")
Back to Journals » Drug Design, Development and Therapy » Volume 8
Drug development strategies for the treatment of obesity: how to ensure efficacy, safety, and sustainable weight loss
Authors Barja-Fernandez S, Leis R, Casanueva F, Seoane L
Received 30 September 2014
Accepted for publication 29 October 2014
Published 1 December 2014 Volume 2014:8 Pages 2391—2400
DOI https://doi.org/10.2147/DDDT.S53129
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Shu-Feng Zhou
S Barja-Fernandez,1–3 R Leis,2 FF Casanueva,3,4 LM Seoane1,3
1Grupo Fisiopatología Endocrina, 2Departamento de Pediatría, Universidad de Santiago de Compostela (USC), Instituto de Investigación Sanitaria de Santiago de Compostela (IDIS), Complexo Hospitalario Universitario de Santiago (CHUS/SERGAS), Santiago de Compostela, Spain; 3CIBER Fisiopatologia de la Obesidad y Nutrición, Instituto de Salud Carlos III, Santiago de Compostela, Spain; 4Laboratorio de Endocrinología Molecular y Celular, Universidad de Santiago de Compostela (USC) Instituto de Investigación Sanitaria de Santiago de Compostela (IDIS), Complexo Hospitalario Universitario de Santiago (CHUS/SERGAS), Santiago de Compostela, Spain
Abstract: The prevalence of obesity has increased worldwide, and approximately 25%–35% of the adult population is obese in some countries. The excess of body fat is associated with adverse health consequences. Considering the limited efficacy of diet and exercise in the current obese population and the use of bariatric surgery only for morbid obesity, it appears that drug therapy is the only available method to address the problem on a large scale. Currently, pharmacological obesity treatment options are limited. However, new antiobesity drugs acting through central nervous system pathways or the peripheral adiposity signals and gastrointestinal tract are under clinical development. One of the most promising approaches is the use of peptides that influence the peripheral satiety signals and brain–gut axis such as GLP-1 analogs. However, considering that any antiobesity drug may affect one or several of the systems that control food intake and energy expenditure, it is unlikely that a single pharmacological agent will be effective as a striking obesity treatment. Thus, future strategies to treat obesity will need to be directed at sustainable weight loss to ensure maximal safety. This strategy will probably require the coadministration of medications that act through different mechanisms.
Keywords: obesity, energy balance, pharmacotherapy
Introduction
Obesity was defined as an epidemic of the 21st century by the World Health Organization (WHO) and has become a serious health problem worldwide. It has been estimated that at least 3.4 million people die each year as a result of being overweight or obese and 35.8 million (2.3%) of global Disability Adjusted Life Years (DALYs) are caused by this pathology.1 DALY is a health gap measure that extends the concept of potential years of life lost due to premature death to include equivalent years of “healthy” life lost by virtue of being in states of poor health or disability.1
The worldwide prevalence of obesity nearly doubled between 1980 and 2008, and it currently affects approximately 30%–35% of the general population in the USA and 25% in the UK.2,3 Moreover, childhood obesity is currently one of the most serious health challenges, and its prevalence has increased worldwide at an alarming rate in recent decades.4,5 In 2012, more than 40 million children under the age of five were overweight or obese. Additionally, 70 million children under age five will be overweight or obese by 2025 if current trends continue. The WHO has estimated that in 2035 more than 300 million adults and children will be obese.6
Obesity is partly regarded as an evolutionary issue. It is postulated that for the human population to survive it was necessary to develop a thrifty genotype, and genes that facilitate fat reserves for use as energy during harder times were selected.7 The ancestors of humans experienced situations that contrasted with the present, which is characterized by availability of abundant food and low physical exercise. These changes have converted an evolutionary benefit into a serious metabolic problem and obesity.
Obesity is associated with increases in morbidity, premature mortality, impaired quality of life, and large health care cost.8–12 The major comorbidities include the following: metabolic syndrome, type 2 diabetes, hypertension, dyslipidemia, myocardial infarction, and certain cancers.12,13 Obesity is considered a chronic medical disease state.14–16 Consequently, more and more strategies focusing on obesity treatment are being explored. However, to develop novel pharmacological therapies effective against obesity, we must understand the main physiological mechanism controlling energy homeostasis. Only then will it be possible to ensure efficacy, safety, and sustainable weight loss.
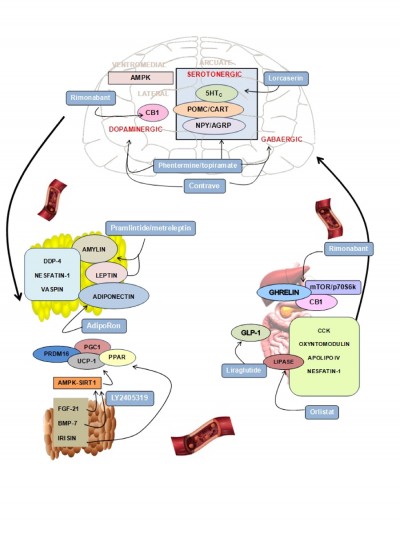
Here we review the main physiological mechanisms that constitute the cornerstone of pharmacological antiobesity strategies. We focus on the most novel and least explored areas of research and describe drugs approved or under study currently targeting the different systems (Figure 1 and Table 1).
