")
Back to Journals » OncoTargets and Therapy » Volume 7
Isothermal multiple displacement amplification: a methodical approach enhancing molecular routine diagnostics of microcarcinomas and small biopsies
Authors Mairinger F, Walter R, Vollbrecht C, Hager T, Worm K, Ting S, Wohlschläger J, Zarogoulidis P, Zarogoulidis K, Schmid K
Received 29 March 2014
Accepted for publication 13 May 2014
Published 13 August 2014 Volume 2014:7 Pages 1441—1447
DOI https://doi.org/10.2147/OTT.S65144
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 7
Fabian D Mairinger,1 Robert FH Walter,2 Claudia Vollbrecht,3 Thomas Hager,1 Karl Worm,1 Saskia Ting,1 Jeremias Wohlschläger,1 Paul Zarogoulidis,4 Konstantinos Zarogoulidis,4 Kurt W Schmid1
1Institute of Pathology, 2Ruhrlandklinik, West German Lung Center, University Hospital Essen, Essen, 3Institute of Pathology, University Hospital Cologne, Cologne, Germany; 4Pulmonary Department, Oncology Unit, G Papanikolaou General Hospital, Aristotle University of Thessaloniki, Thessaloniki, Greece
Background and methods: Isothermal multiple displacement amplification (IMDA) can be a powerful tool in molecular routine diagnostics for homogeneous and sequence-independent whole-genome amplification of notably small tumor samples, eg, microcarcinomas and biopsies containing a small amount of tumor. Currently, this method is not well established in pathology laboratories. We designed a study to confirm the feasibility and convenience of this method for routine diagnostics with formalin-fixed, paraffin-embedded samples prepared by laser-capture microdissection.
Results: A total of 250 µg DNA (concentration 5 µg/µL) was generated by amplification over a period of 8 hours with a material input of approximately 25 cells, approximately equivalent to 175 pg of genomic DNA. In the generated DNA, a representation of all chromosomes could be shown and the presence of elected genes relevant for diagnosis in clinical samples could be proven. Mutational analysis of clinical samples could be performed without any difficulty and showed concordance with earlier diagnostic findings.
Conclusion: We established the feasibility and convenience of IMDA for routine diagnostics. We also showed that small amounts of DNA, which were not analyzable with current molecular methods, could be sufficient for a wide field of applications in molecular routine diagnostics when they are preamplified with IMDA.
Keywords: isothermal multiple displacement amplification, isothermal, whole-genome amplification, routine diagnostics, biopsies, microcarcinomas, microdissection
Introduction
The modern molecular diagnostic field is faced with the problem of an increasing spectrum of analysis from decreasing amounts of tissue, providing the opportunity for “tailored” therapy beneficial for the patient. The current approaches focus on collecting more material for the preferred molecular analysis, eg, by using cryobiopsies for evaluation of lymph node tissue in lung cancer.1–3 With respect to the patients health, a better way would be to improve the preamplification methods already in use to accumulate an adequate amount of DNA/RNA for all types of analysis required. Although many different types of preamplification methods have been published and evaluated, none of them have been accepted for routine diagnostics.4–17
In our opinion, a promising and powerful DNA preamplification method for this application is isothermal multiple displacement amplification (IMDA). This is an isothermal preamplification method that combines the advantages of non-thermostable DNA polymerases with a high yield reaction.10,11,14,18–21 Whole-genome amplification methods (WGA) using thermostable DNA polymerases may often generate non-specific amplification artifacts, resulting in an incomplete coverage of loci and/or generating very short DNA fragments.18 Compared with other whole-genome amplification methods, IMDA, reported for the first time by Dean et al in 2002,18 shows a highly uniform coverage across the genome and generates high molecular weight genomic DNA (up to >70 kb in length). This method is already validated for clinical specimens10,11 as well as for formalin-fixed paraffin-embedded (FFPE) tissue.5,10 In microbiology, IMDA has become state of the art as a prepolymerase chain reaction (PCR) method to amplify difficult samples and low copy number sequences.22 It is sufficient to start with an input of only a few laser-capture microdissected cells for whole-genome amplification to generate an adequate amount of DNA for further molecular analysis.
