")
Back to Journals » OncoTargets and Therapy » Volume 8
The effects of PGC-1α on the proliferation and energy metabolism of malignant endometrial cancer cells
Authors Ren Z, Yang H, Wang C, Ma X
Received 29 December 2014
Accepted for publication 25 February 2015
Published 9 April 2015 Volume 2015:8 Pages 769—774
DOI https://doi.org/10.2147/OTT.S79960
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Daniele Santini
Zhongqian Ren, Hui Yang, Cuicui Wang, Xiaoxin Ma
Department of Obstetrics and Gynecology, Shengjing Hospital of China Medical University, Shenyang, People’s Republic of China
Background: It is well known that peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α) plays an important role in tissue energy metabolism. However, the roles of PGC-1α in malignant endometrial cancer remain unknown.
Methods: Forty cases of endometrial carcinoma, 15 cases with proliferative endometrial tissues, and 21 cases with normal endometrial tissues were collected. Real-time polymerase chain reaction was used to detect the mRNA levels of PGC-1α and estrogen-related receptor gamma (ERRγ). ELISA (enzyme-linked immunosorbent assay) was used to detect the concentrations of pyruvate kinase and isocitrate dehydrogenase. The results were analyzed using medical statistical methods.
Results: The mRNA levels of PGC-1α and ERRγ in the endometrial carcinoma tissues and hyperplasic endometrial tissues were significantly greater than those in the normal endometria. The mRNA levels of PGC-1α and ERRγ in the endometrial carcinoma patients with type 2 diabetes were higher than those in patients without diabetes. The mRNA levels of PGC-1α and ERRγ in the endometrial adenocarcinomas increased with clinical staging, depth of myometrial invasion, and increases in the number of metastatic lymph nodes. The PGC-1α mRNA level was positively correlated with ERRγ in the endometrial carcinoma tissues. The mRNA levels of PGC-1α were positively correlated with the concentrations of pyruvate kinase and isocitrate dehydrogenase in the endometrial carcinoma tissues, and similar results were found for ERRγ.
Conclusion: Our results suggested that the upregulation of PGC-1α and ERRγ in endometrial cancer might be a requirement for cancer cell energy metabolism, which contributes to the development of endometrial cancer.
Keywords: endometrial cancer, peroxisome proliferator-activated receptor gamma coactivator-1 alpha, estrogen-related receptor gamma, energy metabolism
Introduction
Endometrial cancer is a common cause of malignant tumors in the female reproductive tract; the incidence of endometrial cancer has increased in recent years, and endometrial cancer has become the most common female genital tract malignancy in some countries.1 Studies have revealed that there are two types of endometrial cancer, ie, estrogen-dependent (I endometrial cancer) and non-estrogen-dependent (II endometrial cancer), and that approximately 80% of clinical endometrial cancer patients have estrogen-dependent endometrial cancer.2,3 Estrogen and estrogen receptors (ERs) play roles in endometrial cancer that have been generally recognized in the doctrine of endometrial cancer, and ERs have become a biological marker of tumors with important clinical implications for guiding treatment.4,5 ER antagonism-based hormone therapy has become an effective adjuvant therapy for endometrial cancer, but this treatment is not effective for all ER-positive endometrial cancer patients; thus, a single receptor theory cannot explain all of the clinical signs of endometrial cancer. Signaling pathways other than the E-ER pathway that mediate the promotion or development of endometrial cancer might exist. Estrogen-related receptors (ERRs) are orphan nuclear receptor family members and include three subtypes: ERRα, ERRβ, and ERRγ. ERRs can bind the same target gene loci as does ERα; thus, in addition to ERs, ERRs might play an important role in the development of endometrial cancer.
