")
Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 7
Effects of EPHX1 and CYP3A4 polymorphisms on carbamazepine metabolism in epileptic patients
Authors Caruso A, Bellia C, Pivetti A, Agnello L, Bazza F, Scazzone C, Bivona G, Lo Sasso B, Ciaccio M
Received 8 October 2013
Accepted for publication 28 November 2013
Published 2 April 2014 Volume 2014:7 Pages 117—120
DOI https://doi.org/10.2147/PGPM.S55548
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Antonietta Caruso, Chiara Bellia, Alessia Pivetti, Luisa Agnello, Federica Bazza, Concetta Scazzone, Giulia Bivona, Bruna Lo Sasso, Marcello Ciaccio
Department of Biopathology and Medical and Forensic Biotechnologies, University of Palermo, Palermo, Italy
Background: The aim of this study was to investigate the effect of two genetic polymorphisms in the coding regions (exon 3 and exon 4) of the EPHX1 gene, ie, 337T>C and 416A>G, respectively, on the metabolism of carbamazepine (CBZ) 10,11-epoxide (the active metabolite of CBZ) by evaluating the variation in serum CBZ 10,11-epoxide levels 4 hours after administration of the drug. Moreover, we reported the genotype frequencies of the CYP3A4*22 (rs 35599367, C>T) variant and its influence on the metabolism of CBZ.
Methods: The analysis was performed in 50 patients receiving CBZ as monotherapy. DNA was extracted from leukocytes using a commercially available kit. Serum CBZ 10,11-epoxide levels were measured by high-performance liquid chromatography. Allelic discrimination was performed using polymerase chain reaction-restriction fragment length polymorphism. Statistical analysis of the difference in mean values for CBZ 10,11-epoxide levels according to genotype was performed using the Student's t-test with Statistical Package for the Social Sciences version 13 software.
Results: Fourteen percent of the study group were CC, 42% were CT, and 44% were TT for the EPHX1 337T>C variant. No GG homozygote was identified for the EPHX1 416A.G variant; 64% were AA and 36% were AG. When we compared serum CBZ 10,11-epoxide levels 4 hours after drug administration, we found no statistically significant difference between the 337 CC, CT, and TT genotypes. Similarly, no difference in serum CBZ 10,11-epoxide levels was found between 416A>G AA and AG. Genotype frequencies for the CYP3A4*22 (rs 35599367 C>T) allelic variant were 94% for CC and 6% for CT, with no statistically significant difference in serum CBZ 10,11-epoxide levels between these genotypes 4 hours after administration of the drug (2.6±1.3 µg/µL and 2.5±1.2 µg/µL, respectively).
Conclusion: Although there is some evidence of involvement of these polymorphisms in enzyme activity in vitro, we found no interference with CBZ metabolism in vivo.
Keywords: EPHX1 gene, CBZ 10,11-epoxide, CYP3A4*22, drug metabolism
Introduction
Epilepsy is the most common chronic neurological disorder and is characterized by recurrent unprovoked epileptic seizures. It has a prevalence of 5–10 per 1,000 and an incidence of 50–120 per 100,000 per year.1 Administration of antiepileptic drugs fails to control epileptic seizures in 70% of patients. Carbamazepine (CBZ) is one of the most commonly used anticonvulsants in the treatment of epilepsy. Two basic mechanisms of action have been proposed, ie, enhancement of sodium channel inactivation by reducing high-frequency repetitive firing of action potentials and an effect on synaptic transmission.2 CBZ is extensively metabolized in the liver, with less than 5% of an oral dose excreted unmodified in the urine. The enzymes involved in the primary metabolism of CBZ belong to the superfamily of cytochrome P450 (CYP) responsible for formation of reactive epoxides. Isoenzyme CYP3A4, the major form of CYP in the adult liver and responsible for the metabolism of most drugs (45%–60%), is involved in the formation of CBZ 10,11-epoxide, the active metabolite of CBZ. The latter is converted to the inactive water-soluble metabolite (CBZ 10,11-diol) by microsomal epoxide hydrolase, which is encoded by the EPHX1 gene. CBZ 10,11-diol is excreted in the urine.3
The aim of the present study was to investigate the effect of two genetic polymorphisms in the coding regions (exon 3 and exon 4) of the EPHX1 gene on the metabolism of CBZ 10,11-epoxide.4 These polymorphisms were carefully selected from previous reports in the literature and databases such as the dbSNP.5,6 We also evaluated the variation in serum CBZ 10,11-epoxide levels 4 hours after administration of the drug. Finally, we analyzed the genotypic frequencies of the CYP3A4*22 (rs 35599367, C>T) variant and its influence on serum levels of the CBZ metabolite.