The aim of this study was to confirm the convenience and feasibility of IMDA for routine molecular diagnostics using very small tumor samples, including small biopsies with a low number of malignant cells or microcarcinomas.
Materials and methods
Study design
To confirm the feasibility of IMDA for routine molecular pathological diagnostics, different FFPE lung tissue samples were microdissected and subsequently preamplified by IMDA. For pretesting of amplification efficiency, only benign tissue was used. To test the homogeneity of the amplification at different chromosomal loci, 33 targets distributed all over the human genome (at least one on every chromosome) were tested by SYBR-green (Life Technologies, Carlsbad, CA, USA) quantitative PCR analysis. Afterwards, human lung adenocarcinoma samples were resequenced after initial DNA preamplification using IMDA for known EGFR exon 21 c.2573T>G (p.L858R) point mutations by Sanger sequencing. Well known activating EGFR exon 19 deletions were determined using agarose gel electrophoresis. The known EGFR exon 19 deletions can be clearly separated by this method. In addition, the same tumor samples were sequenced by pyrosequencing for possible ERCC1 exon 4 c.19007T>C (p.N118N) and 3′-UTR c.8092C>A polymorphisms, respectively.
Samples and patients
Only FFPE tissue was used. FFPE preparation was performed according to the institutional standards. The fresh tissue was fixed in 4% buffered formalin for 24 hours, then diced to approximately 0.5 × 1.0 × 1.0 cm dimensions and embedded in paraffin. For diagnostic classification, one to four, <1 μm thick sections were used for immunohistochemistry and staining with hematoxylin and eosin.
For evaluation of IMDA, ten benign lung tissue samples were selected for pretesting. Additionally, for sequence analysis, four lung cancer specimens were selected for evaluation. One sample contained a known deletion in exon 19 of the EGFR gene and another sample contained a known p.L858R point mutation in exon 21 of the EGFR gene. The remaining two tumor samples included in the analysis show a wild-type EGFR gene locus. Cells for subsequent IMDA were separated from inflammation and leukocytes by microdissection.
Microdissection
Sample slides were microdissected using laser-capture microdissection technology (PALM RoboMover; Zeiss, Jena, Germany). Zeiss MembraneSlide NF 1.0 PEN glass slides were used due to the plastic film affixed on the slides. The Robo-LPC computer program was used to assist the microdissection procedure.
For tissue lysis prior to IMDA amplification, Qiagen FFPE lysis solution (Qiagen, Hilden, Germany), which is applied in the REPLI-g FFPE kit (Qiagen), was used; 20 μL was directly dropped into the cap of the microdissection tube. The microdissected tumor areas had dimensions in the range of 50 × 50 μm to 150 × 100 μm correlating with an average of approximately 25 tumor cell nuclei.
IMDA preamplification
The commercial REPLI-g FFPE kit (Qiagen) was used for IMDA. This assay combines a ligase reaction to generate high molecular weight DNA templates from degraded FFPE tissue with IMDA DNA preamplification. Genomic DNA from the lysed tissue was denatured at room temperature for 3 minutes and then incubated with random primers. Strand displacement synthesis was performed at 30°C according to the manufacturer’s recommendations. As the amplification reaction reaches a plateau after approximately 8 hours, the reaction was terminated by incubation at 60°C for 10 minutes.
DNA quantification was done using a Qubit dsDNA BR assay kit (Invitrogen, Carlsbad, CA, USA).
Amplification products were stored at 4°C until further use.