The α subunit of peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α) is a recently identified transcription coactivator. This subunit can act on a variety of nuclear receptors, such as the peroxisome proliferator-activated receptor (PPAR), the thyroid hormone receptor, the ERR, the glucocorticoid receptor, hepatocyte nuclear factor 4α, and other transcription factors, including those of the nuclear respiratory factor pathway, to transmit the cell signal; thus, this subunit participates in energy metabolism in various tissues.6–9 Studies have found that, in metabolically active tissues, ERRγ is the main ligand of PGC-1α and regulates energy homeostasis via a complex interaction.10 The roles of abnormal energy metabolism in the pathogenesis of endometrial cancer are receiving increasing attention, but the exact mechanisms are not clear. The metabolism of cancer cells differs from the metabolic processes of normal tissue. The glycolysis pathway is the primary metabolic pathway for energy even under aerobic conditions. Pyruvate kinase (PK) is a key human anaerobic enzyme in the glycolysis pathway, and isocitrate dehydrogenase (IDH) is the rate-limiting enzyme in the Krebs cycle. The expressions and activities of these enzymes in tumor cells directly determine the supply of energy, which is essential for tumor cell proliferation.11
In this study, the role of PGC-1α in the malignant proliferation and energy metabolism of endometrial cancer was experimentally elucidated. We first observed the changes in the mRNA levels of PGC-1α and ERRγ in endometrial carcinomas, proliferative endometrial tissues, and normal endometrial tissues and subsequently detected the concentrations of PK and IDH. We found that the activities of PK and IDH were positively correlated with the expressions of PGC-1α and ERRγ. Thus, we suggest that PGC-1α might play an important role in the malignant proliferation and energy metabolism of endometrial cancer.
Materials and methods
Patients
From October 2011 to July 2013, a total of 40 cases of endometrial carcinoma (11 diabetic cases and 29 nondiabetic cases), including 15 cases with proliferative endometrial tissues and 21 cases with normal endometrial tissues, were collected from the Department of Obstetrics and Gynecology, Shengjing Hospital, China Medical University. This study protocol was approved by the Ethics Committee of China Medical University, and written informed consent was obtained from all subjects. Among the endometrial carcinoma cases, 10 were menopausal and 30 were premenopausal. The patients were grouped according to the International Federation of Gynecology and Obstetrics (FIGO) standard surgical staging as follows: 22 level I cases, 11 level II cases, and 7 level III cases. The gradings according to histological differentiations were as follows: 25 cases that were well differentiated, 9 cases that were moderately differentiated, and 6 cases that were poorly differentiated. The grouping according to myometrial invasion was as follows: 6 cases with no myometrial invasion, 25 cases with myometrial invasion ≤1/2, and 9 cases with myometrial invasion >1/2. The grouping according to lymphatic metastases was as follows: 7 cases with lymph node metastasis and 33 cases without lymph node metastasis.
RNA isolation and quantitative real-time PCR analysis
The tissues were flash frozen in liquid nitrogen immediately upon harvesting and stored at –80°C. Total RNA was extracted with commercially available Trizol reagent (TaKaRa, Takara Holdings, Kyoto, Japan). DNAse was added to eliminate the genome DNA. A 100 mg sample was used for total RNA isolation according to the manufacturer’s protocol. 1 mL Trizol was added to the homogenized sample for 5 minutes at room temperature. After adding 200 μL chloroform, the sample was incubated for 2–3 minutes at room temperature. Then the sample was centrifuged at 12,000× g for 15 minutes at 4°C. Approximately 1 mL isopropanol was added to the aqueous phase and incubated at room temperature for 10 minutes. Then the sample was again centrifuged at 12,000× g for 10 minutes at 4°C. After removing the supernatant, 1 mL of 75% ethanol was used to wash the RNA pellet. The sample was centrifuged at 12,000× g for 5 minutes at 4°C. After drying the RNA pellet in the air, it was resuspended in DEPC water. First-strand cDNA was synthesized using a PrimeScript RT reagent Kit (TaKaRa, Takara Holdings). The polymerase chain reaction (PCR) conditions were as follows: stage 1, 30 seconds at 95°C; stage 2, 40 cycles of 3 seconds of melting at 95°C, followed by DNA synthesis for 30 seconds at 60°C. The ABI Fast 7500 (Applied Biosystems, Foster City, CA, USA) was used. PGK-1 gene expression was quantified relative to the endogenous expression level of β-actin with real-time reverse transcription (RT)-PCR using the following primer sets and probes: for PGC-1α, forward primer, 5′-GACACAACACGGACAGAA-3′, reverse primer, 5′-CACAGGTATAACGGTAGGTAA-3′; for ERRγ, forward primer, 5′-GCCCTCACTACACTGTGTGAC-3′, reverse primer, 5′-CCTGCTAATTTGGACTGGTCTT-3′; and for β-actin, forward primer, 5′-TCGTCACCAACTGGGACGACATGG-3′, reverse primer, 5′-GATCTTGATCTTCATTGTGCT-3′. A negative control for each set of PCR reaction contained sterile water instead of the cDNA template. After amplification, real-time data acquisition and analysis were performed. The point at which the amplification plot crossed the threshold was defined as Ct.
