using a polymerase chain reaction (PCR)-based assay with primers listed in Table S1. The PCR-positive isolate was then subjected to further PCR and sequencing analysis of the 16S rRNA, gyrB, and rpoB loci for species identification as per Kupfer et al.15
Antimicrobial Susceptibility Testing
The minimum inhibitory concentrations (MICs) of 13 antimicrobial agents (listed in Table S2) against the mcr-3-positive isolate were then determined by broth microdilution according to the recommendations of the Clinical and Laboratory Standards Institute (document VET01-A4). The breakpoints for each antimicrobial agent were interpreted according to CLSI (document M45-A2) and European Committee on Antimicrobial Susceptibility Testing (EUCAST) (http://www.eucast.org) guidelines.16 E. coli ATCC 25922 was used as a quality control strain.
Genomic DNA Sequencing and Analysis of Antibiotic Resistance Genes
Genomic DNA from the mcr-3-positive isolate was subjected to sequence using both the Illumina Hiseq platform and the PacBio RSII System (Sinobiocore, Beijing, China). CLC Genomics Workbench 9.0 (CLC Bio, Aarhus, Denmark) was used for assembling data from Illumina Hiseq platform, and all contigs were screened for the presence of mcr-3 using standalone BLAST analysis. The data from PacBio RSII System were assembled using the Hierarchical Genome Assembly Process (HGAP) and Quiver as part of the SMRT analysis program (version 2.3) using the HGAP3 protocol, and then corrected using Pilon. Reference sequences for antibiotic resistance genes were obtained from the ARG-ANNOT database (https://omictools.com/antibiotic-resistance-gene-annotation-tool). Multilocus sequence typing (MLST) was carried out using the MLST tool from the Center for Genomic Epidemiology (http://genomicepidemiology.org/), and annotation of the genomic sequences was performed using ORF Finder (www.ncbi.nlm.nih.gov/gorf/orfig.cgi), Vector NTI (Invitrogen, Carlsbad, CA, USA), and ISFinder (http://www-is.biotoul.fr).
S1 Nuclease Pulsed-Field Gel Electrophoresis (S1-PFGE) and Southern Blotting
S1-PFGE and Southern blotting were performed to locate the mcr-3 gene within the genome. Briefly, whole cells were embedded in agarose gel plugs and digested with S1 nuclease (TaKaRa, Dalian, China). The DNA was then separated by PFGE as described previously.17 For Southern blotting, a 450-bp digoxigenin-labelled mcr-3 PCR amplicon was used as a probe. Genomic DNA from Salmonella enterica serovar Braenderup strain H9812 restricted with XbaI was used as the DNA marker.
Detection of Circular Intermediates
Based on the knowledge that IS30-family elements can form circular DNA intermediates, and that ISAs2 belongs to the IS30 family,18,19 a set of reverse primers (P-F: GGCTGGTTGTGATTGTAGAG and P-R: TCGCTCGGTCTGTTTGCTTT) were designed to investigate the potential of the ISAs2-ISAhy2-ISAs20-mcr-3.12-mcr-3-like-dgkA-ISAs2 segment to circularize. The designed primers allowed us to obtain a complete map of the circular intermediate.
Functional Cloning of mcr-3.6 and mcr-3-Like Genes in E. coli and Aeromonas Species
To determine the function of mcr-3 variant genes, 1983-bp and 2176-bp DNA fragments, corresponding to the mcr-3.6 and mcr-3-like regions along with their upstream sequences, respectively, were amplified and cloned into pUC19, as previously reported.8 The resulting recombinant plasmids, pUC19-mcr-3.6 and pUC19-mcr-3-like, were then separately electroporated into E. coli DH5α (TaKaRa, Dalian, China) and Aeromonas salmonicida cells originally isolated from chicken cloaca swabs. The recombinant strains were then subjected to antibiotic susceptibility testing.