Analysis of chromosomal coverage
To test for the presence of all chromosomes/target sequences in the amplified DNA, randomly chosen genes spanning regions on every chromosome were quantitatively tested using Platinum SYBR Green quantitative PCR SuperMix-UDG (Invitrogen). In addition, on four chromosomes, multiple gene loci were amplified to test the relative representation of different regions on these chromosomes. Quantitative analysis was performed on a 7500 fast real-time PCR system (Applied Biosystems, Foster City, CA, USA). The 2^-ΔCt method was used to normalize target Ct’s versus reference Ct’s. To ensure amplification of the probably highly degraded genomic DNA from the FFPE tissue, amplicons for quantitative PCR reaction were designed with a maximum length of 100 bp. All primer sequences are listed in Table 1, and the PCR protocol is shown in Table 2.
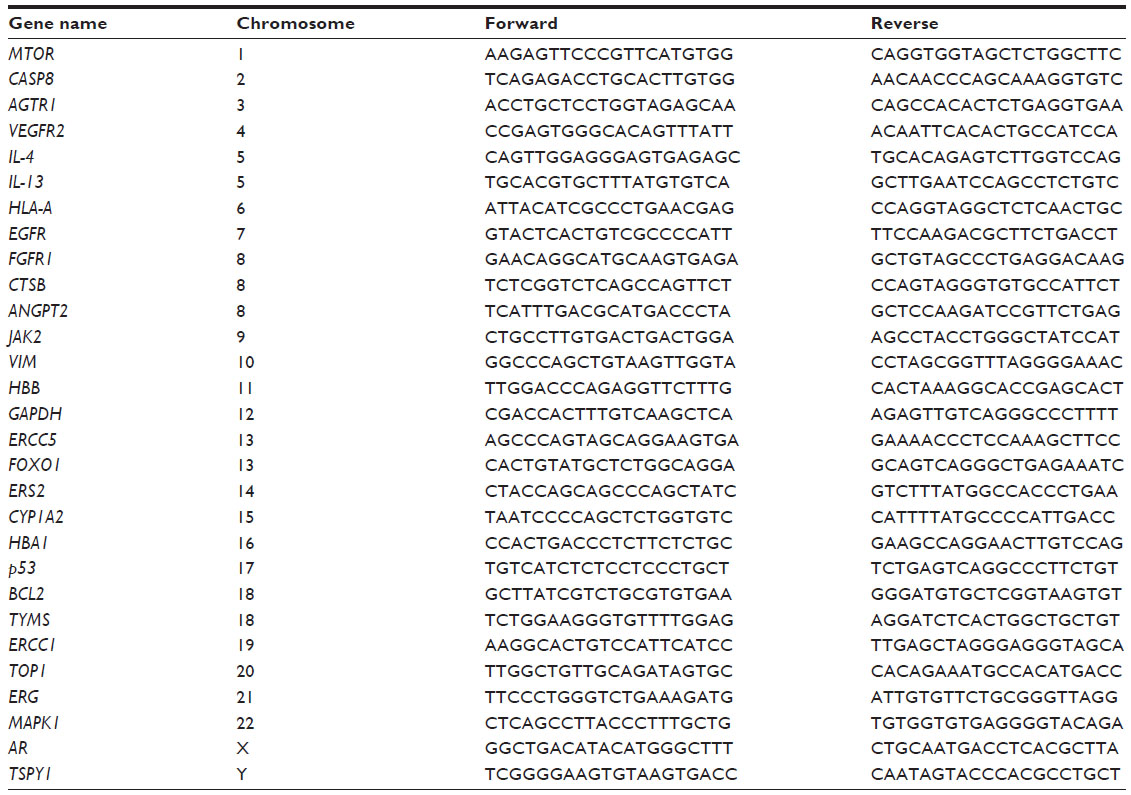 | Table 1 List of all primer sequences used for amplification for relative chromosomal representation analysis |
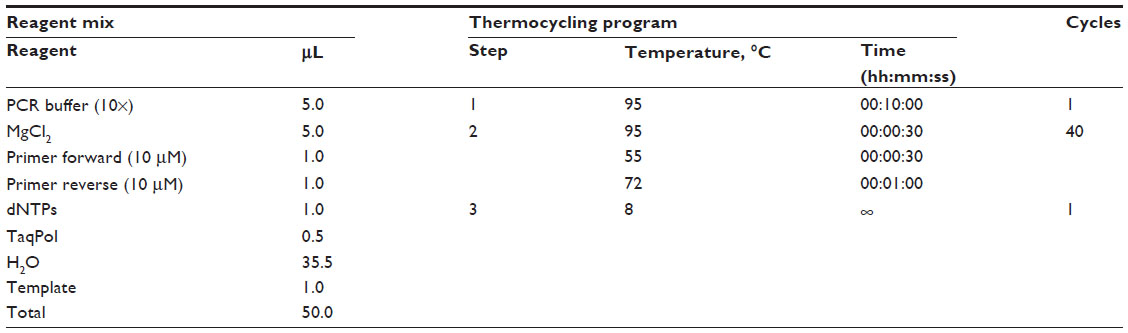 | Table 2 Details regarding the PCR conditions |
Mutational analysis
Sanger sequencing was performed on an Applied Biosystems 3500 genetic analyzer according to the manufacturer’s protocol (BigDye Terminator v3.1 cycle sequencing kit; Applied Biosystems) with a halved reaction volume. Pyrosequencing was performed on a PyroMark Q24 (Qiagen). For data analysis, the Qiagen PyroMark Q24 software was used. Sequencing was done according to the Qiagen PyroMark Q24 protocol. Streptavidin sepharose (GE Healthcare, Chalfont, UK) was used instead of streptavidin-coated beads. Sequences of all sequencing primers are listed in Table 3.