Materials and methods
The patient characteristics are described in Table 1. Thirty-two of the 50 patients included in this study were male, the mean age of the group was 41±14 years, and all were from South Italy. All of the patients had partial or generalized tonic-clonic epileptic seizures. Blood samples for measurement of serum CBZ and metabolite levels, and molecular analysis were obtained from patients who were on maintenance monotherapy with CBZ. The dosage was established on the basis of clinical features and frequency of seizures. All patients had reached steady-state, and the average CBZ dose was 13.5±7.4 mg/kg/day. None of the patients were being treated with other drugs (antiepileptic or nonantiepileptic) that could interfere with the metabolism of CBZ, and none reported significant ingestion of food known to interact with CBZ. The study was approved by the Ethics Committee of the Department of Biopathology and Medial and Forensic Biotechnologies and all patients gave their written informed consent.
 | Table 1 Characteristics of patients on CBZ monotherapy, with serum CBZ/CBZ 10,11-epoxide ratio indicated in the basal state and 4 hours after administration of the drug |
Serum CBZ and CBZ metabolite measurement, which is performed routinely for therapeutic drug monitoring in patients with epilepsy, was performed by high-performance liquid chromatography. Separation was obtained by isocratic elution with ultraviolet detection at 220 nm. The mobile phase was 63% phosphate buffer, 32% acetonitrile, 3.5% methyl alcohol, and 1.5% tetrahydrofuran. The limit of detection was 0.5 μg/mL for CBZ, the intra-assay coefficient of variation was <3%, and the interassay coefficient of variation was <5%.
Genomic DNA was extracted from leukocytes using a commercial kit (Roche Diagnostics Corporation, Indianapolis, IN, USA). Allelic discrimination for EPHX1 337T>C, EPHX1 416A>G, and the CYP3A4*22 (rs 35599367 C>T) variant was carried out by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP).7,8 For PCR amplification, we used the following primers: forward 5’-GATCGATAAGTTCCGTTTCACC-3’ and reverse 5’-CTTAGTCTTGAAGTGAGGAT-3’ for identification of EPHX1 337T>C; forward 5’-ACATCCACTTCATCCACGT-3’ and reverse 5’-ATGCCTCTGAGAAGCCAT-3’ for identification of EPHX1 416A>G; and forward 5’-CTGAAGAGGAATCGGCTCTG-3’ and reverse 5’-TTTTTACCATCCTTCCTCTATGC-3’ for identification of the allelic variant CYP3A4*22 (rs 35599367 C>T). The PCR mixture was performed in a total volume of 50 μL, containing 150 ng DNA, 0.2 mM of each deoxynucleotide triphosphate; 25 nmol of each primer, 5 μL of 10 × reaction buffer, and 1.5 mM MgCl2, with two units of Taq polymerase for EPHX1 337T>C and EPHX1 416A>G, and 1.5 units for CYP3A4*22. The PCR conditions were 94°C for 3 minutes, 35 cycles of 94°C for 30 seconds, 55°C for 25 seconds, 72°C for 20 seconds, and a final extension of 72°C for 5 minutes. This protocol was used for both allelic variants of the EPHX1 gene. CYP3A4*22 was analyzed using the following protocol: 94°C for 3 minutes, 35 cycles of 94°C for one minute, 55°C for one minute, 72°C for one minute, and a final extension of 72°C for 5 minutes. The amplified PCR products were digested with 10 U EcoRV for EPHX1 337T>C, 5 U RSAI for EPHX1 416A>G, and 6 U Dra III for CYP3A4*22 (rs 35599367, C>T) and overnight incubation at 37°C. The RFLP analysis was performed on 3% agarose gel and confirmed by direct sequencing. Statistical analysis of the difference in mean CBZ 10,11-epoxide levels between the genotypes was done with the Student’s t-test using Statistical Package for the Social Sciences version 13 software (SPSS Inc., Chicago, IL, USA). Differences in adjusted basal CBZ 10,11-epoxide levels between the genotypes were tested for using the Kruskal–Wallis test.