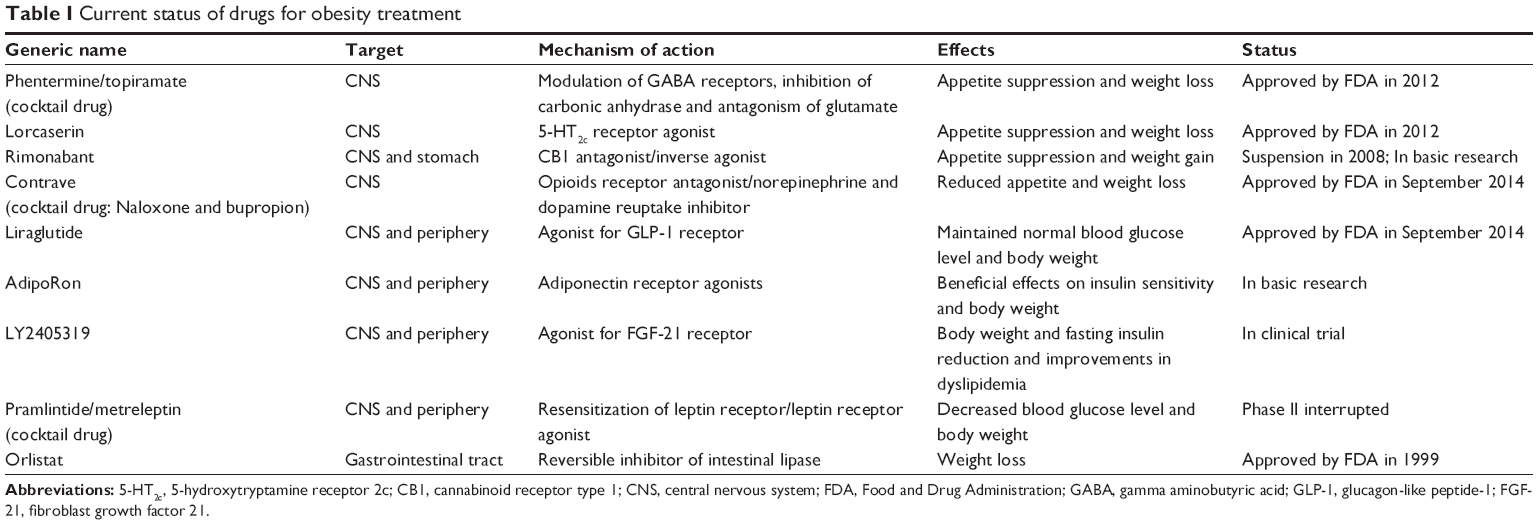 | Table 1 Current status of drugs for obesity treatment |
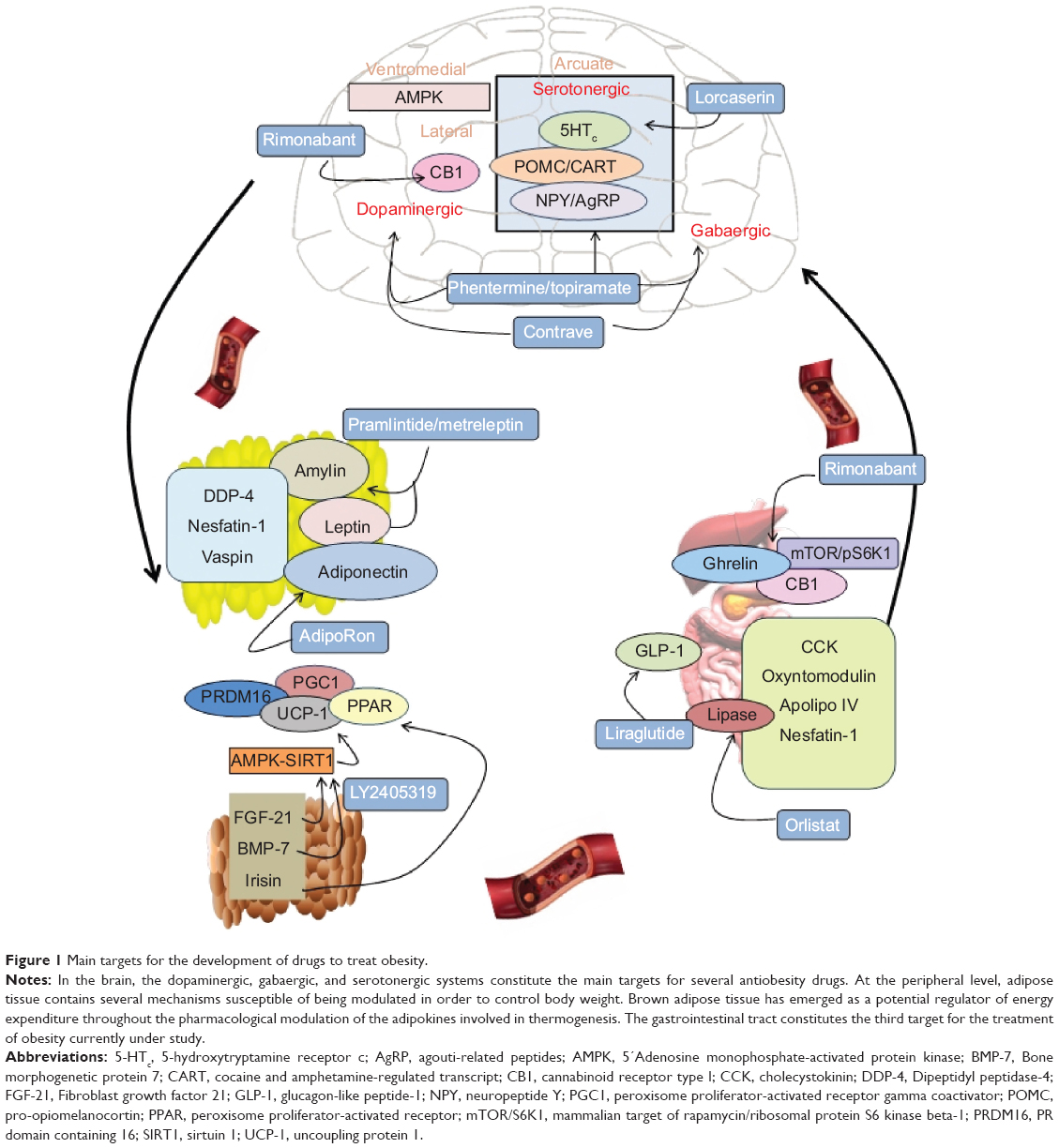 | Figure 1 Main targets for the development of drugs to treat obesity. |
Regulation of food intake, energy balance, and body fat mass
Obesity is defined as abnormal and excessive fat accumulation caused by an imbalance between energy intake and caloric expenditure. Body weight control consists of a complex mechanism regulated by hormonal, metabolic, and nervous pathways. A body weight “set point” existence has been suggested for every individual, and it is regulated by physiological mechanisms that use energy sensors located in the peripheral organs to inform the brain about the energy status of the organism. An effective treatment for obesity would require an efficient knowledge of factors and mechanisms potentially regulating food intake, energy balance, and body fat mass.17
Central regulation of energy homeostasis
The hypothalamus is the central brain structure responsible for food intake regulation. Neuronal clusters constitute the hypothalamic nuclei interconnecting neuronal circuits capable of responding to changes in energy status by modifying the expression of specific neuropeptides, which cause changes in energy intake and expenditure. Crucial relevance in food intake regulation is exerted by the arcuate nucleus in the hypothalamus, where there coexist neurons coexpressing orexigenic peptides such as neuropeptide Y (NPY) and agouti-related peptides (AgRP) with an adjacent set of neurons coexpressing anorexigenic peptides pro-opiomelanocortin (POMC) and cocaine and amphetamine-regulated transcript (CART).18 Hypothalamic neurons respond to peripheral signals such as leptin, ghrelin, glucose, insulin, and the main neurotransmitters by modifying the synthesis of anorexigenic/orexigenic neuropeptides and finally regulating food intake.