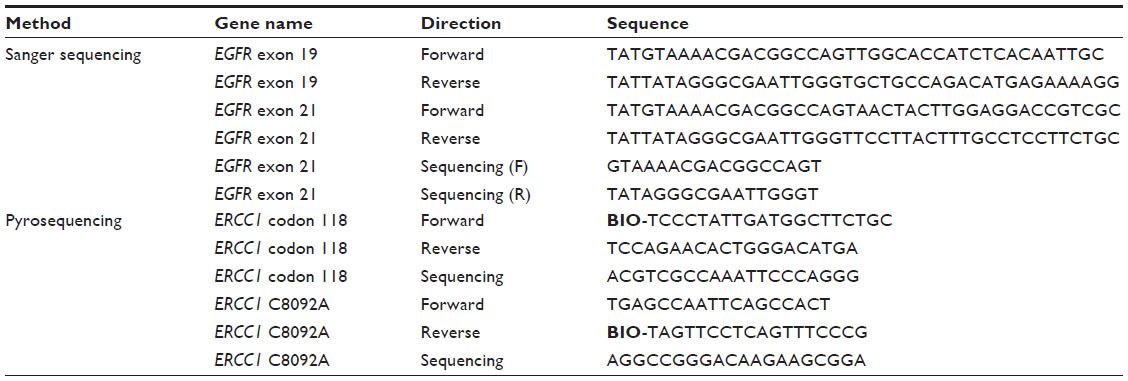 | Table 3 List of primers used for pyrosequencing and amplification |
Results
IMDA preamplification
IMDA was successful in all (malignant and benign) samples. With an input of approximately 20–25 cells (equaling approximately 175 pg of genomic DNA) for laser-capture microdissection, 3 μg/μL up to more than 5 μg/μL DNA was generated with a reaction volume of 50 μL. The actual amount of suitable DNA amplified strongly depends on the duration of the final amplification step.
The best results were obtained with an amplification duration of 8 hours. At this point, all quantitative PCR analyses showed sufficient results; nevertheless the amplification efficiency showed small disparities. The age of the tissue slides, did not show an influence on the qualtity of the quantitative PCR template. All negative controls showed no results.
Chromosomal coverage
All tested genes on all chromosomes could be detected. However, the detected gene loci differed in their amplification (Figure 1). Both the chi-squared test and Fisher’s Exact test showed a significant independency for amplification (P equals nearly 1). Also, within the chromosomes, there were clear differences (Figure 1). Differences in amplification efficiency between the analyzed gene loci ranged up to 3.8-fold between the median 2^-ΔCt values of each locus.