Results
Prevalence of mcr-1 to mcr-9 in Aeromonas Species Isolates
The use of colistin as animal growth enhancers has been banned in China since April, 2017. However, prior to that time, the extensive use of colistin in food animal production, especially in commercial poultry farming, increased the risk of transfer of colistin resistance determinants in bacteria. Our retrospective survey of mcr-1 to mcr-9 in 55 Aeromonas spp. isolates showed that only one ESBL-producing A. veronii isolate w55 was mcr-3-positive, indicating that the presence of mcr-3 in Aeromonas in China for at least 10 years.
WGS Analysis and Functional Identification of mcr-3.6 and mcr-3-Like Genes
The mcr-3.6 variant has been deposited into NCBI with GenBank number: MF598076.1, and was first detected from A. allosaccharophila strain IMT43045 isolated from German Leuciscus idus.19 Illumina HiSeq 2500-based WGS revealed the presence of two adjacent mcr-3 variants, mcr-3.6 and mcr-3-like separated by only 66 bp, on a 36.9-kb contig of A. veronii w55. The two variants showed 93.7% and 82.8% nucleotide sequence identity, respectively, to the original mcr-3 from porcine E. coli.20 Both variants carried several missense mutations, resulting in 94.8% and 84.2% amino acid sequence similarity, respectively, to MCR-3.
In order to determine the two mcr-3 variants function, we expressed mcr-3.6 and mcr-3-like in E. coli DH5ɑ. Transformants containing pUC19-mcr-3.6 or pUC19-mcr-3-like had colistin MIC of 2 µg/mL and 0.25 µg/mL, respectively, which was 8- and 1-fold higher MIC than that of E. coli DH5ɑ containing pUC19 alone (0.25 µg/mL), respectively. Furthermore, to investigate the function of the two variants in Aeromonas species, recombinant plasmids pUC19-mcr-3.6 and pUC19-mcr-3-like were separately electroporated into competent A. salmonicida cells, and the MICs of colistin against these two transformants were 32 µg/mL (carrying pUC19-mcr-3.6) and 1 µg/mL (carrying pUC19-mcr-3-like), which were 32- and 1-fold higher MIC than that of the wild-type A. salmonicida strain (1 µg/mL), respectively. These results suggested that only mcr-3.6 could confer colistin resistance in both E. coli and other Aeromonas species, which agrees with only pUC19-mcr-3.3 strain confers colistin resistance.8 Although most of the identified mcr-3 variants in Aeromonas species so far derived from animals, the surveillance of these potential high-colistin resistance genes in clinical Aeromonas isolates are also needed.
Antimicrobial Susceptibility Profiles
Antimicrobial susceptibility testing showed that A. veronii w55 was resistance to amoxicillin-clavulanate (MIC, 32/16 μg/mL), cefotaxime (8 μg/mL), ceftriaxone (16 μg/mL), ceftazidime (16 μg/mL), gentamycin (16 μg/mL), and chloramphenicol (32 μg/mL), and was susceptible to aztreonam (2 μg/mL), imipenem (0.03 μg/mL), meropenem (0.03 μg/mL), ciprofloxacin (0.5 μg/mL) and tetracycline (4 μg/mL) (Table S2). Interestingly, despite the presence of the two mcr-3 variants, the MIC value for both colistin and polymyxin B against A. veronii w55 was 2 μg/mL, which was determined as “susceptible” based on the EUCAST clinical breakpoints (http://www.eucast.org).
MLST and Other Resistance Genes
MLST analysis revealed that A. veronii w55 belonged to novel sequence type ST514 (https://cge.cbs.dtu.dk/services/MLST/). WGS analysis showed that A. veronii w55 not only carried colistin resistance genes mcr-3.6 and mcr-3-like but also contained ESBL-producing gene blaPER-3. PER-3 was initially characterised from an Aeromonas punctata in France (GenBank accession no. AY740681.1) and recently in a medical centre in Taiwan,21 sharing 99% homology to blaPER-1. Here we report the first identification of a blaPER-3-producing A. veronii isolate. In addition, aminoglycoside resistance genes strA, strB, aac(6ʹ)Ib-cr, and aacA4, macrolide resistance gene mph(A), phenicol resistance gene catB3, sulfonamide resistance gene sul1, and tetracycline resistance gene tet(E) were also detected in the genome of A. veronii w55. In these resistance genes, mcr-3.6, mcr-3-like, strA, mph(A), aacA4, catB3, sul1, tet(E) located on chromosome, and others located on plasmid.