ELISA
The PK and IDH concentrations in the tissues were measured using a sensitive enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer’s protocol (Cloud-Clone Corp., Houston, TX, USA).
Statistical analyses
Statistical comparisons were performed with one-way ANOVAs (analyses of variance), Student’s t-tests, and LSD tests. Correlation analyses were performed with Pearson’s correlation analyses. The data are presented as the means ± the standard errors (SEs). Statistical significance was defined by P-values less than 0.05. The statistical analyses were performed using the SPSS software (version 17.0).
Results
PGC-1α and ERRγ mRNA levels in the tissues
The PGC-1α and ERRγ mRNA levels in the different tissues were measured with quantitative real-time RT-PCR (Figure 1). Compared to the PGC-1α mRNA levels in the normal endometria, the PGC-1α mRNA levels in the endometrial carcinoma tissues and the hyperplasic endometrial tissues were elevated by 2.02±0.74-fold (P<0.01 vs the normal endometrium) and 1.63±0.74-fold (P<0.05 vs normal endometrium), respectively, and ERRγ was elevated to 2.15±0.60-fold (P<0.01 vs normal endometrium) and 1.81±0.74-fold (P<0.05 vs normal endometrium), respectively.
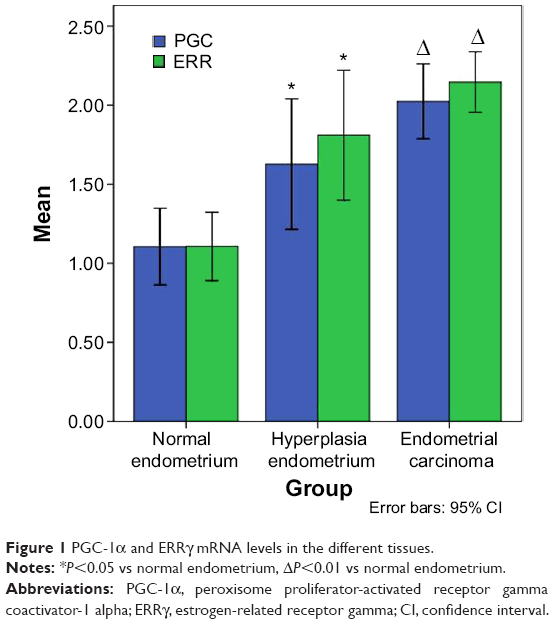 | Figure 1 PGC-1α and ERRγ mRNA levels in the different tissues. |
PK and IDH concentrations in the tissues
The PK and IDH concentrations in the endometrial carcinoma tissues were significantly higher than those in the normal endometria and hyperplasic endometrial tissues (Figure 2). The levels of PK and IDH in the endometrial carcinoma tissues were increased to 41.43±4.89 and 168.11±19.98 ng/mL, respectively, and these levels were significantly greater than those in the endometria (8.47±3.07, 143.87±18.83 ng/mL) and the hyperplasic endometria (36.75±4.29, 61.16±17.79 ng/mL; P<0.01 vs normal endometria and the hyperplasic endometria).
