")
Back to Journals » Infection and Drug Resistance » Volume 12
A Newly Discovered Drug Resistance Gene rfaF In Helicobacter pylori
Authors Lin J , Zhang X, Wen Y , Chen H, She F
Received 16 September 2019
Accepted for publication 30 October 2019
Published 12 November 2019 Volume 2019:12 Pages 3507—3514
DOI https://doi.org/10.2147/IDR.S231152
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eric Nulens
Jiansheng Lin,1,2 Xiaoyan Zhang,1,2 Yancheng Wen,1,2 Hao Chen,1,2 Feifei She1,2
1Key Laboratory of Ministry of Education for Gastrointestinal Cancer, School of Basic Medical Sciences, Fujian Medical University, Fuzhou 350122, People’s Republic of China; 2Fujian Key Laboratory of Tumor Microbiology, School of Basic Medical Sciences, Fujian Medical University, Fuzhou 350122, People’s Republic of China
Correspondence: Feifei She
Key Laboratory of Ministry of Education For Gastrointestinal Cancer, School of Basic Medical Sciences, Fujian Medical University, Fuzhou 350122, People’s Republic of China
Tel +86 591 22862863
Email [email protected]
Background: The purpose of this study was to understand the function of rfaF gene in Helicobacter pylori antibiotic resistance.
Methods: The gene homologous recombination method was used for knockout and complementation of H. pylori rfaF gene. Various constructed strains were analysed for drug sensitivity to amoxicillin (AMO), tetracycline (TET), clarithromycin (CLA), metronidazole (MET), levofloxacin (LEV), and chloramphenicol (CHL) by agar plate dilution method. Drug sensitivity was further confirmed using a growth inhibition curve. Ethidium bromide (EB) accumulation experiments were performed to assess cell membrane permeability. PCR and sequence analysis were used to detect the rfaF gene.
Results: The minimum inhibitory concentrations (MIC) of TET, CHL, AMO, and CLA in 11,637 rfaF knockout strain (ΔrfaF strain) were 4, 4, 2, and 2 times higher than those in 11,637 wild type (WT) strain, respectively. A multidrug-resistant (MDR) ΔrfaF strain also displayed the same trend; however, the degrees of increase were relatively small. Growth inhibition experiments indicated that the growth of the 11,637 ΔrfaF strain was higher with antibiotics at the MIC of the 11,637 WT strain than that of 11,637 rfaF-complemented strain (ΔrfaF/rfaF strain), whereas the 11,637 WT strain did not exhibit any growth. The 11,637 ΔrfaF strain was significantly reduced compared with the cumulative EB fluorescence intensity of the 11,637 WT and of 11,637ΔrfaF/rfaF strain, and the same trend appeared in the MDR strain. Among the 10 clinical strains, 9 clinical strains were found to have mutations in the conserved sequence of rfaF amino acids.
Conclusion: We found a new drug resistance gene, rfaF, in H. pylori, which changes the permeability of cell membrane to confer cross-resistance to AMO, TET, CLA, and CHL and is involved in clinical strain drug resistance. It can be used as a drug target.
Keywords: rfaF, amoxicillin, clarithromycin, tetracycline, resistance
Introduction
Helicobacter pylori (H. pylori), which is a spiral, gram-negative bacterium, has affected more than half of the world population. It is known to be involved in the pathogenesis of chronic gastritis, peptic ulcer disease, gastric cancer, and mucosa-associated lymphoid tissue lymphoma.1,2 Clinically, two or three antibiotics out of the following list: amoxicillin (AMO), tetracycline (TET), clarithromycin (CLA), metronidazole (MET), and levofloxacin (LEV) combined with a proton-pump inhibitor (PPI) or bismuth salts are typically used to kill H. pylori.3,4 With the widespread use of a large number of antibiotics, H. pylori has displayed high antibiotic resistance rate in recent years.5,6
In addition to drug target-binding point mutations and effective efflux, an important mechanism of H. pylori antibiotic resistance is to reduce drug permeability.7–10 The rfaF (waaF) gene, mutated to produce a bacterium with a deep coarse lipopolysaccharide (LPS) phenotype, was identified to function as a core heptose transferase.11 Previous research has demonstrated that the LPS core biosynthetic pathway gene rfaF is involved in drug resistance.12–14 Structural analysis identified the activity of HP1191 in H. pylori 26,695 genome as a heptosyltransferase and a rfaF homolog. The rfaF was confirmed to be involved in biosynthesis of the inner-core region of H. pylori LPS.15–17 However, the role of rfaF in drug tolerance has been unclear in H. pylori. Therefore, the purpose of this study was to investigate the mechanism by which rfaF function affects drug sensitivity.