Location and Genetic Environment of mcr-3.6
S1-nuclease PFGE and Southern blotting using an mcr-3-specific probe showed that mcr-3.6 was located on the chromosome of A. veronii w55 (Figure 1). The segment of mcr-3.6-mcr-3-like-dgkA in a 36.9-kb-size contig was surrounded by five IS elements (two ISAs2, one ISAs20 and two truncated ISAhy2, all of these elements were originally identified from various Aeromonas species, https://www-is.biotoul.fr/scripts/search-db.php), which was different from that in the original mcr-3.6-carrying A. allosaccharophila IMT43045, where this segment was flanked by a truncated IS3 element (Figure 2). In detail, the upstream of mcr-3.6-mcr-3-like-dgkA segment contained one copy of ISAs2, ISAs20, and truncated Δ1ISAhy2 elements. The ISAs2 and ISAs20 elements harbored 50-bp imperfect inverted repeats (IR) at both ends (as shown in Figure 2). In addition, direct target site duplications (5′-CTT-3′ and 5′-AAGG-3′) were also observed immediately up- and downstream of the ISAs2 and ISAs20, respectively (Figure 2). While immediate downstream of mcr-3.6-mcr-3-like-dgkA segment, another copy of ISAs2 and a truncated Δ2ISAhy2 were observed. The IR elements of the downstream ISAs2 were identical to that in upstream; however, the direct target site duplications of the downstream ISAs2 (5′-AAG-3′) were differed from that in the upstream (5′-CCT-3′) (Figure 2).
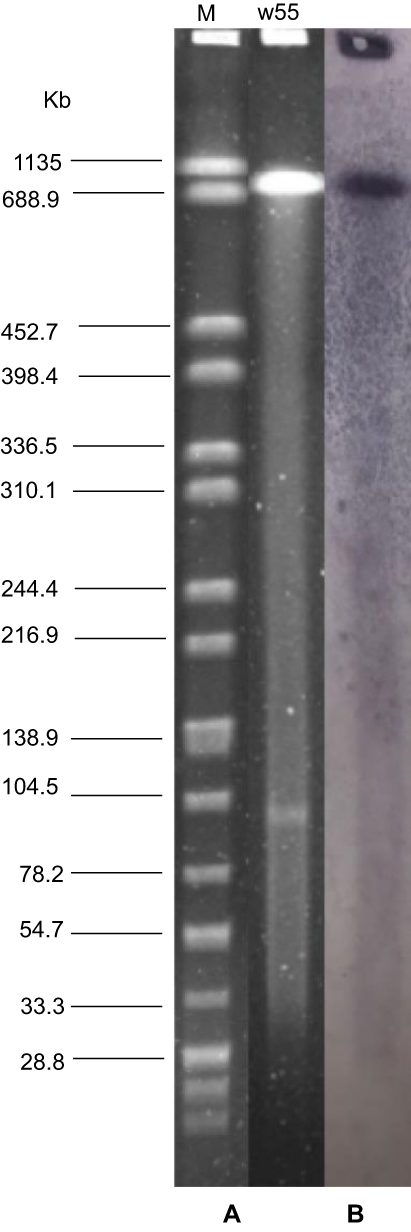 |
Figure 1 Location of mcr-3.6 on the chromosome of Aeromonas veronii w55. (A) S1-PFGE and (B) the corresponding Southern hybridization using the mcr-3.6 probe. Lane M, XbaI-digested Salmonella enterica serovar Braenderup strain H9812 genomic DNA marker; lane w55, A. veronii strain w55 genomic DNA. |
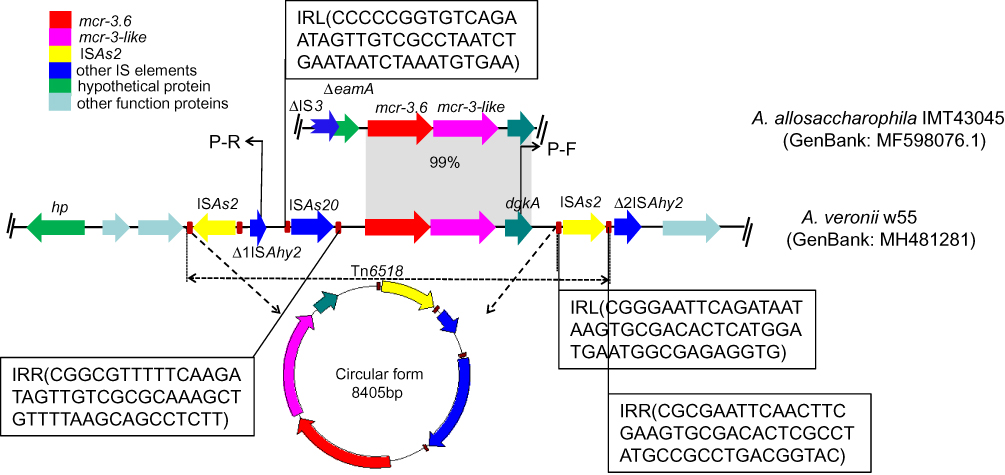 |
Figure 2 Genetic organization of scaffolds containing mcr-3.6 obtained from Aeromonas veronii strain w55 in this study, and structural comparison with A. allosaccharophila IMT43045 (GenBank accession no. MF598076.1). The positions and orientations of the genes are indicated by arrows, with the direction of transcription shown by the arrowhead. Grey shading indicates >99% nucleotide sequence identity. The genetic structure of transposon Tn6518 is shown in circular form. |
Formation of a Circular Intermediate Containing the mcr-3.6 Gene