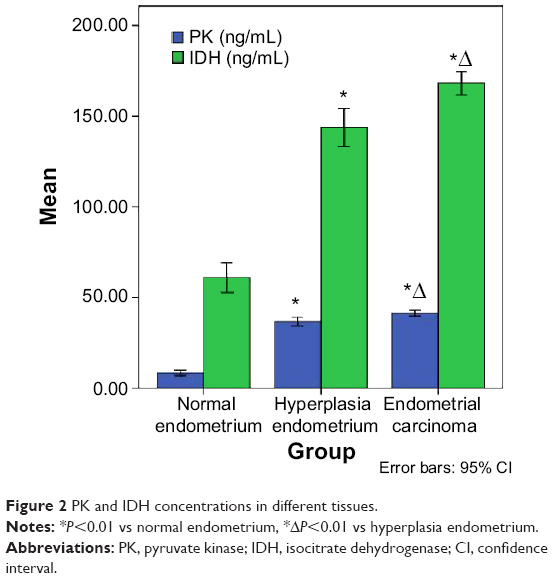 | Figure 2 PK and IDH concentrations in different tissues. |
PGC-1α and ERRγ mRNA levels in the endometrial carcinomas with different clinicopathological features
The PGC-1α and ERRγ mRNA levels in the endometrial carcinoma patients with type 2 diabetes were higher than those in the patients without diabetes. The PGC-1α and ERRγ mRNA levels in the endometrial adenocarcinomas increased with clinical staging, the depth of myometrial invasion, and increases in the number of metastatic lymph nodes. There were no significant correlations of PGC-1α or ERRγ mRNA levels with the degree of cell differentiation among the patients regardless of menopausal status (Table 1).
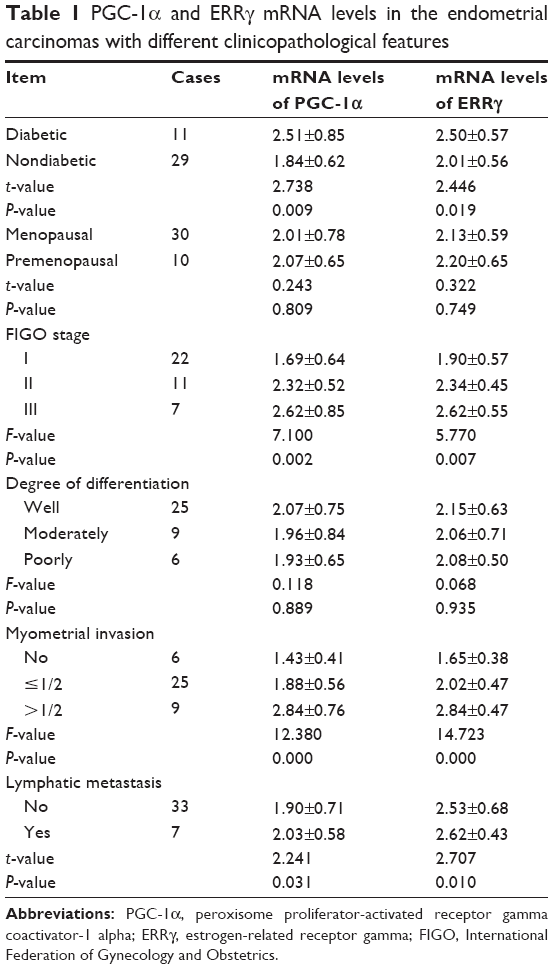 | Table 1 PGC-1α and ERRγ mRNA levels in the endometrial carcinomas with different clinicopathological features |
Correlation analyses of PGC-1α with ERRγ and their respective correlations with PK and IDH concentrations in the endometrial carcinomas
The PGC-1α mRNA levels were positively correlated with ERRγ in the endometrial carcinoma tissues (R=0.713, P<0.01). The PGC-1α mRNA levels were also positively correlated with the concentrations of PK and IDH in the endometrial carcinoma tissues (R=0.854 and 0.865, respectively, P<0.01), and similar results were found for ERRγ (R=0.713 and 0.738, respectively, P<0.01).