Methods
Bacterial Culture
H. pylori 11,637 and 10 clinical isolates of H. pylori, which were kindly offered by the Department of Medical Microbiology of Fujian Medical University, were used in this study. The strains were conserved at −80°C at 20% sucrose and 50% fetal bovine serum (FBS). The strains were cultured under a microaerobic environment (5% O2, 10% CO2, 85% N2) at 37°C on Campylobacter agar plates containing 7% sheep blood. The liquid culture medium for H. pylori consisted of Brucella broth and 10% FBS, and the cells were incubated in a shaker at 120 rpm at 37°C. The mutant strains were cultured on agar plates with kanamycin (MP Biomedicals, LLC) at 5 µg/mL or CHL (Inalco S.p.A., Milano Italy) at 8 µg/mL.
Construction Of rfaF Mutant And Complemented Strains By Allelic Exchange Mutagenesis
To construct a rfaF knockout mutant of H. pylori 11,637, an upstream 718 bp fragment and a downstream 777 bp fragment of rfaF were amplified from wild type (WT) 11,637 genomic DNA with primer F1, R1, F2, and R2, respectively. These two fragments and a kanamycin resistance cassette, which was amplified with F3 and R3, were ligated into a pBluescript II vector by ligase independent cloning (LIC) with ClonExpress MultiS One Step Cloning Kit (Vazyme Biotech Co., Ltd). The vectors were subsequently introduced into H. pylori 11,637 strains by electrotransformation. Chromosomal DNA of the transformants was checked by PCR with primers F4 and R4. The primers used in this study are listed in Table 1.
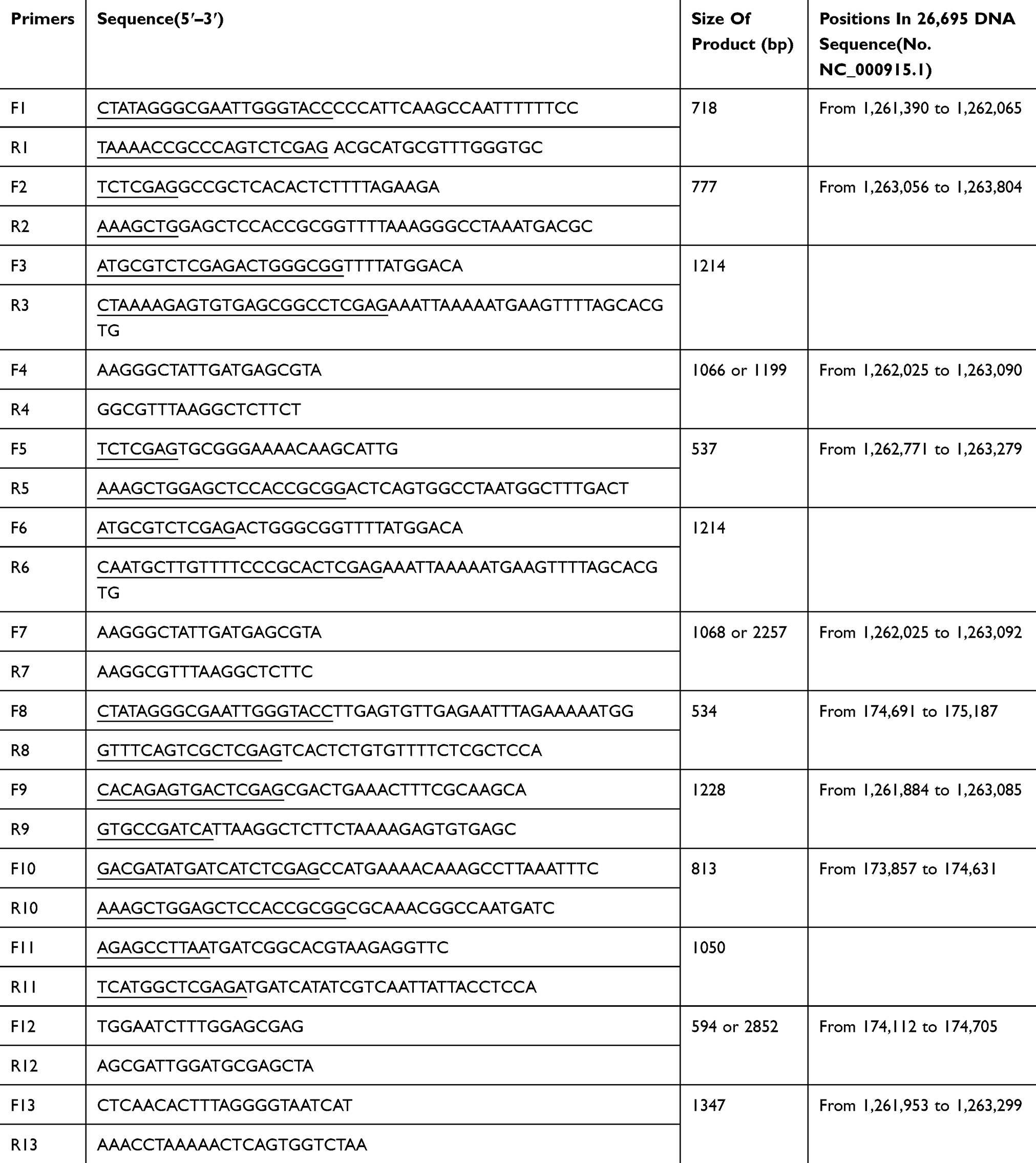 |
Table 1 PCR Primers Used For Construction In This Study |
To construct an rfaF partial knockout mutant of a H. pylori multidrug-resistant (MDR) strain, which was randomly selected from 10 clinical isolates and numbered as FZ068, the same upstream fragment and a 537 bp downstream fragment from +754 to +1263 of rfaF ORF were amplified from WT MDR genomic DNA with primer F1, R1, F5, and R5, respectively. These two fragments and a kanamycin resistance cassette, which was amplified with F6 and R6, were ligated into a pBluescript II vector by LIC with ClonExpress MultiS One Step Cloning Kit (Vazyme Biotech Co., Ltd). The vectors were subsequently introduced into H. pylori MDR strains by electrotransformation. Chromosomal DNA of the transformants was checked by PCR with primers F7 and R7 (Table 1).
To construct a H. pylori 11,637 rfaF-complemented strain, an upstream 534 bp fragment, a 1228 bp fragment of rfaF (containing rfaF 150 bp promoter region and the whole rfaF ORF), and a downstream 813 bp fragment were amplified from wild type 11,637 genomic DNA with primer F8, R8, F9, R9, F10, and R10, respectively. These three fragments and a chloramphenicol resistance cassette which was amplified with F11 and R11 were ligated into a pBluescript II vector by LIC with ClonExpress MultiS One Step Cloning Kit (Vazyme Biotech Co., Ltd). The vectors were then introduced into 11,637 rfaF knockout strains by electrotransformation. Chromosomal DNA of the transformants was checked by PCR with primers F12 and R12 (Table 1).