The regulation of food intake and energy expenditure at the central nervous system (CNS) involves the action of neurotransmitters and neuromodulators. Among the neurotransmitters, it was widely described that dopamine exhibits relevant effects on food intake. Animal models of dopamine deficiency show decreased food intake.19 In fact, it has been shown that dopamine activates brain centers of reward and pleasure that affect food intake.20 In addition, central serotoninergic innervations of the hypothalamus exert anorexigenic actions, especially through the receptors 5-hydroxytryptamine receptor 2B (5-HT2B) and C (5-HT2C).21 Signaling by opioid peptides leads the hedonic components of food and opioid agonist to induce intake of palatable food. In this context, targeting the main central neurotransmitters capable of modulating the main hypothalamic neuropetides production now constitutes one of the most promising therapies to fight against obesity.
The most recent findings in the field of central regulation of energy homeostasis have revealed that hypothalamic lipid metabolism is a crucial mechanism regulating energy balance.18 Accordingly, the main peripheral signals that are crucial for the regulation of energy balance (leptin, ghrelin, and cannabinoids) modulate key enzymes of the lipid metabolism such as AMP-activated protein kinase (AMPK) and acetil-CoA carboxylase (ACC).22
Another system involved in energy balance regulation at the central level is the endocannabinoid system (EC), and, specifically, cannabinoid receptor type 1 (CB1) agonism has been shown to ensure palatable food consumption. On the contrary, the antagonist for CB1 induces weight loss in obesity possibly through the improvement of leptin sensitivity at the central level.23
Energy balance regulation by adipose tissue
The adipose tissue, classically considered as a lipid reservoir, has gained increased attention as an endocrine organ involved in energy homeostasis regulation. In addition to the fatty acids, the liberation from the adipose tissue of bioactive factors named adipokines has been revealed to be altered in obesity. Adipokines mediate physiological actions contributing to energy homeostasis maintenance such as food intake regulation, energy expenditure, and insulin sensitivity.24
The isolation of leptin in 1999 represents an important starting point in the study of the adipose tissue as an endocrine organ. Leptin is produced mainly in the white adipose tissue (WAT) and circulates in plasma correlating with the fat content. It acts through the leptin receptors in the arcuate nucleus of the hypothalamus regulating appetite. At the hypothalamic level, the mechanism of leptin action involves the inhibition of orexigenic neuropeptides in parallel with the increase in anorexigenic peptides.25 In addition to leptin, an increasing number of novel adipokines were identified, some of whose functions still remain unknown. Among the adipokines, the adiponectin has attracted considerable attention because of its insulin-sensitizing properties.26 It has been found that adiponectin levels are downregulated under adverse fat distribution and associated with adipose tissue dysfunction.26
Dipeptidyl peptidase-4 (DPP-4) has been recently identified as an adipokine that shows an elevated production from adipose tissue in obese patients when compared with lean controls.27 Several studies support the relationship among elevated DPP-4 release and obesity and insulin resistance.28 Another signal identified as an adipokine is nesfatin-1, which is secreted from different tissues such as brain, stomach, and adipose tissue. Several studies focusing on the mechanism of action of nesfatin-1 have proposed it as a novel satiety molecule with a direct effect on glucose metabolism.29 Vaspin has also emerged as an adipokine with the potential to improve insulin resistance, although the exact mechanism of action is still not elucidated.30
Two types of adipose tissue have been described: WAT, which stores fat, and brown adipose tissue (BAT), which dissipates energy in the form of heat through nonshivering thermogenesis. The adipokines produced by BAT have proved to be capable of exerting beneficial effects on metabolism. The capacity of the BAT-derived adipokines include differentiation of adipose stem cells into brite adipocytes, which induce thermogenic activation, increase energy expenditure and finally, reduce fat composition and body wight. The adipokines derived from BAT are the subject of a wide range of studies. Among them, fibroblast growth factor 21 (FGF-21) was discovered as an important metabolic regulator primarily produced by the liver and adipose tissue.31 FGF-21 exerts significant glucose- and lipid-lowering actions and has thermogenic effects. The administration of FGF-21 produced beneficial metabolic effects in animal models.31 The mechanism of action for the metabolic effects of FGF-21 is associated with its ability to activate peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1-α), which is a master regulator of mitochondrial biogenesis and oxidative metabolism.32 In addition, there is a positive role for FGF-21 in regulating the interaction between AMPK and sirtuin 1 (SIRT1), which is essential for brown thermogenesis.33
Bone morphogenetic protein 7 (BMP-7) induces brown adipogenesis in subcutaneous adipose tissue, increases energy expenditure, reduces food intake, and decreases weight gain.34,35 These effects might be mediated by its capacity to induce the activation of the p18-AMPK pathway that regulates the expression of thermogenic factors such as PR domain containing 16 (PRDM16), PGC1-α, and uncoupling protein 1 (UCP-1).