Discussion
The PGC-1 family consists of PGC-1α, PGC-1β, and PGC-1-associated factor (PRC). All three of these family members play important roles in many metabolic processes in the body. This family can regulate mitochondrial function as transcriptional activators, and roles for the members of this family in thermogenesis in brown fat have been identified.12 PGC-1α is a key factor in the regulation of mitochondrial metabolism. Studies have found that PGC-1α is overexpressed in endometrial carcinomas. Cormio detected the expression of PGC-1α with PCR and Western blots of 36 cases of type 1 endometrial adenocarcinoma and 16 cases of normal proliferative endometrial tissue, and this author also found that PGC1-α levels are increased in endometrial carcinomas when compared to those in normal proliferative tissues.6 In our study, we also found that PGC-1α mRNA levels in human endometrial carcinomas were significantly increased compared to those in hyperplasic endometria and normal endometria; moreover, the levels of PGC-1α were associated with the clinical features of endometrial cancer, which suggested that PGC-1α might be involved in the development of endometrial cancer.
ERRs are members of the orphan family of nuclear receptors and were recently identified. ERRs share homologies in the DNA-binding domain with ER that are as high as 68%.13 ERRγ is one of the ERRs and regulates the transcription of many genes that are involved in cell proliferation, differentiation, apoptosis, mitochondrial energy metabolism, and other processes. Our experiments revealed that the ERRγ expression in endometrial carcinomas was not only significantly greater than those in normal endometrial and hyperplasic endometrial tissues, but also positively correlated with the levels of PGC-1α in endometrial carcinomas, and both of them are related to the malignancy grade of endometrial cancer. In our study, the PGC-1α and ERRγ mRNA levels in the endometrial adenocarcinomas increased with clinical staging, the depth of myometrial invasion, and increases in the number of metastatic lymph nodes, but there were no significant correlations of the PGC-1α or ERRγ mRNA levels with the degree of cell differentiation among our patients regardless of menopausal status. These findings might suggest that PGC-1α and ERRγ play more important roles in the process of the appearance of tumors in endometrial cancer as opposed to invasion and metastasis. Connaughton et al14 found that the coactivator PGC-1α can work with ERRγ to promote the mRNA and protein expression of glucokinase. Recent studies have indicated that PGC-1α is involved in diabetes,15 which was known as an important risk factor for endometrial cancer. Similar to these results, our experiment revealed that the PGC-1α and ERRγ mRNA levels in the endometrial carcinoma patients with type 2 diabetes were higher than those in the patients without diabetes.
Currently, little research exists on the mechanism of the interaction between PGC-1α and ERRγ. PGC-1α is a key regulator of cellular energy metabolism. Rangwal et al16 found that ERRγ can be activated by PGC-1α; thus, ERRγ can regulate gene expression that is associated with energy metabolism via binding to PGC-1α.10,17 In our study, PGC-1α and ERRγ were significantly positively correlated in human endometrial carcinomas, which suggested that there is a relation between PGC-1α and ERRγa in the development of endometrial cancer.
The glycolysis pathway is the main metabolic energy pathway even in aerobic conditions in cancer. PK is a key human anaerobic enzyme in the glycolysis pathway, and IDH is a rate-limiting enzyme in the Krebs cycle. The expressions and activities of these enzymes in tumor cells directly determine the supply of energy, which is essential for tumor cell proliferation. Our experiments revealed that, in endometrial cancer tissues, the expressions of these two enzymes were significantly increased compared to those in normal endometria and hyperplasic endometria. PGC-1α plays important roles in the regulation of mitochondrial function and generation. We found that the expressions of PGC-1α and ERRγ in endometrial carcinomas were significantly increased and positively correlated with the concentrations of PK and IDH, which are closely related to the clinical features of endometrial carcinoma. These suggested that PGC-1α might be involved in the abnormal energy metabolisms of endometrial carcinoma. It is necessary to investigate how these factors interact with each other. Additionally, the function of other members of the PGC-1 family in endometrial cancer development should be elucidated. Therefore, we will further investigate these issues using an in vitro experimental endometrial carcinoma cell model, in our future studies.