Antibiotic Susceptibility Testing
The MICs of AMO, TET, CLA, MET, LEV, and CHL for all strains were determined by the agar dilution method reported by Osato, et al.18 Briefly, 2 µL of the 2.0 McF suspension cultures was inoculated on a Mueller-Hinton agar plate containing twofold dilutions of an antibiotic. All the plates were incubated at 37°C under microaerobic conditions, and the MIC values were determined after 72 hrs. H. pylori 11,637 was used as a reference strain for quality control of antibiotic susceptibility testing. Each experiment was repeated at least three times.
Determination Of Growth Curves And Growth Inhibition Curves
The growth curve was determined as described in previous literature.19 To monitor the growth curve of bacteria, the initial optical density (OD600) value of the bacterial suspension in Brucella broth was measured as 0.1 and then the bacteria were shake cultured at 37°C for 96 hrs. The OD600 value of the bacterial suspension was recorded every 8 h. Each experiment was repeated at least three times.
To analyse the growth inhibition curve, the H. pylori strains were inoculated at initial OD600 value of 0.1 in Brucella broth, which contained different antibiotics, and then the bacteria were further cultured for 96 hrs at 37°C with shaking, and the OD600 value of the bacterial suspension was recorded every 8 hrs. Each experiment was repeated at least three times.
Determination Of The Accumulation Of Ethidium Bromide (EB)
The EB assay was modified as described previously.20 H. pylori was cultured on Campylobacter blood agar plates for 48 hrs. The cells were subsequently washed twice with PBS (pH 7.0). The cells were resuspended in 1 mL of PBS (pH 7.0) to an OD600 of 0.4 and were incubated at 37°C for 30 mins in the presence of 10 μM carbonyl cyanide m-chlorophenylhydrazine (CCCP) to deplete cells of metabolic energy. Subsequently, the cells were washed three times with PBS (pH 7.0) and 10 μg/mL of EB was added. At 30 time points in 30 mins, EB fluorescence intensity was measured by using Synergy™ HT (BioTek®) with an excitation measurement wavelength at 530/25 nm and an emission wavelength at 590/35 nm.
Sequence Data Analysis
The rfaF HS PCR products amplified from DNA of 11,637 and 10 clinical strains with primers F13 and R13 were ligated into the PLB plasmids. The ligation plasmids were sent to SunyBiotech Co., Ltd (Fuzhou) for sequence analysis. Finding conserved domains in 26,695 rfaF amino acid sequence (NCBI Reference Sequence: NP_207982.1) was performed by NCBI Conserved Domain Search (CD-Search). We used CLC Sequence Viewer 7 software for amino acid conserved domain comparison analysis.