Some members of the IS family, including IS3, IS30, IS110, IS26, and ISCR1 elements, utilize circular DNA intermediates containing accessory genes to undergo gene translocation via copy-and-paste mechanisms.22 ISAs2, which is first identified in A. salmonicida, disrupts a range of genes in Aeromonas species,23 also belongs to the IS3 family and is flanked by left and right inverted repeats. Therefore, we hypothesize that mcr-3.6 translocation could be mediated via a circular intermediate with the assistance of ISAs2 element. The reverse primers P-F and P-R targeting the Δ1ISAhy2 and dgkA loci, respectively (Figure 2), were used to amplify a putative circular intermediate harboring mcr-3.6 in A. veronii w55. PCR products with a size of ~2.0 kb were obtained and composed of the partial sequence of Δ1ISAhy2 and dgkA and one ISAs2 element. When combined with the sequence between primers P-F and P-R, we concluded that the circular intermediate was 8405-bp in size, and contained one copy of ISAs2, Δ1ISAhy2, ISAs20, and a segment of mcr-3.6-mcr-3-like-dgkA (Figure 2). This new composite transposon was thereafter designated as Tn6518 (ISAs2-Δ1ISAhy2-ISAs20-mcr-3.6-mcr-3-like-dgkA-ISAs2). Nevertheless, the molecular mechanisms underlying the formation of the circular intermediate form of Tn6518, and how it evolves to become a genetically stable fragment in the chromosome, require further investigation.
Discussion
Aeromonas spp. species are recognized enteric pathogens, and it is possible that the presence of Aeromonas isolates that carry these mcr-3 variants in the intestine will facilitate the spread of these genes to other bacterial species living in the same habitat.15,24 Until now, there are several mcr-3 variants, including mcr-3.3 and mcr-3.13 to mcr-3.18, identified among 10 Aeromonas isolates from different sources.12 Shen et al reported samples from the aquatic environment had the highest prevalence (10.0%) of mcr-3-positive Aeromonas isolates,12 which indicated that mcr-3 have been disseminated throughout various environments, the aquatic environment might be the major reservoir. For this study, it is the first case about a novel transposon, Tn6518, mediated transfer of mcr-3.6 in ESBL-producing Aeromonas veronii isolated from chicken faeces.