Irisin was recently described as a myokine and an adipokine produced by cleavage of the fibronectin type 3 domain containing 5 (FNDC5) protein.36,37 It has been proposed as a regulator of peroxisome proliferator-activated receptor gamma (PPARγ) and is attributed to browning properties such as increases of brown fat, UCP-1 levels, and oxygen consumption. Several studies have reported an increase in irisin production after exercise. However, recent studies in humans indicate that neither acute nor chronic exercise increases the endogenous concentration of FNDC5/irisin.38,39
Energy balance regulation by gastrointestinal tract
More than 30 peptides are secreted from the enteroendocrine cells of the gastrointestinal tract in response to ingested food. These peptides are involved in hunger/satiety perception by interacting with long-acting hormones involved in body weight regulation such as leptin and insulin to maintain energy homeostasis.40 The majority of satiation-inducing gut peptides mediate their effects via vagal afferent fibers, although some enter the brain from the circulation and exert their effects directly.41 In addition, there is a fundamental role for these hormones in several digestive functions, including gastric emptying and energy intake regulation.42 Thus, the gastrointestinal tract-derived peptides involved in the control of energy homeostasis have recently garnered a notable degree of attention. Among these peptides, the most studied include cholecystokinin (CCK), glucagon-like peptide-1 (GLP-1), oxyntomodulin, peptide YY (PYY), apolipoprotein A.IV, and enterostatin.43 Since its isolation in 1999, ghrelin has been considered the most relevant hormone because it is the only stomach-derived peptide with orexigenic actions.44,45 Another gastrointestinal hormone, GLP-1, is postprandially released primarily from the distal small intestine and colon L cells.44 In humans, GLP-1 reduces energy intake, gastric emptying rates, and energy consumption.46 It has been suggested that there is an antagonistic effect of ghrelin on GLP-1 in the regulation of food intake.43 In addition, an inverse relationship between circulating levels of ghrelin and GLP-1 has been described.46 This result may indicate an interaction between these two peptides. Nesfatin-1 is a stomach-derived peptide involved in food intake regulation that was recently discovered, and it is receiving increasing interest as a regulator of energy homeostasis.47
The EC system, and particularly CB1, has emerged as a relevant target to treat obesity and its pathologies. It is known that EC production is regulated by nutritional status at specific areas of the brain involved in modulating feeding behavior. However, the EC system is not only present at the central level. It was shown that the major peripheral organs involved in metabolism regulation such as adipose tissue, the liver, the endocrine pancreas, and the skeletal muscles are targets for cannabinoid actions.48 The EC system plays a role in glucose homeostasis, lipogenesis, and insulin sensitivity regulation in the periphery.48 Moreover, it was recently demonstrated that CB1 receptors are localized in the stomach in the neuroendocrine gastric cells producing ghrelin.49 All of the recent studies regarding the involvement of the gastric EC system suggest that any regulator mechanism at this level can be implicated in gastric–brain communication and the mediation of energy balance control.
Recent studies of the metabolic control of energy balance focused on the intracellular pathway mammalian target of rapamycin/ribosomal protein S6 kinase beta-1 (mTOR/S6K1), which has been proposed as an intracellular energy sensor. This system is regulated by nutritional status and pathological situations such as obesity and diabetes in the brain and several peripheral tissues such as the liver and skeletal muscle.50
This intracellular pathway participates in the brain–stomach connection and regulation of energy balance indirectly through the action of gastric peptides on hypothalamic neurons, which regulates food intake. Recently, a novel gastric mechanism regulating central food intake was described involving a connection between gastric cannabinoid receptor CB1 and ghrelin through the intracellular mTOR pathway via the neural control of the vagus nerve.51 The work showed that the pharmacological blockade of the cannabinoid receptor in fasting states by the use of two antagonists (rimonabant and AM281) is sensed by the gastric cells as a satiety signal comparable to food intake. This signal consequently decreases ghrelin secretion from the stomach and decreases food intake and body weight.
Future studies focusing on this novel mechanism will reveal whether additional gastrokines besides ghrelin might be involved in this gastric system to regulate food intake by the EC system and via mTOR. Previous studies have indicated that the production of the adipokine nesfatin-1 opposes the effects of ghrelin and it is also regulated by the gastric intracellular mTOR pathway.52
The classical idea of a central regulation of food intake and body weight has led to the concept that energy balance results from the interaction between the CNS and peripheral organs directly involved in body weight regulation such as the gastrointestinal tract and adipose tissue. Novel therapeutic strategies are being designed to treat obesity by acting on the main pathways communicating with the brain and periphery.
New pharmacological management interventions for obesity
The strategies recommended for weight loss include lifestyle interventions, pharmacotherapy, and bariatric surgery.14 The foundation of obesity care consists in lifestyle changes that include diet, physical activity, and behavior change therapies. The most important strategy for sustainable body weight loss is caloric intake reduction together with increased and maintained physical activity.53,54
With the exception of bariatric surgery, individual pharmacological interventions have shown limited success.41,55,56 An effective pharmacological intervention to treat obesity should provide sustained weight loss and avoid adverse effects. However, it is difficult to achieve this goal because energy balance regulation has redundancy and overlapping physiological functions. Thus, weight loss is affected by external factors that limit the effectiveness of pharmacological interventions.
The antiobesity drugs currently under investigation act mainly at two different levels. The first level is the central level, and the second level is the peripheral organs with relevant roles in energy homeostasis such as adipose tissue and the gastrointestinal tract.
Pharmacotherapy is approved for patients with a body mass index (BMI) ≥30 kg/m2 or ≥27 kg/m2 when complicated by obesity-related comorbidity (FDA: Food and Drug Administration).14
The brain as a target for the development of antiobesity drugs
The role of the brain in appetite regulation and body weight control is well known.57 Thus, drugs targeting the CNS might represent the most promising obesity therapy.
The drugs acting through the CNS affect neurotransmitters with anorexiants or appetite suppressant actions. These treatments have targeted three monoamine receptor systems in the hypothalamus, namely, the noradrenergic, dopaminergic, and serotonergic systems.58 The first compound introduced as an appetite suppressant was amphetamine, which was addictive and had euphoric side effects. To reduce these side effects, amphetamine-derived anorexiants such as phentermine were produced, and this is the most commonly prescribed agent in the USA but is prohibited in Europe. The most common side effects of the anorexiant compounds are restlessness, insomnia, dry mouth, constipation, and increased blood pressure and heart rate. The phentermine/topiramate combination contains a stimulator of noradrenaline, dopamine, and serotonin release with an anticonvulsant drug. This combination treatment was approved by FDA because present synergic effects allow the use of lower doses of each substance and reduced the toxicity. The mechanism responsible for weight loss is uncertain, but it is thought to be mediated through its modulation of gamma aminobutyric acid receptors, inhibition of carbonic anhydrase, and antagonism of glutamate to reduce food intake.14 Clinical trials have shown the efficacy and safety of the treatment. The major concern about this medication is teratogenicity, and hence it should never be administered to pregnant women.