Statistical Analysis
Data are presented as mean ± standard errors of mean. Unpaired t-test was used for statistical analysis of the two groups. P < 0.05 was considered statistically significant. GraphPad Prism software 6.07 was used to analyse the results.
Ethical Approval
This study was approved by the Fujian Medical University Biomedical Research Ethics Committee ([2012] Fujian Medical University Ethics Review No. (52)).
Results
rfaF Conferred Cross-Resistance To AMO, TET, CLA, And CHL
Assessment Of Susceptibility To Antimicrobial Agents In Various Types Of Strains
In order to assess the drug tolerance of rfaF, we constructed knockout and complemented strains of rfaF. The MIC level was determined by agar plate dilution method. Using CHL as a screening marker for the construction of rfaF-complemented strain, it was unexpectedly found that 11,637 ΔrfaF was resistant to CHL; therefore, we increased the amount of CHL to 8 µg/mL. It was identified that in 11,637 and MDR strains, MIC levels of ΔrfaF strain did not change compared with those of MEM and LEV of WT strain, but interestingly, MIC levels of ΔrfaF strain increased to varying degrees compared with those of AMO, TET, CLA, and CHL of WT strain (Table 2). The MICs of TET, CHL, AMO, and CLA of 11,637 ΔrfaF strain were 4, 4, 2, and 2 times higher than those of 11,637 WT strain, respectively. By successfully constructing the 11,637 ΔrfaF-complemented strain, MIC values of AMO, TET, CLA, and CHL in 11,637 ΔrfaF/rfaF strain decreased compared with those in 11,637 ΔrfaF. These results imply that rfaF confers cross-resistance to AMO, TET, CLA, and CHL but not to MEM and LEV, pointing out a unique resistance mechanism.
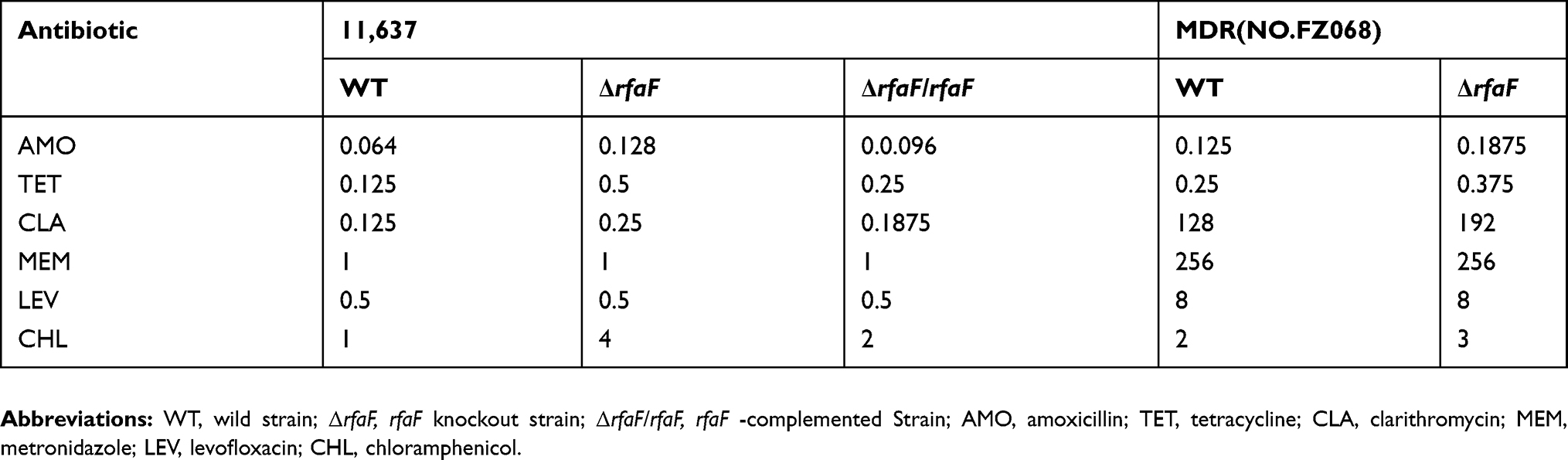 |
Table 2 MIC Values Of Various Types Of H. Pylori To Antimicrobial Agents |
Growth Inhibition Experiment
These drug susceptibility results were further confirmed by growth inhibition experiments. Growth curve showed that 11,637 ΔrfaF strain grew faster than 11,637 WT and 11,637 ΔrfaF/rfaF strain (Figure 1A). We treated the 11,637 WT, 11,637 ΔrfaF, and 11,637 ΔrfaF/rfaF strain with three antibiotics (AMO, TET, and CLA) at the MIC of the 11,637 WT strain and then observed their growth inhibition. We found that compared with the growth curve of the 11,637 ΔrfaF strain, 11,637 WT was significantly inhibited, and 11,637 ΔrfaF/rfaF strain was slightly inhibited (Figure 1B–D).