Conclusion
In summary, our results suggested that the upregulation of PGC-1α and ERRγ in endometrial cancer might be a requirement for cancer cell energy metabolism, which contributes to the development of endometrial cancer.
Acknowledgments
The study was supported by the National Natural Science Foundation of China (82172874), National Natural Science Foundation of China (81472438), Shenyang City Science and Technology Bureau (F21-227-1-62), Science Research Foundation of Shengjing Hospital of China Medical University.
Author contributions
All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel R, Ma JM, Zou ZH, Jemal A. Cancer Statistics, 2014. CA Cancer J Clin. 2014;64(1):9–29. | ||
Bokhman JV. Two pathogenetic types of endometrial carcinoma. Gynecol Oncol. 1983;15(1):10–17. | ||
Hecht JL, Mutter GL. Molecular and pathologic aspects of endometrial carcinogenesis. J Clin Oncol. 2006; 24(29):4783–4791. | ||
Fournier DB, Chisamore M, Lurain JR, Rademaker AW, Jordan VC, Tonetti DA. Protein kinase C alpha expression is inversely related to ER status in endometrial carcinoma: possible role in AP-1-mediated proliferation of ER-negative endometrial cancer. Gynecol Oncol. 2001;81(3): 366–372. | ||
Henderson BE, Feigelson HS. Hormonal carcinogenesis. Carcinogenesis. 2000;21(3):427–433. | ||
Cormio A, Guerra F, Cormio G, et al. The PGC-1alpha-dependent pathway of mitochondrial biogenesis is upregulated in type I endometrial cancer. Biochem Biophys Res Commun. 2009;390(4):1182–1185. | ||
Ascenzi P, Bocedi A, Marino M. Structure-function relationship of estrogen receptor alpha and beta: impact on human health. Mol Aspects Med. 2006;27(4):299–402. | ||
Huppunen J, Aarnisalo P. Dimerization modulates the activity of the orphan nuclear receptor ERRgamma. Biochem Biophys Res Commun. 2004;314(4):964–970. | ||
Lanvin O, Bianco S, Vanacker JM. Estrogen-receptor-related receptors and hormone-dependent cancers. Adv Exp Med Biol. 2008;617:235–243. | ||
Giguère V. Transcriptional control of energy homeostasis by the estrogen-related receptors. Endocr Rev. 2008;29(6):677–696. | ||
Teicher BA, Linehan WM, Helman LJ. Targeting cancer metabolism. Clin Cancer Res. 2012;18(20):5537–5545. 12.Girnun GD. The diverse role of the PPARγ coactivator 1 family of transcriptional coactivators in cancer. Semin Cell Dev Biol. 2012;23(4): 381–388. | ||
Giguère V. To ERR in the estrogen pathway. Trends Endocrinol Metab. 2002;13(5):220–225. | ||
Connaughton S, Chowdhury F, Attia RR, et al. Regulation of pyruvate dehydrogenase kinase isoform 4 (PDK4) gene expression by glucocorticoids and insulin. Mol Cell Endocrinol. 2010;315(1–2):159–167. 15.Patti ME, Butte AJ, Crunkhorn S, et al. Coordinated reduction of genes of oxidative metabolism in humans with insulin resistance and diabetes: potential role of PGC1 and NRF1. Proc Natl Acad Sci U S A. 2003;100(14):8466–8471. | ||
Rangwala SM, Wang X, Calvo JA, et al. Estrogen-related receptor γ is a key regulator of muscle mitochondrial activity and oxidative capacity. J Biol Chem. 2010;285(29):22619–22629. | ||
17.Sun P, Wei L, Denkert C, Lichtenegger W, Sehouli J. The orphan nuclear receptors, estrogen receptor-related receptors: their role as new biomarkers in gynecological cancer. Anticancer Res. 2006;26(2C): 1699–1706. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.