Specific central serotonin receptor agonism represents an effective pharmacological approach to body weight loss. The first drugs developed to target serotonin receptors were fenfluramine and dexfenfluramine. They were used in combination with phentermine. However, the drugs were withdrawn because of adverse cardiovascular and lung effects. These adverse effects were produced by its 5-HT2B agonism, which is expressed in the cardiovascular system. Novel and safe strategies are necessary to develop drugs acting specifically in another family member of the 5-HT such as 5-HT2C. This family member is only expressed in the brain and participates in energy balance regulation. One appetite suppressant approved by the FDA in 2012 was lorcaserin, which is a selective 5-HT2C receptor agonist that is thought to decrease food intake through the POMC system of neurons.59
Another drug that suppresses appetite and weight gain is rimonabant. It is a CB1 antagonist/inverse agonist developed in the mid-1990s. This drug was licensed in Europe as an antiobesity agent in 2006. However, it was never approved in the US. The presence of serious psychiatric problems led to its suspension in 2008. This decision rapidly led to the termination of several CB1-receptor-antagonist antiobesity drug development programs.60
In September 2014, the FDA has approved two new drugs for obesity. One of them is Contrave, a CNS-level-acting drug that is a combination of naloxone and bupropion formulated for extended release. Naloxone is a nonselective antagonist for opioids, and bupropion a norepinephrine and dopamine reuptake inhibitor.61 The second approved drug is liraglutide 3 mg, an agonist of GLP-1 receptor acting at the central and peripheral levels. All these drugs must be used as adjuncts to diet and exercise.
Adipose tissue as a target to treat obesity
Obesity is characterized by excessive fat accumulation predominantly in visceral depots, and it is associated with the development of several pathologies such as cardiovascular diseases, type 2 diabetes, and fatty liver disease.62
The main well-known adipokines are deregulated in obesity, and they are involved in the regulation of energy balance through their role in food intake control, fat distribution, insulin sensitivity, and energy expenditure.24 Adipokines have emerged as potential biomarkers and pharmacological treatment strategies for obesity. A number of adipokines are currently being studied as potential candidates for the development of antiobesity therapies. Such adipokines include leptin, adiponectin, DPP-4, FGF-21, nesfatin-1, BMP-7, tumor necrosis factor alpha (TNF-α), apelin, and vaspin.62 The majority of these adipokines act in the brain to increase energy expenditure and cause weight loss.63,64 Thus, the concept of an interconnection between the CNS and adipose tissue has promoted the search for novel adipokines that affect obesity and may be potential targets for pharmacological strategies.
Among the known adipokines, only leptin has reached advanced clinical study phases, and the remaining adipokines are currently under preclinical studies. Leptin secretion from adipose tissue was discovered in 1994, and it was proposed as an obesity treatment on the basis of its anorexigenic effect. Leptin acts on the hypothalamic nucleus, which regulates body weight and appetite.25 However, obese patients present elevated serum levels of this adipokine, and there is central resistance to the weight-lowering effects of exogenously administered leptin.65,66 Thus, recombinant leptin is not used for clinical treatment, but only in a small number of centers as a protocol for research in patients with congenital leptin deficiency.67 Metreleptin is an analog of human leptin that has been recently approved for the treatment of lipodystrophy in Japan. However, it has not yet been approved by the US and European regulatory agencies.68 Recently, leptin was combined with other peptides such as amylin, exendin-4, and FGF-21 as promising antiobesity drugs.69 However, the last clinical study, based on the administration of a combination of the amylin agonist and leptin analogs (pramlintide/metreleptin), showed efficacy in reducing body weight. However, this trial was suspended because of relevant adverse effects. Future antiobesity strategies investigating leptin analogs in diet-induced obese rodents and human obesity will require increased potency and sustained action to obtain sustainable weight loss, ensure safety, and minimize adverse effects.
Adiponectin is another adipokine proposed as a promising therapy for obesity, in view of its peripheral and central effects on improving insulin sensitivity, lowering body weight, and increasing energy expenditure.26 Several studies have been conducted with recombinant adiponectin and adiponectin analogs. These studies showed beneficial effects on insulin sensitivity and body weight.26,70,71 Recent research suggests a more promising strategy includes the development of adiponectin receptor agonists, as shown by the initial studies with AdipoRon. AdipoRon was shown to have beneficial effects on insulin resistance in mice receiving a high-fat diet.72 However, the main limitation of this therapy is the lack of clinical trial data in humans.
An additional adipokine that has recently attracted special interest is DPP-4, which was initially studied as an incretin inhibitor. Several DPP-4 antagonists are clinically used as antidiabetic drugs.73 However, a new pathway has been identified by specifically targeting actions of DPP-4 directly at the adipose tissue. The effects of DPP-4 on metabolism were produced independently of their actions mediated by incretins. The main limitation of this therapeutic strategy is that human data on DPP-4 action as an adipokine are not available.
Other candidate adipokines were examined in the search for therapeutic targets able to provide a stable and sustained weight loss. These candidates must be accompanied by an improvement of the obesity-associated pathologies. Examples of candidate adipokines include nesfatin-1, vaspin, and amylin. However, the main limitation to advancements in the development of these candidates is the lack of human data. In addition, the mechanisms of action controlling effects in regulating homeostasis have not been identified.
Harnessing the thermogenic effects of new adipokines is a novel strategy to treat obesity
A new trend has recently emerged in treating obesity by targeting energy expenditure. The novel strategies are focused on the findings that human BAT, which dissipates energy by increased energy expenditure, is inversely correlated with BMI.74 The adipokines produced by BAT have beneficial effects on metabolism and browning action on WAT that reduces fat composition and body weight.
LY2405319 is an FGF-21 analog that has been produced and tested in a proof-of-concept trial. The trial reported beneficial effects on body weight reduction and fasting insulin. It also improved dyslipidemia in subjects with obesity and type 2 diabetes.33 Therefore, FGF-21-based therapies might be effective for the treatment of obesity and related disorders.
BMP-7 has been suggested as a novel therapeutic approach for obesity and metabolic diseases. However, there are no clinical data available on the effects of this adipokine in obesity and metabolic diseases.