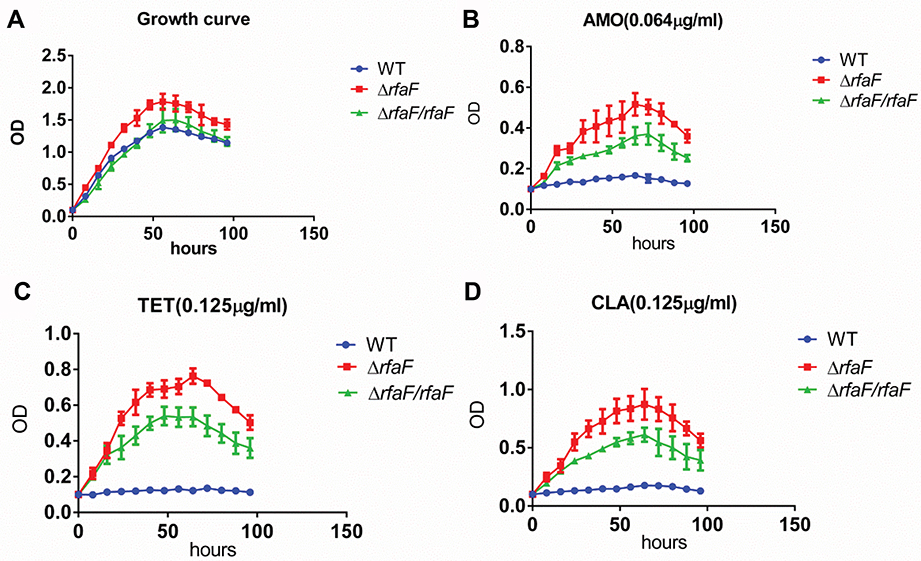 |
Figure 1 Growth curve of various types of H. pylori 11,637 with no drug or with drug. Notes: (A) Growth curve without drug or with (B) AMO (0.064 µg/mL); (C) TET (0.125 µg/mL); and (D) CLA (0.125 µg/mL). |
Overall, these results confirm that rfaF mutation leads to more resistance to AMO, TET, and CLA.
rfaF Changed Cell Membrane Permeability
rfaF is involved in lipopolysaccharide (LPS) synthesis of H. pylori. In addition, changes in LPS structure cause changes in cell membrane permeability leading to changes in drug concentration in the cell membrane, which suggests that H. pylori rfaF may alter cell membrane permeability to drugs. Therefore, in order to figure out the resistance mechanism of rfaF, we compared the cumulative fluorescence intensity of EB on the cell membranes of 11,637 WT, 11,637 ΔrfaF, 11,637 ΔrfaF/rfaF, MDR WT, and MDR ΔrfaF strains. We found that compared with WT strain, 11,637 ΔrfaF and MDR ΔrfaF strains showed reduced EB cumulative fluorescence intensity. The change in degree of EB fluorescence cumulative intensity between 11,637 WT strain and 11,637 ΔrfaF strain is greater than that between MDR WT strain and MDR ΔrfaF strain, which was consistent with drug sensitivity change trends (Figure 2). Compared with 11,637 ΔrfaF strain, EB cumulative fluorescence intensity of 11,637 ΔrfaF/rfaF strain was significantly increased. These results indicate that rfaF knockout resulted in a decrease in EB permeability through the cell membrane. Therefore, we conclude that rfaF knockout reduces drug sensitivity because its mutation decreases the permeability of the cell membrane to the drug, which results in a decrease in drug concentration in the cell membrane.