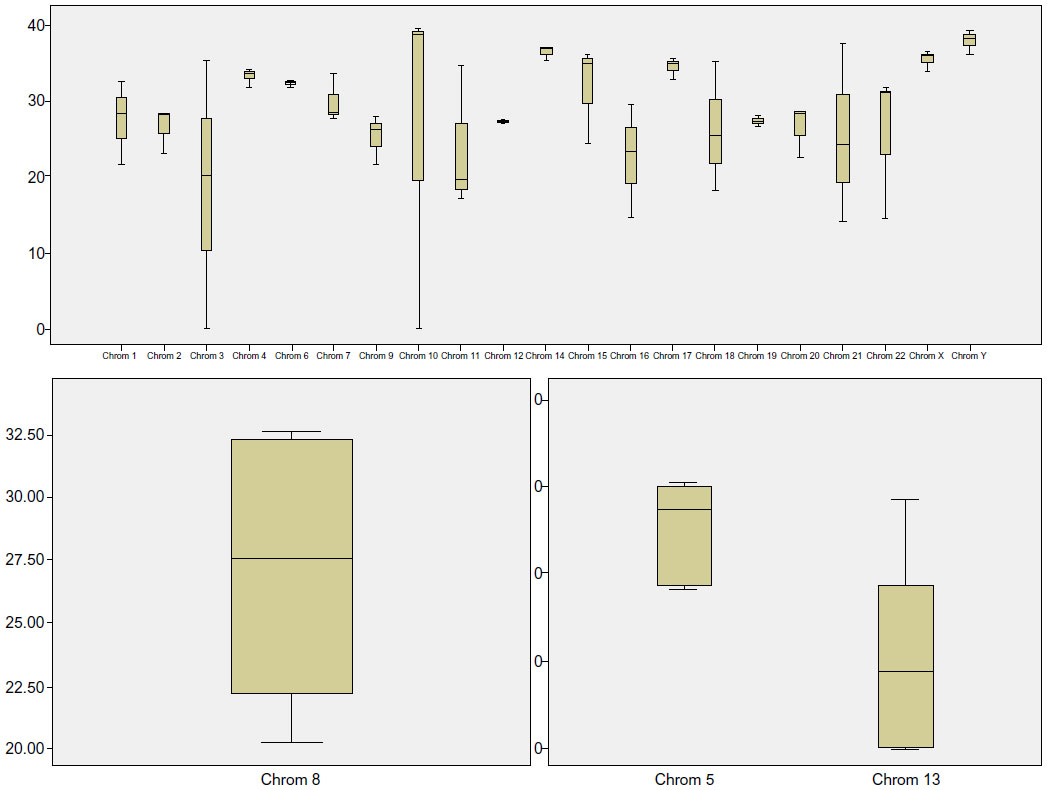 | Figure 1 Amplification strength of tested genes normalized to GAPDH and displayed by 2^-deltaCt values. Six genes show an extraordinary deviation (VIM, FOXO1, HBA1, AR, MAPK1, TSPY1), but most of them have a lower amplification rate (except TSPY1). The variances can be explained by the difference in amplification product length and the quality of the template before ligation. |
The amplified sequence seems not to have a major impact on this phenomenon; it rather depends on the quality of DNA in the tissue slides being investigated than on the parameters of the target sequence. Most of the amplified DNA fragments were of a small size. This effect becomes increased by the ligation step, which links together several fragments of distinct genes/regions. Small-sized DNA fragments like these “patchwork fragments” would still be long enough for next-generation sequencing, and the merging of different gene sequences is not problematic, but also for normal “qualitative” analysis, like mutational analysis, the amount of useful amplified DNA is adequate.