Despite the initial expectation of using irisin as an antiobesity drug, the in vivo and in vitro data obtained in rodents has not been reproduced in humans. This lack of data suggests that we should be careful when using pharmacological applications of irisin to induce browning.
There are currently no adipokines approved for weight loss, and only an analog of amylin is in phase I study. The drug is a combination of leptin and an amylin analog (pramlintide, metreleptin). The drug was moved to a phase II study but had to be interrupted owing to the generation of antibodies and severe adverse effects.
In summary, the novel strategies to treat obesity by targeting adipose tissue have not been completely addressed. The main limitation of the drugs targeting adipokines is the lack of knowledge regarding the exact mechanisms in controlling obesity. The major problem in the development of drugs targeting browning and thermogenesis is a safety issue. The main risk is that thermogenesis activation involves adrenergic activity and might induce cardiovascular effects. In the future it will be important to produce thermogenic analogs with mechanisms independent of adrenergic activation.
Current and future strategies targeting the gastrointestinal tract in obesity treatment
Gastric surgery is the most effective treatment for obesity and has led to the investigation of signals derived from the gastrointestinal tract as promising targets in treating obesity. Recent antiobesity research has been focused on peripheral hunger and satiety signals that require central integration to communicate the organism’s nutritional status to the brain centers and allow efficient energy homeostasis.75
The gastrointestinal tract and the gastrokines together with novel systems such as the EC system and the intracellular mTOR pathway represent promising pharmacological targets for the development of therapies against obesity.
Orlistat is currently the only approved drug that acts at the gastrointestinal level. It is a potent lipase inhibitor that reduces intestinal fat absorption and is recommended for long-term obesity treatment. The safety of this drug is based on data showing fat is not absorbed and consequently does not display nondesirable systemic effects. However, the main limitation of orlistat is gastrointestinal symptoms that, in general, can be avoided by patients if they limit their high-fat diet. Another limitation is that in addition to reducing fat absorption, the drug also inhibits vitamins. However, this problem can be solved with vitamin supplementation.76
Another novel therapy based on gut–brain communication takes advantage of the development of GLP-1 analogs such as liraglutide, which is an agonist for the GLP-1 receptor. This drug was previously used for diabetes treatment. A new strategy currently being explored is the use of liraglutide at higher doses than those used for diabetes. The objective is to induce weight loss and reduce obesity. In addition, the rate of liraglutide metabolism is much lower than for GLP-1, which ensures a longer duration of action. Occasionally, liraglutide treatment can induce gastrointestinal effects such as nausea and vomiting.61 As mentioned above, the use of liraglutide 3 mg for obesity treatment was very recently approved (September 2014).
One very promising strategy to treat obesity is the modulation of the EC system, but this was scuttled with the withdrawal of rimonabant due to adverse CNS-related side effects. However, the peripheral antagonist of CB1 receptors might still represent a new possibility with beneficial metabolic effects for cannabinoid-derived drugs. Drugs that act on the periphery rather than on the CNS should be explored in the future. However, a predictable limitation of this type of therapy is the short half-life characteristics of drugs at the gastric level, which complicates their oral administration. In addition, the use of drugs acting on a single molecular target provides only modest reductions in body weight.
Incretins or other pharmacological agents may play an important role in combating the obesity epidemic. Although any antiobesity drug may affect one or several of the systems that control food intake and energy expenditure, it is unlikely that a single pharmacological agent will be an effective obesity treatment. Thus, future strategies to treat obesity will have to induce effective weight loss and will likely require the coadministration of medications that act through different mechanisms.
Therapeutic weight loss interventions have provided only limited long-term success. An effective obesity treatment would require an evaluation of factors potentially affecting energy intake, expenditure, and metabolism.17 Because the factors causing obesity are incompletely understood, weight loss strategies may not address the root causes of energy imbalance.17,77 Therefore, there is an urgent need to develop novel pharmacological treatment strategies targeting the mechanisms underlying positive energy balance and excessive and adverse fat accumulation. One key priority at present is to delineate how mechanisms governing food intake and body fat content are altered in an obesogenic environment. This information will allow the development of effective new obesity prevention and treatment strategies.
The reduction of body weight might not be the only or best approach to improve obesity-related diseases. Novel treatment concepts may include changing metabolically unhealthy patients into metabolically healthy obese individuals. Such a phenotype switch could be achieved by a reduction in fat mass, improvement of adipose tissue function, or normalization of the adipokine, diabetogenic, and proinflammatory profiles.62
Conclusion
In conclusion, owing to the limited efficacy of dietary restrictions, physical exercise, and lifestyle modifications in controlling obesity, the only realistic approach available to clinicians is pharmacotherapy. The drugs currently in use all lead to an approximately 5 kg weight loss compared with placebo, and no striking differences are observed. A better understanding of the mechanisms controlling appetite, satiety, and energy homeostasis may allow the development of more effective drugs in the future.
Acknowledgments
This research has been funded by Instituto de Salud Carlos III (PS09/02075 and PI12/02021); Xunta de Galicia (10 PXIB 918 273PR), Fundación Mutua Madrileña. CIBER Fisiopatologia de la Obesidad y Nutrición is an ISCIII initiative. SB-F is funded by Xunta de Galicia.
Disclosure
The authors report no conflicts of interest in this work.