The first reported mcr-3-carrying E. coli WJ1 exhibited MICs of 8 ug/mL for both colistin and polymyxin B.14 However, five previously identified Aeromonas isolates carrying mcr-3.3 to mcr-3.7 exhibited colistin MICs ranging from 2 to128 ug/mL. In the original report on mcr-3.3 in A. veronii, the mcr-3.3-positive isolate demonstrated borderline susceptibility to colistin (MIC of 2 mg/liter).11 However, E. coli and A. salmonicida transformants carrying pUC19-mcr-3.3 exhibited 8- and 64-fold higher MIC values (MICs of 2 mg/liter and 64 mg/liter, respectively) than the transformant carrying pUC19 alone, which was similar with our findings. These observations not only confirmed that the mcr-3 variants can confer colistin resistance in both Enterobacteriaceae strains and Aeromonads but also suggested that the colistin resistance levels conferred by these genes may vary among different Aeromonas species. In the previous report, the mcr-3 variants were surrounded by many different types of IS, like IS26, IS6100, ISKpn3 and ISAS17.12,15 However, in our study, the genetic environment of mcr-3.6 was ISAs2-ISAhy2-ISAs20-mcr-3.6-mcr-3-like-dgkA-ISAs2, which can form a circular intermediate, and may increase the transmission of colistin resistance.
In conclusion, two mcr-3 variants, mcr-3.6 and mcr-3-like, were identified in ESBL-producing A. veronii isolate w55. This isolate has the novel sequence type ST514 and was isolated from a chicken cloacal sample in 2008. In addition, WGS analysis revealed that the segment of mcr-3.6-mcr-3-like-dgkA was surrounded by multi-copy IS elements and termed as a novel transposon Tn6518, which may be responsible for the transmission of mcr-3.6 among various Aeromonas species. More worrying, Aeromonas species are prevalent in aquatic environments, where they can interact with different bacterial species and genus. Overall, the presence of transposon-carrying mcr-3 variants in Aeromonas species is highly concerning with regards to public health and highlights the need to investigate the prevalence of mcr-3 in high-level colistin-resistant clinical Aeromonas isolates.
Nucleotide Sequence Accession No
The nucleotide sequence reported in this study has been submitted to GenBank with NCBI no. MH481281.
Acknowledgments
We thank Dr. Adam Roberts from UCL Eastman Dental Institute for assistance in registering Tn6518 in the Tn registry.
Funding
This work was supported in part by the grants from the National Key Research and Development Program of China (2016YFD0501304 and 2018YFD0500300), National Natural Science Foundation of China (81661138002), and Qingdao Science and Technology Project (No. 17-3-3-69-nsh).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Yang S, He T, Sun J, et al. distinct antimicrobial resistance profiling of clinically important Aeromonas spp. in Southwest China: a seven-year surveillance study. Infect Drug Resist. 2019;12:2971–2978. doi:10.2147/IDR
2. Castelo-Branco DS, Silva AL, Monteiro FO, et al. Aeromonas and Plesiomonas species from scarlet ibis (Eudocimus ruber) and their environment: monitoring antimicrobial susceptibility and virulence. Antonie Van Leeuwenhoek. 2017;110(1):33–43. doi:10.1007/s10482-016-0771-9
3. Dong HT, Techatanakitarnan C, Jindakittikul P, et al. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L.). J Fish Dis. 2017;40:1395–1403.
4. Dias C, Borges A, Saavedra MJ, et al. Biofilm formation and multidrug-resistant Aeromonas spp. from wild animals. J Glob Antimicrob Resist. 2018;12:227–234. doi:10.1016/j.jgar.2017.09.010
5. Li J, Nation RL, Turnidge JD, et al. Colistin: the re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infect Dis. 2006;6(9):589–601. doi:10.1016/S1473-3099(06)70580-1
6. Liu Y, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-7
7. AbuOun M, Stubberfield EJ, Duggett NA, et al. mcr-1 and mcr-2 variant genes identified in Moraxella species isolated from pigs in Great Britain from 2014 to 2015. J Antimicrob Chemother. 2017;72(10):2745–2749. doi:10.1093/jac/dkx286
8. Ling Z, Yin W, Li H, et al. Chromosome-mediated mcr-3 variants in Aeromonas veronii from chicken meat. Antimicrob Agents Chemother. 2017;61(11):e01272–17.