Mutational analysis
After an initial IMDA preamplification, two specimens were tested for known EGFR mutations (exon 19 deletions and exon 21 p.L858R point mutations) by capillary sequencing. Both mutations could be detected reproducibly in all replications (Figure 2). In two patient samples that contained the EGFR wild-type sequence and served as negative controls, no mutations could be detected after IMDA preamplification.
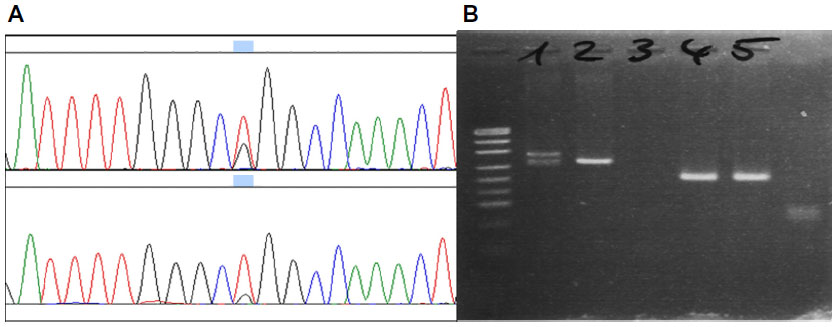 | Figure 2 Chromatograms of Sanger sequencing for EGFR mutations in clinical specimens (A) exon 21 p.L858R wild mutated sample (B) exon 19 deletion in the agarose gel (1) compared with an exon 19 wild-type (2), a non-template control (3), as well as amplicons for EGFR exon 21 sequencing (4 and 5). |
The same four specimens were used for pyrosequencing of ERCC1 point mutations at the c.19007T>C and c.8092C>A mutation hotspot of ERCC1. Sequencing was successful in all four cases. One case showed an ERCC1 exon 4 c.19007T>C, whereas the others were classified as wild-type (Figure 3). For ERCC1 c.8092C>A mutation status, all cases showed a wild-type genotype in comparison with a mutated positive control.
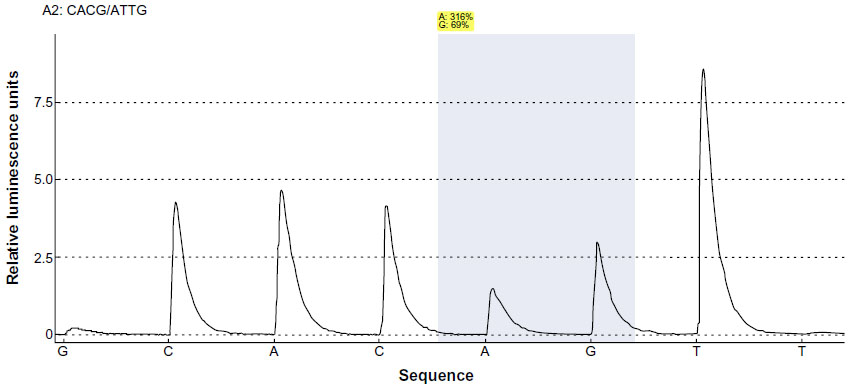 | Figure 3 Pyrosequencing flowgram showing an ERCC1 exon 4 c.19007T>C heterogeneous genotype (p.N118N). The first injected base (G) was used to identify the background, and the last injected base (T) was used for determination of phasing effects. The shaded area marks the position analyzed for possible variants. |
Discussion
The aim of our study was to check the feasibility and convenience of IMDA as an isothermal preamplification method for molecular diagnostics involving notably small sample material.
Our results show outstanding amplification efficiency (250 μg of genomic DNA for approximately 25 cells) and that IMDA is a valuable preamplification method for all further molecular analysis of these clinical specimens. In contrast, recent studies show a five- to ten-fold lower efficiency of IMDA, but these deviances may be based on differences between the quantification methods used.18 The additional preceding ligation step to generate DNA templates of higher genomic weight could also be a possible explanation for this phenomenon.