Results
Fourteen percent of the study group were CC, 42% were CT, and 44% were TT for the EPHX1 337T>C variant, and the frequency of the C allele was 0.35 (Table 2B); none were homozygous for the EPHX1 416A>G variant, with 64% and 36% being AA and AG, respectively (Table 2A). The frequency of the G allele was 0.18. We compared the distribution of genotype frequencies with that of a representative control population used in previous studies in our laboratory and found no differences (data not shown). Patient characteristics were not different between the EPHX1 and CYP3A4 genotypes when we performed an analysis for daily CBZ dose, serum CBZ and CBZ 10,11-epoxide levels in the basal state and 4 hours after ingestion of the drug.
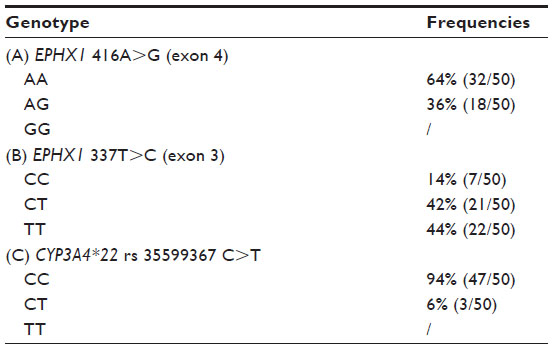 | Table 2 Genotype frequencies for EPHX1 416A>G (A), EPHX1 337T>C (B), and CYP3A4*22 rs 35599367 (C) in epileptic patients on carbamazepine therapy |
When we compared the mean values for CBZ 10,11-epoxide at 4 hours between 337 CC (1.9±0.84 μg/μL), 337 CT (2.9±1.3 μg/μL), and 337 TT (2.5±1.4 μg/μL), we found no statistically significant differences (P>0.05). The difference was also not statistically significant between AA (2.7±1.4 μg/μL) and AG (2.4±1.2 μg/μL) for the EPHX1 416A>G variant. Moreover, we compared basal serum CBZ 10,11-epoxide levels adjusted for the daily dose between EPHX1 337C>T and 416A>G genotypes to evaluate whether these variants can modulate serum levels of the active form of the drug, but no statistically significant differences were found (P=0.23 for EPHX1 377T>C and P=0.71 for EPHX1 416A>G). The combined effect of the two EPHX1 polymorphisms on serum concentrations of CBZ, CBZ 10,11-epoxide, and the corresponding value adjusted for daily dose was evaluated, but the analysis revealed no statistically significant differences (P=0.63).
The genotype frequencies were 94% CC and 6% CT for the CYP3A4*22 (rs 35599367, C>T) variant (Table 2C). The T allele frequency was 0.03. As for the EPHX1 variants, we found no differences in genotype distribution between the study group and the control group (data not shown). No statistically significant differences were found comparing serum CBZ 10,11-epoxide levels between the genotype CC (2.6±1.3 μg/μL) and genotype CT (2.5±1.2 μg/μL) groups 4 hours after administration of the drug.
Discussion
Epilepsy is a common neurological disease characterized by recurrent seizures. The therapeutic goal is to maximize seizure control while minimizing adverse drug effects, thus preserving the patient’s quality of life. However, despite the introduction of a number of new antiepileptic drugs during recent years, about one third of the epilepsy population continues to show drug-resistance.9,10 It has been shown that the variability in individual response to antiepileptic drugs depends not only on environmental factors such as diet and smoking, but also on age, sex, weight, liver and renal function, disease heterogeneity, and genetic background.11 Genetic background can predict drug efficacy and toxicity, so the effect of genetic polymorphisms on the metabolism of drugs is significant.
In the present study, we investigated the two best known polymorphisms of the EPHX1 gene coding for microsomal epoxide hydrolase, ie, 337T>C within exon 3 (resulting in substitution of Tyr with His at amino acid position 113) and 416A>G within exon 4 (which codes for substitution of His with Arg at amino acid position 139).
Our results demonstrate that EPHX1 and CYP3A4 polymorphisms do not affect serum CBZ 10,11-epoxide levels. To our knowledge, involvement of these variants in the metabolism of CBZ has not been well investigated, and there are conflicting reports in the literature. Makmor-Bakry et al demonstrated that EPHX1 337C>T and 416A>G polymorphisms were associated with the maintenance dose of CBZ, but no serum drug or metabolite levels were reported.12 Moreover, the 377C>T polymorphism has been associated with a higher diol/epoxide ratio and the 416A>G polymorphism with a lower ratio, suggesting altered activity for the corresponding variant enzyme.13
It was reported that the frequency of the C allele for EPHX1 337T>C was 0.29 in a population from northern Italy, while the frequency of the G allele for EPHX1 416A>G was 0.16 in the same population.6 These values are comparable with our data. Indeed, the frequency of the C allele for EPHX1 337T>C was 0.35 while the frequency of the G allele for EPHX1 416A>G was 0.18.