9. Roschanski N, Falgenhauer L, Grobbel M, et al. Retrospective survey of mcr-1 and mcr-2 in German pig-fattening farms, 2011–2012. Int J Antimicrob Agents. 2017;50(2):266–271. doi:10.1016/j.ijantimicag.2017.03.007
10. Carroll LM, Gaballa A, Guldimann C, et al. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica Serotype Typhimurium Isolate. mBio. 2019;10(3):e00853–19.
11. Wang X, Zhai W, Li J, et al. Presence of an mcr-3 variant in Aeromonas caviae, Proteus mirabilis, and Escherichia coli from one domestic duck. Antimicrob Agents Chemother. 2018;62(2):e02106–17.
12. Shen Y, Xu C, Sun Q, et al. Prevalence and genetic analysis of mcr-3-positive Aeromonas species from humans, retail meat, and environmental water samples. Antimicrob Agents Chemother. 2018;62(9):e00404–18.
13. Ma S, Sun C, Hulth A, et al. Mobile colistin resistance gene mcr-5 in porcine Aeromonas hydrophila. J Antimicrob Chemother. 2018;73(7):1777–1780. doi:10.1093/jac/dky110
14. Eichhorn I, Feudi C, Wang Y, et al. Identification of novel variants of the colistin resistance gene mcr-3 in Aeromonas spp. from the national resistance monitoring programme GERM-Vet and from diagnostic submissions. J Antimicrob Chemother. 2018;73(5):1217–1221. doi:10.1093/jac/dkx538
15. Kupfer M, Kuhnert P, Korczak BM, Peduzzi R, Demarta A Peduzzi R, Demarta A. Genetic relationships of Aeromonas strains inferred from 16S rRNA, gyrB and rpoB gene sequences. Int J Syst Evol Microbiol. 2006;56(Pt 12):2743–2751.
16. Shen Z, Wang Y, Shen Y, et al. Early emergence of mcr-1 in Escherichia coli from food-producing animals. Lancet Infect Dis. 2016;16(3):293. doi:10.1016/S1473-3099(16)00061-X
17. Hu YY, Wang YL, Sun QL, et al. Colistin resistance gene mcr-1 in gut flora of children. Int J Antimicrob Agents. 2017;50(4):593–597. doi:10.1016/j.ijantimicag.2017.06.011
18. He S, Hickman AB, Varani AM, et al. Insertion sequence IS26 reorganizes plasmids in clinically isolated multidrug-resistant bacteria by replicative transposition. mBio. 2015;6(3):e762. doi:10.1128/mBio.00762-15
19. Olasz F, Farkas T, Kiss J, et al. Terminal inverted repeats of insertion sequence IS30 serve as targets for transposition. J Bacteriol. 1997;179(23):7551–7558. doi:10.1128/JB.179.23.7551-7558.1997
20. Yin W, Li H, Shen Y, et al. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio. 2017;8:e543.
21. Wu CJ, Chuang YC, Lee MF, et al. Bacteremia due to extended-spectrum-β-lactamase-producing Aeromonas spp. at a Medical Center in Southern Taiwan. Antimicrob Agents Chemother. 2011;55(12):5813–5818. doi:10.1128/AAC.00634-11
22. Li R, Xie M, Zhang J, et al. Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J Antimicrob Chemother. 2017;72(2):393–401. doi:10.1093/jac/dkw411
23. Gustafson CE, Chu S, Trust TJ. Mutagenesis of the paracrystalline surface protein array of Aeromonas salmonicida by endogenous insertion elements. J Mol Biol. 1994;237(4):452–463. doi:10.1006/jmbi.1994.1247
24. Batra P, Mathur P, Misra MC. Aeromonas spp.: an emerging nosocomial pathogen. J Lab Physicians. 2016;8(1):1–4. doi:10.4103/0974-2727.176234
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.