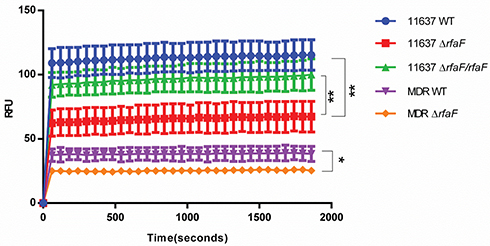 |
Figure 2 Comparison of EB accumulation in various types of H. pylori strains. Notes: The data presented are the means ± standard errors of the means from three separate experiments. *P<0.05; **P<0.01. |
High Mutation Rate Of Amino Acid Conserved Domains Of rfaF From Clinical Isolates Associated With Antibiotic Resistance
To determine whether the target gene rfaF mutation is also involved in the resistance of clinical isolates, we compared the amino acid sequence of the target protein of the clinical strains with the conserved amino acid sequence of the target protein in NCBI. Insertions, deletions, and amino acid substitutions found only in highly conserved portions of the target protein in clinical isolates are believed to impart a change in susceptibility reduction.12 We screened 10 clinical strains with relatively high MIC levels of AMO, TET, and CLA and 11,637 WT strain, and sequenced rfaF. After alignment analysis, although no deletions or insertions were found, there were conservative amino acid substitutions in 9 out of 10 clinical strains, and no mutations were found in 11,637 WT strain (Table 3). In addition, interestingly, K331R mutation type accounted for 44.44% of all mutants. Therefore, we conclude that the high mutation rate of the rfaF in clinical strains is an important factor causing resistance to AMO, TET, and CLA.
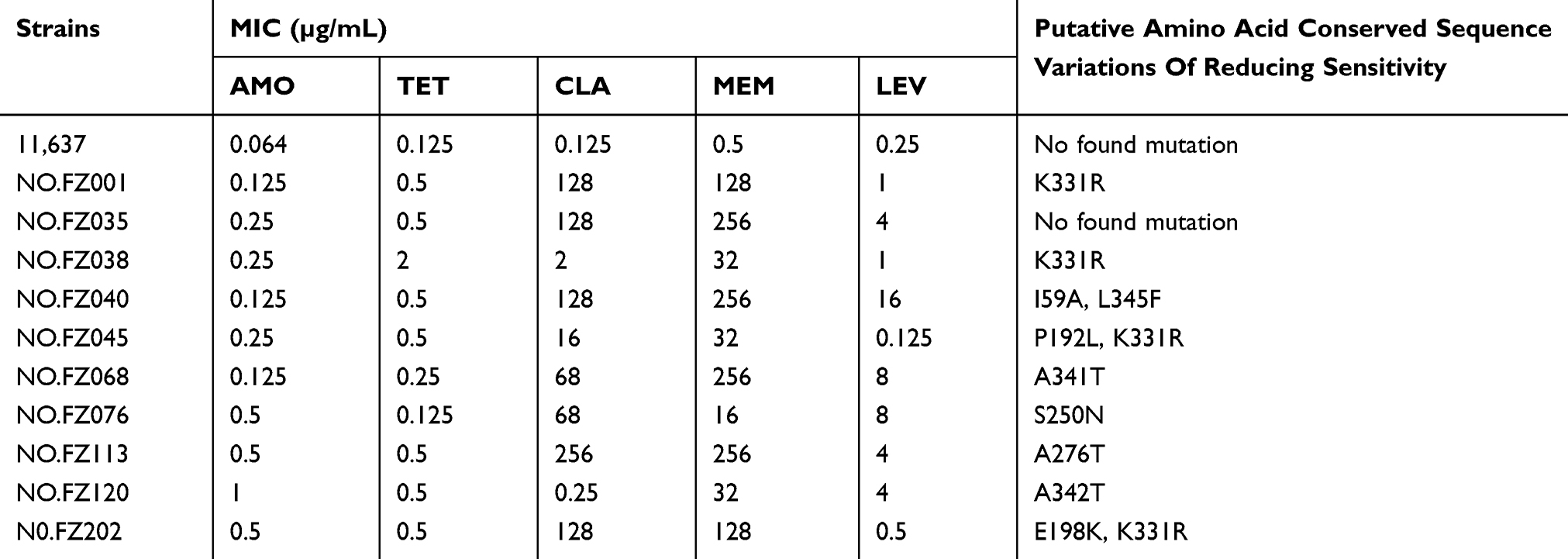 |
Table 3 rfaF Amino Acid Mutation Analysis |
Discussion
Although the resistance to AMO and TET is low, the resistance rates of these two antibiotics and CLA have increased in recent years.21–24 The rfaF (waaF) gene encodes a protein that transfers a second heptose residue and integrates it into the core of the LPS. Disruption of heptose transport results in a herpes-free phenotype called “deep roughness”.25 To evaluate the function of rfaF, we generated knockout strains from both 11,673 WT and MDR WT strain (No. FZ068) simultaneously. The results revealed that their ΔrfaF was cross-resistant to AMO, TET, and CLA. The role of rfaF in causing drug resistance is universal. In agreement with our results, previous studies have reported that Escherichia coli rfaF mutation reduced drug sensitivity to AMO and TET.12 This may be due to the mutation that causes a decline in the expression of a drug-dependent pore protein or a transferring protein, resulting in the decrease of cell membrane permeability to the drug, thus decreasing the concentration of the drug in the cell. However, chloramphenicol susceptibility results are contradictory. The reason for this difference is that rfaF mutation led to the expression of chloramphenicol-dependent porins to be completely reversed. Our results show that compared with the MIC values of AMO, TET, CLA, and CHL of MDR WT strain, MDR ΔrfaF strain only increased by 0.5-fold. One of the possible reasons may be that the knockout strategy of MDR strain was incomplete, which retained a part of the ORF sequence, and this part may continue to express partially functional proteins. The other more likely reason is that rfaF in MDR may have been mutated before it was knocked out, so the rfaF protein activity may be partially inactivated. In line with our inference, the A341T mutation was found in the MDR rfaF strain by alignment analysis of amino acid conserved sequences. Coincidentally, in another study, the rfaF mutation in clinical E. coli isolates also occurred at position 341 where A was replaced by T to confer antibiotic resistance.12 If this mutation can lead to a partial inactivation of the protein, then the basic drug MIC value of the MDR strain will increase. Consistent with our hypothesis, the results showed that the MICs of AMO, CLA, TET, and CHL of MDR WT strain were higher than that of 11,637 WT strain.