In vitro expression studies of the EPHX1 gene have shown that the 337T>C variant confers a 40% decrease in hydrolase activity and that the 416A>G variant confers a 25% increase in activity. In our study, subjects with 337 TT did not show higher levels of circulating CBZ 10,11-epoxide 4 hours after administration of the drug. Similarly, no significant decrease was observed in subjects heterozygous for the 416 AG variant. The influence of microsomal epoxide hydrolase variants on the metabolism of CBZ is still under investigation.
In our study population, the frequency of the CYP3A4*22 (rs 35599367, C>T) allele was 0.03, as reported in the literature for the Caucasian population.14 Although recently associated with low hepatic CYP3A4 expression and activity, this allelic variant did not appear to affect CBZ metabolism in the epileptic patients in our study. One of the limitations of this study was its relatively small sample size, so our results require validation in a larger cohort. However, it is challenging to obtain patients on monotherapy with CBZ, given that this drug is commonly used in combination with other antiepileptic drugs.
Conclusion
Although the literature contains some evidence of involvement of these polymorphisms in CBZ metabolism in vitro, our results show no interference in vivo. However, our preliminary findings require confirmation in a larger population of epileptic patients receiving CBZ. We intend to extend this study to include epileptic patients on polytherapy in order to assess the influence of different drugs on the activity of microsomal epoxide hydrolase, which is involved in the metabolism of CBZ.
Disclosure
The authors report no conflicts of interest in this work.
References
Shovoron SD. Handbook of the Treatment of Epilepsy. Oxford, UK: Blackwell Science; 2000. | |
Tolou-Ghamari Z, Zare M, Habibabadi JM, et al, A quick review of carbamazepine pharmacokinetics in epilepsy from 1953 to 2012. J Res Med Sci. 2013;18 Suppl 1:S81–S85. | |
Kerr BM, Thummel KE, Wurden CJ, et al. Human liver carbamazepine metabolism. Role of CYP3A4 and CYP2C8 in 10,11-epoxide formation. Biochem Pharmacol. 1994;47:1969–1979. | |
Puranik YG, Birnbaum AK, Marino SE, et al. Association of carbazepine major metabolism and transport pathway gene polymorphisms and pharmacokinetics in patients with epilepsy. Pharmagenomics. 2013;14:35–45. | |
Junji S, Takateru I, Kazuko N. Update on the genetic polymorphisms of drug-metabolizing enzymes in antiepileptic drug therapy. Pharmaceuticals. 2010;3:2709–2732. | |
Ensembl. Variation: rs1051740 SNP. Available from: http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=1:226019133-226020133;v=rs1051740;vdb=variation;vf=835988. Accessed January 31, 2014. | |
Wang D, Guo Y, Wrighton SA, et al. Intronic polymorphism in CYP3A4 affects hepatic expression and response to statin drugs. Pharmacogenomics J. 2011;11:274–286. | |
Harrison DJ, Hubbard AL, MacMillan J, et al. Microsomal epoxide hydrolase gene polymorphism and susceptibility to colon cancer. B J Cancer. 1999;79:168–171. | |
Deng H, Xiu X, Song Z. The molecular biology of genetic-based epilepsies. Mol Neurobiol. August 10, 2013. [Epub ahead of print.] | |
Nakken KO, Taubøll E. [Drug-resistant epilepsy]. Tidsskr Nor Laegeforen. 2009;129:1986–1989. Norwegian. | |
Tomalik-Scharte D, Lazar A, Fuhr U, et al. The clinical role of genetic polymorphisms in drug-metabolizing enzymes. Pharmacogenomics J. 2008;8:4–15. | |
Makmor-Bakry M, Sills G, Hitiris N, et al. Genetic variants in microsomial epoxide hydrolase influence carbamazepine dosing. Clin Neuropharmacol. 2009;32:205–211. | |
Nakajima Y, Saito Y, Shiseki K, et al. Haplotype structure of EPHX1 and their effects on metabolism of carbamazepine-10,11-epoxide in Japanese epileptic patients. Eur J Clin Pharmacol. 2005;61:25–34. | |
Elens L, van Gelder T, Hesselink DA, Haufroid V, van Schaik RH. CYP3A4*22: promising newly identified CYP3A4 variant allele for personalizing pharmacotherapy. Pharmacogenomics. 2013;14:47–62. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.