All 33 investigated genes could be amplified and detected by subsequent analysis, although the amplification efficiency was heterogeneous between the tested gene loci, ranging up to 3.8-fold between each locus.
Although a reaction time of 3 hours generated a feasible amount of DNA, the average fragment size seemed to be too short for efficient use as a template for further PCR reactions, even with an amplicon size ranging from 100 bp to 200 bp. In contrast with that, published data for subsequent whole-genome sequencing suggests that this relatively small amplicon size may be sufficient.8,10,14,22 The published range bias generated with whole-genome amplification methods lies between six-fold and eight-fold.8,10,14,18,22 In comparison with that, the methodical bias of IMDA, even for FFPE tissue, is, significantly below.5,10
The relatively high bias in our experiment can be explained by the thickness of the tissue slides (1–4 μm); during microdissection, nucleoli may be fragmented, leading to a loss of genetic loci in individual cells. Moreover, all diagnostically relevant genes and mutations were detected after IMDA preamplification. Subsequent sequencing gave evaluable results using two independent sequencing methods (capillary sequencing and pyrosequencing).
New challenges in the upcoming years will require new molecular biological techniques that are currently not established in routine pathology laboratories. Therefore, isothermal amplification methods, originally derived from microbiological work-up processes,12,22–24 are merely examples of the potential new techniques for modern pathology that can be derived from other fields of research. In this context, the outstanding role of isothermal amplification methods has to be mentioned. Additionally, it is of note that non-thermostable DNA polymerases avoid artificial variants because of their proofreading ability and simultaneously enable the advantages of non-thermostable DNA polymerases.4,6,11,13,15–19,21 Nevertheless, IMDA is not suitable for rapid on-site evaluation during the biopsy process because of the long reaction time. Molecular diagnostics is an evolving field in routine pathology. Therefore, the development and integration of feasible, sensitive, robust and efficient techniques like IMDA is mandatory.
Conclusion
With a reaction time of 8 hours, a feasible amount of amplicons with adequate size for qualitative PCR can be generated using IMDA, starting with input material equaling 25 human cells. Further, tissue slides can be stained with hematoxylin and eosin for microdissection, making it much easier for the pathologist/technical assistant to find tumor cells. Additionally, this method also works with macrodissected (microscope and scalpel) tissue of a surface extension less than 1 mm3. For larger tissue samples, routine amplification without IMDA remains the first choice, due to the relatively high costs and additional processing time of IMDA. However, it needs to be borne in mind that every additional processing step promotes more or less bias. Nevertheless, IMDA has proven to be a feasible and robust molecular pathological method to preamplify hitherto hardly analyzable clinical specimens.
Disclosure
The authors report no conflicts of interest in this work.
References
Hetzel J, Eberhardt R, Herth FJ, et al. Cryobiopsy increases the diagnostic yield of endobronchial biopsy: a multicentre trial. Eur Respir J. 2012;39(3):685–690. | |
Aktas Z, Gunay E, Hoca NT, et al. Endobronchial cryobiopsy or forceps biopsy for lung cancer diagnosis. Ann Thorac Med. 2010;5(4):242–246. | |
Schumann C, Hetzel J, Babiak AJ, et al. Cryoprobe biopsy increases the diagnostic yield in endobronchial tumor lesions. J Thorac Cardiovasc Surg. 2010;140(2):417–421. | |
Guatelli JC, Whitfield KM, Kwoh DY, Barringer KJ, Richman DD, Gingeras TR. Isothermal, in vitro amplification of nucleic acids by a multienzyme reaction modeled after retroviral replication. Proc Natl Acad Sci U S A. 1990;87(19):7797. | |