References
World Health Organization. The SuRF Report 2. Geneva, Switzerland: World Health Organization, 2005. | ||
Wang Y, Beydoun MA. The obesity epidemic in the United States–gender, age, socioeconomic, racial/ethnic, and geographic characteristics: a systematic review and meta-regression analysis. Epidemiol Rev. 2007;29:6–28. | ||
Rodgers RJ, Tschop MH, Wilding JP. Anti-obesity drugs: past, present and future. Dis Model Mech. 2012;5:621–626. | ||
Carotenuto M, Bruni O, Santoro N, et al. Waist circumference predicts the occurrence of sleep-disordered breathing in obese children and adolescents: a questionnaire-based study. Sleep Med. 2006;7:357–361. | ||
Carotenuto M, Santoro N, Grandone A, et al. The insulin gene variable number of tandemrepeats (INS VNTR) genotype and sleep disordered breathing in childhood obesity. J Endocrinol Invest. 2009;32:752–755. | ||
World Health Organization. Obesity: Preventing and Managing the Global Epidemic (WHO/NUT/NCD/98.1). Geneva, Switzerland: World Health Organization, 1997. | ||
O’Dea K. Overview of the thrifty genotype hypothesis. Asia Pac J Clin Nutr. 1995;4:339–340. | ||
Kopelman PG. Obesity as a medical problem. Nature. 2000;404:635–643. | ||
Fontaine KR, Redden DT, Wang C, Westfall AO, Allison DB. Years of life lost due to obesity. JAMA. 2003;289:187–193. | ||
Haslam DW, James WP. Obesity. Lancet. 2005;366:1197–1209. | ||
Esposito M, Antinolfi L, Gallai B, et al. Executive dysfunction in children affected by obstructive sleep apnea syndrome: an observational study. Neuropsychiatr Dis Treat. 2013;9:1087–1094. | ||
Carotenuto M, Esposito M, Parisi L, et al. Depressive symptoms and childhood sleep apnea syndrome. Neuropsychiatr Dis Treat. 2012;8:369–373. | ||
Flegal KM, Graubard BI, Williamson DF, Gail MH. Cause-specific excess deaths associated with underweight, overweight, and obesity. JAMA. 2007;298:2028–2037. | ||
Kushner RF. Weight loss strategies for treatment of obesity. Prog Cardiovasc Dis. 2014;56:465–472. | ||
Verrotti A, Carotenuto M, Altieri L, et al. Migraine and obesity: metabolic parameters and response to a weight loss programme. Pediatr Obes. Epub 2014 July 3. | ||
Verrotti A, Agostinelli S, D’Egidio C, et al. Impact of a weight loss program on migraine in obese adolescents. Eur J Neurol. 2013;20:394–397. | ||
Sharma AM, Padwal R. Obesity is a sign–over-eating is a symptom: an aetiological framework for the assessment and management of obesity. Obes Rev. 2010;11:362–370. | ||
Dieguez C, Vazquez MJ, Romero A, Lopez M, Nogueiras R. Hypothalamic control of lipid metabolism: focus on leptin, ghrelin and melanocortins. Neuroendocrinology. 2011;94:1–11. | ||
Hnasko TS, Szczypka MS, Alaynick WA, During MJ, Palmiter RD. A role for dopamine in feeding responses produced by orexigenic agents. Brain Res. 2004;1023:309–318. | ||
Cawley EI, Park S, aan het Rot M, et al. Dopamine and light: dissecting effects on mood and motivational states in women with subsyndromal seasonal affective disorder. J Psychiatry Neurosci. 2013;38:388–397. | ||
Singh M. Mood, food, and obesity. Front Psychol. 2014;5:925. | ||
Lage R, Dieguez C, Vidal-Puig A, Lopez M. AMPK: a metabolic gauge regulating whole-body energy homeostasis. Trends Mol Med. 2008;14:539–549. | ||
Guyenet SJ, Schwartz MW. Clinical review. Regulation of food intake, energy balance, and body fat mass: implications for the pathogenesis and treatment of obesity. J Clin Endocrinol Metab. 2012;97:745–755. | ||
Bluher M. Clinical relevance of adipokines. Diabetes Metab J. 2012;36:317–327. | ||
Seoane LM, Lopez M, Tovar S, et al. Agouti-related peptide, neuropeptide Y, and somatostatin-producing neurons are targets for ghrelin actions in the rat hypothalamus. Endocrinology. 2003;144:544–551. | ||
Turer AT, Scherer PE. Adiponectin: mechanistic insights and clinical implications. Diabetologia. 2012;55:2319–2326. | ||
Sell H, Bluher M, Kloting N, et al. Adipose dipeptidyl peptidase-4 and obesity: correlation with insulin resistance and depot-specific release from adipose tissue in vivo and in vitro. Diabetes Care. 2013;36:4083–4090. | ||
Kloting N, Fasshauer M, Dietrich A, et al. Insulin-sensitive obesity. Am J Physiol Endocrinol Metab. 2010;299:E506–E515. | ||
Oh IS, Shimizu H, Satoh T, et al. Identification of nesfatin-1 as a satiety molecule in the hypothalamus. Nature. 2006;443:709–712. | ||
Kloting N, Kovacs P, Kern M, et al. Central vaspin administration acutely reduces food intake and has sustained blood glucose-lowering effects. Diabetologia. 2011;54:1819–1823. | ||
Gaich G, Chien JY, Fu H, et al. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell Metab. 2013;18:333–340. | ||
Fisher FM, Kleiner S, Douris N, et al. FGF21 regulates PGC-1alpha and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012;26:271–281. | ||
Canto C, Auwerx J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr Opin Lipidol. 2009;20:98–105. | ||
Tseng YH, Kokkotou E, Schulz TJ, et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature. 2008;454:1000–1004. | ||
Townsend KL, Suzuki R, Huang TL, et al. Bone morphogenetic protein 7 (BMP7) reverses obesity and regulates appetite through a central mTOR pathway. FASEB J. 2012;26:2187–2196. | ||
Bostrom P, Wu J, Jedrychowski MP, et al. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. 2012;481:463–468. | ||
Roca-Rivada A, Castelao C, Senin LL, et al. FNDC5/irisin is not only a myokine but also an adipokine. PLoS One. 2013;8:e60563. | ||
Raschke S, Elsen M, Gassenhuber H, et al. Evidence against a beneficial effect of irisin in humans. PLoS One. 2013;8:e73680. | ||
Timmons JA, Baar K, Davidsen PK, Atherton PJ. Is irisin a human exercise gene? Nature. 2012;488:E9–E10; discussion E10–E11. | ||
Neary MT, Batterham RL. Gut hormones: implications for the treatment of obesity. Pharmacol Ther. 2009;124:44–56. | ||