EB cumulative fluorescence was used to reflect drug permeability: the lower MIC value, the stronger the EB cumulative fluorescence intensity.8,26 We found that the cumulative EB fluorescence intensity of MDR WT was lower than that of 11,637 WT, which is consistent with the result that the basic MIC values of AMO, TET, and CLA in MDR WT strain were higher than that of 11,637 WT strain. The cumulative fluorescence intensity of EB of ΔrfaF was significantly lower in 11,637 and in MDR than that of the WT strain, which was consistent with the significant increase in MIC levels of AMO, TET, and CLA of ΔrfaF strains. Our findings indicate that the higher the MIC level of the drug, the lower the EB cumulative fluorescence. A drug permeability mechanism study similar to ours indicates that TET-resistant strains reduce the accumulation of TET.10 We conclude that the mechanism of reducing drug sensitivity of ΔrfaF is the decrease in drug permeability of the cell membrane.
This study discovered that H. pylori rfaF gene was directly related to drug sensitivity, and its mechanism of drug resistance was the decrease in drug permeability of the cell membrane. Interestingly, we found that in the clinical strains, rfaF amino acid conserved sequence had high mutation rate and the K331R mutation was the most frequent (44.44% of all mutations). This result suggests that rfaF confers resistance to drugs in clinical strains. The high mutation rate of rfaF accounted for the relatively high levels of MIC of AMO, TET, and CLA. Further analysis needs to be performed to confirm the active site of rfaF protein. In conclusion, this study provides a useful target gene for finding solutions to antibiotic resistance.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Peek RM, Blaser MJ. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat Rev Cancer. 2002;2(1):28–37. doi:10.1038/nrc703
2. Delaney BC, Paul M, David F. Helicobacter pylori infection. N Engl J Med. 2005;2007(15):518–534.
3. Garza-González E, Perez-Perez G, Maldonado-Garza H, et al. A review of Helicobacter pylori diagnosis, treatment, and methods to detect eradication. World J Gastroenterol. 2014;20(6):1438–1449. doi:10.3748/wjg.v20.i6.1438
4. Goderska K, Pena SA, Alarcon T. Helicobacter pylori treatment: antibiotics or probiotics. Appl Microbiol Biotechnol. 2018;102(1):1–7. doi:10.1007/s00253-017-8535-7
5. Song Z, Zhou L, Zhang J, He L, Bai P, Xue Y. Hybrid therapy as first-line regimen for Helicobacter pylori eradication in populations with high antibiotic resistance rates. Helicobacter. 2016;21(5):382–388. doi:10.1111/hel.2016.21.issue-5
6. Hashemi SJ, Sheikh A, Goodarzi H, et al. Genetic basis for metronidazole and clarithromycin resistance in Helicobacter pylori strains isolated from patients with gastroduodenal disorders. Infect Drug Resist. 2019;12:535–543. doi:10.2147/IDR.S192942
7. Hu Y, Zhang M, Lu B, Dai J. Helicobacter pylori and antibiotic resistance, a continuing and intractable problem. Helicobacter. 2016;21(5):349–363. doi:10.1111/hel.2016.21.issue-5
8. Chiu HC, Lin TL, Yang JC, Wang JT. Synergistic effect of imp/ostA and msbA in hydrophobic drug resistance of Helicobacter pylori. BMC Microbiol. 2009;9(1):136. doi:10.1186/1471-2180-9-136
9. Co EMA, Schiller NL. Resistance mechanisms in an in vitro-selected amoxicillin-resistant strain of Helicobacter pylori. Antimicrob Agents Chemother. 2006;50(12):4174–4176. doi:10.1128/AAC.00759-06
10. Wu JY, Kim JJ, Rita R, Wang WM, Graham DY, Kwon DH. Tetracycline-resistant clinical Helicobacter pylori isolates with and without mutations in 16S rRNA-encoding genes. Antimicrob Agents Chemother. 2005;49(2):578–583. doi:10.1128/AAC.49.2.578-583.2005
11. Allen AG, Isobe T, Maskell DJ. Identification and cloning of waaF (rfaF) from Bordetella pertussis and use to generate mutants of Bordetella spp. with deep rough lipopolysaccharide. J Bacteriol. 1998;180(1):35–40.