Aviel-Ronen S, Qi Zhu C, Coe BP, et al. Large fragment Bst DNA polymerase for whole genome amplification of DNA from formalin-fixed paraffin-embedded tissues. BMC Genomics. 2006;7:312. | |
Fire A, Xu SQ. Rolling replication of short DNA circles. Proc Natl Acad Sci U S A. 1995;92(10):4641–4645. | |
Gingeras TR, Richman DD, Kwoh DY, Guatelli JC. Methodologies for in vitro nucleic acid amplification and their applications. Vet Microbiol. 1990;24(3–4):235–251. | |
Lage JM, Leamon JH, Pejovic T, et al. Whole genome analysis of genetic alterations in small DNA samples using hyperbranched strand displacement amplification and array-CGH. Genome Res. 2003;13(2):294–307. | |
Lee CI, Leong SH, Png AE, et al. An isothermal primer extension method for whole genome amplification of fresh and degraded DNA: applications in comparative genomic hybridization, genotyping and mutation screening. Nat Protoc. 2006;1(5):2185–2194. | |
Lee CI, Leong SH, Png AE, et al. An isothermal method for whole genome amplification of fresh and degraded DNA for comparative genomic hybridization, genotyping and mutation detection. DNA Res. 2006;13(2):77–88. | |
Luthra R, Medeiros LJ. Isothermal multiple displacement amplification: a highly reliable approach for generating unlimited high molecular weight genomic DNA from clinical specimens. J Mol Diagn. 2004;6(3):236–242. | |
Nelson JR, Cai YC, Giesler TL, et al. TempliPhi, phi29 DNA polymerase based rolling circle amplification of templates for DNA sequencing. Biotechniques. 2002;Suppl:44–47. | |
Notomi T, Okayama H, Masubuchi H, et al. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000;28(12):E63. | |
Rook MS, Delach SM, Deyneko G, Worlock A, Wolfe JL. Whole genome amplification of DNA from laser capture-microdissected tissue for high-throughput single nucleotide polymorphism and short tandem repeat genotyping. Am J Pathol. 2004;164(1):23–33. | |
Vincent M, Xu Y, Kong H. Helicase-dependent isothermal DNA amplification. EMBO Rep. 2004;5(8):795–800. | |
Yi J, Zhang W, Zhang DY. Molecular Zipper: a fluorescent probe for real-time isothermal DNA amplification. Nucleic Acids Res. 2006;34(11):e81. | |
Zhang DY, Brandwein M, Hsuih TC, et al. Amplification of target- specific, ligation-dependent circular probe. Gene. 1998;211(2):277–285. | |
Dean FB, Hosono S, Fang L, et al. Comprehensive human genome amplification using multiple displacement amplification. Proc Natl Acad Sci U S A. 2002;99(8):5261–5266. | |
Gadkar VJ, Arseneault T, Filion M. Fidelity and representativeness of two isothermal multiple displacement amplification systems to preamplify limiting amounts of total RNA. Mol Biotechnol. 2014;56(4):377–385. | |
Kaewmanee M, Phoksawat W, Romphruk A, Romphruk AV, Jumnainsong A, Leelayuwat C. Development of a multiplex polymerase chain reaction-sequence-specific primer method for NKG2D and NKG2F single-nucleotide polymorphism typing using isothermal multiple displacement amplification products. Tissue Antigens. 2013;81(6):419–427. | |
Gill P, Ghaemi A. Nucleic acid isothermal amplification technologies: a review. Nucleosides Nucleotides Nucleic Acids. 2008;27(3):224–243. | |
Gonzalez JM, Portillo MC, Saiz-Jimenez C. Multiple displacement amplification as a pre-polymerase chain reaction (pre-PCR) to process difficult to amplify samples and low copy number sequences from natural environments. Environ Microbiol. 2005;7(7):1024–1028. | |
Kim KH, Chang HW, Nam YD, et al. Amplification of uncultured single-stranded DNA viruses from rice paddy soil. Appl Environ Microbiol. 2008;74(19):5975–5985. | |
Groathouse NA, Brown SE, Knudson DL, Brennan PJ, Slayden RA. Isothermal amplification and molecular typing of the obligate intracellular pathogen Mycobacterium leprae isolated from tissues of unknown origins. J Clin Microbiol. 2006;44(4):1502–1508. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.