Folgueira C, Seoane LM, Casanueva FF. The brain-stomach connection. Front Horm Res. 2014;42:83–92. | ||
Inui A, Asakawa A, Bowers CY, et al. Ghrelin, appetite, and gastric motility: the emerging role of the stomach as an endocrine organ. FASEB J. 2004;18:439–456. | ||
Cummings DE, Overduin J. Gastrointestinal regulation of food intake. J Clin Invest. 2007;117:13–23. | ||
Kojima M, Hosoda H, Date Y, et al. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature. 1999;402:656–660. | ||
Seoane LM, Al-Massadi O, Barreiro F, Dieguez C, Casanueva FF. Growth hormone and somatostatin directly inhibit gastric ghrelin secretion. An in vitro organ culture system. J Endocrinol Invest. 2007;30:RC22–RC25. | ||
Zhang X, Li W, Li P, et al. Intraportal infusion of ghrelin could inhibit glucose-stimulated GLP-1 secretion by enteric neural net in Wistar rat. Biomed Res Int. 2014;2014:923564. | ||
Garcia-Galiano D, Tena-Sempere M. Emerging roles of NUCB2/nesfatin-1 in the metabolic control of reproduction. Curr Pharm Des. 2013;19:6966–6972. | ||
Cota D, Marsicano G, Lutz B, et al. Endogenous cannabinoid system as a modulator of food intake. Int J Obes Relat Metab Disord. 2003;27:289–301. | ||
Pazos MR, Tolon RM, Benito C, et al. Cannabinoid CB1 receptors are expressed by parietal cells of the human gastric mucosa. J Histochem Cytochem. 2008;56:511–516. | ||
Khamzina L, Veilleux A, Bergeron S, Marette A. Increased activation of the mammalian target of rapamycin pathway in liver and skeletal muscle of obese rats: possible involvement in obesity-linked insulin resistance. Endocrinology. 2005;146:1473–1481. | ||
Senin LL, Al-Massadi O, Folgueira C, et al. The gastric CB1 receptor modulates ghrelin production through the mTOR pathway to regulate food intake. PLoS One. 2013;8:e80339. | ||
Li Z, Xu G, Li Y, et al. mTOR-dependent modulation of gastric nesfatin-1/NUCB2. Cell Physiol Biochem. 2012;29:493–500. | ||
United States Department of Health and Human Services. United States. Department of Agriculture. Dietary Guidelines Advisory Committee. Dietary Guidelines for Americans, 2010. Washington, DC: Government Printing Office, 2010, xi, 95. | ||
Jakicic JM. The effect of physical activity on body weight. Obesity (Silver Spring). 2009;17(Suppl 3):S34–S38. | ||
Kral JG, Naslund E. Surgical treatment of obesity. Nat Clin Pract Endocrinol Metab. 2007;3:574–583. | ||
Sjostrom L, Narbro K, Sjostrom CD, et al. Effects of bariatric surgery on mortality in Swedish obese subjects. N Engl J Med. 2007;357:741–752. | ||
Lopez M, Seoane LM, Tovar S, et al. Orexin-A regulates growth hormone-releasing hormone mRNA content in a nucleus-specific manner and somatostatin mRNA content in a growth hormone-dependent fashion in the rat hypothalamus. Eur J Neurosci. 2004;19:2080–2088. | ||
Ioannides-Demos LL, Proietto J, McNeil JJ. Pharmacotherapy for obesity. Drugs. 2005;65:1391–1418. | ||
Bai B, Wang Y. The use of lorcaserin in the management of obesity: a critical appraisal. Drug Des Devel Ther. 2010;5:1–7. | ||
Kirilly E, Gonda X, Bagdy G. CB1 receptor antagonists: new discoveries leading to new perspectives. Acta Physiol (Oxf). 2012;205:41–60. | ||
Yanovski SZ, Yanovski JA. Long-term drug treatment for obesity: a systematic and clinical review. JAMA. 2014;311:74–86. | ||
Bluher M. Adipokines–removing road blocks to obesity and diabetes therapy. Mol Metab. 2014;3:230–240. | ||
Bauche IB, El Mkadem SA, Pottier AM, et al. Overexpression of adiponectin targeted to adipose tissue in transgenic mice: impaired adipocyte differentiation. Endocrinology. 2007;148:1539–1549. | ||
Yamauchi T, Kamon J, Waki H, et al. Globular adiponectin protected ob/ob mice from diabetes and ApoE-deficient mice from atherosclerosis. J Biol Chem. 2003;278:2461–2468. | ||
Ahima RS, Flier JS. Leptin. Annu Rev Physiol. 2000;62:413–437. | ||
Savage DB, O’Rahilly S. Leptin: a novel therapeutic role in lipodystrophy. J Clin Invest. 2002;109:1285–1286. | ||
Foo JP, Mantzoros CS. Leptin in congenital or HIV-associated lipodystrophy and metabolic syndrome: a need for more mechanistic studies and large, randomized, placebo-controlled trials. Metabolism. 2012;61:1331–1336. | ||
Chou K, Perry CM. Metreleptin: first global approval. Drugs. 2013;73:989–997. | ||
Muller TD, Sullivan LM, Habegger K, et al. Restoration of leptin responsiveness in diet-induced obese mice using an optimized leptin analog in combination with exendin-4 or FGF21. J Pept Sci. 2012;18:383–393. | ||
Okamoto Y, Folco EJ, Minami M, et al. Adiponectin inhibits the production of CXC receptor 3 chemokine ligands in macrophages and reduces T-lymphocyte recruitment in atherogenesis. Circ Res. 2008;102:218–225. | ||
Kusminski CM, Scherer PE. The road from discovery to clinic: adiponectin as a biomarker of metabolic status. Clin Pharmacol Ther. 2009;86:592–595. | ||
Heiker JT, Kloting N, Kovacs P, et al. Vaspin inhibits kallikrein 7 by serpin mechanism. Cell Mol Life Sci. 2013;70:2569–2583. | ||
Lotfy M, Singh J, Kalasz H, Tekes K, Adeghate E. Medicinal Chemistry and Applications of Incretins and DPP-4 Inhibitors in the Treatment of Type 2 Diabetes Mellitus. Open Med Chem J. 2011;5:82–92. | ||
Saito M, Okamatsu-Ogura Y, Matsushita M, et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes. 2009;58:1526–1531. | ||
Al-Massadi O, Pardo M, Casanueva FF, Seoane LM. The stomach as an energy homeostasis regulating center. An approach for obesity. Recent Pat Endocr Metab Immune Drug Discovery. 2010;4:75–84. | ||
Kim GW, Lin JE, Blomain ES, Waldman SA. Antiobesity pharmacotherapy: new drugs and emerging targets. Clin Pharmacol Ther. 2014;95:53–66. | ||
Ravussin E, Gautier JF. Metabolic predictors of weight gain. Int J Obes Relat Metab Disord. 1999;23(Suppl 1):37–41. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.