12. Linkevicius M, Sandegren L, Andersson DI. Mechanisms and fitness costs of tigecycline resistance in Escherichia coli. J Antimicrob Chemother. 2013;68(12):2809–2819. doi:10.1093/jac/dkt263
13. Burtnick MN, Woods DE. Isolation of polymyxin B-susceptible mutants of Burkholderia pseudomallei and molecular characterization of genetic loci involved in polymyxin B resistance. Antimicrob Agents Chemother. 1999;43(11):2648–2656. doi:10.1128/AAC.43.11.2648
14. Jeon B, Muraoka W,A, Zhang Q. Roles of lipooligosaccharide and capsular polysaccharide in antimicrobial resistance and natural transformation of Campylobacter jejuni. J Antimicrob Chemother. 2009;63(3):462–468. doi:10.1093/jac/dkn529
15. Vandana C, Logan SM, Harrison BA, et al. Characterization of a waaF mutant of Helicobacter pylori strain 26695 provides evidence that an extended lipopolysaccharide structure has a limited role in the invasion of gastric cancer cells. Biochem Cell Biol. 2007;85(5):582–590. doi:10.1139/O07-056
16. Stead CM, Jinshi Z, Raetz CRH, Trent MS. Removal of the outer Kdo from Helicobacter pylori lipopolysaccharide and its impact on the bacterial surface. Mol Microbiol. 2010;78(4):837–852. doi:10.1111/j.1365-2958.2010.07304.x
17. Khamri W, Moran AP, Worku ML, et al. Variations in Helicobacter pylori lipopolysaccharide to evade the innate immune component surfactant protein D. Infect Immun. 2005;73(11):7677. doi:10.1128/IAI.73.11.7677-7686.2005
18. Osato MS, Reddy R, Reddy SG, Penland RL, Graham DY. Comparison of the Etest and the NCCLS-approved agar dilution method to detect metronidazole and clarithromycin resistant Helicobacter pylori. Int J Antimicrob Agents. 2001;17(1):39–44. doi:10.1016/S0924-8579(00)00320-4
19. Geng X, Li W, Chen Z, et al. Bifunctional enzyme SpoT is involved in the clarithromycin tolerance of Helicobacter pylori by up-regulating the transporters HP0939, HP1017, HP0497, and HP0471. Antimicrob Agents Chemother. 2017;61(5):
20. Giraud E, Cloeckaert A, Kerboeuf D, Chaslus-Dancla E. Evidence for active efflux as the primary mechanism of resistance to ciprofloxacin in Salmonella enterica serovar typhimurium. Antimicrob Agents Chemother. 2000;44(5):1223–1228. doi:10.1128/AAC.44.5.1223-1228.2000
21. Mollison LC, Stingemore N, Wake RA, Cullen DJ, Mcgechie DB. Antibiotic resistance in Helicobacter pylori. Med J Aust. 2000;173(10):521–523. doi:10.5694/mja2.2000.173.issue-10
22. Lee JW, Kim N, Kim JM, et al. Prevalence of primary and secondary antimicrobial resistance of Helicobacter pylori in Korea from 2003 through 2012. Helicobacter. 2013;18(3):206–214. doi:10.1111/hel.2013.18.issue-3
23. Shiota S, Reddy R, Alsarraj A, El-Serag HB, Graham DY. Antibiotic resistance of helicobacter pylori among male United States veterans. Clin Gastroenterol Hepatol. 2015;13(9):1616–1624. doi:10.1016/j.cgh.2015.02.005
24. Kocazeybek B, Tokman HB. Prevalence of primary antimicrobial resistance of H. pylori in Turkey: a systematic review. Helicobacter. 2016;21(4):251–260. doi:10.1111/hel.2016.21.issue-4
25. Schnaitman CA, Klena JD. Genetics of lipopolysaccharide biosynthesis in enteric bacteria. Microbiol Rev. 1993;57(3):655.
26. Rodrigues L, Viveiros M, Aínsa JA. Measuring efflux and permeability in mycobacteria. Methods Mol Biol. 2015;1285:227–239.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.